
Genetic Evidence for the Presence of Wild-Caught Sturgeons in Commercial Markets in Georgia

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
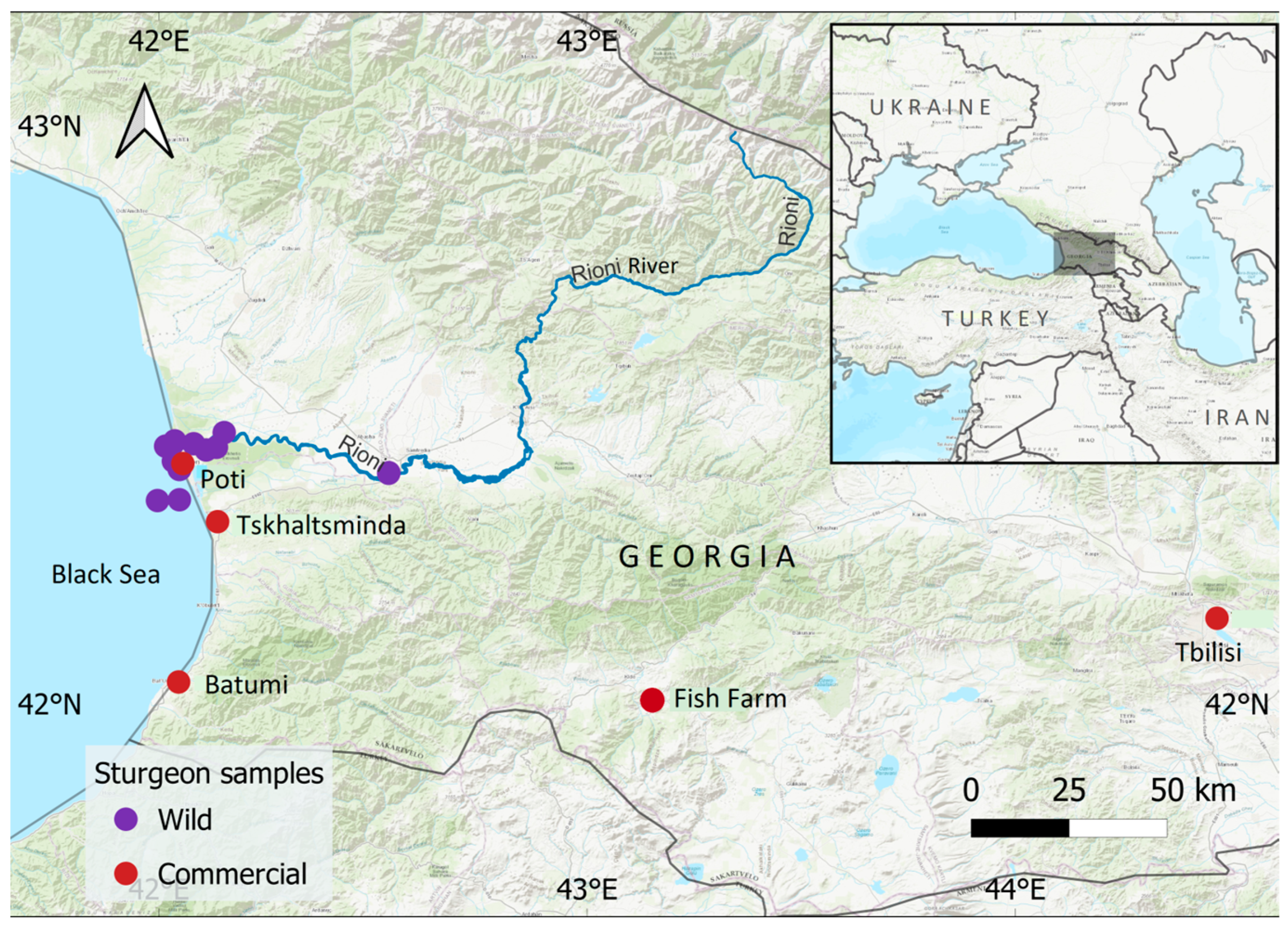
2.1. Sample Collection
2.2. Mitochondrial Analysis
2.3. Microsatellite Analysis
3. Results
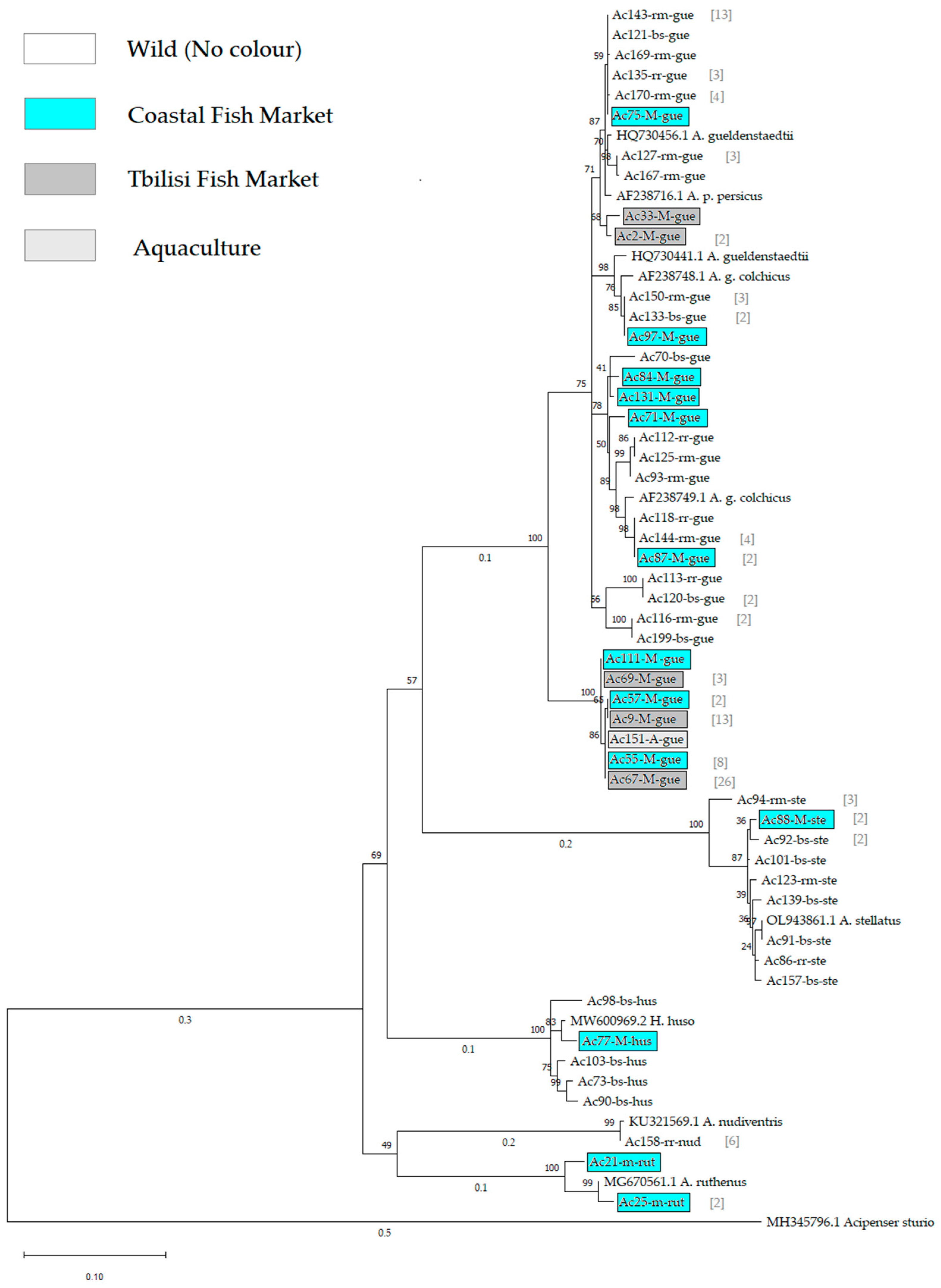
3.1. Mitochondrial DNA Analysis
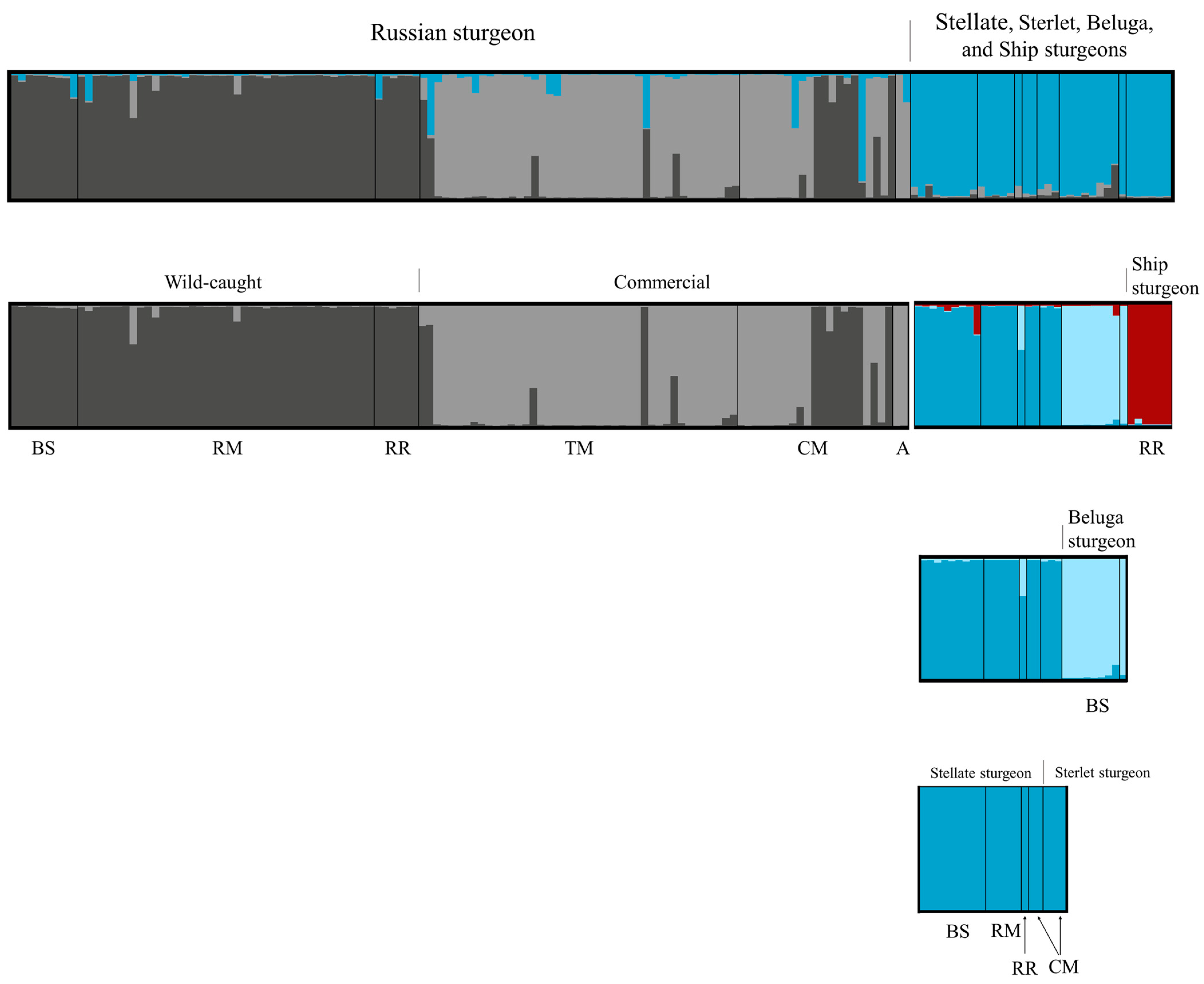
3.2. Microsatellite Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bemis, W.E.; Kynard, B. Sturgeon Rivers: An Introduction to Acipenseriform Biogeography and Life History. In Sturgeon Biodiversity and Conservation; Birstein, V.J., Waldman, J.R., Bemis, W.E., Eds.; Developments in Environmental Biology of Fishes; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; Volume 17, pp. 167–183. ISBN 978-0-7923-4517-6. [Google Scholar]
- Peng, Z.; Ludwig, A.; Wang, D.; Diogo, R.; Wei, Q.; He, S. Age and Biogeography of Major Clades in Sturgeons and Paddlefishes (Pisces: Acipenseriformes). Mol. Phylogenetics Evol. 2007, 42, 854–862. [Google Scholar] [CrossRef]
- Congiu, L.; Striebel-Greiter, B.; Gessner, J.; Boscari, E.; Boner, M.; Jahrl, J.; Dalle Palle, S.; Ludwig, A. Identification and Tracking of Sturgeons and Paddlefish Products in Trade: Implications for Trade Control and Biodiversity Management. Aquaculture 2023, 574, 739708. [Google Scholar] [CrossRef]
- Ludwig, A. Identification of Acipenseriformes Species in Trade. J. Appl. Ichthyol. 2008, 24, 2–19. [Google Scholar] [CrossRef]
- Bronzi, P.; Rosenthal, H.; Gessner, J. Global Sturgeon Aquaculture Production: An Overview. J. Appl. Ichthyol. 2011, 27, 169–175. [Google Scholar] [CrossRef]
- Banarescu, P.; Holcik, J. Acipenseriformes. In The Freshwater Fishes of Europe; Aula Verlag: Wiesbaden, Germany, 1989; Volume 1. [Google Scholar]
