
Aggregate Size Mediated the Changes in Soil Microbial Communities After the Afforestation of a Former Dryland in Northwestern China

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Sampling
2.3. Determination of Distribution, Stability, and Chemical Properties of Soil Aggregates
2.4. DNA Extraction, PCR Amplification, and High-Throughput Sequencing
2.5. Data Analysis
3. Results
3.1. Distribution, Stability, and Chemical Properties of Soil Aggregates
3.2. Composition of Microbial Communities Within Soil Aggregates
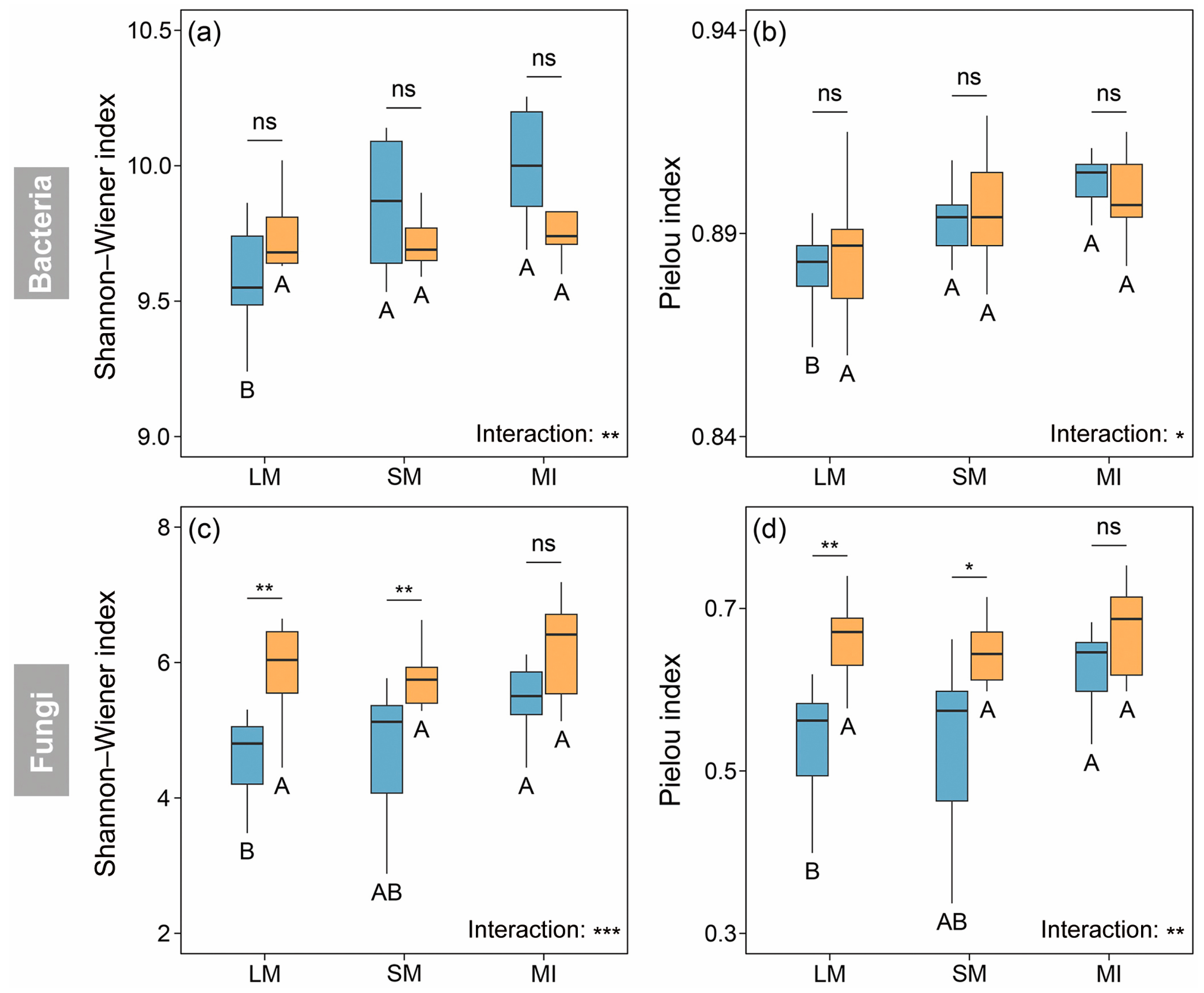
3.3. Diversity of Microbial Communities Within Soil Aggregates
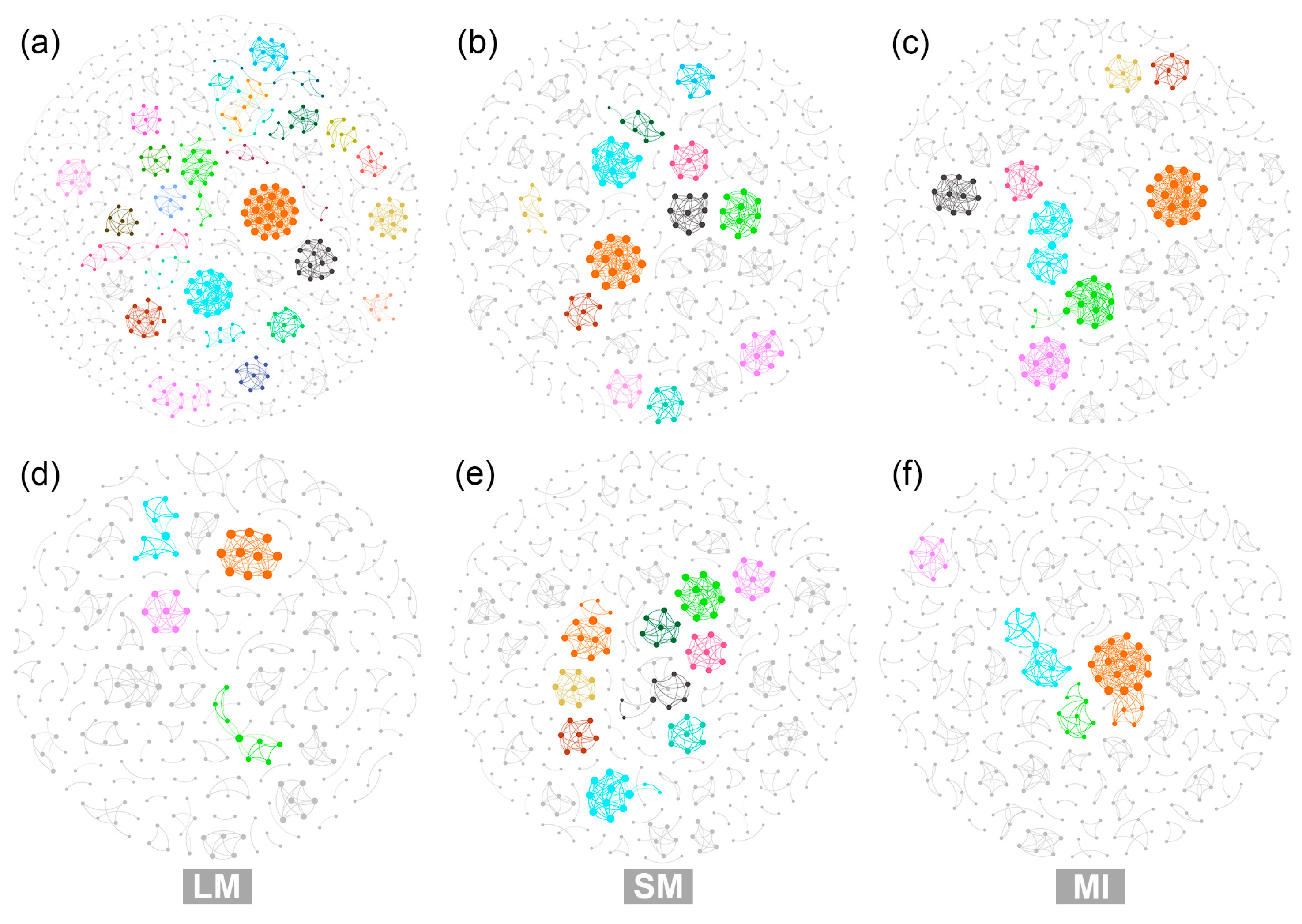
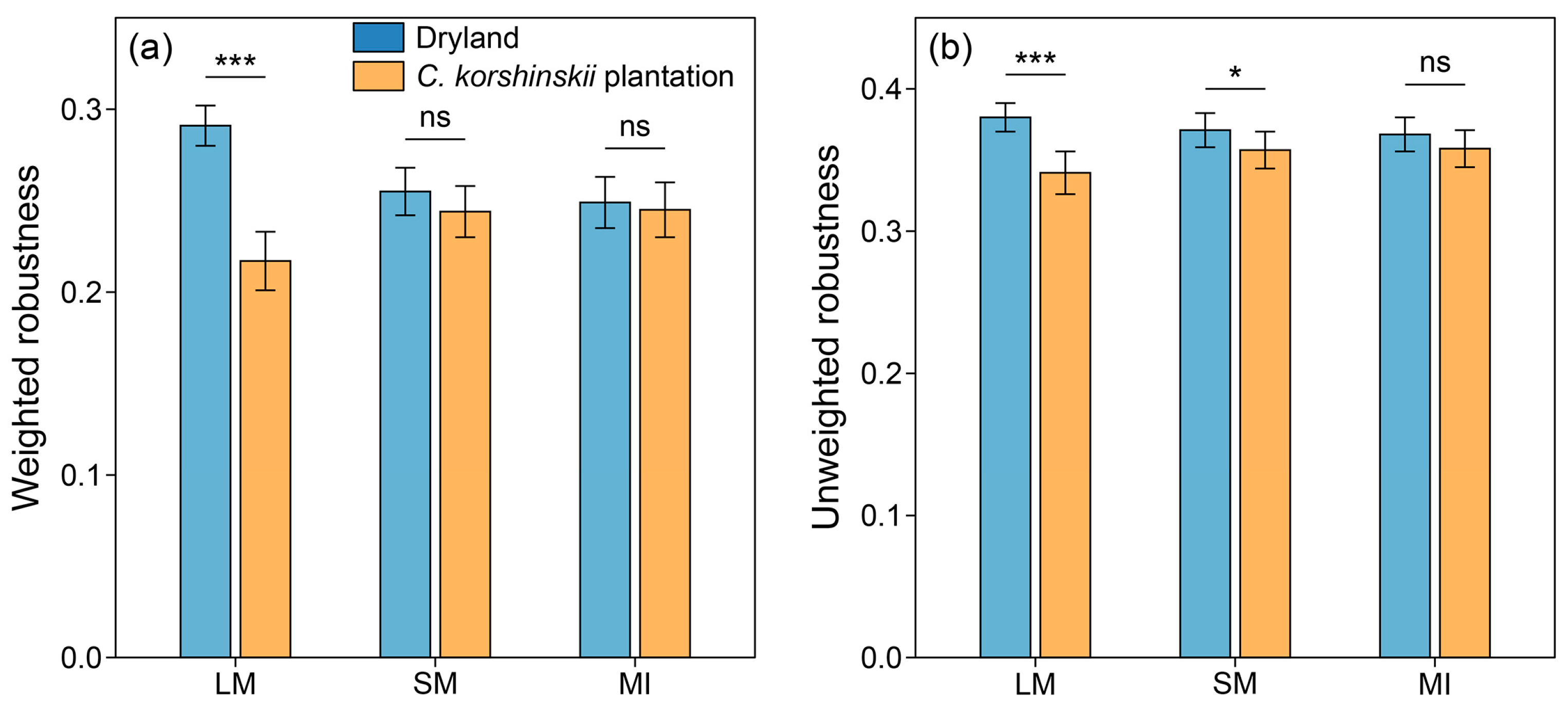
3.4. Characteristics of Microbial Co-Occurrence Networks Within Soil Aggregates
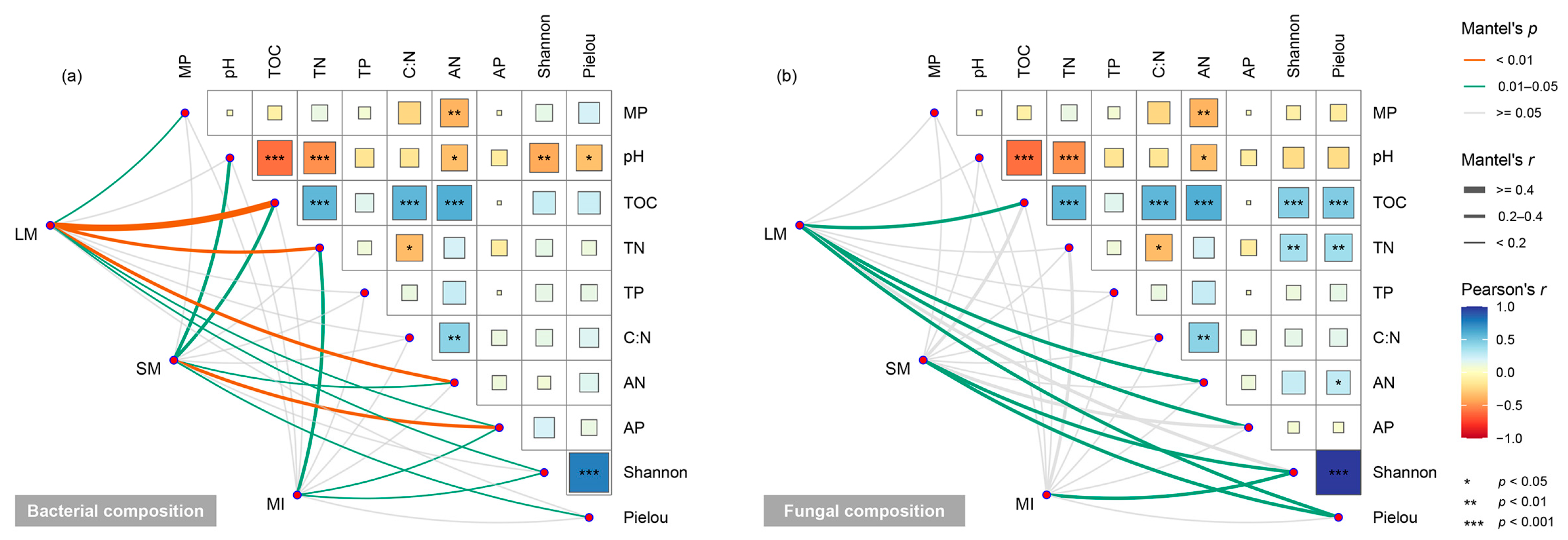
3.5. Factors Influencing Microbial Communities Within Soil Aggregates
4. Discussion
4.1. Impacts of Afforestation on Distribution, Stability, and Chemical Properties of Soil Aggregates
4.2. Impacts of Afforestation on Composition and Diversity of Microbial Communities Within Soil Aggregates
4.3. Impacts of Afforestation on Complexity and Stability of Microbial Co-Occurrence Networks Within Soil Aggregates
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Global Forest Resources Assessment 2020: Main Report; FAO: Rome, Italy, 2020. [Google Scholar]
- La Manna, L.; Tarabini, M.; Gomez, F.; Rostagno, C.M. Changes in soil organic matter associated with afforestation affect erosion processes: The case of erodible volcanic soils from Patagonia. Geoderma 2021, 403, 115265. [Google Scholar] [CrossRef]
- Nadal-Romero, E.; Llena, M.; Cortijos-López, M.; Lasanta, T. Afforestation after land abandonment as a nature-based solution in Mediterranean mid-mountain areas: Implications and research gaps. Curr. Opin. Environ. Sci. Health 2023, 34, 100481. [Google Scholar] [CrossRef]
- Guo, Y.; Abdalla, M.; Espenberg, M.; Hastings, A.; Hallett, P.; Smith, P. A systematic analysis and review of the impacts of afforestation on soil quality indicators as modified by climate zone, forest type and age. Sci. Total Environ. 2021, 757, 143824. [Google Scholar] [CrossRef]
- Rong, G.; Zhang, X.; Wu, H.; Ge, N.; Yao, Y.; Wei, X. Changes in soil organic carbon and nitrogen mineralization and their temperature sensitivity in response to afforestation across China’s Loess Plateau. Catena 2021, 202, 105226. [Google Scholar] [CrossRef]
