
Reconstructing the Paleoenvironmental Evolution of Lake Kolon (Hungary) through Palaeoecological, Statistical and Historical Analyses




Abstract
:1. Introduction
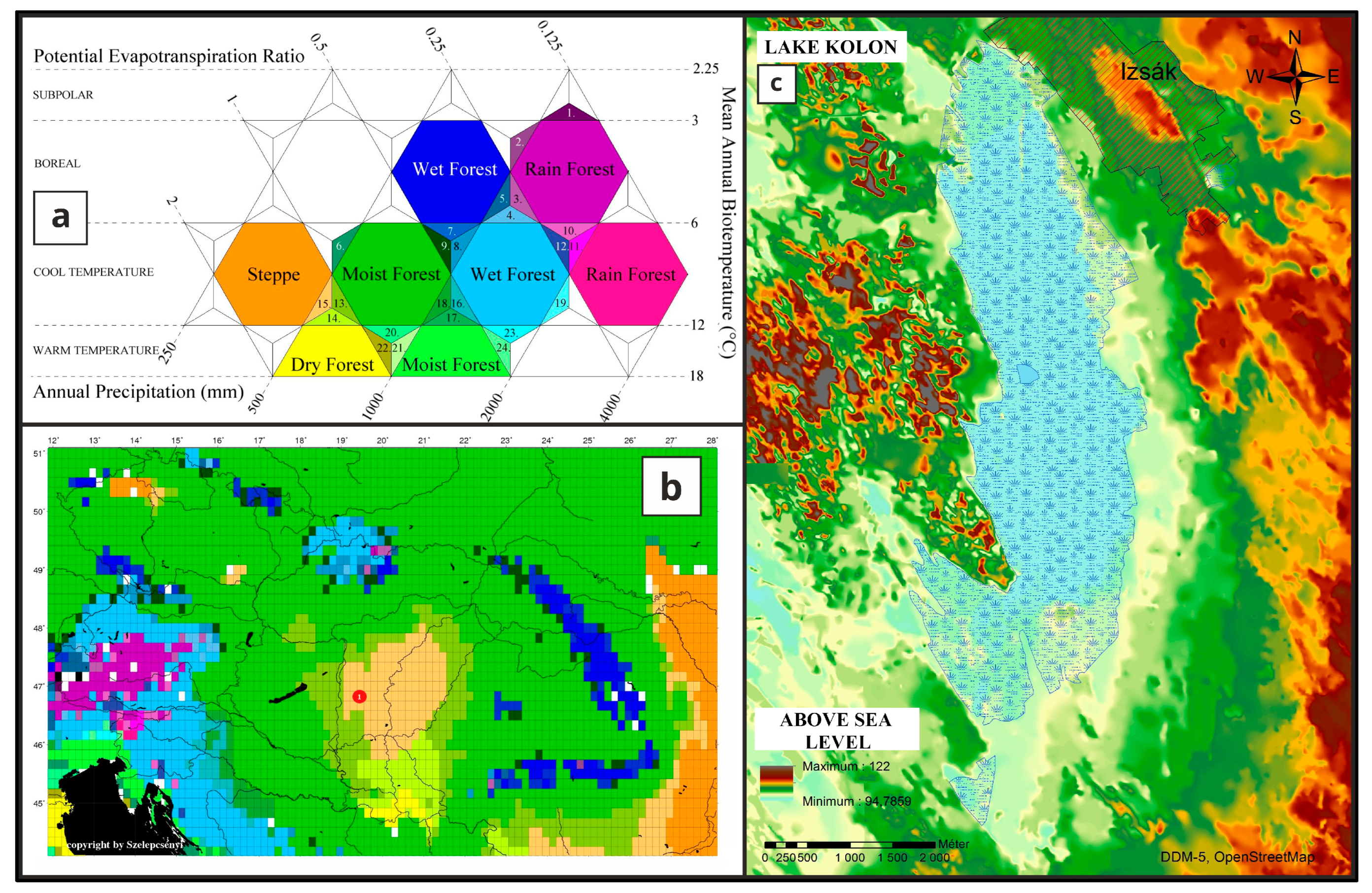
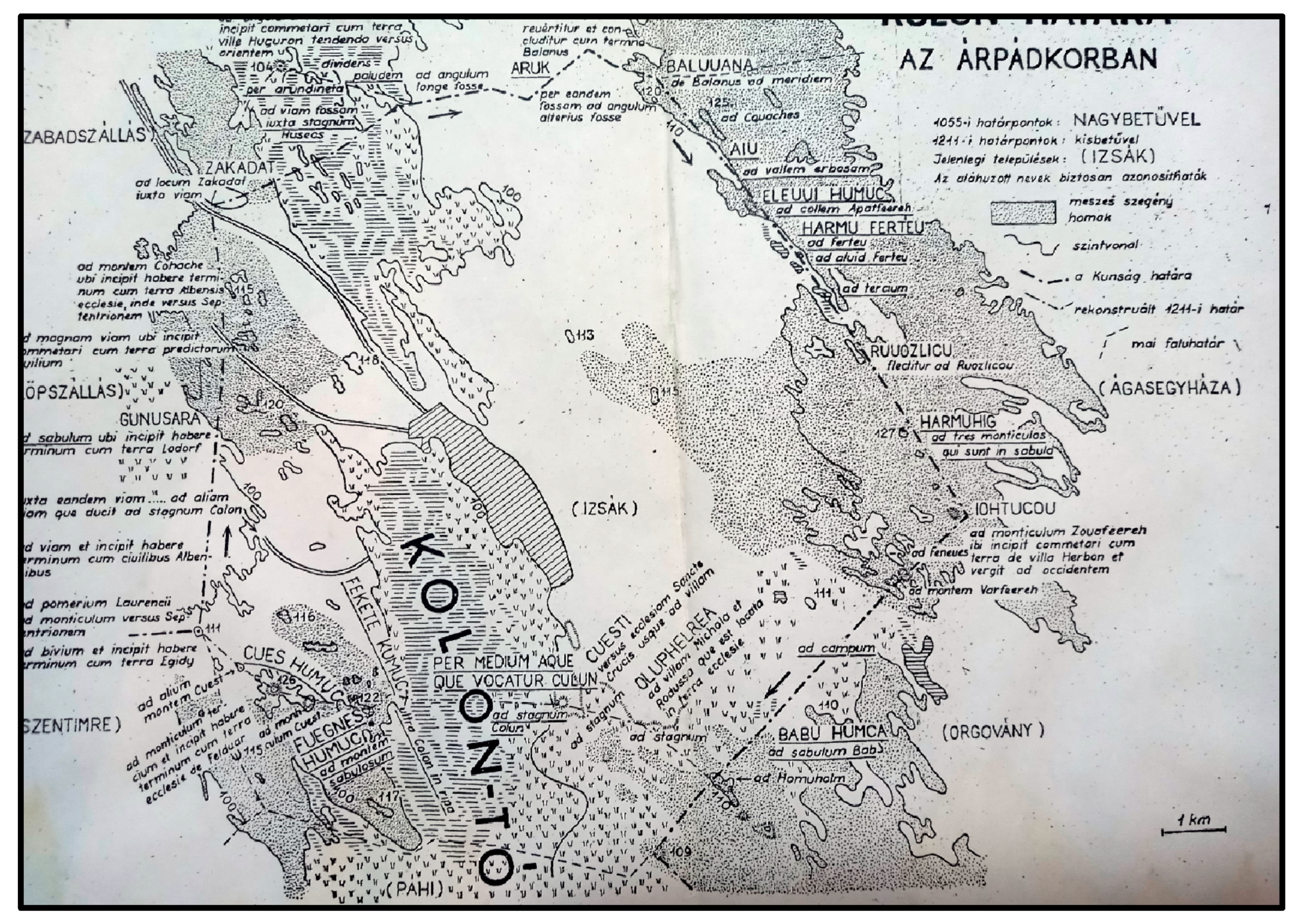
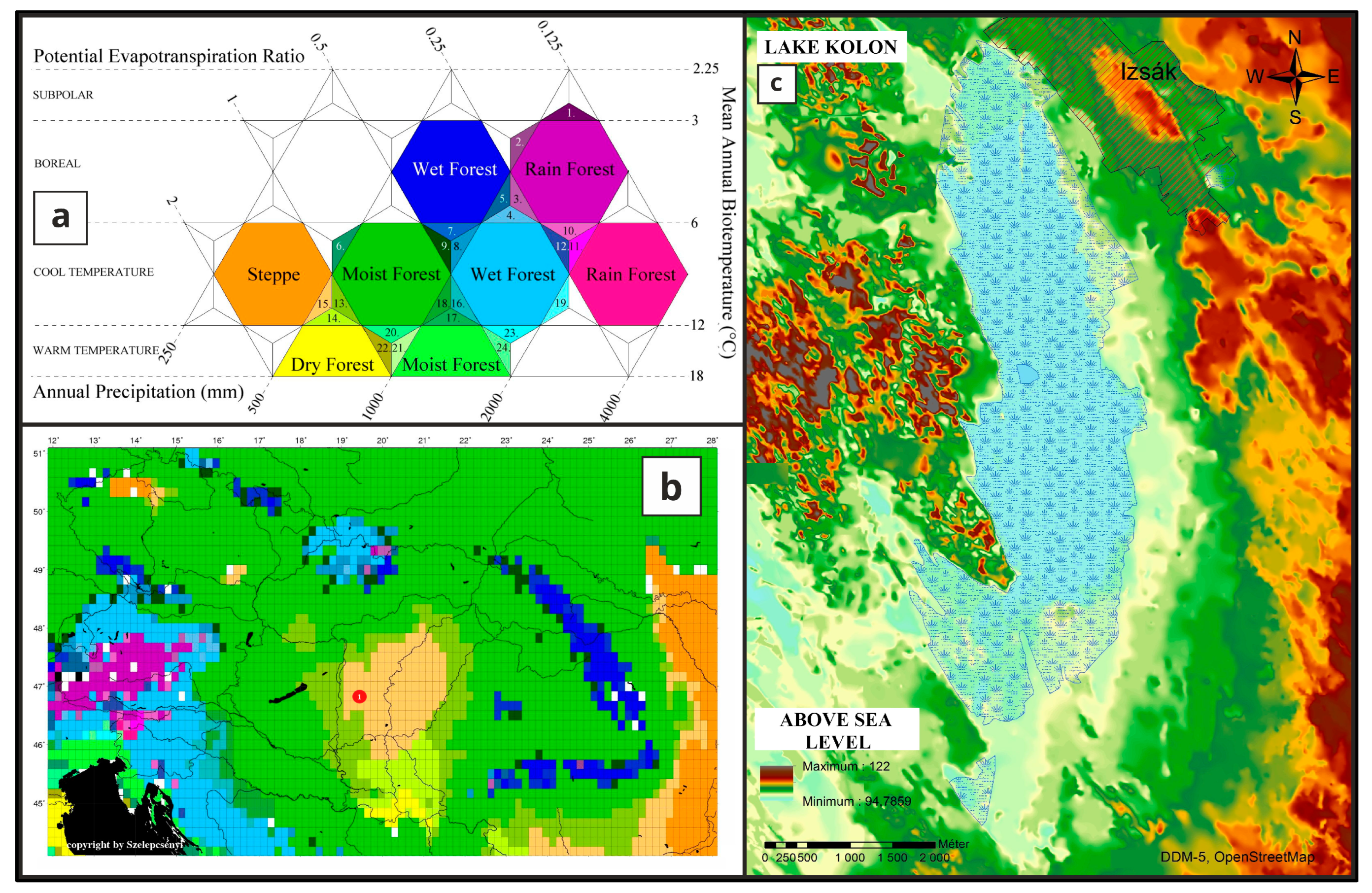
Study Area
2. Materials and Methods
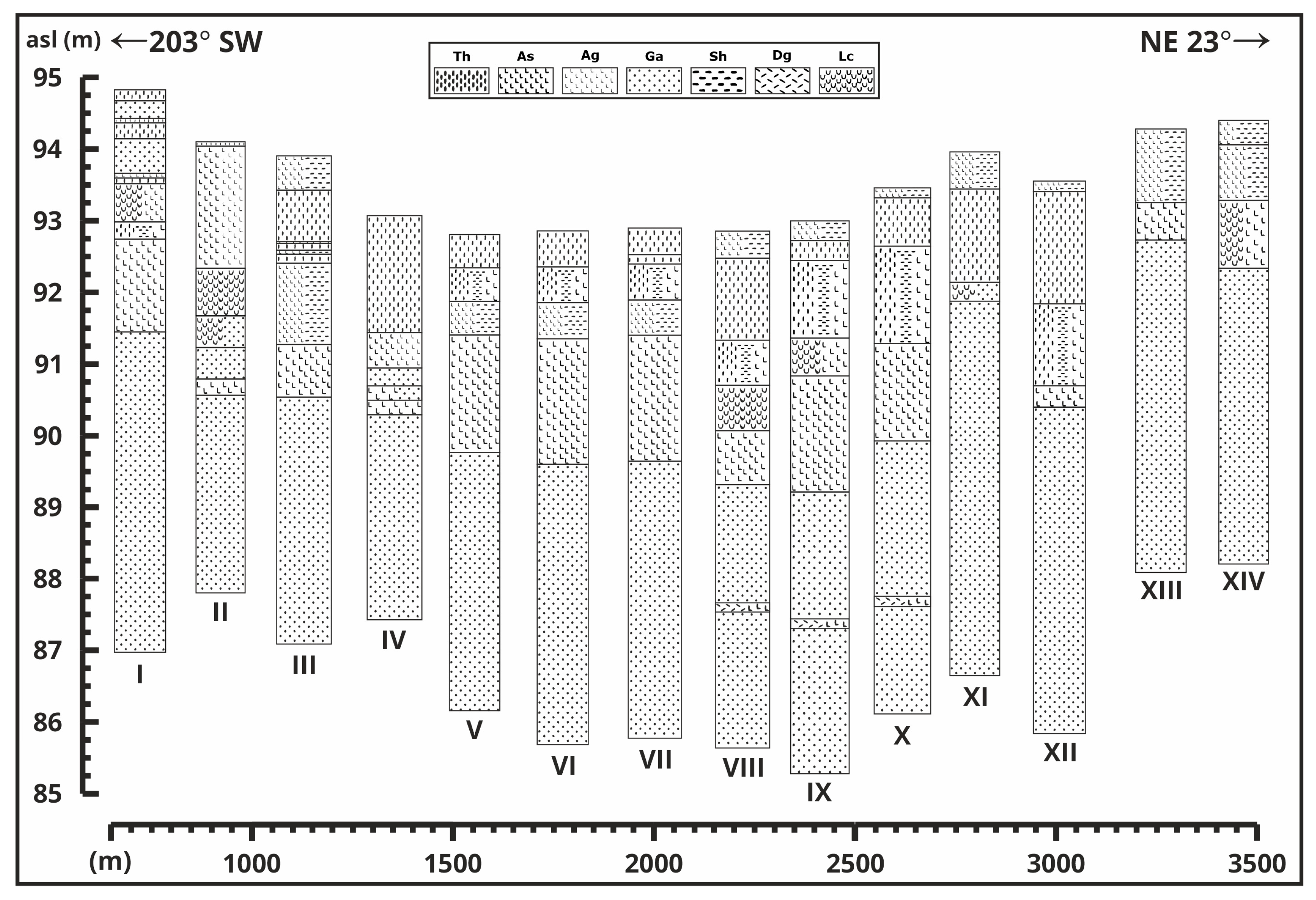
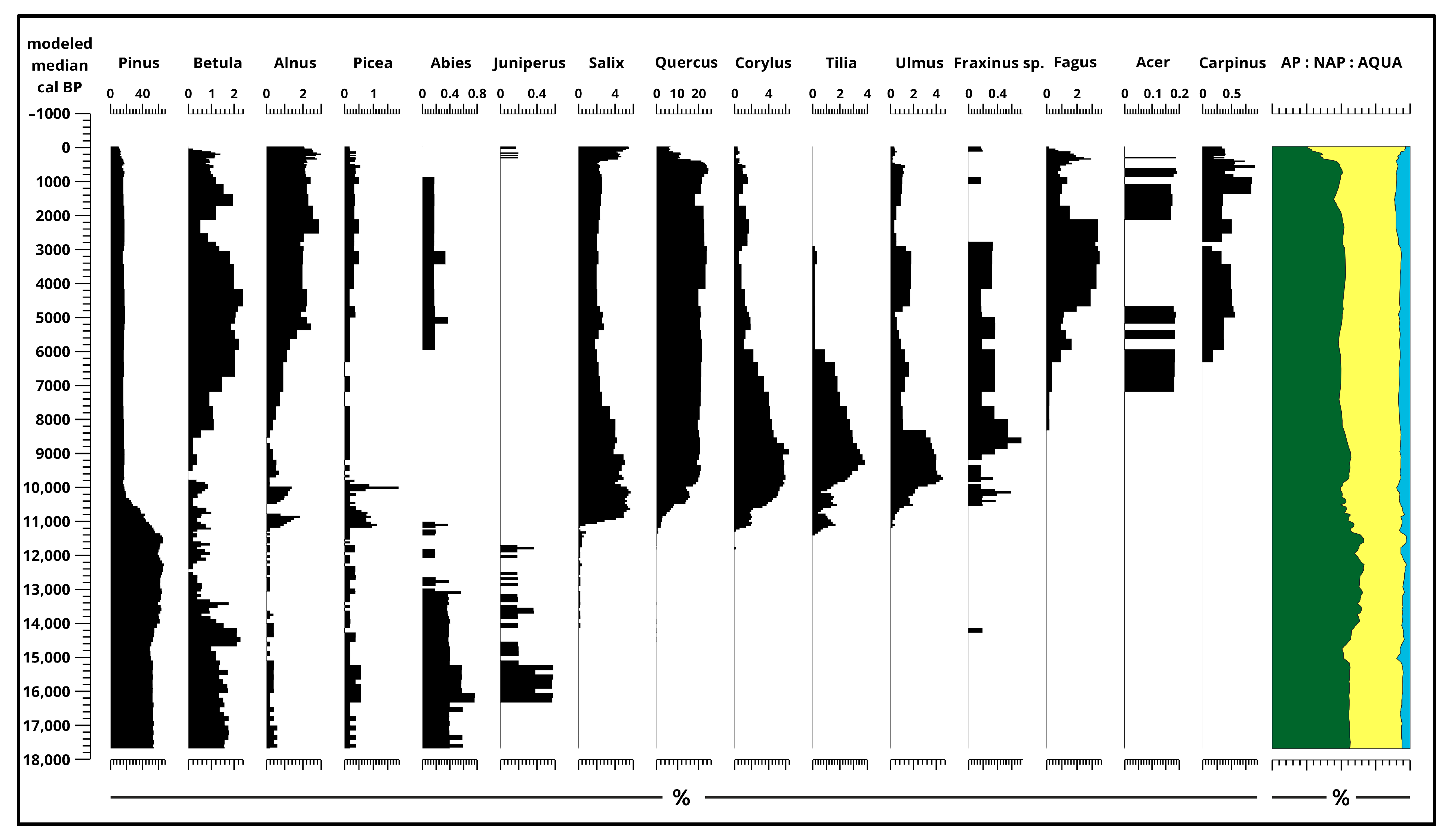
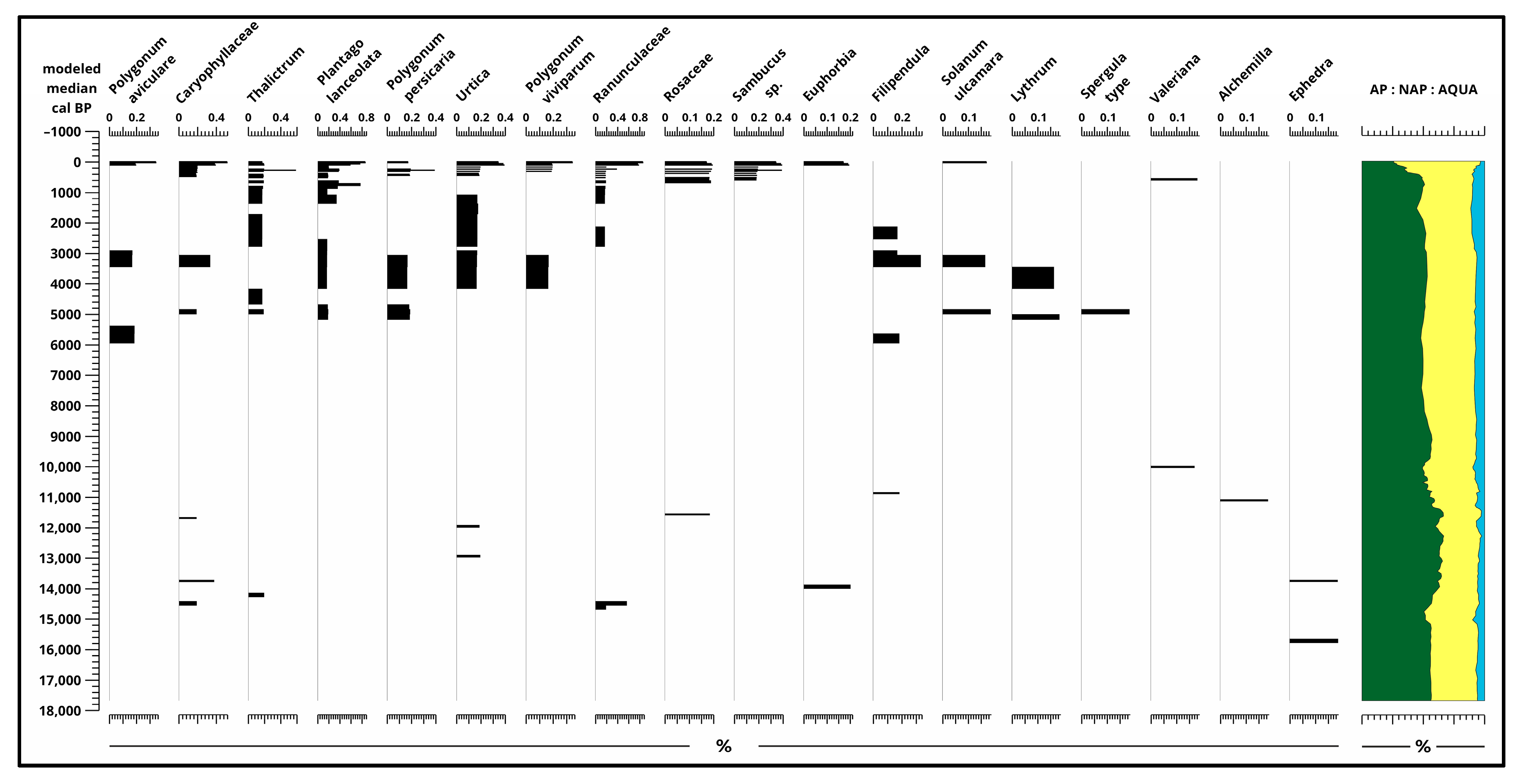
3. Results
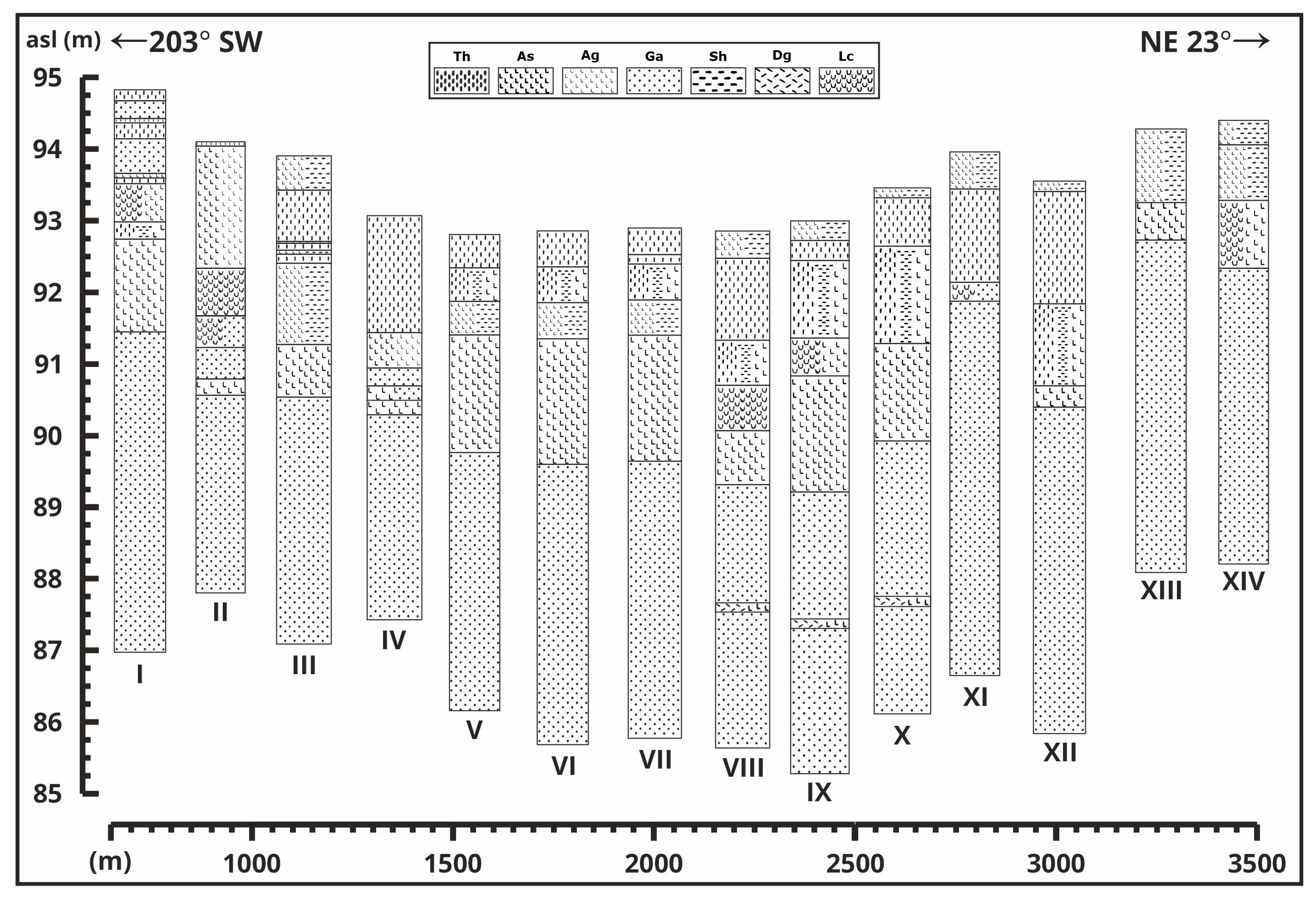
3.1. Cross-Sections
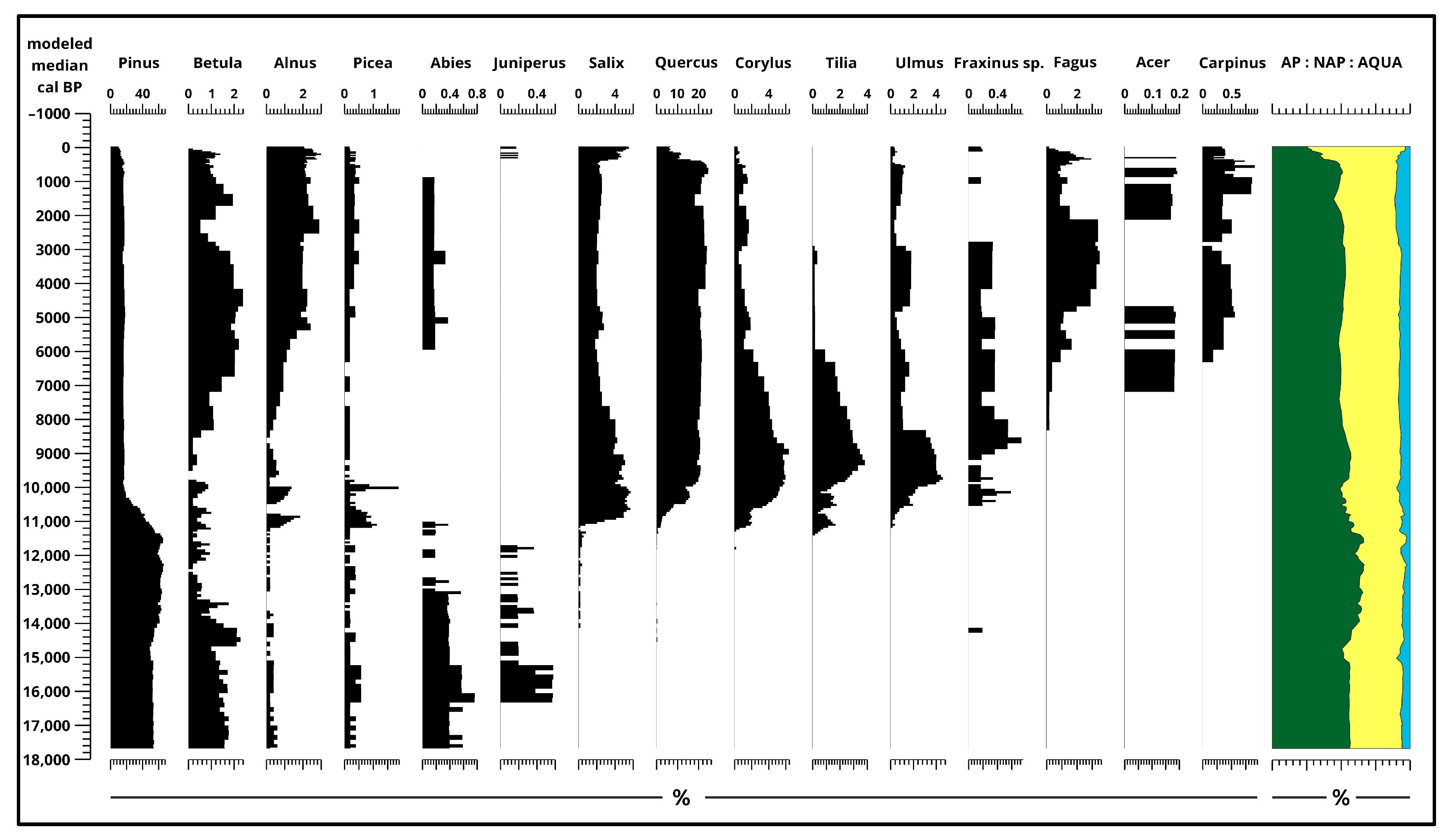
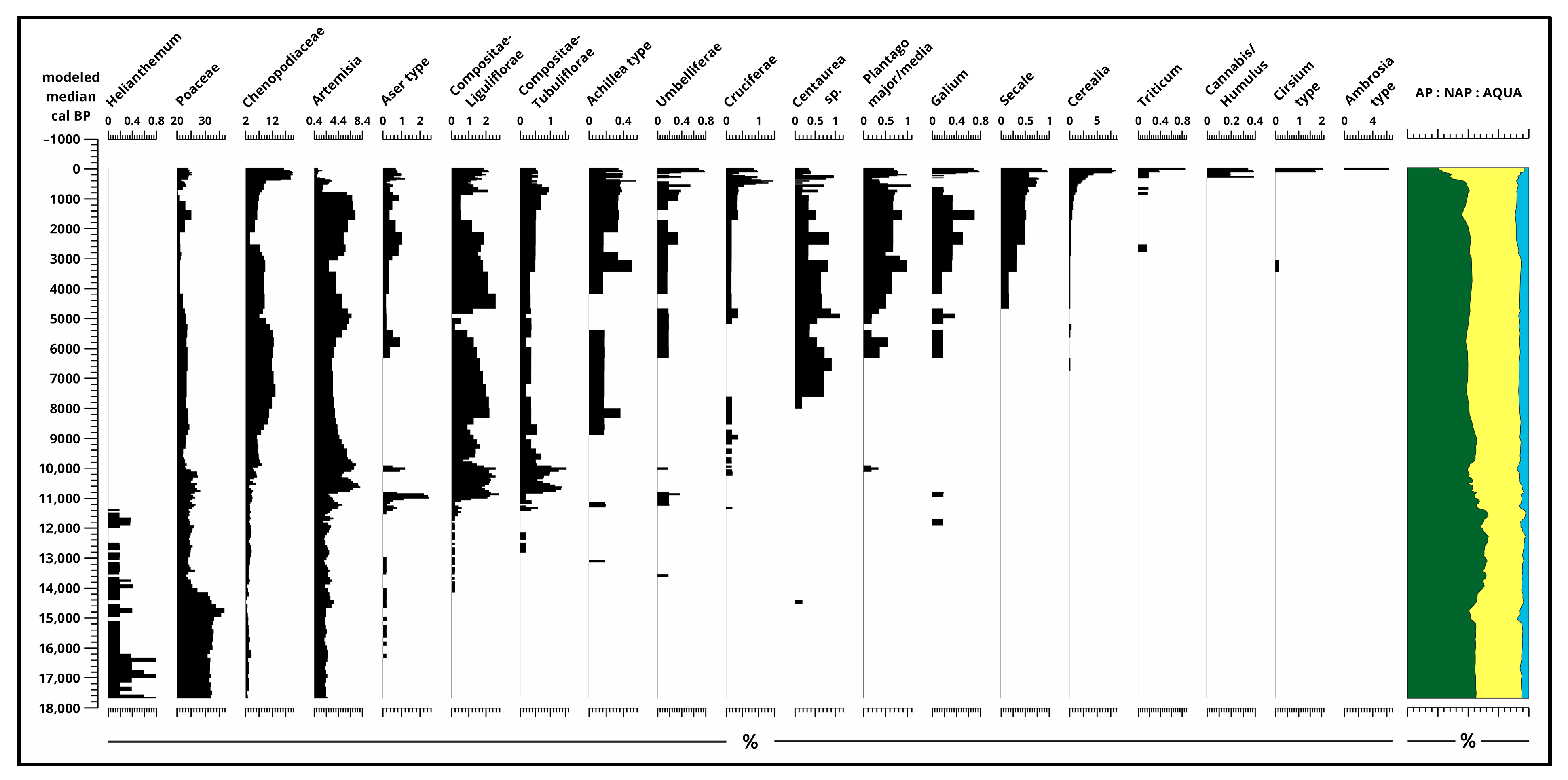
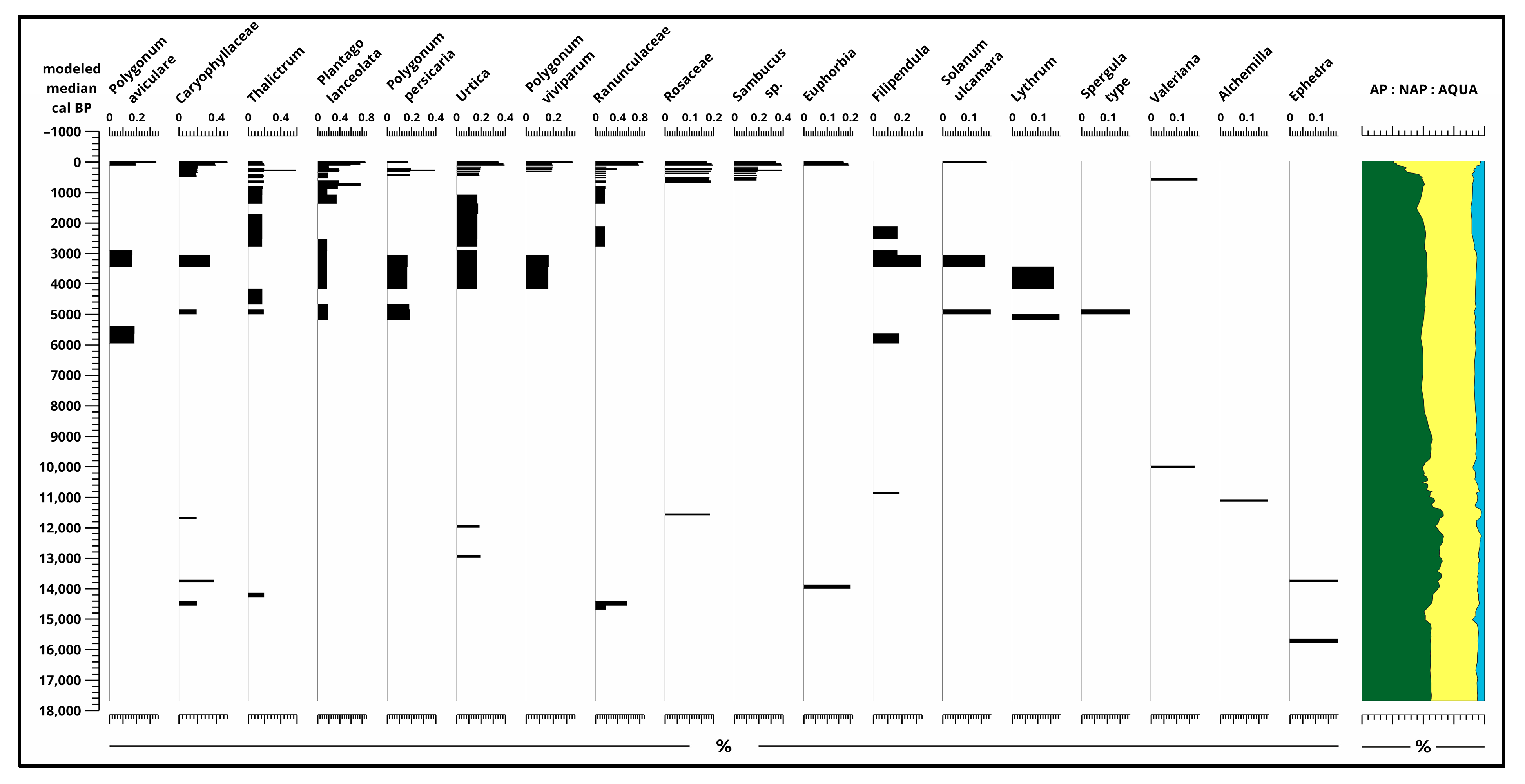
3.2. Pollen Analysis
3.3. Macrobotanical Analysis
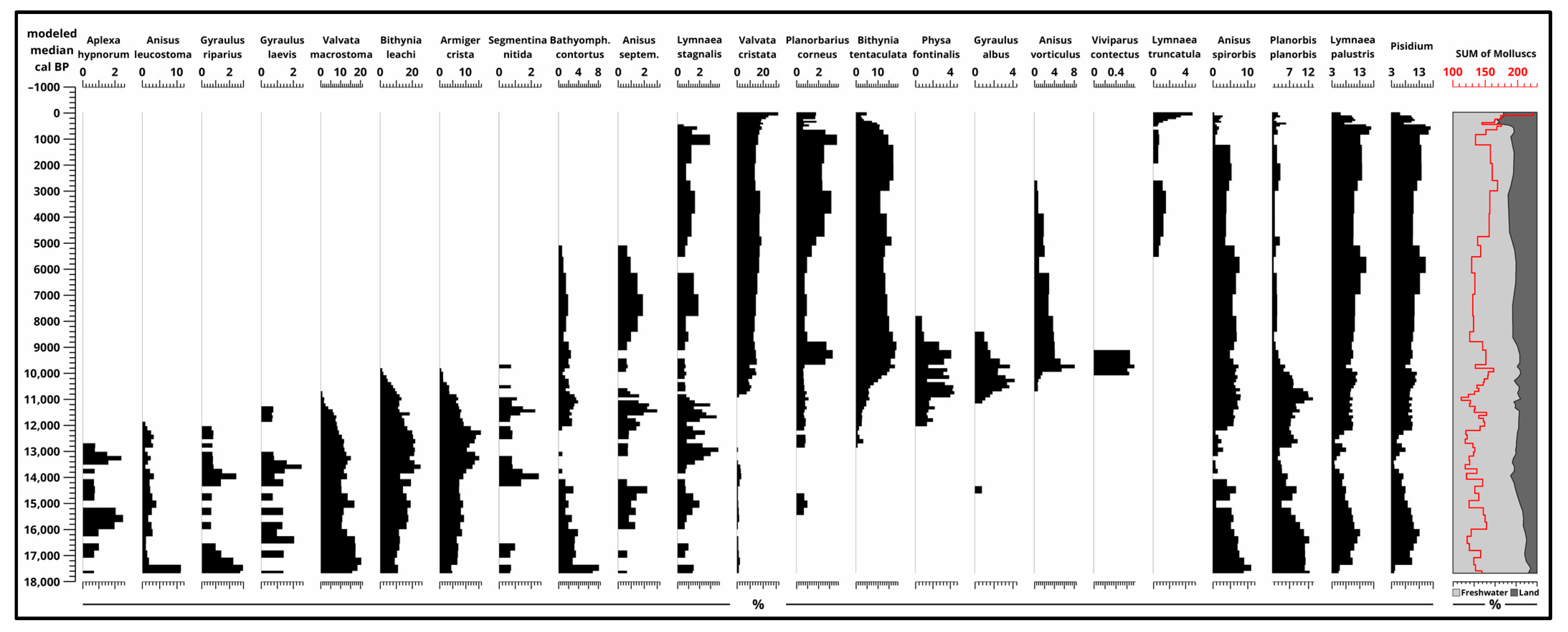
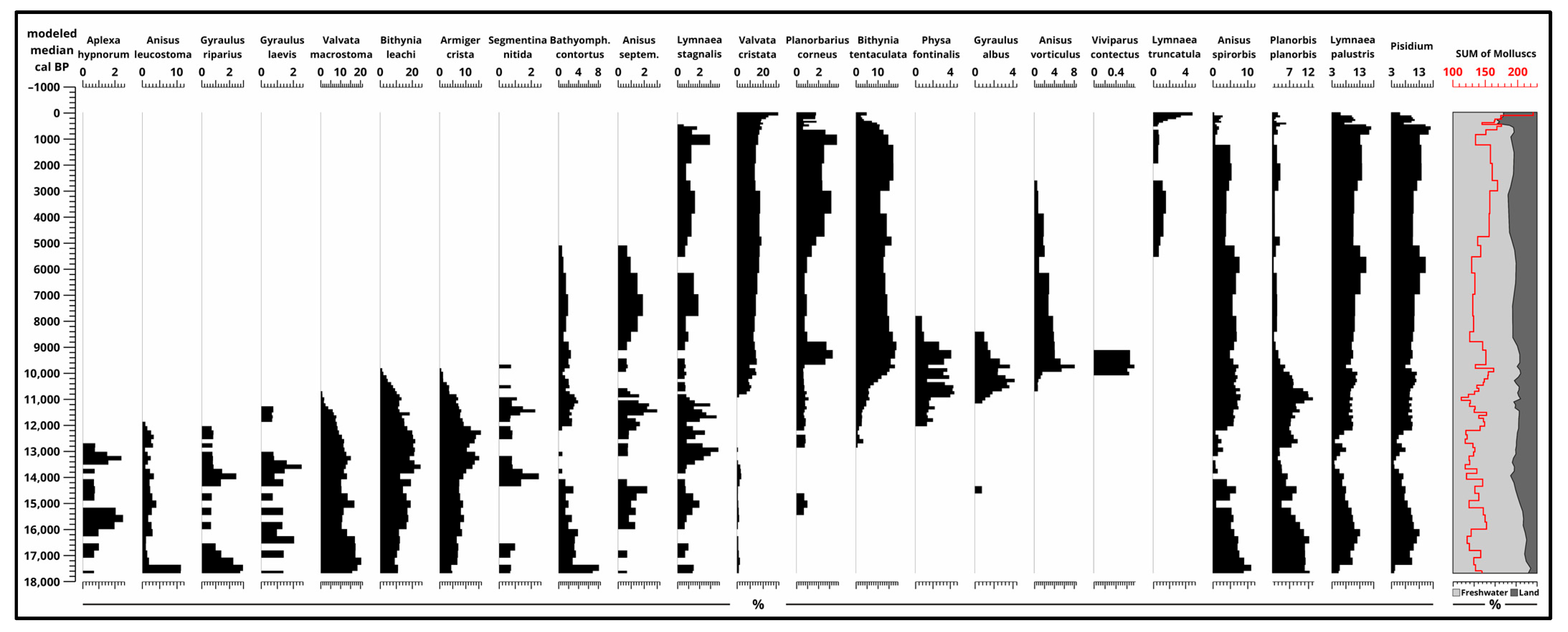
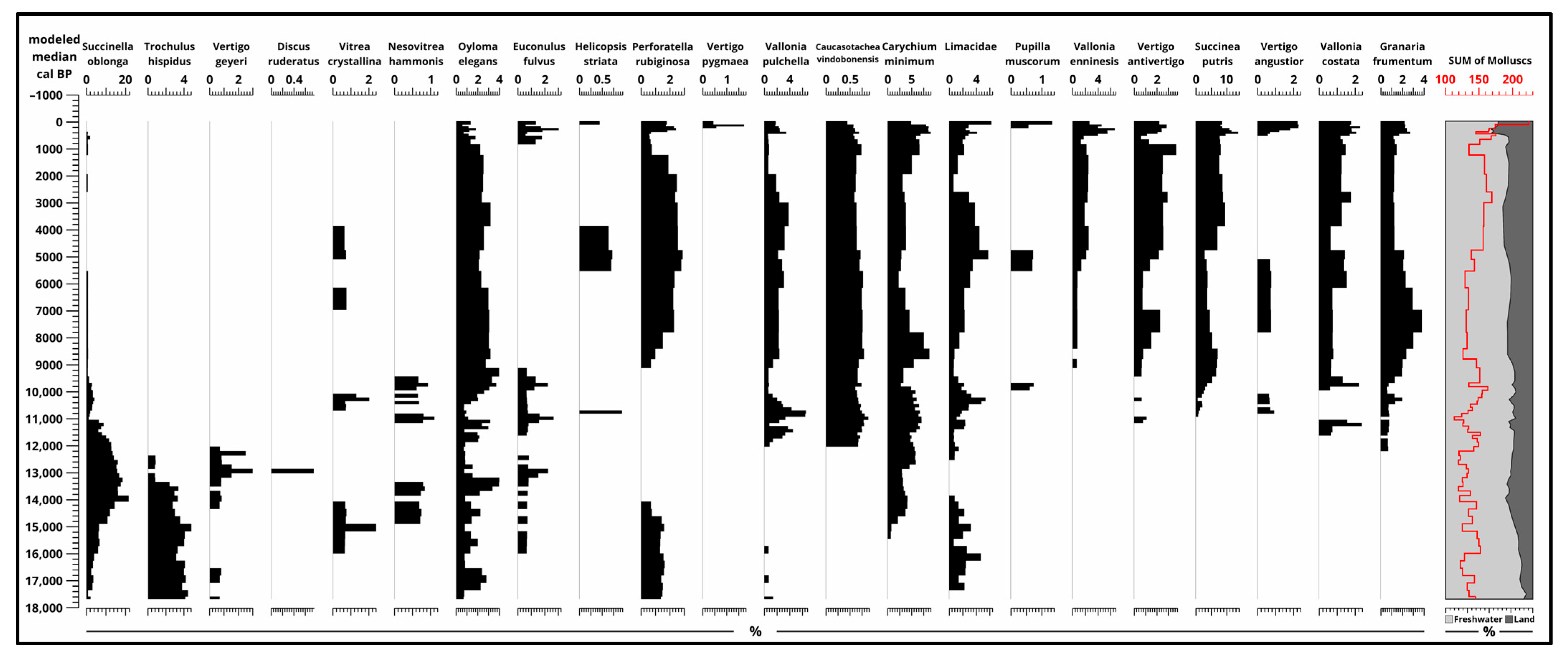
3.4. Malacological Analysis
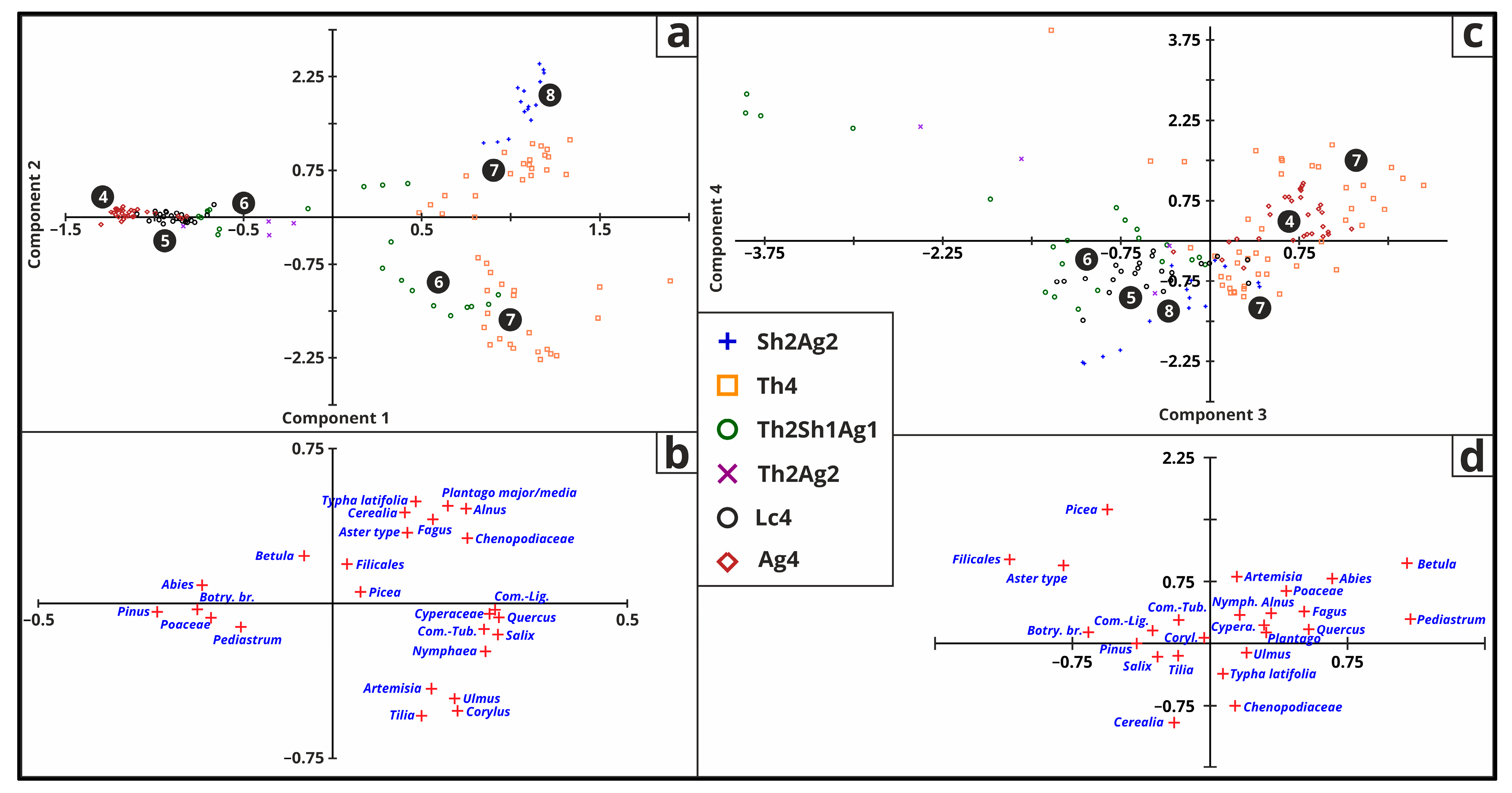
3.5. Statistical Analysis
3.5.1. Arbor Pollen
3.5.2. Non-Arbor Pollen
3.5.3. Aquatic Pollen
3.5.4. All Pollen
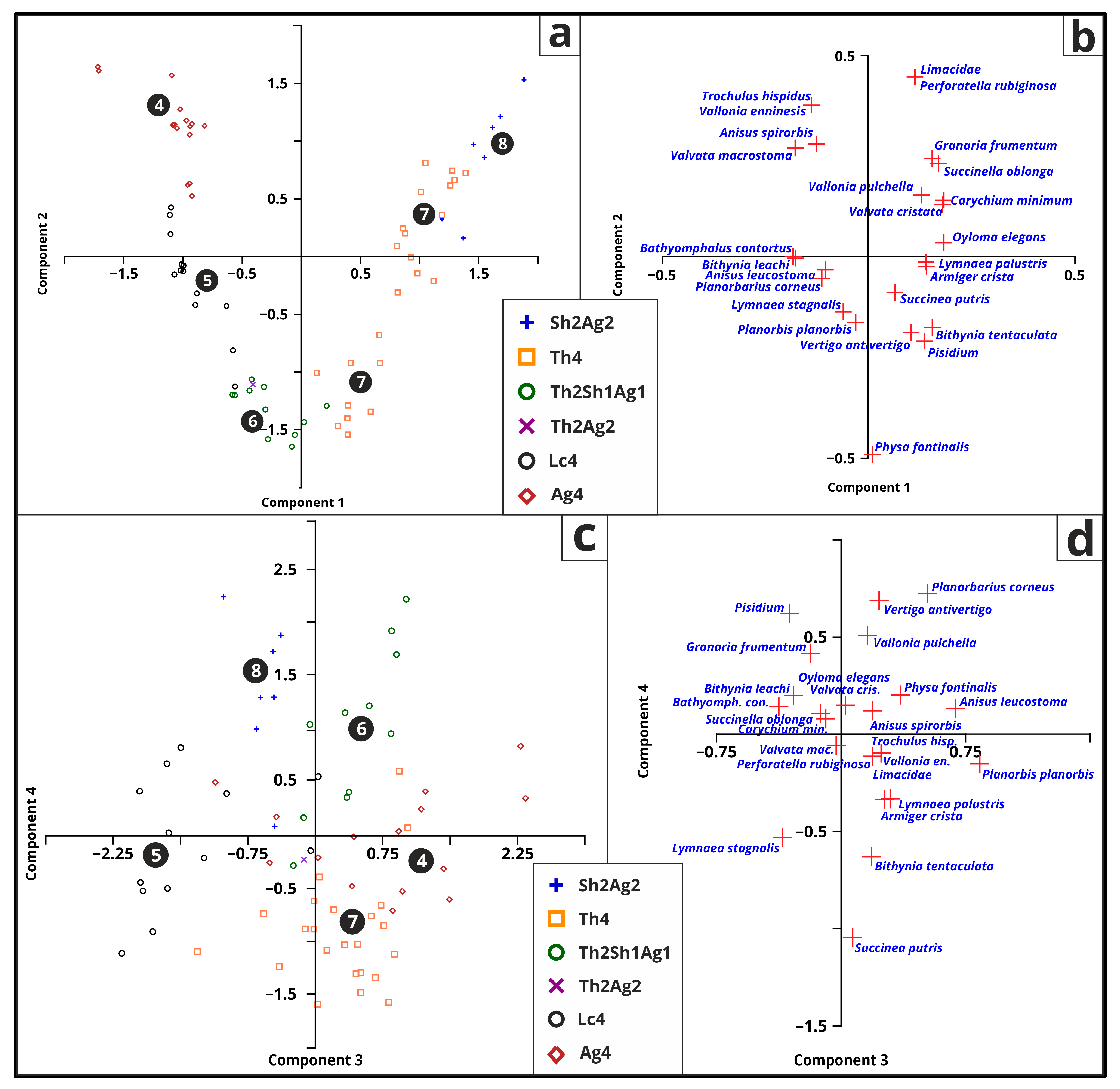
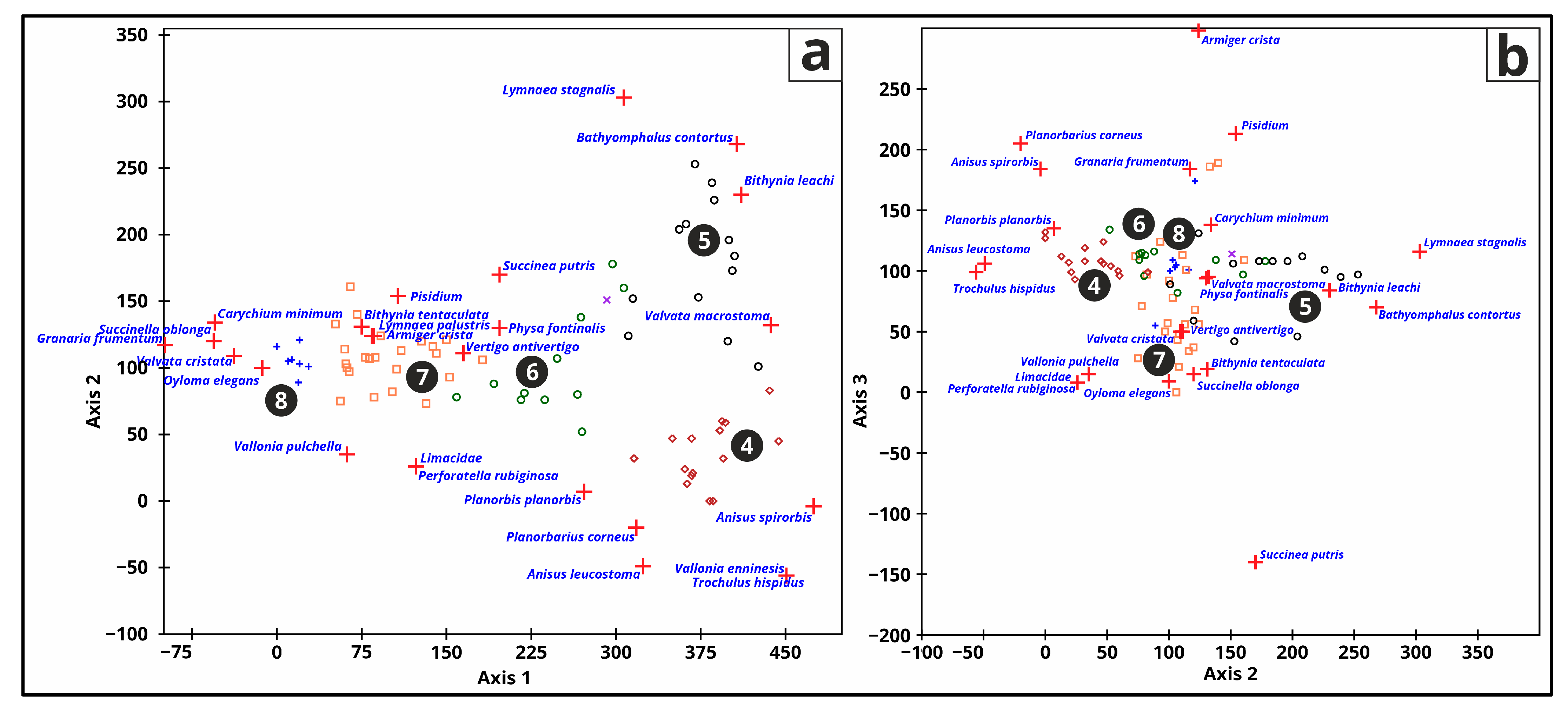
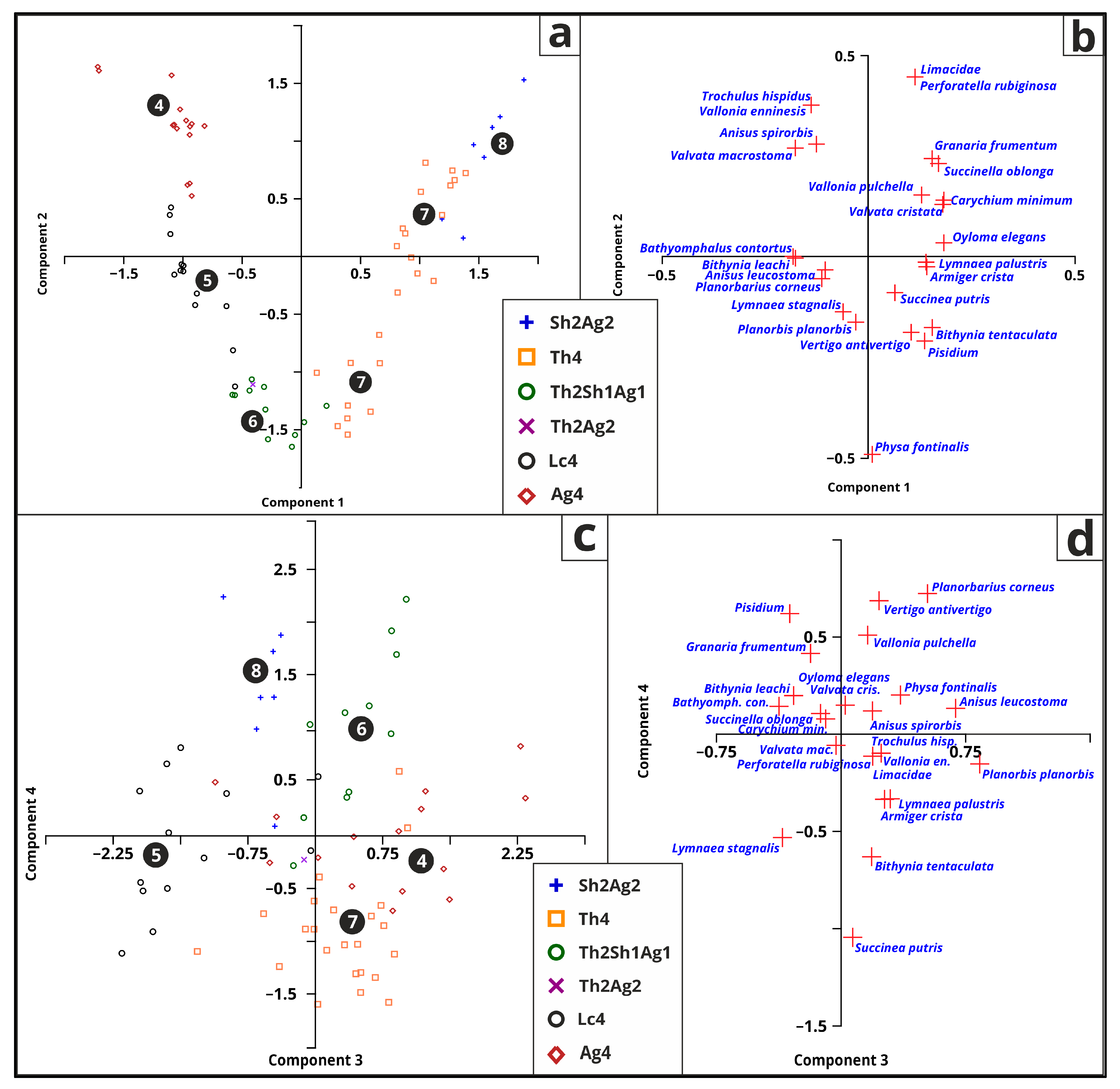
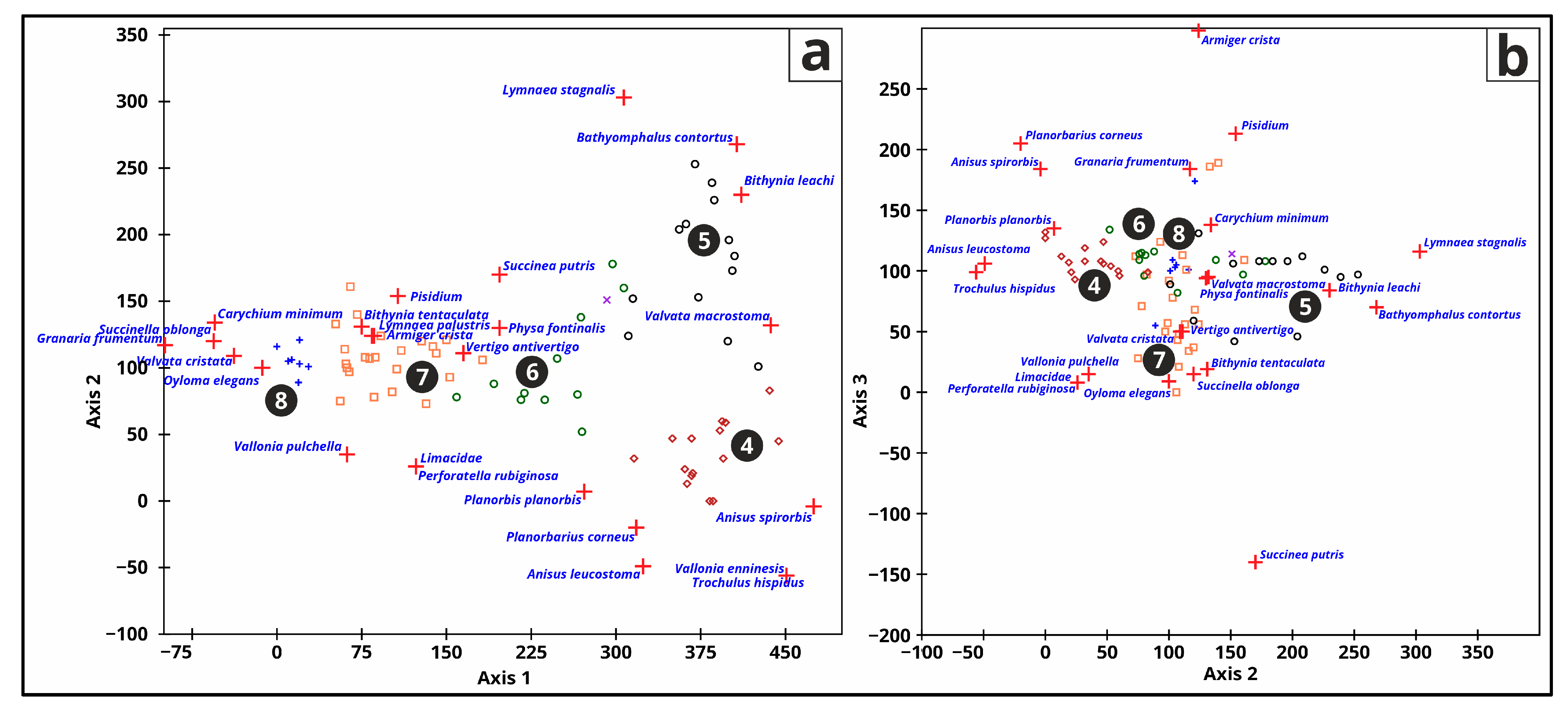
3.5.5. Malacology
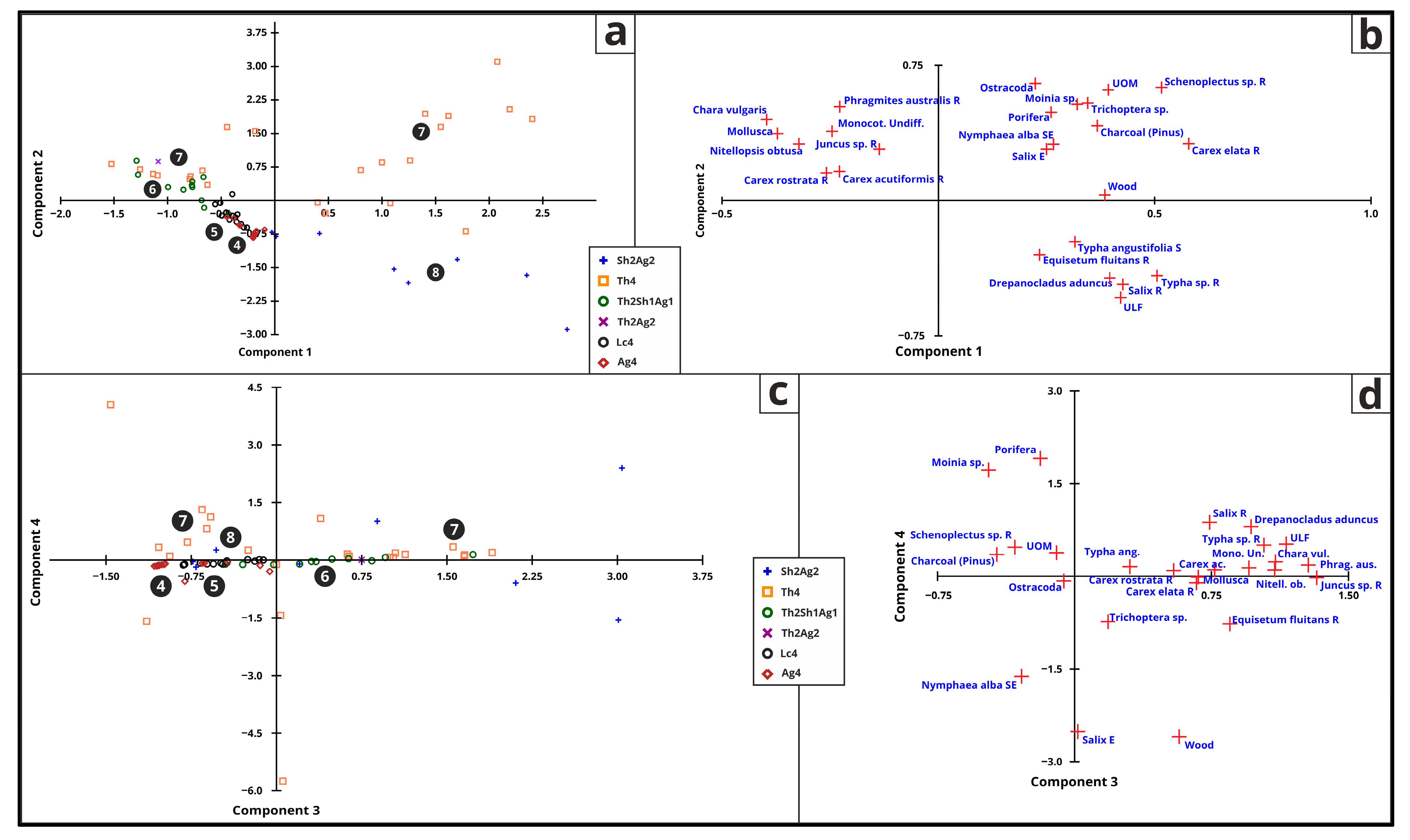
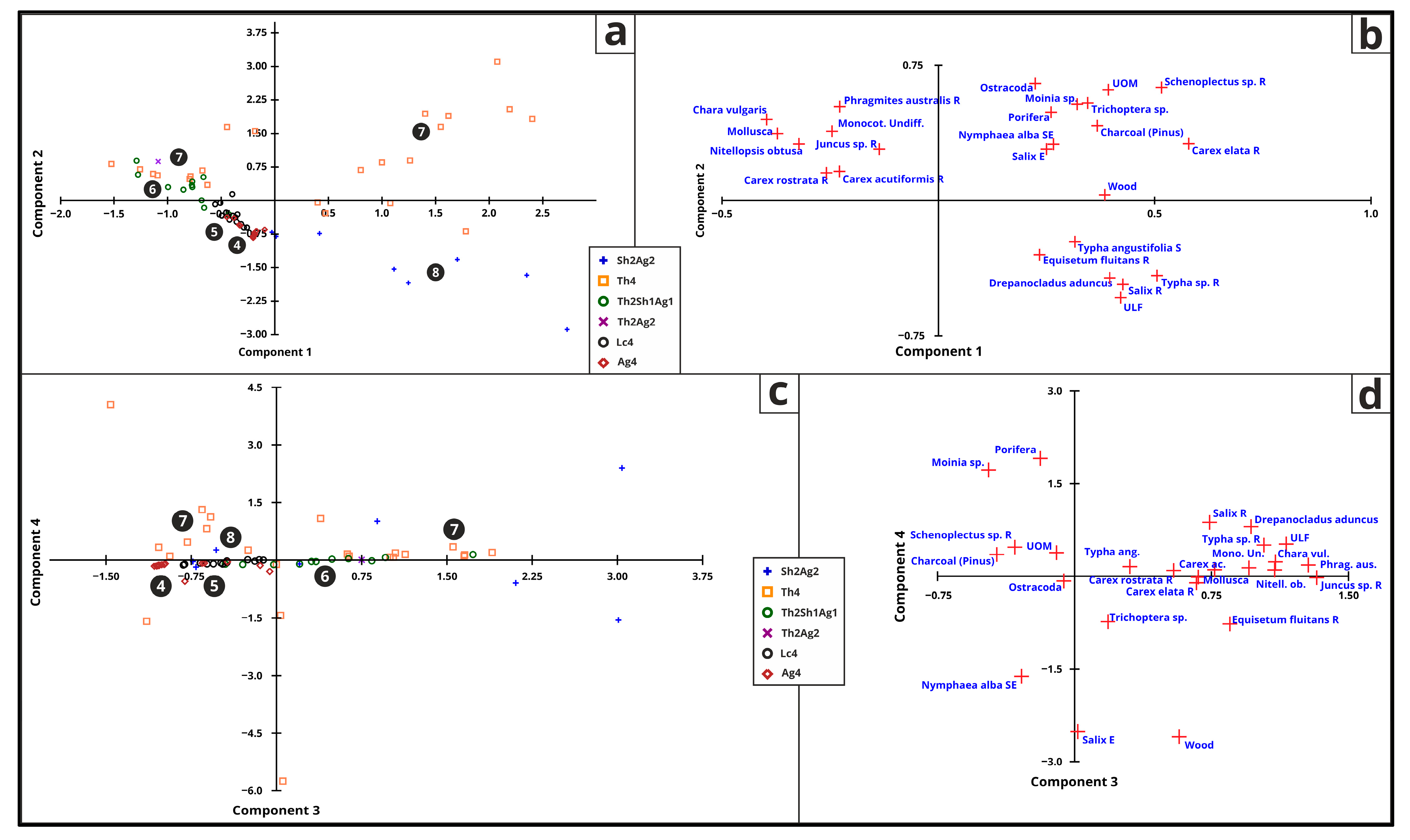
3.5.6. Macrofossil
4. Discussion
4.1. Paleoecological Changes
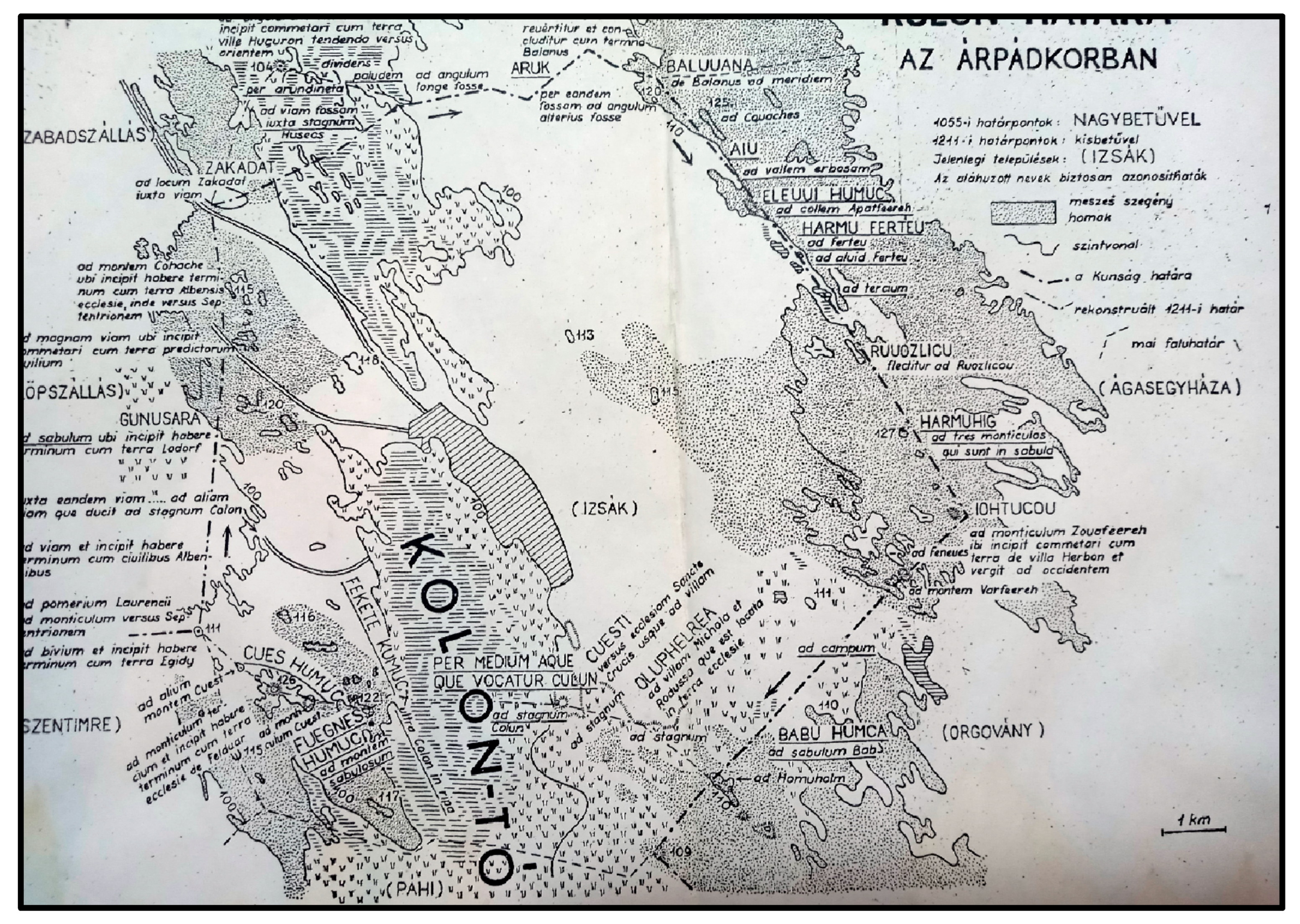
4.2. Medieval History
4.3. XX. Century History
4.4. Rehabilitation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Varga, Z.; Borhidi, A.; Fekete, G.; Debreczy, Z.; Bartha, D.; Bölöni, J.; Molnár, A.; Kun, A.; Molnár, Z.; Lendvai, G.; et al. Az erdőssztyepp fogalma, típusai és jellemzésük. In Alföld Erdőssztyepp Maradványok Magyarországon; Molnár, Z., Kun, A., Eds.; WWF Kiadvány: Budapest, Hungary, 2000; pp. 7–19. (In Hungarian) [Google Scholar]
- Vári, T.Z.; Gulyás, S.; Sümegi, P. Reconstructing the Paleoenvironmental Evolution of Lake Kolon (Hungary) through Integrated Geochemical and Sedimentological Analyses of Quaternary Sediments. Quaternary 2023, 6, 39. [Google Scholar] [CrossRef]
