
Allelopathic Potential of Nicotiana glauca Aqueous Extract on Seed Germination and Seedlings of Acacia gerrardii


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials Collection
2.2. Purification, Treatment, and Viability Test of A. gerrardii Seeds
2.3. Preparation of Aqueous Extracts and Germination Bioassay
- Germination percentage (GP): GP was measured using the formula below:
- Coefficient of velocity of germination (CVG) or germination speed (GS): CVG was computed according to the formula below [31].where N1, N2, N3, …, Nk represent the number of germinated seeds perceived at time (days) T1, T2, T3, ⋯⋯, Tk after sowing, while k represents the total number of time intervals.
- Mean germination time (MGT): MGT was measured according to the formula described in [32].where Ti is the number of days from the beginning of the test, Ni is the number of germinated seeds per day, and S is the total number of germinated seeds.
- Germination rate index [33]: GRI was measured using the following formula:where G1 represents the percentage of germination in the first day after seed sowing, and G2 represents the percentage of germination in the second day after seed sowing.
- First day of germination (FDG) and last day of germination (LDG) were recorded.
- Time spread of germination (TSG): TSG was calculated by different of time intervals between the first and last day of germination [34].
- Radicle length (RL), shoot length (SL), and total length (TL) of hypocotyl were measured using a ruler.
- Vigor Index (VI): = VI was measured using the formula described by [35].
- Fresh weights of young hypocotyls were determined using electrical balance.
2.4. Estimation of Total Phenols and Tannins of N. glauca Aqueous Extract
2.5. Statistical Analysis
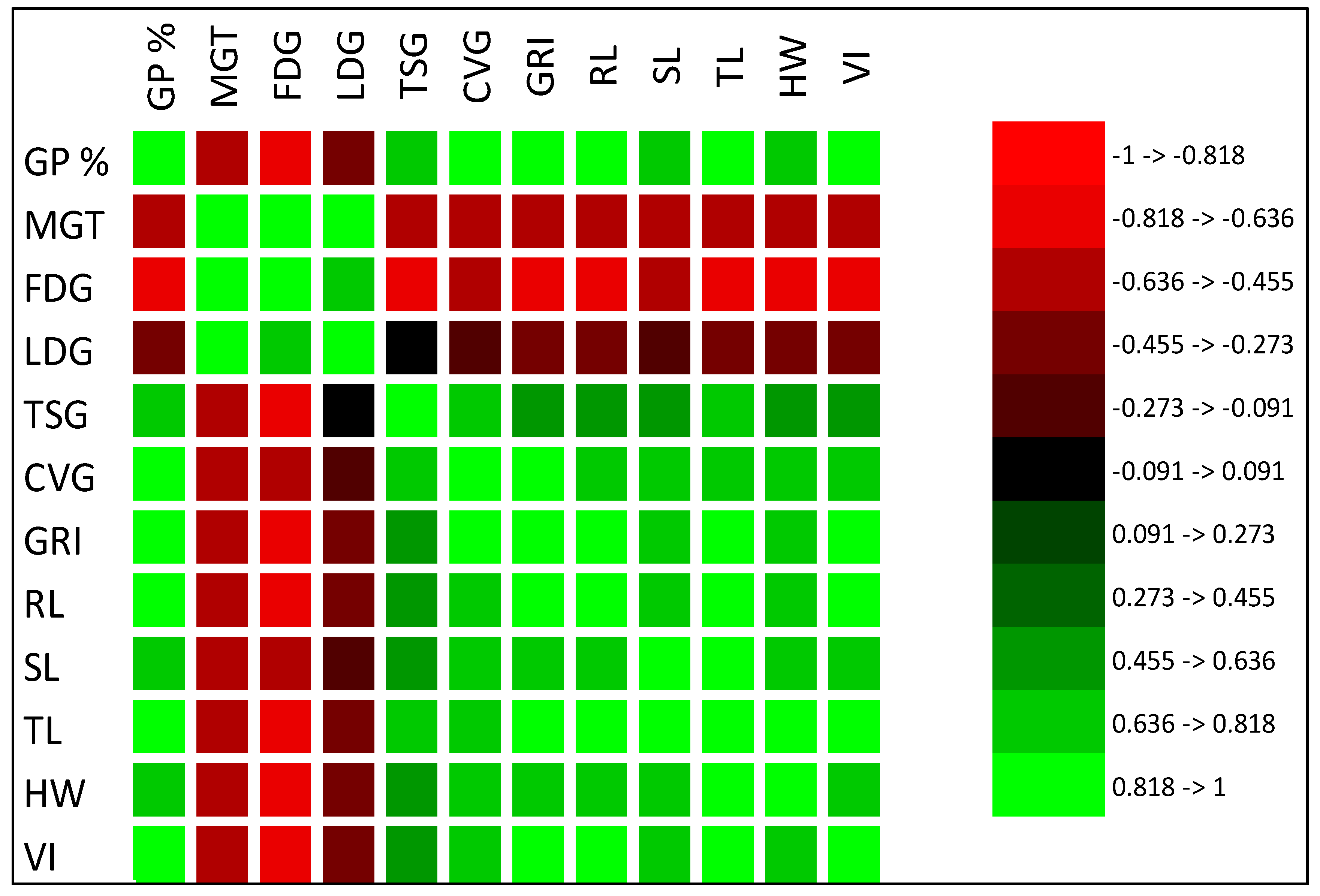
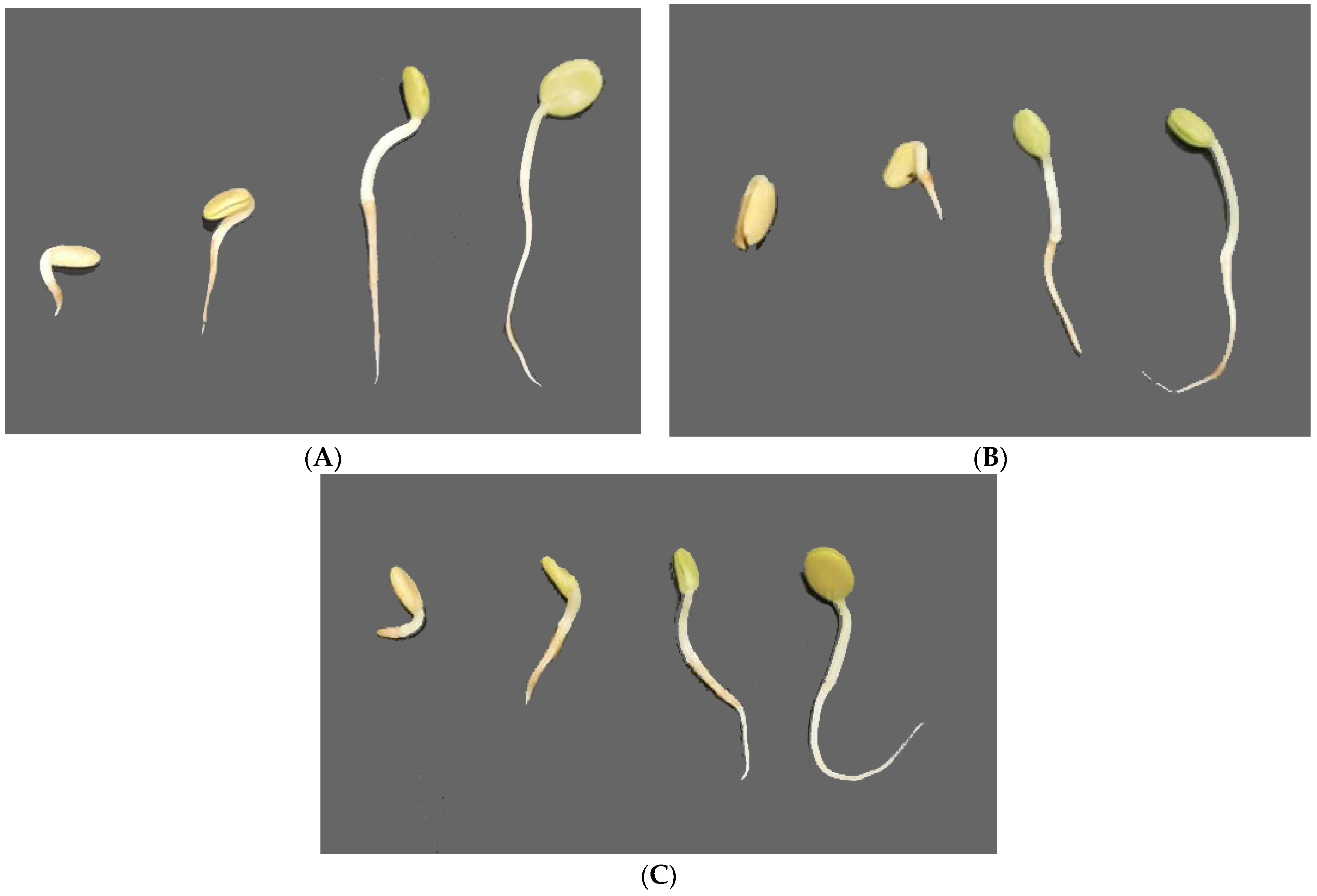
3. Results
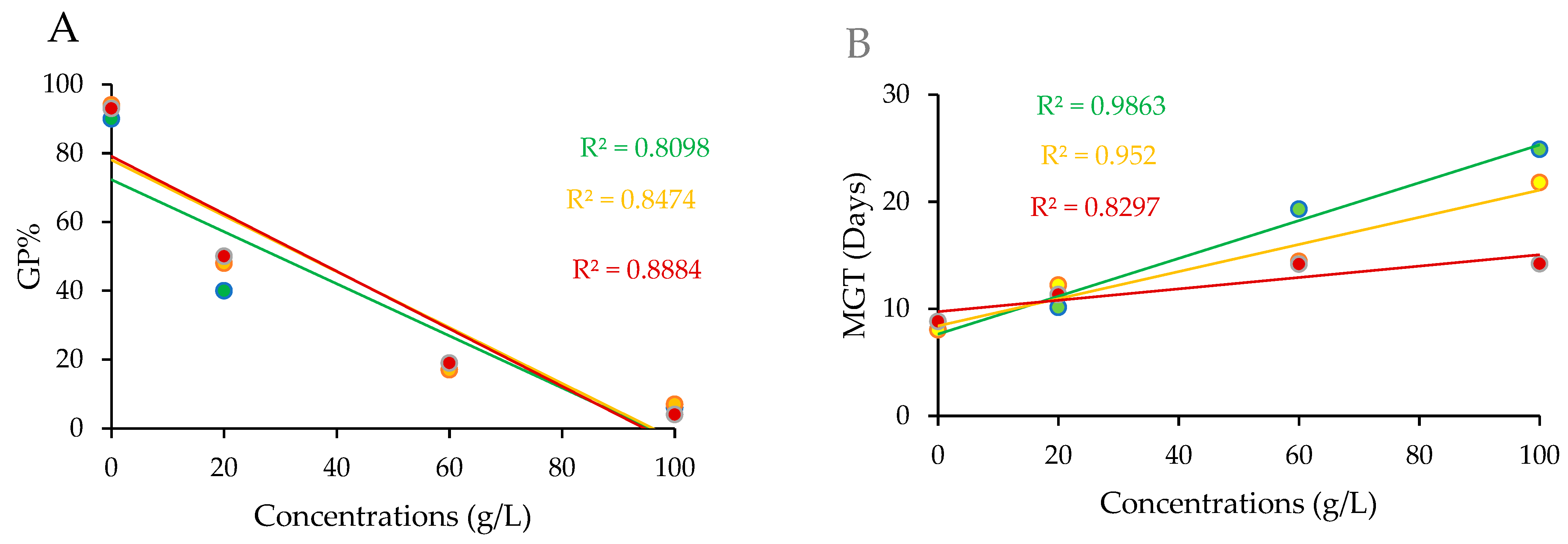
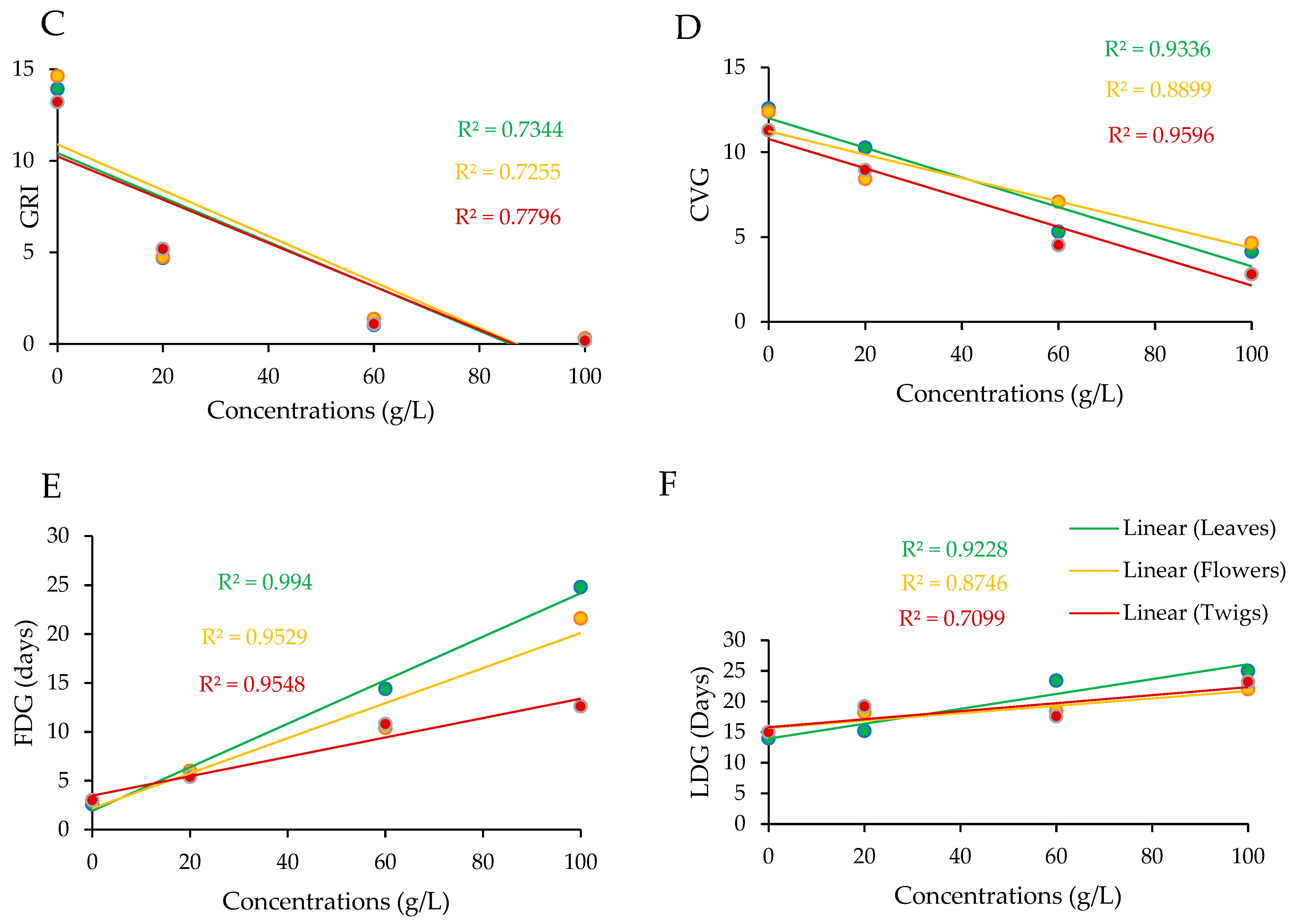
3.1. Effects of Aqueous Extracts on Germination Indices of A. gerrardii
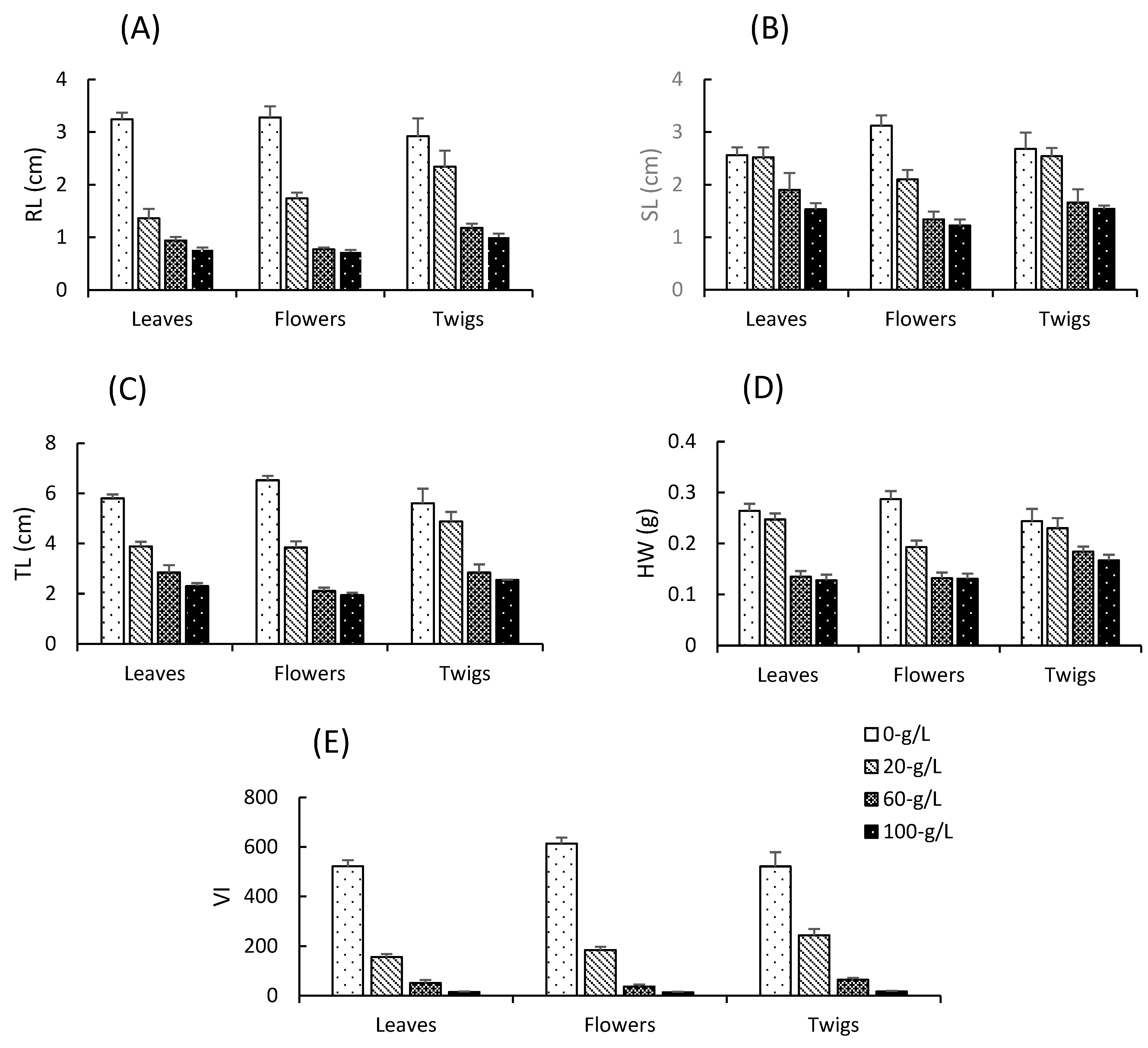
3.2. Effects of Aqueous Extracts on A. gerrardii Hypocotyl Indices
3.3. Total Phenols and Tannins in Aqueous Extract Sources of Invasive N. glauca
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linders, T.E.W.; Schaffner, U.; Eschen, R.; Abebe, A.; Choge, S.K.; Nigatu, L.; Mbaabu, P.R.; Shiferaw, H.; Allan, E. Direct and indirect effects of invasive species: Biodiversity loss is a major mechanism by which an invasive tree affects ecosystem functioning. J. Ecol. 2019, 107, 2660–2672. [Google Scholar] [CrossRef]
