


Species Composition and Distribution of Terrestrial Herbs in a High Montane Forest in Ecuador

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Species Data
2.3. Species Endemism and Conservation Status
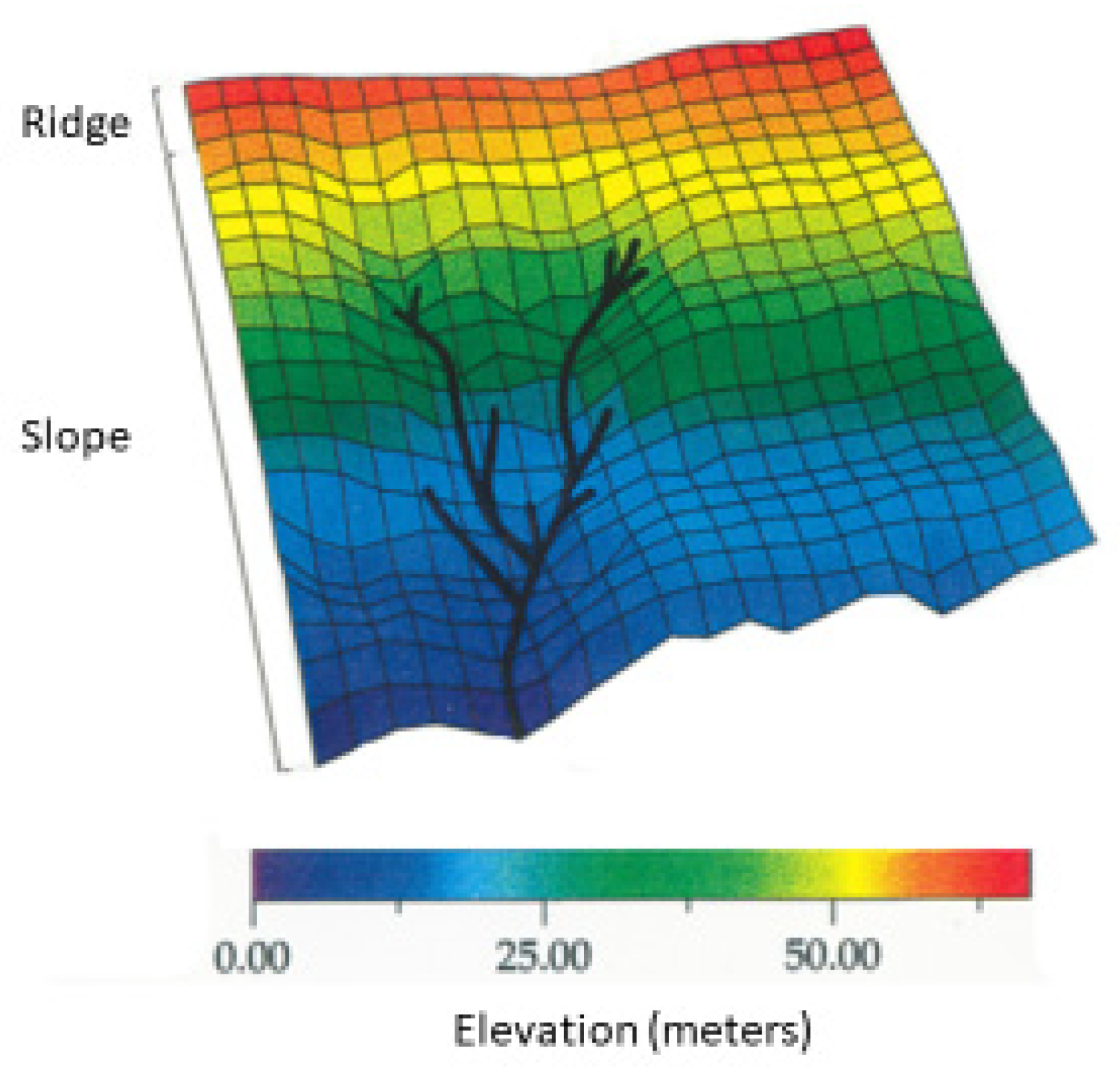
2.4. Topographic Microhabitats
2.5. Light Microhabitats
2.6. Statistical Analysis
3. Results
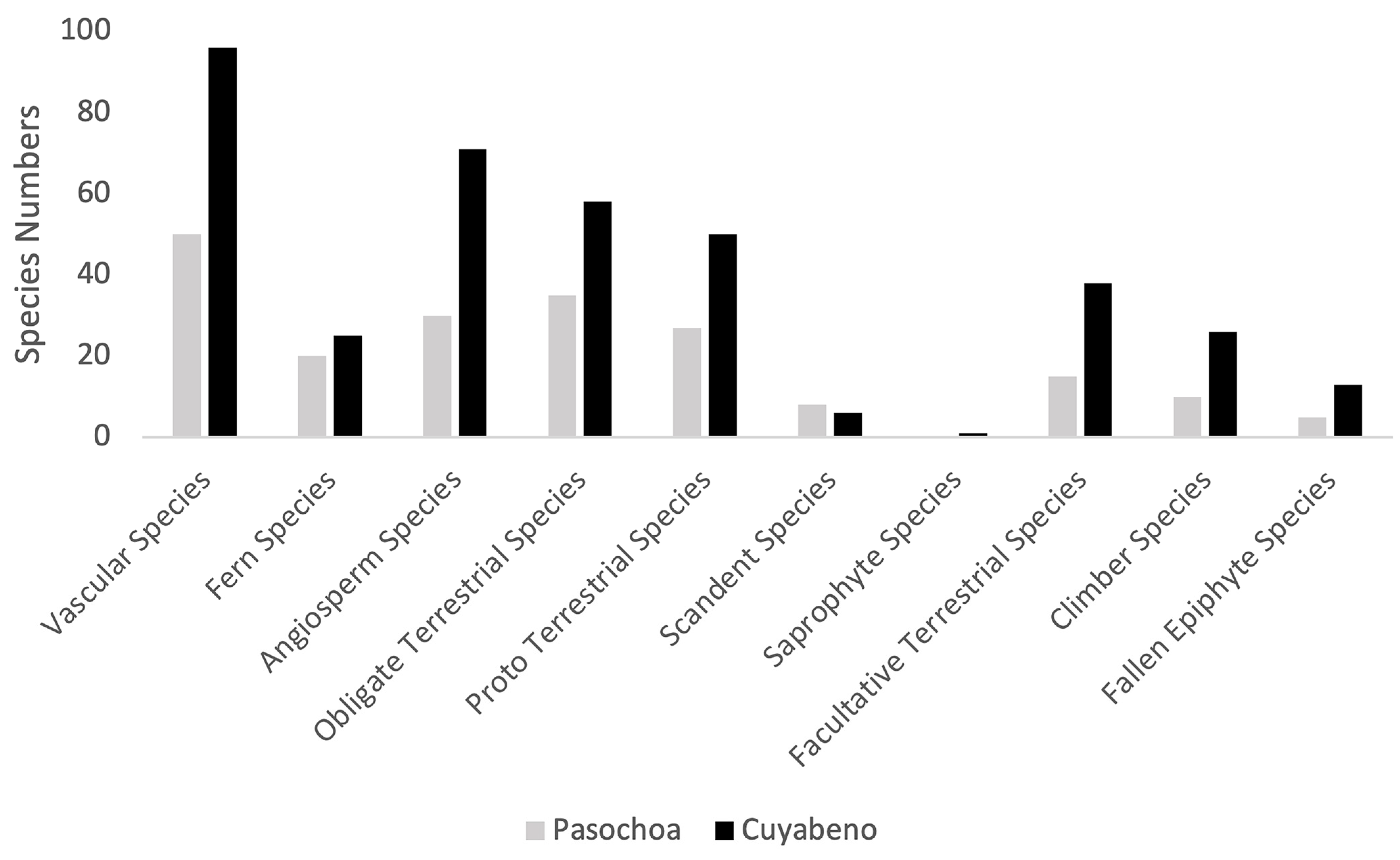
3.1. Species Composition and Richness
3.2. Life Forms
3.3. Species Endemism and Conservation Status
3.4. Density
3.5. Cover
3.6. Indices of Importance Value
3.7. Microhabitats and Species Preferences
3.8. Statistical Analysis
4. Discussion
4.1. Density
4.2. Cover
4.3. Importance Value of Species and Microhabitat Associations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gentry, A.H.; Dodson, C. Contribution of Nontrees to Species Richness of a Tropical Rain Forest. Biotropica 1987, 19, 149–156. [Google Scholar] [CrossRef]
- Galeano, G.; Suárez, S.; Balslev, H. Vascular Plant Species Count in a Wet Forest in the Choco Area on the Pacific Coast of Colombia. Biodivers. Conserv. 1998, 7, 1563–1575. [Google Scholar] [CrossRef]
- Wright, J.S. Plant Diversity in Tropical Forests: A Review of Mechanisms of Species Coexistence. Oecologia 2002, 130, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Valencia, R.; Condit, R.; Foster, R.B.; Romoleroux, K.; Villa Munoz, G.; Svenning, J.C.; Magard, E.; Bass, M.; Losos, E.C.; Balslev, H. Yasuni Forest Dynamics Plot, Ecuador. In Tropical Forest Diversity and Dynamism: Findings from a Large-Scale Plot Network; The University of Chicago Press: Chicago, IL, USA, 2004. [Google Scholar]
- Esquivel-Muelbert, A.; Pugh, T.; Baker, T.; Dexter, K.; Lewis, S.; Galbraith, D.; Phillips, O. Vulnerability of Amazonian Tree Communities to Global Change. In Proceedings of the EGU General Assembly Conference, Online, 4–8 May 2020. [Google Scholar]
- Banda, K.R.; Delgado-Salinas, A.; Dexter, K.G.; Linares-Palomino, R.; Oliveira-Filho, A.; Prado, D.; Pullan, M.; Quintana, C.; Riina, R.; Rodríguez, G.M.; et al. Plant Diversity Patterns in Neotropical Dry Forests and Their Conservation Implications. Science 2016, 353, 1383–1387. [Google Scholar] [CrossRef]