- Sümegi, P.; Molnár, M.; Jakab, G.; Persaits, G.; Majkut, P.; Páll, D.; Gulyás, S.; Jull, A.J.T.; Törőcsik, T. Radiocarbon-Dated Paleoenvironmental Changes on a Lake and Peat Sediment Sequence from the Central Great Hungarian Plain (Central Europe) During the Last 25,000 Years. Radiocarbon 2011, 53, 85–97. [Google Scholar] [CrossRef]
- Sümegi, P.; Molnár, D.; Náfrádi, K.; Makó, L.; Cseh, P.; Törőcsik, T.; Molnár, M.; Zhou, L. Vegetation and land snail-based reconstruction of the palaeocological changes in the forest steppe eco-region of the Carpathian Basin during last glacial warming. Glob. Ecol. Conserv. 2022, 33, e01976. [Google Scholar] [CrossRef]
- Magyari, E.K.; Chapman, J.C.; Passmore, D.G.; Allen, J.R.; Huntley, J.P.; Huntley, B. Holocene persistence of wooded steppe in the Great Hungarian Plain. J. Biogeogr. 2010, 37, 915–935. [Google Scholar] [CrossRef]
- Magyari, E.K.; Chapman, J.; Fairbairn, A.S.; Francis, M.; de Guzman, M. Neolithic human impact on the landscapes of North-East Hungary inferred from pollen and settlement records. Veg. Hist. Archaeobotany 2012, 21, 279–302. [Google Scholar] [CrossRef]
- Holdridge, L.R. Determination of world plant formations from simple climatic data. Science 1947, 105, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Holdridge, L.R. Life Zone Ecology; Tropical Science Center: San Jose, Costa Rica, 1967; p. 146. [Google Scholar]
- Szelepcsényi, Z.; Breuer, H.; Ács, F.; Kozma, I. Biofizikai klímaklasszifikációk. Légkör 2009, 54, 21–26. [Google Scholar]
- Szelepcsényi, Z.; Breuer, H.; Sümegi, P. The climate of Carpathian Region in the 20th century based on the original and modified Holdridge life zone system. Cent. Eur. J. Geosci. 2014, 6, 293–307. [Google Scholar] [CrossRef]
- Szelepcsényi, Z.; Breuer, H.; Kis, A.; Pongrácz, R.; Sümegi, P. Assessment of projected climate change in the Carpathian Region using the Holdridge life zone system. Theor. Appl. Climatol. 2018, 31, 1–18. [Google Scholar] [CrossRef]