- Alharthi, A.S.; Abd-ElGawad, A.M.; Assaeed, A.M. Influence of the invasive shrub Nicotiana glauca Graham on the plant seed bank in various locations in Taif region, western of Saudi Arabia. Saudi J. Biol. Sci. 2021, 28, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-J.; Chen, D.; Yan, R.; Yu, F.-H.; van Kleunen, M. Invasive alien clonal plants are competitively superior over co-occurring native clonal plants. Perspect. Plant Ecol. Evol. Syst. 2019, 40, 125484. [Google Scholar] [CrossRef]
- Alqarni, A.S.; Awad, A.M.; Owayss, A.A. Evaluation of Acacia gerrardii Benth.(Fabaceae: Mimosoideae) as a honey plant under extremely hot-dry conditions: Flowering phenology, nectar yield and honey potentiality. J. Anim. Plant Sci. 2015, 25, 1667–1674. [Google Scholar]
- Furer, V.; Hersch, M.; Silvetzki, N.; Breuer, G.S.; Zevin, S. Nicotiana glauca (tree tobacco) intoxication—Two cases in one family. J. Med. Toxicol. 2011, 7, 47–51. [Google Scholar] [CrossRef]
- Issaly, E.A.; Sersic, A.N.; Pauw, A.; Cocucci, A.A.; Traveset, A.; Benitez-Vieyra, S.M.; Paiaro, V. Reproductive ecology of the bird-pollinated Nicotiana glauca across native and introduced ranges with contrasting pollination environments. Biol. Invasions 2020, 22, 485–498. [Google Scholar] [CrossRef]