- Kumar, N.; Khamzina, A.; Knöfel, P.; Lamers, J.P.A.; Tischbein, B. Afforestation of degraded croplands as a water-saving option in irrigated region of the aral sea basin. Water 2021, 13, 1433. [Google Scholar] [CrossRef]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The role of soil microorganisms in plant mineral nutrition—Current knowledge and future directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef]
- Basu, S.; Kumar, G.; Chhabra, S.; Prasad, R. Role of soil microbes in biogeochemical cycle for enhancing soil fertility. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2021; pp. 149–157. [Google Scholar]
- Liu, J.; Le, T.H.; Zhu, H.; Yao, Y.; Zhu, H.; Cao, Y.; Zhao, Z. Afforestation of cropland fundamentally alters the soil fungal community. Plant Soil 2020, 457, 279–292. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Fan, X.D.; Ren, C.J.; Zhang, L.; Han, X.H.; Yang, G.H.; Wang, J.; Doughty, R. Changes of the organic carbon content and stability of soil aggregates affected by soil bacterial community after afforestation. Catena 2018, 171, 622–631. [Google Scholar] [CrossRef]
- Kong, W.; Wei, X.; Wu, Y.; Shao, M.; Zhang, Q.; Sadowsky, M.J.; Ishii, S.; Reich, P.B.; Wei, G.; Jiao, S.; et al. Afforestation can lower microbial diversity and functionality in deep soil layers in a semiarid region. Glob. Chang. Biol. 2022, 28, 6086–6101. [Google Scholar] [CrossRef] [PubMed]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Matchado, M.S.; Lauber, M.; Reitmeier, S.; Kacprowski, T.; Baumbach, J.; Haller, D.; List, M. Network analysis methods for studying microbial communities: A mini review. Comput. Struct. Biotechnol. J. 2021, 19, 2687–2698. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.M.; Guo, X.; Wu, L.; Zhang, Y.; Xiao, N.; Ning, D.; Shi, Z.; Zhou, X.; Wu, L.; Yang, Y.; et al. Climate warming enhances microbial network complexity and stability. Nat. Clim. Chang. 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Yang, Y.; Chai, Y.; Xie, H.; Zhang, L.; Zhang, Z.; Yang, X.; Hao, S.; Gai, J.; Chen, Y. Responses of soil microbial diversity, network complexity and multifunctionality to three land-use changes. Sci. Total Environ. 2023, 859, 160255. [Google Scholar] [CrossRef]