- Piti, F. A tihanyi monostor alapítólevele (1055). In Írott Források az 1050-1116 Közötti Magyar Történelemről; Makk, F., Thoroczkay, G., Eds.; Szegedi Középkortörténeti Könyvtár: Szeged, Hungary, 2006; pp. 16–25. (In Hungarian) [Google Scholar]
- Molnár, B. The Geology and Hydrogeology of Kiskunsági (Little Cumanian) National Park; JATE Press: Szeged, Hungary, 2015; p. 534. (In Hungarian) [Google Scholar]
- Molnár, B.; Iványosi-Szabó, A.; Fényes, J. Die Entstehung des Kolon-Sees und seine limnogeologische Entwicklung. Hidrol. Közlöny 1979, 59, 549–560, (In Hungarian with German Summary). [Google Scholar]
- Tölgyesi, I. The Flora of the Kolon Lake and Its Environs at Izsák (Hungary). Ph.D. Thesis, Eötvös Lóránd University, Budapest, Hungary, 1981; p. 95. (In Hungarian). [Google Scholar]
- Szujkó-Lacza, J. The Flora of the Kiskunság National Park; Magyar Természet-Tudományi Múzeum Kiadványa: Budapest, Hungary, 1993; p. 467. [Google Scholar]
- Vadász, C. The Effect of Reedbed Management on the Breeding Passerine Assemblage. Ph.D. Thesis, Eötvös Loránd University, Budapest, Hungary, 2009; p. 120, (In Hungarian with English Summary). [Google Scholar]
- Hollósi, A.; Biró, C.; Biró, M.; S.-Falusi, E. Landscape historical aspects and macropyte monitoring of the habitat restoration on the Lake Kolon at Izsák. Hidrol. Közlöny 2015, 95, 100–101. [Google Scholar]
- Zólyomi, B. Budapest és környékének természetes növénytakarója. In Budapest Természeti Képe; Pécsi, M., Marosi, S., Szilárd, J., Eds.; Akadémiai Kiadó: Budapest, Hungary, 1958; pp. 509–642. [Google Scholar]
- Zólyomi, B.; Kéri, M.; Horváth, F. A szubmediterrán éghajlati hatások jelentősége a Kárpát-medence klímazonális növénytársuásainak összetételére. In Hegyfoky Kabos Klimatológus Születésének 145. Évfordulája Alkalmából Rendezett Tudományos Emlékülés Előadásai; Tar, K., Ed.; MTA Debreceni Területi Bizottságának Kiadványa: Debrecen-Túrkeve, Hungary, 1992; pp. 60–74. [Google Scholar]