- Al-Robai, S.A.; Mohamed, H.A.; Ahmed, A.A.; Al-Khulaidi, A.W.A. Effects of elevation gradients and soil components on the vegetation density and species diversity of Alabna escarpment, southwestern Saudi Arabia. Acta Ecol. Sin. 2019, 39, 202–211. [Google Scholar] [CrossRef]
- Thomas, J.; El-Sheikh, M.A.; Alfarhan, A.H.; Alatar, A.A.; Sivadasan, M.; Basahi, M.; Al-Obaid, S.; Rajakrishnan, R. Impact of alien invasive species on habitats and species richness in Saudi Arabia. J. Arid Environ. 2016, 127, 53–65. [Google Scholar] [CrossRef]
- Alshahrani, S.T. Effect of aqueous extract of the invasive species tobacco (Nicotiana glauca L.) on seedlings growth of Juniper (Juniperus procera L.). Emir. J. Food Agric. 2008, 20, 10–17. [Google Scholar] [CrossRef]
- Inderjit; Cahill, J.F. Linkages of plant–soil feedbacks and underlying invasion mechanisms. AoB Plants 2015, 7, plv022. [Google Scholar]
- Wang, S.; Wei, M.; Wu, B.; Cheng, H.; Wang, C. Combined nitrogen deposition and Cd stress antagonistically affect the allelopathy of invasive alien species Canada goldenrod on the cultivated crop lettuce. Sci. Hortic. 2020, 261, 108955. [Google Scholar] [CrossRef]