- Guevara Andino, J.E.; Hernández, C.; Valencia, R.; Forrister, D.; Endara, M.J. Accelerating the Discovery of Rare Tree Species in Amazonian Forests: Integrating Long Monitoring Tree Plot Data with Metabolomics and Phylogenetics for the Description of a New Species in the Hyperdiverse Genus Inga Mill. PeerJ 2022, 10, e13767. [Google Scholar] [CrossRef] [PubMed]
- Spicer, M.E.; Mellor, H.; Carson, W.P. Seeing beyond the Trees: A Comparison of Tropical and Temperate Plant Growth Forms and Their Vertical Distribution. Ecology 2020, 101, e02974. [Google Scholar] [CrossRef] [PubMed]
- Cicuzza, D.; Kessler, M.; Pitopang, R.; Tjitrosoedirdjo, S.; Gradstein, S.R. Terrestrial Herb Communities of Tropical Submontane and Tropical Montane Forests in Central Sulawesi, Indonesia. In Environmental Science and Engineering; Springer Science and Business Media Deutschland GmbH: Berlin/Heidelberg, Germany, 2010; pp. 377–390. [Google Scholar]
- Willinghöfer, S.; Cicuzza, D.; Kessler, M. Elevational Diversity of Terrestrial Rainforest Herbs: When the Whole Is Less than the Sum of Its Parts. Plant Ecol. 2012, 213, 407–418. [Google Scholar] [CrossRef]
- Vieira, L.T.A.; Polisel, R.T.; Ivanauskas, N.M.; Shepherd, G.J.; Waechter, J.L.; Yamamoto, K.; Martins, F.R. Geographical Patterns of Terrestrial Herbs: A New Component in Planning the Conservation of the Brazilian Atlantic Forest. Biodivers. Conserv. 2015, 24, 2181–2198. [Google Scholar] [CrossRef]
- Whitmore, T.C.; Peralta, R.; Brown, K. Total Species Count in a Costa Rican Tropical Rain Forest. J. Trop. Ecol. 1985, 1, 375–378. [Google Scholar] [CrossRef]
- Poulsen, A.D.; Balslev, H. Abundance and Cover of Ground Herbs in an Amazonian Rain Forest. J. Veg. Sci. 1991, 2, 315–322. [Google Scholar] [CrossRef]
- Poulsen, A.D.; Pendry, C.A. Inventories of Ground Herbs at Three Altitudes on Bukit Belalong, Brunei, Borneo. Biodivers. Conserv. 1995, 4, 745–757. [Google Scholar] [CrossRef]
- Poulsen, A.D. Species Richness and Density of Ground Herbs within a Plot of Lowland Rainforest in North-West Borneo. J. Trop. Ecol. 1996, 12, 177–190. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K. Distribution of Pteridophyta and Melastomataceae along an Edaphic Gradient in an Amazonian Rain Forest. J. Veg. Sci. 1994, 5, 25–34. [Google Scholar] [CrossRef]
- Grubb, P.J.; Lloyd, J.R.; Pennington, T.D.; Whitmore, T.C. A Comparison of Montane and Lowland Rain Forest in Ecuador I. The Forest Structure, Physiognomy, and Floristics. J. Ecol. 1963, 51, 567–601. [Google Scholar] [CrossRef]