- Zólyomi, B.; Fekete, G. The Pannonian loess steppe: Differentation in space and time. Abstr. Bot. 1994, 18, 29–41. [Google Scholar]
- Belokopytov, I.E.; Beresnevich, V.V. Giktorf’s peat borers. Torfyanaya Promyfhlennost 1955, 8, 9. [Google Scholar]
- Aaby, B.; Digerfeldt, G. Sampling techniques for lakes and mires. In Handbook of Holocene Palaeoecology and Palaeohydrology; Berglund, B.E., Ed.; John Wiley & Sons: Chichester, UK, 1986; pp. 181–194. [Google Scholar]
- Vleeschouwer, F.; Chambers, F.; Swindles, G. Coring and sub-sampling of peatlands for palaeoenvironmental research. Mires Peat 2010, 7, 10. [Google Scholar]
- Eriksson, L.; Johansson, E.; Kettaneh-Wold, N.; Wold, S. Introduction to Multi- and Megavariate Data Analysis Using Projection Methods (PCA & PLS); Umetrics AB: Umea, Sweden, 1999. [Google Scholar]
- Troels-Smith, J. Characterization of unconsolidated sediments. Dan. Geol. Unders. 1955, 4, 10. (In Danish) [Google Scholar]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9–18. [Google Scholar]
- Davis, J.C. Statistics and Data Analysis in Geology; John Wiley & Sons Inc.: New York, NY, USA, 1986; p. 656. [Google Scholar]
- Bánffy, E. The Early Neolithic in the Danube-Tisza Interfluve; British Archaeological Reports; Archaeopress: Oxford, UK, 2013; Volume 2584, p. 189. [Google Scholar]
- Behre, K.E. Evidence for Mesolithic agriculture in and around central Europe? Veg. Hist. Archaeobotany 2007, 16, 203–219. [Google Scholar] [CrossRef]
- Sümegi, P. The results of paleoenvironmental reconstruction and comparative geoarcheological analysis for the examined area. In The Geohistory of Bátorliget Marhsland; Sümegi, P., Gulyás, S., Eds.; Archaeolingua Press: Budapest, Hungary, 2004; pp. 301–348. [Google Scholar]
- Koszta, L.; Capitulum, I. Fejezetek a Középkori Magyar Egyház Történetéből; Szegedi Középkorász Műhely: Szeged, Hungary, 1998; p. 183. (In Hungarian) [Google Scholar]