- Yu, P.; Tang, H.; Sun, X.; Shi, W.; Pan, J.; Liu, S.; Jia, H.; Ding, Z.; Tang, X.; Chen, M. Afforestation alters soil microbial community composition and reduces microbial network complexity in a karst region of Southwest China. Land Degrad. Dev. 2024, 35, 2926–2939. [Google Scholar] [CrossRef]
- Wang, M.; Masoudi, A.; Wang, C.; Yang, J.; Zhai, Y.; Wu, C.; Yu, Z.; Liu, J. Plantation type and afforestation age disclose variable influences on soil microbial compositions in man-made forests in the Xiong’an New Area, China. Land Degrad. Dev. 2022, 33, 3058–3073. [Google Scholar] [CrossRef]
- Upton, R.N.; Bach, E.M.; Hofmockel, K.S. Spatio-temporal microbial community dynamics within soil aggregates. Soil Biol. Biochem. 2019, 132, 58–68. [Google Scholar] [CrossRef]
- Bach, E.M.; Williams, R.J.; Hargreaves, S.; Yang, F.; Hofmockel, K.S. Greatest soil microbial diversity found in micro-habitats. Soil Biol. Biochem. 2018, 118, 217–226. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Lal, R. Mechanisms of carbon sequestration in soil aggregates. Crit. Rev. Plant Sci. 2004, 23, 481–504. [Google Scholar] [CrossRef]
- Sey, B.K.; Manceur, A.M.; Whalen, J.K.; Gregorichm, E.G.; Rochette, P. Small-scale heterogeneity in carbon dioxide, nitrous oxide and methane production from aggregates of a cultivated sandy-loam soil. Soil Biol. Biochem. 2008, 40, 2468–2473. [Google Scholar] [CrossRef]
- Six, J.; Elliott, E.T.; Paustian, K. Soil macroaggregate turnover and microaggregate formation: A mechanism for C sequestration under no-tillage agriculture. Soil Biol. Biochem. 2000, 32, 2099–2103. [Google Scholar] [CrossRef]
- Yu, P.; Liu, J.; Tang, H.; Ci, E.; Tang, X.; Liu, S.; Ding, Z.; Ma, M. The increased soil aggregate stability and aggregate-associated carbon by farmland use change in a karst region of Southwest China. Catena 2023, 231, 107284. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Q.; Wang, H.; Wang, W.; Zhong, Z.; Di, G. Effects of poplar shelterbelt plantations on soil aggregate distribution and organic carbon in northeastern China. Forests 2022, 13, 1546. [Google Scholar] [CrossRef]
- Stewart, B.A.; Thapa, S. Dryland farming: Concept, origin and brief history. In Innovations in Dryland Agriculture; Farooq, M., Siddique, K.H.M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 3–29. [Google Scholar]
- Zhuo, Z.; Chen, Q.; Zhang, X.; Chen, S.; Gou, Y.; Sun, Z.; Huang, Y.; Shi, Z. Soil organic carbon storage, distribution, and influencing factors at different depths in the dryland farming regions of Northeast and North China. Catena 2022, 210, 105934. [Google Scholar] [CrossRef]