- Young, K.R.; León, B. Pteridophyte Species Diversity in the Central Peruvian Amazon: Importance of Edaphic Specialization. Brittonia 1989, 41, 388–395. [Google Scholar] [CrossRef]
- Linares-Palomino, R.; Cardona, V.; Hennig, E.I.; Hensen, I.; Hoffmann, D.; Lendzion, J.; Soto, D.; Herzog, S.K.; Kessler, M. Non-Woody Life-Form Contribution to Vascular Plant Species Richness in a Tropical American Forest. Plant Ecol. 2009, 201, 87–99. [Google Scholar] [CrossRef][Green Version]
- Tuomisto, H.; Poulsen, A.D.; Ruokolainen, K.; Moran, R.C.; Quintana, C.; Celi, J.; Cañas, G. Linking Floristic Patterns with Soil Heterogeneity and Satellite Imagery in Ecuadorian Amazonia. Ecol. Appl. 2003, 13, 352–371. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K.; Poulsen, A.D.; Moran, R.C.; Quintana, C.; Cañas, G.; Celi, J. Distribution and Diversity of Pteridophytes and Melastomataceae along Edaphic Gradients in Yasuní National Park, Ecuadorian Amazonia. Biotropica 2002, 34, 516–533. [Google Scholar] [CrossRef]
- Quintana, C.; Girardello, M.; Barfod, A.S.; Balslev, H. Diversity Patterns, Environmental Drivers and Changes in Vegetation Composition in Dry Inter-Andean Valleys. J. Plant Ecol. 2017, 10, rtw036–475. [Google Scholar] [CrossRef]
- Llambí, L.D.; Rada, F. Ecological Research in the Tropical Alpine Ecosystems of the Venezuelan Páramo: Past, Present and Future. Plant Ecol. Divers. 2019, 12, 519–538. [Google Scholar] [CrossRef]
- Esteban Suárez, R.; Medina, G. Vegetation Structure and Soil Properties in Ecuadorian Páramo Grasslands with Different Histories of Burning and Grazing. Arctic, Antarct. Alp. Res. 2001, 33, 158–164. [Google Scholar] [CrossRef]
- Grubb, P.J.; Lloyd, J.R.; Pennington, T.D.; Páez-Bimos, S. A Historical Baseline Study of the Páramo of Antisana in the Ecuadorian Andes Including the Impacts of Burning, Grazing and Trampling. Plant Ecol. Divers. 2020, 13, 225–256. [Google Scholar] [CrossRef]
- Cruz, M.; Lasso, E. Insights into the Functional Ecology of Páramo Plants in Colombia. Biotropica 2021, 53, 1415–1431. [Google Scholar] [CrossRef]
- Keating, P.L.; Young, K.R.; León, B. Variation in High Andean Vegetation at a Site in Southwestern Ecuador. Pa. Geogr. 2002, 40, 15–35. [Google Scholar]