- Kőfalvi, T. A hiteles helyi oklevelek egyháztörténeti tanulságai. Egyháztörténeti Szemle 2000, 1, 49–64. (In Hungarian) [Google Scholar]
- Reszegi, K. Hegynevek a Középkori Magyarországon. Ph.D. Thesis, University of Debrecen, Debrecen, Hungary, 2008; p. 180. (In Hungarian). [Google Scholar]
- Gyárfás, I. A Jász-Kunok Története. III. Kötet (Oklevéltár); Bakos Nyomda: Szolnok, Hungary, 1883; pp. 498–500, 578–580, 626–651. (In Hungarian) [Google Scholar]
- Hornyik, J. Kecskemét Város Története, Oklevéltárral. 1. Kötet. Kecskemét, Szilády Károly, Hungary. 1860, pp. 201–206, 222–225. (In Hungarian). Available online: http://real-eod.mtak.hu/5588/1/000909780.pdf (accessed on 10 October 2023).
- Bártfai-Szabó, L. Pest Megye Történetének Okleveles Emlékei 1002-1599-ig: Függelékül az Inárchi Farkas, az Irsai Irsay, Valamint a Szilasi és Pilisi Szilassy Családok Története; Ablaka János és Ferenc Nyomdája: Gyöngyös, Hungary, 1938; p. 636. (In Hungarian) [Google Scholar]
- Györffy, G. A tihanyi alapítólevél földrajzinév-azonosításához. In Emlékkönyv Pais Dezső Hetvenedik Születésnapjára; Bárczi, G., Benkő, L., Eds.; Akadémiai Kiadó: Budapest, Hungary, 1956; pp. 407–415. (In Hungarian) [Google Scholar]
- Holler, L. Az 1055. évi tihanyi oklevélben említett két birtok lokalizálása. Javaslat a lacus segisti és a bagat mezee határú birtok elhelyezkedésére. Helynévtörténeti Tanulmányok 2010, 5, 47–82. (In Hungarian) [Google Scholar]
- Miháltz, I. Duna-Tisza-közi futóhomok. Földtani Értesítő 1938, 3, 1–8. (In Hungarian) [Google Scholar]
- Miháltz, I. A Duna-Tisza Köze Déli Részének Földtani Felvétele; MÁFI Évi Jelentése 1950; Magyar Állami Földtani Intézet: Budapest, Hungary, 1953; pp. 113–144. (In Hungarian) [Google Scholar]
- Miháltz, I.; Mucsi, M. A kiskunhalasi Kunfehértó hidrogeológiája. Hidrol. Közlöny 1964, 44, 463–471. (In Hungarian) [Google Scholar]