- Li, P.; Chen, J.; Zhao, G.; Holden, J.; Liu, B.; Chan, F.K.S.; Hu, J.; Wu, P.; Mu, X. Determining the drivers and rates of soil erosion on the Loess Plateau since 1901. Sci. Total Environ. 2022, 823, 153674. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, P.; Liu, X.; Xiao, L.; Shi, P.; Zhao, B. Effects of farmland conversion on the stoichiometry of carbon, nitrogen, and phosphorus in soil aggregates on the Loess Plateau of China. Geoderma 2019, 351, 188–196. [Google Scholar] [CrossRef]
- Rong, G.; Li, W.; Zhu, H.; Zhou, J.; Qiu, L.; Ge, N.; Wei, X.; Shao, M. Dynamics of new- and old-organic carbon and nitrogen in bulk soils and aggregates following afforestation on farmland. Catena 2020, 195, 104838. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, G.B.; Zhang, J.Y.; Xue, S. Long-term effects of vegetational restoration on soil microbial communities on the Loess Plateau of China. Restor. Ecol. 2016, 24, 794–804. [Google Scholar] [CrossRef]
- Adingo, S.; Yu, J.-R.; Liu, X.; Jing, S.; Li, X.; Zhang, X. Land-use change influence soil quality parameters at an ecologically fragile area of YongDeng County of Gansu Province, China. PeerJ 2021, 9, e12246. [Google Scholar] [CrossRef]
- Ju, W.; Fang, L.; Shen, G.; Delgado-Baquerizo, M.; Chen, J.; Zhou, G.; Ma, D.; Bing, H.; Liu, L.; Liu, J.; et al. New perspectives on microbiome and nutrient sequestration in soil aggregates during long-term grazing exclusion. Glob. Chang. Biol. 2024, 30, e17027. [Google Scholar] [CrossRef]
- Yan, Z.; Zhou, J.; Yang, L.; Gunina, A.; Yang, Y.; Peixoto, L.; Zeng, Z.; Zang, H.; Kuzyakov, Y. Diversified cropping systems benefit soil carbon and nitrogen stocks by increasing aggregate stability: Results of three fractionation methods. Sci. Total Environ. 2022, 824, 153878. [Google Scholar] [CrossRef]
- Liu, X.; Ma, J.; Ma, Z.W.; Li, L.H. Soil nutrient contents and stoichiometry as affected by land-use in an agro-pastoral region of northwest China. Catena 2017, 150, 146–153. [Google Scholar] [CrossRef]
- Li, X.W.; Li, X.L.; Shi, Y.; Zhao, S.J.; Liu, J.L.; Lin, Y.Y.; Li, C.L.; Zhang, C.H. Effects of microtopography on soil microbial communities in alpine meadows on the Qinghai-Tibetan Plateau. Catena 2024, 239, 107945. [Google Scholar] [CrossRef]