- Ramsay, P.M.; Oxley, E.R.B. The Growth Form Composition of Plant Communities in the Ecuadorian Paramos. Plant Ecol. 1997, 131, 173–192. [Google Scholar] [CrossRef]
- Leon-Garcia, I.V.; Lasso, E. High Heat Tolerance in Plants from the Andean Highlands: Implications for Paramos in a Warmer World. PLoS ONE 2019, 14, e0224218. [Google Scholar] [CrossRef] [PubMed]
- Campos, P.V.; Villa, P.M.; Nunes, J.A.; Schaefer, C.E.G.R.; Porembski, S.; Neri, A.V. Plant Diversity and Community Structure of Brazilian Páramos. J. Mt. Sci. 2018, 15, 1186–1198. [Google Scholar] [CrossRef]
- Lozano, P.; Cabrera, O.; Peyre, G.; Cleef, A.; Toulkeridis, T. Plant Diversity and Composition Changes along an Altitudinal Gradient in the Isolated Volcano Sumaco in the Ecuadorian Amazon. Diversity 2020, 12, 229. [Google Scholar] [CrossRef]
- Sklenár, P.; Ramsay, P.M. Diversity of Zonal Páramo Plant Communities in Ecuador. Divers. Distrib. 2001, 7, 113–124. [Google Scholar] [CrossRef]
- Ramsay, P.M. Gradients in a Tropical Mountain Ecosystem of Ecuador. Mt. Res. Dev. 2008, 28, 340–341. [Google Scholar] [CrossRef]
- Tokarz, E.L.; Álvia, P.; Valencia, R.; Queenborough, S.A. Habitat Filtering of Six Coexisting Heliconia Species in a Lowland Tropical Rain Forest in Amazonian Ecuador. J. Trop. Ecol. 2019, 35, 91–94. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, J.; Xiao, L.; Zhong, Z.; Wang, Q.; Wang, W. Dominant Species Abundance, Vertical Structure and Plant Diversity Response to Nature Forest Protection in Northeastern China: Conservation Effects and Implications. Forests 2020, 11, 295. [Google Scholar] [CrossRef]
- Dar, J.A.; Sundarapandian, S. Patterns of Plant Diversity in Seven Temperate Forest Types of Western Himalaya, India. J. Asia-Pacific Biodivers. 2016, 9, 280–292. [Google Scholar] [CrossRef]
- Royo, A.A.; Carson, W.P. The Herb Community of a Tropical Forest in Central Panamá: Dynamics and Impact of Mammalian Herbivores. Oecologia 2005, 145, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.Y.; Yang, K.C.; Hsieh, C.F.; Hsieh, C.-H.; Su, M.H. Local Neighborhood Communities in the Understory Play a Critical Role by Affecting Regeneration Niches and Subsequent Community Assembly in a Montane Cloud Forest. Ecol. Res. 2017, 32, 821–833. [Google Scholar] [CrossRef]