- Molnár, B. A Duna-Tisza közi eolikus rétegek felszíni és felszínalatti kiterjedése. Földtani Közlöny 1961, 91, 303–315. (In Hungarian) [Google Scholar]
- Tóber, M. “Sárga homokdombok emelkednek miket épít s dönt a szélvész…” homoktípusok homokformák futóhomok a középkori homokhátság területén az egykorú írott források tükrében, V.I. In Proceedings of the Magyar Földrajzi Konferencia, Szeged, Hungary, 5–7 September 2012. (In Hungarian). [Google Scholar]
- HTóth, E. The equestrian grave of Izsák—Balázspuszta from the period of the Hungarian Conquest. Cumania 1976, 4, 141–173. [Google Scholar]
- Parádi, N. Pénzleletekkel keletezett XIII. századi ékszerek. Folia Archaeol. 1975, 11, 119–157. (In Hungarian) [Google Scholar]
- Treitz, P. A Duna-Tisza közének agrogeolgóiai leírása. Földtani Közlöny 1903, 33, 297–316. (In Hungarian) [Google Scholar]
- Treitz, P. Homok Vizsgálatok. Magyar Királyi Földtani Intézet Évi Jelentése az 1916-ról; Magyar Állami Földtani Intézet: Budapest, Hungary, 1917; pp. 113–144. (In Hungarian) [Google Scholar]
- Kádár, L. Futóhomok-tanulmányok a Duna-Tisza-közén. Földrajzi Közlemények 1935, 63, 4–15. (In Hungarian) [Google Scholar]
- Borsy, Z. A szélerózió vizsgálata a magyarországi futóhomok területeken. Földrajzi Közlemények 1972, 20, 156–160. (In Hungarian) [Google Scholar]
- Stefanovits, P.; Filep, G.; Füleky, G. Talajtan; Mezőgazda Kiadó: Budapest, Hungary, 2010; p. 415. (In Hungarian) [Google Scholar]
- Kiss, I. Szikes Területek Alga-Tömegprodukciós Jelzései a Foltos Regradáció Vízfeltöréses Folyamatáról; Szegedi Tanárképzős Főiskola Tudományos Közleményei: Szeged, Hungary, 1969; Volume 3. (In Hungarian) [Google Scholar]
- Molnár, B.; Makádi, M. A Duna—Tisza fejlődéstörténete. Iskolakultúra Pedagógusok Szakmai-Tudományos Folyóirata 1995, 5, 119–129. (In Hungarian) [Google Scholar]
- Hoffmann, I. A Tihanyi Alapítólevél, Mint Helynévtörténeti Forrás. Ph.D. Thesis, University of Debrecen, Debrecen, Hungary, 2007; p. 306. (In Hungarian). [Google Scholar]