- Bronzi, P.; Chebanov, M.; Michaels, J.T.; Wei, Q.; Rosenthal, H.; Gessner, J. Sturgeon Meat and Caviar Production: Global Update 2017. J. Appl. Ichthyol. 2019, 35, 257–266. [Google Scholar] [CrossRef]
- Import of Live Sturgeon for Aquaculture; Department of Agriculture, Fisheries and Forestry DAFF: Canberra, Australia, 2023.
- White, S.L.; Fox, D.A.; Beridze, T.; Bolden, S.K.; Johnson, R.L.; Savoy, T.F.; Scheele, F.; Schreier, A.D.; Kazyak, D.C. Decades of Global Sturgeon Conservation Efforts Are Threatened by an Expanding Captive Culture Industry. Fisheries 2023, 48, 54–61. [Google Scholar] [CrossRef]
- Jahrl, J.; Boner, M.; Striebel, B.; Ludwig, A. Evidence for Trafficking of Critically Endangered Sturgeon in the Lower Danube Region; WWF Austria & WWF CEE: Vienna, Austria, 2021. [Google Scholar]
- Ninua, L. Baseline Study of Sturgeon Catch and Illegal Trade in Georgia; Fauna & Flora Caucasus Programme: Tbilisi, Georgia, 2018. [Google Scholar]
- Ludwig, A. A Sturgeon View on Conservation Genetics. Eur. J. Wildl. Res. 2006, 52, 3–8. [Google Scholar] [CrossRef]
- Boscari, E.; Barmintseva, A.; Pujolar, J.M.; Doukakis, P.; Mugue, N.; Congiu, L. Species and Hybrid Identification of Sturgeon Caviar: A New Molecular Approach to Detect Illegal Trade. Mol. Ecol. Resour. 2014, 14, 489–498. [Google Scholar] [CrossRef]
- Boscari, E.; Vitulo, N.; Ludwig, A.; Caruso, C.; Mugue, N.S.; Suciu, R.; Onara, D.F.; Papetti, C.; Marino, I.A.M.; Zane, L.; et al. Fast Genetic Identification of the Beluga Sturgeon and Its Sought-after Caviar to Stem Illegal Trade. Food Control 2017, 75, 145–152. [Google Scholar] [CrossRef]
- Havelka, M.; Bytyutskyy, D.; Symonová, R.; Ráb, P.; Flajšhans, M. The Second Highest Chromosome Count among Vertebrates Is Observed in Cultured Sturgeon and Is Associated with Genome Plasticity. Genet. Sel. Evol. 2016, 48, 12. [Google Scholar] [CrossRef]