- Radhamoni, H.V.N.; Queenborough, S.A.; Arietta, A.Z.A.; Suresh, H.S.; Dattaraja, H.S.; Kumar, S.S.; Sukumar, R.; Comita, L.S. Local- and Landscape-Scale Drivers of Terrestrial Herbaceous Plant Diversity along a Tropical Rainfall Gradient in Western Ghats, India. J. Ecol. 2023, 111, 1021–1036. [Google Scholar] [CrossRef]
- Sierra, R. Patrones Y Factores De Deforestación En El Ecuador Continental. Conserv. Int. Ecuador For. Trends 2013, 1, 22. [Google Scholar]
- Valencia, R.; Jorgensen, P.M. Composition and Structure of a Humid Montane Forest on the Pasochoa Volcano, Ecuador. Nord. J. Bot. 1992, 12, 239–247. [Google Scholar] [CrossRef]
- Hall, M.L.; Samaniego, P.; Le Pennec, J.L.; Johnson, J.B. Ecuadorian Andes Volcanism: A Review of Late Pliocene to Present Activity. J. Volcanol. Geotherm. Res. 2008, 176, 1–6. [Google Scholar] [CrossRef]
- Ministerio del Ambiente Areas Protegidas Del Ecuador. Available online: http://areasprotegidas.ambiente.gob.ec/ (accessed on 13 April 2024).
- Stern, M.J. An Inter-Andean Forest Relict: Vegetation Change on Pasochoa Volcano, Ecuador. Mt. Res. Dev. 1995, 15, 339–348. [Google Scholar] [CrossRef]
- Jorgensen, P.M.; Ulloa Ulloa, C.; Madsen, J.E.; Valencia, R., R. A Floristic Analysis of the High Andes of Ecuador. In Biodiversity and Conservation of Neotropical Montane Forests; New York Botanical Garden: Bronx, NY, USA, 1995. [Google Scholar]
- Curtis, J.T.; McIntosh, R.P. An Upland Forest Continuum in the Prairie-Forest Border Region of Wisconsin. Ecology 1951, 32, 476–496. [Google Scholar] [CrossRef]
- IUCN Standards and Petitions Committee. Guidelines for Using the IUCN Red List Categories and Criteria. Eur. J. Taxon. 2021, 2021, 1–122. [Google Scholar] [CrossRef]
- León-yánez, S.; Valencia, R.; Pitman, N.; Endara, L.; Ulloa, C.U.; Navarrete, H. Libro Rojo de Las Plantas Endémicas del Ecuador; Ediciones del Herbario QCA, Pontificia Universidad Católica del Ecuador, Quito 2011; Pages 1-440.
- Legendre, P. Comparison of Permutation Methods for the Partial Correlation and Partial Mantel Tests. J. Stat. Comput. Simul. 2000, 67, 37–73. [Google Scholar] [CrossRef]
- IUCN IUCN Red List Categories and Criteria: Version 3.1.; Gland. 2001.
- UNEP World Conservation Monitoring Centre Checklist of CITES Species; UNEP World Conservation Monitoring Centre: Cambidge, UK, 2003.