- Iványosi-Szabó, A. A Kiskunsági Nemzeti Park Igazgatóság 40 Éve; Kiskunsági Nemzeti Park Igazgatóságának Kiadványa: Kecskemét, Hungary, 2015; p. 424. (In Hungarian) [Google Scholar]
- Bálint, M. Az Árpád-Kori Településhálózat Rekonstrukciója a Dorozsma-Majsai Homokhát területén. Ph.D. Thesis, Eötvös Lóránd University, Budapest, Hungary, 2007; p. 414. [Google Scholar]
- Rosta, S. A Kiskunsági Homokhátság 13-16. Századi Településtörténete. Ph.D. Thesis, Eötvös Lóránd University, Budapest, Hungary, 2014; p. 378. (In Hungarian). [Google Scholar]
- Vajda, T. Az Izsáki Kolon-tó és Környezete Tájökológiai Vizsgálata. Ph.D. Thesis, University of Szeged, Szeged, Hungary, 1999; p. 61. (In Hungarian). [Google Scholar]
- Gácsér, I. Az 1211. évi tihanyi összeírás helyesírasa és hangtani sajátságai. Pannonhalmi Füzetek 1941, 29, 1–40. (In Hungarian) [Google Scholar]
- Györffy, G. Diplomata Hungariae Antiquissima I. Kötet (DHA I); In aedibus Academiae Scientiarium Hungaricae: Budapest, Hungary, 1992; pp. 149–152. (In Latin) [Google Scholar]
- Györffy, G. Árpád Kori Oklevelek; Balassa Kiadó: Budapest, Hungary, 1997; p. 148. (In Hungarian) [Google Scholar]
- Szabó, I. A Falu Rendszer Kialakulása Magyarországon (X–XV. Század); Akadémiai Kiadó: Budapest, Hungary, 1966; p. 215. (In Hungarian) [Google Scholar]
- Sümegi, P. Little Cuman region in the Medieval Age—Through the eyes of a geologist. In The Leader and His People of the Cuman; Horváth, F., Ed.; Archaeolingua Kiadó: Budapest, Hungary, 2001; pp. 313–317. (In Hungarian) [Google Scholar]
- Iványosi-Szabó, A. Az Izsáki Kolon-tó Üledék-és Környezetföldtani Vizsgálata. Ph.D. Thesis, University of Szeged, Szeged, Hungary, 1984. (In Hungarian). [Google Scholar]
- Treitz, P. A Kurjantói Nádasrét és a Kolon-tó Környékének Agrogeológiai Leírása—Manuscript; MÁFI: Budapest, Hungary, 1931; p. 90. (In Hungarian) [Google Scholar]
- Kolon-tó [Lake Kolon]. Available online: https://www.kolon-to.com (accessed on 8 September 2023).