- Ludwig, A.; Lippold, S.; Debus, L.; Reinartz, R. First Evidence of Hybridization between Endangered Sterlets (Acipenser ruthenus) and Exotic Siberian Sturgeons (Acipenser baerii) in the Danube River. Biol. Invasions 2009, 11, 753–760. [Google Scholar] [CrossRef]
- Ninua, N.; Guchmanidze, A. Sturgeons of Georgia; Georgian National Museum: Tbilisi, Georgia, 2013; ISBN 978-9941-9311-2-3. [Google Scholar]
- Zarkua, Z.; Bolkvadze, I.; Khintibidze, M.; Gogotishvili, M.; Variadilis, L. The Current State of the Georgian Black Sea Sturgeons; ASEN2; Springer: Dordrecht, The Netherlands, 1998; Volume 46. [Google Scholar]
- Beridze, T.; Boscari, E.; Scheele, F.; Edisherashvili, T.; Anderson, C.; Congiu, L. Interspecific Hybridization in Natural Sturgeon Populations of the Eastern Black Sea: The Consequence of Drastic Population Decline? Conserv. Genet. 2022, 23, 211–216. [Google Scholar] [CrossRef]
- Rogava, J.; Potskhishvili, M.; Seskuria, G. Report on the Sturgeon Farms in Georgia; Fauna & Flora Caucasus Programme: Tbilisi, Georgia, 2020. [Google Scholar]
- Friedrich, T.; Gessner, J.; Reinartz, R.; Greiter, S.B. Pan-European Action Plan for Sturgeons, Convention on the Conservation of European Wildlife and Natural Habitats; World Sturgeon Conservation Society and WWF: Strasbourg, France, 2018. [Google Scholar]
- Fox, D.A.; Hightower, J.E.; Parauka, F.M. Gulf Sturgeon Spawning Migration and Habitat in the Choctawhatchee River System, Alabama–Florida. Trans. Am. Fish. Soc. 2000, 129, 811–826. [Google Scholar] [CrossRef]
- Kazyak, D.C.; White, S.L.; Lubinski, B.A.; Johnson, R.; Eackles, M. Stock Composition of Atlantic Sturgeon (Acipenser oxyrinchus oxyrinchus) Encountered in Marine and Estuarine Environments on the U.S. Atlantic Coast. Conserv. Genet. 2021, 22, 767–781. [Google Scholar] [CrossRef]