- Jørgensen, P.M.; Ulloa Ulloa, C. Seed Plants of the High Andes of Ecuador—A Checklist. AAU reports 1994, 34, 1–443. [Google Scholar]
- Renner, S.S. Reproduction and Evolution in Some Genera of Neotropical Melastomataceae. Mem. N. Y. Bot. Gard. 1990, 55, 143–152. [Google Scholar]
- Zizka, A.; ter Steege, H.; Pessoa, M.d.C.R.; Antonelli, A. Finding Needles in the Haystack: Where to Look for Rare Species in the American Tropics. Ecography 2018, 41, 321–330. [Google Scholar] [CrossRef]
- Prather, L.A.; Alvarez-Fuentes, O.; Mayfield, M.H.; Ferguson, C.J. Implications of the Decline in Plant Collecting for Systematic and Floristic Research. Syst. Bot. 2004, 29, 216–220. [Google Scholar] [CrossRef]
- Lendemer, J.; Thiers, B.; Monfils, A.K.; Zaspel, J.; Ellwood, E.R.; Bentley, A.; Levan, K.; Bates, J.; Jennings, D.; Contreras, D.; et al. The Extended Specimen Network: A Strategy to Enhance US Biodiversity Collections, Promote Research and Education. Bioscience 2020, 70, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Monfils, A.K.; Powers, K.E.; Marshall, C.J.; Martine, C.T.; Smith, J.F.; Prather, L.A. Natural History Collections: Teaching about Biodiversity Across Time, Space, and Digital Platforms. Southeast. Nat. 2017, 16, 47–57. [Google Scholar] [CrossRef]
- Moran, R.C. The Importance of Mountains to Pteridophytes, with Emphasis on Neotropical Montane Forests. In Biodiversity and Conservation of Neotropical Montane Forests; New York Botanical Garden: Bronx, NY, USA, 1995; pp. 359–363. [Google Scholar]
- Ollgaard, B. Diversity of Huperzia (Lycopodiaceae) in Neotropical Montane Forests. In Biodiversity and Conservation of Neotropical Montane Forests; New York Botanical Garden: Bronx, NY, USA, 1995; pp. 349–358. [Google Scholar]
- Mandl, N.; Lehnert, M.; Kessler, M.; Gradstein, S.R. A Comparison of Alpha and Beta Diversity Patterns of Ferns, Bryophytes and Macrolichens in Tropical Montane Forests of Southern Ecuador. Biodivers. Conserv. 2010, 19, 2359–2369. [Google Scholar] [CrossRef]
- Hubbell, S.P. Tropical Rain Forest Conservation and the Twin Challenges of Diversity and Rarity. Ecol. Evol. 2013, 3, 3263–3274. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.B.; Swaine, M.D. Distribution and Ecology of Vascular Plants in a Tropical Rain Forest; Springer Science and Business Media LLC: Dordrecht, Netherlands, 1981. [Google Scholar]
- YOUNG, K. Floristic Diversity on the Eastern Slopes of the Peruvian Andes. Candollea 1991, 46, 125–143. [Google Scholar]
- Pérez-Escobar, O.A.; Zizka, A.; Bermúdez, M.A.; Meseguer, A.S.; Condamine, F.L.; Hoorn, C.; Hooghiemstra, H.; Pu, Y.; Bogarín, D.; Boschman, L.M.; et al. The Andes through Time: Evolution and Distribution of Andean Floras. Trends Plant Sci. 2022, 27, 364–378. [Google Scholar] [CrossRef] [PubMed]
- G.S.P.H. The Tropical Rain Forest. By, P.W. Richards. Cambridge University Press, 1952. Pp. Xviii, 450; 43 Figs., 15 Plates, 39 Tables. 63s. Q. J. R. Meteorol. Soc. 1953, 79, 312. [Google Scholar] [CrossRef]
- Kabakoff, R.P.; Chazdon, R.L. Effects of Canopy Species Dominance on Understorey Light Availability in Low-Elevation Secondary Forest Stands in Costa Rica. J. Trop. Ecol. 1996, 12, 779–788. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Life Form | N | C | Topographic Micro Habitat | Light Micro Habitat | Iucn Conservation Status | Proposed Conservation Status | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | ARALIACEAE | Hydrocotyle alchemilloides A. Rich. (CQ 60) | O-P | 260 | 1.3 | s,r | h | - | SC |