- Amézketa, E. Soil Aggregate Stability: A Review. J. Sustain. Agric. 1999, 14, 83–151. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, W.W.; Wang, G.; Yang, S.Q.; Pereira, P. Effects of long-term afforestation and natural grassland recovery on soil properties and quality in Loess Plateau (China). Sci. Total Environ. 2021, 770, 144833. [Google Scholar] [CrossRef]
- Zhong, Z.; Wu, S.; Lu, X.; Ren, Z.; Wu, Q.; Xu, M.; Ren, C.; Yang, G.; Han, X. Organic carbon, nitrogen accumulation, and soil aggregate dynamics as affected by vegetation restoration patterns in the Loess Plateau of China. Catena 2021, 196, 104867. [Google Scholar] [CrossRef]
- Guo, L.K.; Shen, J.; Li, B.; Li, Q.Q.; Wang, C.Q.; Guan, Y.; D’Acqui, L.P.; Luo, Y.L.; Tao, Q.; Xu, Q.; et al. Impacts of Agricultural Land Use Change on Soil Aggregate Stability and Physical Protection of Organic C. Sci. Total Environ. 2020, 707, 136049. [Google Scholar] [CrossRef]
- Han, X.; Zhao, F.; Tong, X.; Deng, J.; Yang, G.; Chen, L.; Kang, D. Understanding soil carbon sequestration following the afforestation of former arable land by physical fractionation. Catena 2017, 150, 317–327. [Google Scholar] [CrossRef]
- Creamer, C.A.; Foster, A.L.; Lawrence, C.; McFarland, J.; Schulz, M.; Waldrop, M.P. Mineralogy dictates the initial mechanism of microbial necromass association. Geochim. Cosmochim. Acta 2019, 260, 161–176. [Google Scholar] [CrossRef]
- Xuan, M.; Ai, L.; Wu, F.; Zhang, X.; Ni, X. Biomarkers evidence shows a preferential occlusion of microbial necromass in mineral-associated and not particle organic matter. Geoderma 2024, 450, 117030. [Google Scholar] [CrossRef]
- Liu, L.; Zhu, Q.; Wan, Y.; Yang, R.; Mou, J.; Li, Y.; Meng, L.; Zhu, T.; Elrys, A.S. Afforestation improves soil organic carbon and total nitrogen stocks mainly through increasing >2 mm aggregate fractions and stimulating carbon and nitrogen transformations within aggregates in subtropical karst region. Catena 2024, 243, 108220. [Google Scholar] [CrossRef]
- Deng, Q.; McMahon, D.E.; Xiang, Y.; Yu, C.-L.; Jackson, R.B.; Hui, D. A global meta-analysis of soil phosphorus dynamics after afforestation. New Phytol. 2017, 213, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Dragone, N.B.; Hoffert, M.; Strickland, M.S.; Fierer, N. The taxonomic and genomic attributes of oligotrophic soil bacteria. ISME Commun. 2024, 4, ycae081. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, A.C.H.; Hartmann, M.; Conz, R.F.; Six, J.; Solly, E.F. Prolonged water limitation shifts the soil microbiome from copiotrophic to oligotrophic lifestyles in Scots pine mesocosms. Environ. Microbiol. Rep. 2024, 16, e13211. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.C.; Wang, S.S.; Wang, J.X.; Qi, X.; Long, Q.X.; Huang, M.Z. The Shift of Soil Bacterial Community After Afforestation Influence Soil Organic Carbon and Aggregate Stability in Karst Region. Front. Microbiol. 2022, 13, 901126. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.; Di Lonardo, D.P.; Bodelier, P.L. Revisiting life strategy concepts in environmental microbial ecology. FEMS Microbiol. Ecol. 2017, 93, fix006. [Google Scholar] [CrossRef]
- He, B.; Li, Q.; Zou, S.; Bai, X.; Li, W.; Chen, Y. Dynamic changes of soil microbial communities during the afforestation of Pinus armandii in a karst region of Southwest China. Microb. Ecol. 2024, 87, 36. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, X.; Shan, Y.; Lu, X.; Cao, J. Effects of land-use patterns on soil microbial diversity and composition in the Loess Plateau, China. J. Arid. Land 2024, 16, 415–430. [Google Scholar] [CrossRef]
- Zeng, Q.; Liu, Y.; Xiao, L.; An, S. Climate and soil properties regulate soil fungal communities on the Loess Plateau. Pedobiologia 2020, 81, 150668. [Google Scholar] [CrossRef]
- Chen, W.; Yu, T.; Zhao, C.; Li, B.; Qin, Y.; Li, H.; Zhang, X. Development and determinants of topsoil bacterial and fungal communities of afforestation by aerial sowing in Tengger Desert, China. J. Fungi 2023, 9, 399. [Google Scholar] [CrossRef]
- Liu, C.G.; Jin, Y.Q.; Lin, F.M.; Jiang, C.; Zeng, X.L.; Feng, D.F.; Huang, F.Z.; Tang, J.W. Land use change alters carbon and nitrogen dynamics mediated by fungal functional guilds within soil aggregates. Sci. Total Environ. 2023, 902, 166080. [Google Scholar] [CrossRef]
- Trivedi, P.; Rochester, I.J.; Trivedi, C.; Van Nostrand, J.D.; Zhou, J.; Karunaratne, S.; Anderson, I.C.; Singh, B.K. Soil aggregate size mediates the impacts of cropping regimes on soil carbon and microbial communities. Soil Biol. Biochem. 2015, 91, 169–181. [Google Scholar] [CrossRef]
- Huang, K.; Guo, Z.; Zhao, W.; Song, C.; Wang, H.; Li, J.; Mumin, R.; Sun, Y.; Cui, B. Response of fungal communities to afforestation and its indication for forest restoration. For. Ecosyst. 2023, 10, 100125. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, H.; Liang, Y.; Liang, Y.; Zhao, Y.; Wang, Z.; Li, Y.; Liu, W.; Wang, X.; Yang, G.; et al. The Multifunctionality of Soil Aggregates Is Related to the Complexity of Aggregate Microbial Community during Afforestation. Catena 2024, 236, 107737. [Google Scholar] [CrossRef]