- Laikre, L.; Palm, S.; Ryman, N. Genetic Population Structure of Fishes: Implications for Coastal Zone Management. AMBIO A J. Hum. Environ. 2005, 34, 111–119. [Google Scholar] [CrossRef]
- Schreier, A.D.; Mahardja, B.; May, B. Hierarchical Patterns of Population Structure in the Endangered Fraser River White Sturgeon (Acipenser transmontanus) and Implications for Conservation. Can. J. Fish. Aquat. Sci. 2012, 69, 1968–1980. [Google Scholar] [CrossRef]
- Congiu, L.; Pujolar, J.M.; Forlani, A.; Cenadelli, S.; Dupanloup, I.; Barbisan, F.; Galli, A.; Fontana, F. Managing Polyploidy in Ex Situ Conservation Genetics: The Case of the Critically Endangered Adriatic Sturgeon (Acipenser naccarii). PLoS ONE 2011, 6, e18249. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Henderson-Arzapalo, A.; King, T.L. Novel Microsatellite Markers for Atlantic Sturgeon (Acipenser oxyrinchus) Population Delineation and Broodstock Management. Mol. Ecol. Notes 2002, 2, 437–439. [Google Scholar] [CrossRef]
- King, T.L.; Lubinski, B.A.; Spidle, A.P. Microsatellite DNA Variation Atlantic Sturgeon (Acipenser oxyrinchus oxyrinchus) and Cross-Species Amplification the Acipenseridae. Conserv. Genet. 2001, 2, 103–119. [Google Scholar] [CrossRef]
- Welsh, A.B.; Blumberg, M.; May, B. Identification of Microsatellite Loci in Lake Sturgeon, Acipenser fulvescens, and Their Variability in Green Sturgeon, A. medirostris. Mol. Ecol. Notes 2002, 3, 47–55. [Google Scholar] [CrossRef]
- Zane, L.; Patarnello, T.; Ludwig, A.; Fontana, F.; Congiu, L. Isolation and Characterization of Microsatellites in the Adriatic Sturgeon (Acipenser naccarii). Mol. Ecol. Notes 2002, 2, 586–588. [Google Scholar] [CrossRef]