| 2 | ARACEAE | Anthurium pulchrum Engl. (CQ 217) | F-C | 276 | 18.9 | s,r | h | - | WC |
| 3 | ASTERACEAE | Asteraceae sp. (CQ 126) | O-S | 41 | 1.5 | s | m | - | - |
| 4 | Ageratum sp. (CQ 233) | F-C | 16 | 0.7 | s,r,g | m,l | - | - | |
| 5 | BASELLACEAE | Tournonia hookeriana Moq. (CQ 211) | F-C | 189 | 7.8 | s | m,l | - | SC |
| 6 | BEGONIACEAE | Begonia sodiroi C. DC. (CQ 206) | F-C | 428 | 39.2 | s,r,g | h,m | NT | SC |
| 7 | CARYOPHYLLACEAE | Arenaria lanuginosa (Michx.) Rohrb. (CQ 110) | O-S | 35 | 1.5 | s,r | h,m,l | - | MC |
| 8 | Stellaria recurvata Willd. ex D.F.K. Schltdl. (CQ 143) | O-S | 59 | 3.1 | s,r,g | h,m,l | LC | WC | |
| 9 | CYPERACEAE | Carex hamata Sw. (CQ 213) | O-P | 7116 | 175 | s,r,g | h,m,l | - | WC |
| 10 | Carex muricata L. (CQ 191) | O-P | 2099 | 28.9 | s,r,g | h,m,l | - | MC | |
| 11 | Rhynchospora macrochaeta Steudel ex Boeckeler (CQ 209) | O-P | 1702 | 90 | s,r,g | h,m | - | WC | |
| 12 | FERNS | Adiantum concinnum Humb. and Bonpl. ex Willd. (CQ 245) | O-P | 6 | 0.7 | r,g | m | - | MC |
| 13 | Asplenium pteropus Kaulf (CQ 150) | O-P | 347 | 19.8 | s,r,g | h,m | - | WC | |
| 14 | Asplenium sessilifolium Desv. (CQ 190) | O-P | 1466 | 46.7 | s,r,g | h,m,l | - | MC | |
| 15 | Asplenium monanthes L. (CQ 153) | O-P | 187 | 7.5 | s,r,g | h,m | - | WC | |
| 16 | Asplenium castaneum Schltdl. and Cham. (CQ 132) | O-P | 475 | 8.1 | s,r,g | h,m | - | SC | |
| 17 | Asplenium peruvianum Desv. (CQ 124) | O-P | 110 | 1.9 | s,r,g | h,m | - | SC | |
| 18 | Asplenium squamosum L. (CQ 247) | O-P | 10 | 0.8 | g | h,m | - | SC | |
| 19 | Athyrium dombeyi Desv. (CQ 116) | O-P | 86 | 9.3 | s,r,g | h,m | - | SC | |
| 20 | Blechnum fragile (Liebm.) C.V. Morton and Lellinger (CQ 255) | O-S | 23 | 1.5 | g | m | - | MC | |
| 21 | Botrychium virginianum (L.) Sw. (CQ 144) | O-P | 41 | 1.4 | s | h,m | - | SC | |
| 22 | Campyloneurum amphostenon (Klotzch) Fée (CQ 155) | F-E | 31 | 0.9 | s,r,g | m | - | MC | |
| 23 | Dryopteris paleacea (Sw.) Hand.-Mazz. (CQ 112) | O-P | 72 | 40.5 | s,r,g | h,m | - | SC | |
| 24 | Elaphoglossum papillosum (Baker) Christ (CQ 100) | F-E | 2 | 0.1 | s | h,m | - | WC | |
| 25 | Equisetum bogotense Kunth (CQ 231) | O-P | 48 | 0.7 | s | h,m | - | WC | |
| 26 | Hypolepis bogotensis H. Karst. (CQ 258) | O-P | 260 | 0.9 | s | h,m | - | MC | |
| 27 | Pecluma divaricata (E. Fourn.) Mickel and Beitel (CQ 229) | O-P | 8 | 1 | s,g | h,m | - | MC | |
| 28 | Polypodium subandinum Sodiro (CQ 193) | F-C | 4620 | 43.7 | s,r,g | h,m,l | - | WC | |
| 29 | Polystichum muricatum (L.) Fée (CQ 67) | O-P | 737 | 50.6 | s,r,g | h,m | - | SC | |
| 30 | Pteris muricata Hook. (CQ 196) | O-P | 35 | 4.5 | g | h,m | - | MC | |
| 31 | Thelypteris rigescens (Sodiro) A.R. Sm. (CQ 23) | O-P | 102 | 68 | s,r,g | h,m,l | - | SC | |
| 32 | GERANIACEAE | Geranium laxicaule R. Knuth (CQ21) | O-S | 27 | 0.5 | s,r | h,m | - | SC |
| 33 | GESNERIACEAE | Alloplectus dielsii (Mansf.) Wiehler (CQ 197) | O-S | 451 | 19.3 | s,r,g | h,m | - | WC |
| 34 | ORCHIDACEAE | Baskervilla auriculata Garay (CQ 113) | O-P | 30 | 0.9 | s,r | h | NT | SC |
| 35 | Cranichis ciliata Kunth (CQ 135) | O-P | 208 | 6.2 | s,r | h | CA II | MC | |
| 36 | Malaxis hoppii (Schltr.)Løjtnant (CQ 119) | O-P | 406 | 6.8 | s,r,g | h | CA II | SC | |
| 37 | Stelis pusilla Kunth (CQ 111) | F-E | 59 | 3.1 | s,g | h,m | CA II | WC | |