- Alqarawi, A.A.; Hashem, A.; Kumar, A.; Al-Arjani, A.-B.F.; Abd_Allah, E.F.; Dar, B.A.; Wirth, S.; Davranov, K.; Egamberdieva, D. Allelopathic effects of the aqueous extract of Rhazya stricta on growth and metabolism of Salsola villosa. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2018, 152, 1263–1273. [Google Scholar]
- Kalisz, S.; Kivlin, S.N.; Bialic-Murphy, L. Allelopathy is pervasive in invasive plants. Biol. Invasions 2021, 23, 367–371. [Google Scholar] [CrossRef]
- Asakawa, C. Nature of interference potential of hairy vetch (Vicia villosa Roth) to radish (Raphanus sativus L.): Does allelopathy play any role? Crop Prot. 2001, 20, 261–265. [Google Scholar]
- Cipollini, K.; Titus, K.; Wagner, C. Allelopathic effects of invasive species (Alliaria petiolata, Lonicera maackii, Ranunculus ficaria) in the Midwestern United States. Allelopath. J. 2012, 29, 63–76. [Google Scholar]
- Kong, C.-H.; Xuan, T.D.; Khanh, T.D.; Tran, H.-D.; Trung, N.T. Allelochemicals and signaling chemicals in plants. Molecules 2019, 24, 2737. [Google Scholar] [CrossRef]
- John, J.; Sarada, S. Role of phenolics in allelopathic interactions. Allelopath. J. 2012, 29, 215–230. [Google Scholar]