- Chen, P.; Wang, L.; Li, J.-Y.; Wan, W.; Zhang, R.-Q.; Liu, Y.; Li, Z.-G. Response of Soil Aggregate-Associated Fertility and Microbial Communities to Afforestation in the Degraded Ecosystem of the Danjiangkou Reservoir, China. Plant Soil 2024, 501, 171–189. [Google Scholar] [CrossRef]
- Yang, P.; van Elsas, J.D. Mechanisms and ecological implications of the movement of bacteria in soil. Appl. Soil Ecol. 2018, 129, 112–120. [Google Scholar] [CrossRef]
- Li, X.; Zhang, H.; Sun, M.; Xu, N.; Sun, G.; Zhao, M. Land use change from upland to paddy field in Mollisols drives soil aggregation and associated microbial communities. Appl. Soil Ecol. 2020, 146, 103351. [Google Scholar] [CrossRef]
- Guseva, K.; Darcy, S.; Simon, E.; Alteio, L.V.; Montesinos-Navarro, A.; Kaiser, C. From diversity to complexity: Microbial networks in soils. Soil Biol. Biochem. 2022, 69, 108604. [Google Scholar] [CrossRef]
- De Gryze, S.; Six, J.; Merckx, R. Quantifying water-stable soil aggregate turnover and its implication for soil organic matter dynamics in a model study. Eur. J. Soil Sci. 2006, 57, 693–707. [Google Scholar]
- Feng, H.; Wang, S.; Gao, Z.; Pan, H.; Zhuge, Y.; Ren, X.; Hu, S.; Li, C. Aggregate Stability and Organic Carbon Stock under Different Land Uses Integrally Regulated by Binding Agents and Chemical Properties in Saline-sodic Soils. Land Degrad. Dev. 2021, 32, 4151–4161. [Google Scholar] [CrossRef]
- MacArthur, R. Fluctuations of animal populations and a measure of community stability. Ecology 1955, 36, 533–536. [Google Scholar] [CrossRef]
- May, R. Stability and Complexity in Model Ecosystems; Princeton University Press: Princeton, NJ, USA, 1973. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land-Use Type | MP (%) | MWD (mm) | ||
|---|---|---|---|---|
| LM | SM | MI | ||
| Dryland | 21.63 (2.69) Ab | 49.48 (2.27) Aa | 28.89 (2.45) Bb | 1.67 (0.12) A |
| C. korshinskii plantation | 16.51 (1.4) Ac | 47.46 (1.89) Aa | 36.03 (1.99) Ab | 1.40 (0.07) A |
| Properties | Dryland | C. korshinskii Plantation | ||||
|---|---|---|---|---|---|---|
| LM | SM | MI | LM | SM | MI | |
| pH | 8.53 (0.02) Aa | 8.53 (0.03) Aa | 8.26 (0.05) Ab | 8.48 (0.03) Aa | 8.42 (0.03) Bab | 8.30 (0.04) Ab |