- Kolon-tó|Kiskunsági Nemzeti Park [Lake Kolon|Little Cumanian National Park]. Available online: https://www.knp.hu/hu/kolon-to (accessed on 10 October 2023).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PC 1 | PC 2 | |
|---|---|---|
| Eigenvalue | 6.0113 | 3.4310 |
| % variance | 40.088 | 22.881 |
| Abies | −0.2733 | 0.0942 |
| Acer | 0.1205 | 0.2520 |
| Alnus | 0.2277 | 0.4002 |
| Betula | −0.0633 | 0.3313 |
| Carpinus | 0.1744 | 0.4233 |
| Corylus | 0.3261 | −0.2836 |
| Fagus | 0.1696 | 0.3973 |
| Fraxinus sp. | 0.2773 | −0.1043 |
| Juniperus | −0.1947 | 0.0139 |
| Picea | 0.0186 | 0.0483 |
| Pinus | −0.3720 | −0.1443 |
| Quercus | 0.3711 | 0.1374 |
| Salix | 0.3519 | −0.0812 |
| Tilia | 0.2643 | −0.3555 |
| Ulmus | 0.3226 | −0.2321 |
| PC 1 | PC 2 | |
|---|---|---|
| Eigenvalue | 12.76 | 3.59 |
| % variance | 51.06 | 14.38 |
| Achillea type | 0.2089 | −0.1710 |
| Ambrosia type | 0.1699 | 0.2278 |
| Artemisia | −0.0419 | −0.3495 |
| Aster type | 0.1231 | −0.1575 |
| Cannabis/Humulus | 0.2320 | 0.1678 |
| Caryophyllaceae | 0.2271 | 0.1437 |
| Centaurea sp. | 0.1490 | −0.2085 |
| Cerealia | 0.2555 | 0.1110 |
| Triticum | 0.2355 | 0.1993 |
| Secale | 0.2458 | −0.0721 |
| Chenopodiaceae | 0.2205 | −0.1427 |
| Cirsium type | 0.2183 | 0.2341 |
| Compositae-Liguliflorae | 0.1338 | −0.3280 |
| Compositae-Tubuliflorae | 0.1024 | −0.3500 |
| Cruciferae | 0.2169 | −0.0742 |
| Galium | 0.2181 | −0.0110 |
| Helianthemum | −0.1050 | 0.3371 |
| Plantago lanceolata | 0.2523 | 0.0610 |
| Plantago major/media | 0.2129 | −0.1808 |
| Poaceae | −0.1066 | 0.3246 |
| Ranunculaceae | 0.2312 | 0.1795 |
| Sambucus sp. | 0.2370 | 0.1341 |
| Thalictrum | 0.1823 | −0.0504 |
| Umbelliferae | 0.2397 | −0.0075 |
| Urtica | 0.2257 | 0.0577 |
| PC 1 | PC 2 | PC 3 | PC 4 | PC 5 | PC 6 | |
|---|---|---|---|---|---|---|
| Eigenvalue | 3.35 | 2.28 | 1.98 | 1.61 | 1.37 | 1.16 |
| % variance | 17.62 | 11.98 | 10.41 | 8.49 | 7.21 | 6.10 |
| Botryococcus braunii | −0.3385 | 0.3299 | 0.0204 | −0.0554 | 0.1167 | −0.1086 |
| Butomus umbellatus | 0.0568 | 0.4004 | −0.0667 | −0.0821 | −0.0946 | 0.1896 |
| Coelastrum | 0.0022 | 0.0855 | 0.0095 | −0.0856 | −0.0174 | −0.3136 |
| Cyperaceae | 0.4008 | −0.2580 | −0.0825 | −0.0429 | −0.1886 | 0.1110 |
| Equisetum | 0.0857 | −0.0721 | 0.4189 | −0.0167 | −0.2920 | −0.4112 |
| Filicales | 0.1583 | 0.5371 | −0.0119 | −0.0723 | 0.0313 | −0.0125 |
| Hottonia palustris | 0.2303 | −0.0320 | 0.4383 | 0.0064 | 0.3959 | 0.1319 |
| Menyanthes | −0.0687 | 0.0630 | 0.1676 | 0.2246 | −0.3474 | 0.4921 |
| Myriophyllum sp. | −0.0899 | 0.0793 | 0.0221 | 0.2637 | 0.0292 | −0.0854 |
| Myriophyllum spicatum | −0.1495 | 0.0501 | 0.0432 | 0.0364 | −0.0460 | −0.1264 |
| Myriophyllum verticillatum | 0.1439 | 0.0621 | −0.1565 | 0.5593 | 0.1601 | −0.2752 |
| Nuphar | −0.0209 | 0.0268 | 0.4602 | 0.2460 | −0.3678 | 0.1182 |
| Nymphaea | 0.3969 | −0.1953 | −0.2842 | −0.0632 | −0.0815 | 0.2467 |
| Pediastrum | −0.3092 | −0.2888 | −0.1121 | 0.2265 | 0.2038 | 0.2222 |
| Pediastrum kawrayskii | −0.3523 | −0.0565 | 0.1062 | 0.2938 | 0.0333 | 0.2260 |
| Sparganium-Typha angustifolia | 0.2106 | 0.4644 | −0.1097 | 0.1617 | −0.0842 | 0.2263 |
| Trilete spora | 0.0951 | 0.0589 | 0.2406 | −0.1015 | 0.5771 | 0.2410 |
| Typha latifolia | 0.3333 | −0.0244 | 0.3603 | 0.1678 | 0.1296 | −0.0764 |
| Utricularia | 0.1951 | 0.0221 | −0.2224 | 0.5240 | 0.0884 | −0.1530 |
| Cyperaceae | −0.3385 | 0.3299 | 0.0204 | −0.0554 | 0.1167 | −0.1086 |
| PC 1 | PC 2 | PC 3 | PC 4 | |
|---|---|---|---|---|
| Eigenvalue | 10.32 | 4.54 | 2.73 | 1.99 |
| % variance | 41.29 | 18.18 | 10.92 | 7.95 |
| Abies | −0.2150 | 0.0568 | 0.2529 | 0.2539 |
| Alnus | 0.2198 | 0.2950 | 0.0613 | 0.1118 |
| Betula | −0.0470 | 0.1485 | 0.4077 | 0.3142 |
| Corylus | 0.2058 | −0.3335 | −0.0123 | 0.0222 |
| Fagus | 0.1651 | 0.2618 | 0.1947 | 0.1256 |
| Picea | 0.0460 | 0.0362 | −0.2121 | 0.5245 |
| Pinus | −0.2889 | −0.0258 | −0.1521 | −0.0003 |
| Quercus | 0.2739 | −0.0431 | 0.2043 | 0.0551 |
| Salix | 0.2723 | −0.0972 | −0.1089 | −0.0520 |
| Tilia | 0.1465 | −0.3488 | −0.0663 | −0.0483 |
| Ulmus | 0.2010 | −0.2954 | 0.0754 | −0.0364 |
| Artemisia | 0.1628 | −0.2642 | 0.0553 | 0.2613 |
| Aster type | 0.1232 | 0.2204 | −0.3030 | 0.3050 |
| Cerealia | 0.1190 | 0.2831 | −0.0745 | −0.3086 |
| Chenopodiaceae | 0.2220 | 0.2036 | 0.0518 | −0.2436 |
| Compositae-Liguliflorae | 0.2676 | −0.0195 | −0.1192 | 0.0503 |
| Compositae-Tubuliflorae | 0.2494 | −0.0803 | −0.0651 | 0.0926 |
| Poaceae | −0.2004 | −0.0443 | 0.1577 | 0.2055 |
| Botryococcus braunii | −0.2228 | −0.0182 | −0.2514 | 0.0435 |
| Cyperaceae | 0.2585 | −0.0341 | 0.1117 | 0.0723 |
| Filicales | 0.0237 | 0.1222 | −0.4145 | 0.3287 |
| Nymphaea | 0.2518 | −0.1494 | 0.1264 | 0.1187 |
| Pediastrum | −0.1511 | −0.0735 | 0.4149 | 0.0963 |
| Typha latifolia | 0.1370 | 0.3172 | 0.0265 | −0.1182 |
| PC 1 | PC 2 | PC 3 | PC 4 | |
|---|---|---|---|---|
| Eigenvalue | 11.27 | 3.76 | 2.85 | 1.80 |
| % variance | 45.07 | 15.04 | 11.40 | 7.19 |
| Valvata cristata | 0.2637 | 0.1090 | 0.0004 | 0.0942 |
| Valvata macrostoma | −0.2570 | 0.2227 | −0.0187 | −0.0286 |
| Bithynia leachi | −0.2645 | 0.0000 | −0.1781 | 0.0981 |
| Bithynia tentaculata | 0.2267 | −0.1484 | 0.1148 | −0.3121 |
| Lymnaea stagnalis | −0.0880 | −0.1154 | −0.2196 | −0.2632 |
| Lymnaea palustris | 0.2047 | −0.0117 | 0.1843 | −0.1646 |
| Physa fontinalis | 0.0146 | −0.4134 | 0.2226 | 0.0996 |
| Planorbarius corneus | −0.1635 | −0.0463 | 0.3246 | 0.3577 |
| Planorbis planorbis | −0.0435 | −0.1371 | 0.5203 | −0.0759 |
| Anisus spirorbis | −0.1820 | 0.2349 | 0.1175 | 0.0591 |
| Anisus leucostoma | −0.1514 | −0.0279 | 0.4301 | 0.0658 |
| Bathyomphalus contortus | −0.2566 | −0.0034 | −0.2330 | 0.0706 |
| Armiger crista | 0.2072 | −0.0215 | 0.1635 | −0.1660 |
| Pisidium | 0.1998 | −0.1764 | −0.1924 | 0.3066 |
| Carychium minimum | 0.2678 | 0.1181 | −0.0576 | 0.0385 |
| Succinea putris | 0.0946 | −0.0756 | 0.0437 | −0.5170 |
| Succinella oblonga | 0.2485 | 0.1943 | −0.0773 | 0.0524 |
| Oyloma elegans | 0.2678 | 0.0287 | 0.0157 | 0.0737 |
| Vertigo antivertigo | 0.1520 | −0.1583 | 0.1426 | 0.3394 |
| Granaria frumentum | 0.2264 | 0.2046 | −0.1143 | 0.2054 |
| Vallonia pulchella | 0.1893 | 0.1286 | 0.0996 | 0.2520 |
| Vallonia enninesis | −0.2008 | 0.3166 | 0.1503 | −0.0485 |
| Limacidae | 0.1657 | 0.3752 | 0.1228 | −0.0565 |
| Trochulus hispidus | −0.2008 | 0.3166 | 0.1503 | −0.0485 |
| PC 1 | PC 2 | PC 3 | PC 4 | |
|---|---|---|---|---|
| Eigenvalue | 5.70 | 3.95 | 3.31 | 1.81 |
| % variance | 22.80 | 15.81 | 13.23 | 7.25 |
| Charcoal (Pinus) | 0.2088 | 0.1965 | −0.1129 | 0.0689 |
| Wood | 0.2187 | 0.0149 | 0.1520 | −0.5093 |
| UOM | 0.2937 | 0.2972 | −0.0263 | 0.0746 |
| ULF | 0.2400 | −0.2548 | 0.3072 | 0.1020 |
| Monocot. Undiff. | −0.1400 | 0.1821 | 0.2916 | 0.0457 |
| Ostracoda | 0.1272 | 0.3075 | −0.0157 | −0.0152 |
| Mollusca | −0.2118 | 0.1761 | 0.1797 | −0.0028 |
| Chara vulgaris | −0.2259 | 0.2130 | 0.2905 | 0.0196 |
| Nitellopsis obtusa | −0.1834 | 0.1484 | 0.2532 | 0.0265 |
| Porifera | 0.1480 | 0.2320 | −0.0498 | 0.3744 |
| Phragmites australis R | −0.1298 | 0.2468 | 0.3393 | 0.0354 |
| Carex rostrata R | −0.1473 | 0.0727 | 0.1441 | 0.0181 |
| Equisetum fluitans R | 0.1340 | −0.1424 | 0.2256 | −0.1507 |
| Carex acutiformis R | −0.1302 | 0.0767 | 0.2034 | 0.0203 |
| Carex elata R | 0.3294 | 0.1493 | 0.1771 | −0.0153 |
| Juncus sp. R | −0.0785 | 0.1351 | 0.3515 | −0.0047 |
| Drepanocladus aduncus | 0.2254 | −0.2034 | 0.2562 | 0.1576 |
| Trichoptera sp. | 0.1962 | 0.2562 | 0.0485 | −0.1436 |
| Typha sp. R | 0.2877 | −0.1972 | 0.2749 | 0.0988 |
| Schenoplectus sp. R | 0.2234 | 0.2911 | −0.0867 | 0.0922 |
| Salix E | 0.1420 | 0.1350 | 0.0046 | −0.4928 |
| Moinia sp. | 0.1824 | 0.2532 | −0.1248 | 0.3377 |
| Nymphaea alba SE | 0.1512 | 0.1478 | −0.0770 | −0.3172 |
| Typha angustifolia S | 0.1794 | −0.1084 | 0.0805 | 0.0305 |
| Salix R | 0.2428 | −0.2202 | 0.1962 | 0.1710 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vári, T.Z.; Pál-Molnár, E.; Sümegi, P. Reconstructing the Paleoenvironmental Evolution of Lake Kolon (Hungary) through Palaeoecological, Statistical and Historical Analyses. Diversity 2023, 15, 1095. https://doi.org/10.3390/d15101095
Vári TZ, Pál-Molnár E, Sümegi P. Reconstructing the Paleoenvironmental Evolution of Lake Kolon (Hungary) through Palaeoecological, Statistical and Historical Analyses. Diversity. 2023; 15(10):1095. https://doi.org/10.3390/d15101095
Chicago/Turabian StyleVári, Tamás Zsolt, Elemér Pál-Molnár, and Pál Sümegi. 2023. "Reconstructing the Paleoenvironmental Evolution of Lake Kolon (Hungary) through Palaeoecological, Statistical and Historical Analyses" Diversity 15, no. 10: 1095. https://doi.org/10.3390/d15101095
APA StyleVári, T. Z., Pál-Molnár, E., & Sümegi, P. (2023). Reconstructing the Paleoenvironmental Evolution of Lake Kolon (Hungary) through Palaeoecological, Statistical and Historical Analyses. Diversity, 15(10), 1095. https://doi.org/10.3390/d15101095