- Tomar, N.S.; Agarwal, R. Influence of treatment of Jatropha curcas L. leachates and potassium on growth and phytochemical constituents of wheat (Triticum aestivum L.). Am. J. Plant Sci. 2013, 4, 1134–1150. [Google Scholar] [CrossRef]
- Mushtaq, W.; Ain, Q.; Siddiqui, M.; Hakeem, K.R. Cytotoxic allelochemicals induce ultrastructural modifications in Cassia tora L. and mitotic changes in Allium cepa L.: A weed versus weed allelopathy approach. Protoplasma 2019, 256, 857–871. [Google Scholar] [CrossRef]
- Chou, C.-H. Allelopathic researches in the subtropical vegetation in Taiwan. Comp. Physiol. Ecol. 1980, 5, 222–234. [Google Scholar]
- Gniazdowska, A.; Bogatek, R. Allelopathic interactions between plants. Multi site action of allelochemicals. Acta Physiol. Plant. 2005, 27, 395–407. [Google Scholar] [CrossRef]
- Findura, P.; Hara, P.; Szparaga, A.; Kocira, S.; Czerwińska, E.; Bartoš, P.; Nowak, J.; Treder, K. Evaluation of the effects of allelopathic aqueous plant extracts, as potential preparations for seed dressing, on the modulation of cauliflower seed germination. Agriculture 2020, 10, 122. [Google Scholar] [CrossRef]
- Nilsson, M.-C.; Högberg, P.; Zackrisson, O.; Fengyou, W. Allelopathic effects by Empetrum hermaphroditum on development and nitrogen uptake by roots and mycorrhizae of Pinus silvestris. Can. J. Bot. 1993, 71, 620–628. [Google Scholar] [CrossRef]