| 38 | Odontoglossum pardinum (Lindl.)Lindl.(CQ 223) | F-E | 10 | 0.1 | s,r | h,m | CA II | MC | |
| 39 | Pleurothallis sp. (CQ 240) | F-E | 2 | 0.01 | s | h,m | - | - | |
| 40 | OXALIDACEAE | Oxalis lotoides Kunth (CQ 75) | O-S | 115 | 7.5 | s,r,g | h,m,l | - | WC |
| 41 | PHYTOLACCACEAE | Phytolacca bogotensis Kunth (CQ 270) | O-P | 14 | 0.6 | s,r | h,m | - | MC |
| 42 | PIPERACEAE | Peperomia rotundata Kunth (CQ 33) | F-C | 4860 | 58.1 | s,r,g | h,m,l | - | WC |
| 43 | Peperomia sp. 1 (CQ 224) | F-C | 102 | 1.5 | s,g | h,m | - | - | |
| 44 | Peperomia sp. 2 (CQ 243) | F-C | 24 | 0.6 | g | m | - | - | |
| 45 | POACEAE | Poa aequatoriensis Hack. (CQ 114) | O-P | 130 | 3.7 | s,r | h,m | - | SC |
| 46 | RUBIACEAE | Galium obovatum Kunth (CQ 133) | O-S | 39 | 0.5 | s,r | h,m | - | SC |
| 47 | Leptostigma pilosum (Benth.) Fosberg (CQ 106) | O-P | 1159 | 19.9 | s,r | h | - | SC | |
| 48 | SOLANACEAE | Solanum dalibardiforme Bitter (CQ 138) | F-C | 97 | 3.6 | s,r,g | m,l | - | SC |
| 49 | URTICACEAE | Pilea pubescens Liebm. (CQ 117) | F-C | 1883 | 7 | s,r | h,m | - | MC |
| 50 | Urtica leptophylla Kunth (CQ 249) | O-P | 8 | 0.9 | s | m | - | MC |
| Herbs Species/Individuals | Obligate Terrestrial Herbs Species/Individuals | Density (Number Individuals/m2) | Plot Area m2 | Altitude m | Study Area | Reference |
|---|---|---|---|---|---|---|
| 0.046 | * | 1.54 | 464 | 1710 | Ecuador (Borja) | [17] |
| 0.087 | * | 1.53 | 464 | 520 | Ecuador (Shinguipino) | [17] |
| - | 0.134 | 1.19 | 100 | 100 | Costa Rica | [12] |
| 0.04 | 0.03 | 0.03 | 100 | 3300 | Ecuador (Pasochoa) | This study |
| - | 0.018 | 2.85 | 1000 | 220 | Ecuador (Capeira) | [1] |
| - | 0.019 | 0.94 | 1000 | 220 | Ecuador (Jauneche) | [1] |
| - | 0.041 | 1.22 | 1000 | 220 | Ecuador (Palenque) | [1] |
| 0.009 | 0.005 | 1.1 | 10,000 | 250 | Ecuador (Cuyabeno) | [13] |
| 0.015 | * | 0.63 | 10,000 | 250 | Borneo (Brunei) | [15] |
| 0.027 | * | 0.24 | 10,000 | 250 | Borneo (Brunei) | [15] |
| 0.014 | 0.008 | 0.31 | 1000 | 3300 | Ecuador (Pasochoa) | This study |
| 0.008 | * | 2.67 | 30,000 | 900–1100 | Bolivia (Los Volcanes Reserve) | [19] |
| 0.002 | 0.011 | 3.11 | 10,000 | 3300 | Ecuador (Pasochoa) | This study |
| Matrix B | r | P | |
|---|---|---|---|
| Topography | 0.2 | 0.01 | |
| Space | 0.09 | 0.04 | |
| Light (presence/absence thicket) | 0.21 | 0.001 | |
| Matrix B | Matrix C | Partial r | P |
| Topography | Space | 0.187 | 0.0001 |
| Space | Topography | 0.02 | 0.33 |
| Topography | Light (presence/absence thicket) | 0.183 | 0.001 |
| Light (presence/absence thicket) | Topography | 0.193 | 0.007 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quintana, C.; Balslev, H.; Valencia, R. Species Composition and Distribution of Terrestrial Herbs in a High Montane Forest in Ecuador. Diversity 2024, 16, 262. https://doi.org/10.3390/d16050262
Quintana C, Balslev H, Valencia R. Species Composition and Distribution of Terrestrial Herbs in a High Montane Forest in Ecuador. Diversity. 2024; 16(5):262. https://doi.org/10.3390/d16050262
Chicago/Turabian StyleQuintana, Catalina, Henrik Balslev, and Renato Valencia. 2024. "Species Composition and Distribution of Terrestrial Herbs in a High Montane Forest in Ecuador" Diversity 16, no. 5: 262. https://doi.org/10.3390/d16050262
APA StyleQuintana, C., Balslev, H., & Valencia, R. (2024). Species Composition and Distribution of Terrestrial Herbs in a High Montane Forest in Ecuador. Diversity, 16(5), 262. https://doi.org/10.3390/d16050262