- Janes, J.K.; Miller, J.M.; Dupuis, J.R.; Malenfant, R.M.; Gorrell, J.C.; Cullingham, C.I.; Andrew, R.L. The K = 2 Conundrum. Mol. Ecol. 2017, 26, 3594–3602. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Stift, M.; Kolář, F.; Meirmans, P.G. Structure Is More Robust than Other Clustering Methods in Simulated Mixed-Ploidy Populations. Heredity 2019, 123, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-L.; Liu, J.-X. StructureSelector: A Web-Based Software to Select and Visualize the Optimal Number of Clusters Using Multiple Methods. Mol. Ecol. Resour. 2018, 18, 176–177. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software Structure: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Ludwig, A.; Congiu, L.; Gessner, J.; Boscari, E.; Striebel-Greiter, B.; Boner, M.; Jahrl, J. Identification of Species and Hybrids, Source and Geographical Origin of Sturgeon and Paddlefish (Acipenseriformes spp.) Specimens and Products in Trade. In Proceedings of the Addendum to Identification and Traceability of Sturgeons and Paddlefish (Acipenseriformes spp.), CITES—Thirty-First Meeting of the Animals Committee, Online, 31 May–22 June 2021. [Google Scholar]
- Ludwig, A.; Lieckfeldt, D.; Jahrl, J. Mislabeled and Counterfeit Sturgeon Caviar from Bulgaria and Romania. J. Appl. Ichthyol. 2015, 31, 587–591. [Google Scholar] [CrossRef]
- Barmintseva, A.E.; Mugue, N.S. The Use of Microsatellite Loci for Identification of Sturgeon Species (Acipenseridae) and Hybrid Forms. Russ. J. Genet. 2013, 49, 950–961. [Google Scholar] [CrossRef]
- Reinartz, R.; Lippold, S.; Lieckfeldt, D.; Ludwig, A. Population Genetic Analyses of Acipenser ruthenus as a Prerequisite for the Conservation of the Uppermost Danube Population: Genotype Structure of the Danube Sterlet. J. Appl. Ichthyol. 2011, 27, 477–483. [Google Scholar] [CrossRef]
- Havelka, M.; Boscari, E.; Sergeev, A.; Mugue, N.; Congiu, L.; Arai, K. A New Marker, Isolated by ddRAD Sequencing, Detects Siberian and Russian Sturgeon in Hybrids. Anim. Genet. 2019, 50, 115–116. [Google Scholar] [CrossRef] [PubMed]