| TOC (g·kg−1) | 7.88 (0.33) Bb | 8.05 (0.36) Bb | 11.13 (0.52) Aa | 11.30 (0.58) Aa | 10.83 (0.78) Aa | 11.86 (0.70) Aa |
| TN (g·kg−1) | 0.82 (0.06) Ba | 1.01 (0.10) Aa | 1.03 (0.06) Aa | 1.01 (0.04) Aa | 1.04 (0.06) Aa | 1.09 (0.03) Aa |
| TP (g·kg−1) | 0.87 (0.03) Aa | 0.91 (0.06) Aa | 0.94 (0.04) Aa | 0.89 (0.05) Aa | 0.88 (0.05) Aa | 0.93 (0.03) Aa |
| C:N | 11.59 (0.86) Aab | 9.82 (0.75) Bb | 12.73 (0.40) Aa | 13.13 (0.69) Aa | 12.21 (0.62) Aa | 12.61 (0.49) Aa |
| AN (mg·kg−1) | 46.79 (3.91) Ba | 19.04 (3.66) Bb | 53.70 (3.92) Aa | 63.75 (3.28) Aa | 47.62 (10.03) Aa | 64.91 (5.87) Aa |
| AP (mg·kg−1) | 1.89 (0.24) Aa | 2.48 (0.40) Aa | 4.98 (1.46) Aa | 1.61 (0.12) Ab | 1.07 (0.17) Bb | 2.86 (0.22) Aa |
| Features | Dryland | C. korshinskii Plantation | ||||
|---|---|---|---|---|---|---|
| LM | SM | MI | LM | SM | MI | |
| Node | 735 | 503 | 500 | 347 | 473 | 439 |
| Edge | 1680 | 817 | 882 | 401 | 674 | 626 |
| Average degree | 4.571 | 3.249 | 3.528 | 2.311 | 2.850 | 2.852 |
| Average path length | 1.220 | 1.031 | 1.095 | 1.076 | 1.083 | 1.152 |
| Modularity | 0.953 | 0.968 | 0.951 | 0.970 | 0.972 | 0.948 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Bai, L.; Wang, W.; Wang, Y.; Chen, T.; Yang, Q.; Chen, H.; Kang, S.; Zhu, Y.; Liu, X. Aggregate Size Mediated the Changes in Soil Microbial Communities After the Afforestation of a Former Dryland in Northwestern China. Diversity 2024, 16, 696. https://doi.org/10.3390/d16110696
Zhang D, Bai L, Wang W, Wang Y, Chen T, Yang Q, Chen H, Kang S, Zhu Y, Liu X. Aggregate Size Mediated the Changes in Soil Microbial Communities After the Afforestation of a Former Dryland in Northwestern China. Diversity. 2024; 16(11):696. https://doi.org/10.3390/d16110696
Chicago/Turabian StyleZhang, Deming, Ling Bai, Wei Wang, Yanhe Wang, Tiankun Chen, Quan Yang, Haowen Chen, Shuning Kang, Yongan Zhu, and Xiang Liu. 2024. "Aggregate Size Mediated the Changes in Soil Microbial Communities After the Afforestation of a Former Dryland in Northwestern China" Diversity 16, no. 11: 696. https://doi.org/10.3390/d16110696
APA StyleZhang, D., Bai, L., Wang, W., Wang, Y., Chen, T., Yang, Q., Chen, H., Kang, S., Zhu, Y., & Liu, X. (2024). Aggregate Size Mediated the Changes in Soil Microbial Communities After the Afforestation of a Former Dryland in Northwestern China. Diversity, 16(11), 696. https://doi.org/10.3390/d16110696