- Káldy, J.; Mozsár, A.; Fazekas, G.; Farkas, M.; Fazekas, D.L.; Fazekas, G.L.; Goda, K.; Gyöngy, Z.; Kovács, B.; Semmens, K.; et al. Hybridization of Russian Sturgeon (Acipenser gueldenstaedtii, Brandt and Ratzeberg, 1833) and American Paddlefish (Polyodon spathula, Walbaum 1792) and Evaluation of Their Progeny. Genes 2020, 11, 753. [Google Scholar] [CrossRef] [PubMed]
- Linhartová, Z.; Havelka, M.; Pšenička, M.; Flajšhans, M. Interspecific Hybridization of Sturgeon Species Affects Differently Their Gonadal Development. Czech J. Anim. Sci. 2018, 63, 1–10. [Google Scholar] [CrossRef]
- Suciu, R.; Molnar, P.; Kynard, B. Rioni River, Georgia Current Status of Sturgeon, and Implications for Future Development in the Catchment; Fauna & Flora Caucasus Programme: Tbilisi, Georgia, 2021; pp. 1–132. [Google Scholar]
- Lanfredi, M.; Congiu, L.; Garrido-Ramos, M.A.; de la Herrán, R.; Leis, M.; Chicca, M.; Rossi, R.; Tagliavini, J.; Ruiz Rejón, C.; Ruiz Rejón, M.; et al. Chromosomal Location and Evolution of a Satellite DNA Family in Seven Sturgeon Species. Chromosome Res. 2001, 9, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.; Debus, L.; Jenneckens, I. A Molecular Approach to Control the International Trade in Black Caviar. Int. Rev. Hydrobiol. 2002, 87, 661–674. [Google Scholar] [CrossRef]
- Meirmans, P.G.; Liu, S.; van Tienderen, P.H. The Analysis of Polyploid Genetic Data. J. Hered. 2018, 109, 283–296. [Google Scholar] [CrossRef]
- Avigliano, E.; Leisen, M.; Duquenoy, C.; Liotta, J.; Volpedo, A. Siberian and Russian Sturgeon Natal Origin in South America: Fish Farm or Established Population? Austral Ecol. 2023, 48, 1121–1131. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Wild-Caught | Commercial | ||||
|---|---|---|---|---|---|---|
| Rioni River | Rioni River Mouth | Black Sea | Coastal Market | Tbilisi Market | Aquaculture | |
| Russian sturgeon | 6 | 40 (34) | 9 (6) | 21 (18) | 43 (45) | 2 (1) |
| Ship sturgeon | 4 | 2 | - | - | - | - |
| Stellate sturgeon | 1 | 5 (4) | 9 (6) | 2 | - | - |
| Sterlet sturgeon | - | - | - | 3 | - | - |
| Beluga sturgeon | - | - | 8 (4) | 1 | - | - |
| Total for each group | 11 | 47 (40) | 26 (16) | 27 (24) | 43 (45) | 2 (1) |
| Total | 84 (67) | 72 (70) | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beridze, T.; White, S.L.; Kazyak, D.C.; Ninua, L.; Fox, D.; Sethuraman, A.; Edisherashvili, T.; Roberts, B.; Potskhishvili, M.; Klailova, M.; et al. Genetic Evidence for the Presence of Wild-Caught Sturgeons in Commercial Markets in Georgia. Diversity 2024, 16, 274. https://doi.org/10.3390/d16050274
Beridze T, White SL, Kazyak DC, Ninua L, Fox D, Sethuraman A, Edisherashvili T, Roberts B, Potskhishvili M, Klailova M, et al. Genetic Evidence for the Presence of Wild-Caught Sturgeons in Commercial Markets in Georgia. Diversity. 2024; 16(5):274. https://doi.org/10.3390/d16050274
Chicago/Turabian StyleBeridze, Tamar, Shannon L. White, David C. Kazyak, Levan Ninua, Dewayne Fox, Arun Sethuraman, Tamari Edisherashvili, Bianca Roberts, Mikheil Potskhishvili, Michelle Klailova, and et al. 2024. "Genetic Evidence for the Presence of Wild-Caught Sturgeons in Commercial Markets in Georgia" Diversity 16, no. 5: 274. https://doi.org/10.3390/d16050274
APA StyleBeridze, T., White, S. L., Kazyak, D. C., Ninua, L., Fox, D., Sethuraman, A., Edisherashvili, T., Roberts, B., Potskhishvili, M., Klailova, M., & Anderson, C. (2024). Genetic Evidence for the Presence of Wild-Caught Sturgeons in Commercial Markets in Georgia. Diversity, 16(5), 274. https://doi.org/10.3390/d16050274