- Fernandez, C.; Monnier, Y.; Ormeño, E.; Baldy, V.; Greff, S.; Pasqualini, V.; Mévy, J.-P.; Bousquet-Mélou, A. Variations in allelochemical composition of leachates of different organs and maturity stages of Pinus halepensis. J. Chem. Ecol. 2009, 35, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Alghamdi, A.A. Phytoconstituents screening and antimicrobial activity of the invasive species Nicotiana glauca collected from Al-Baha region of Saudi Arabia. Saudi J. Biol. Sci. 2021, 28, 1544–1547. [Google Scholar] [CrossRef] [PubMed]
- El-Kenany, E.T.; El-Darier, S.M.; Abdellatif, A.A.; Shaklol, S.M. Allelopathic potential of invasive species: Nicotiana glauca Graham on some ecological and physiological aspects of Medicago sativa L. and Triticum aestivum L. Rend. Lincei 2017, 28, 159–167. [Google Scholar] [CrossRef]
- Turnbull, J. Forest tree seed testing. In Report on FAO/DANIDA Training Course on Forest Seed Collection and Handling; FAO: Rome, Italy, 1975. [Google Scholar]
- Aref, I.M.; Atta, H.A.E.; Shahrani, T.A.; Mohamed, A.I. Effects of seed pretreatment and seed source on germination of five Acacia spp. Afr. J. Biotechnol. 2011, 10, 15901–15910. [Google Scholar] [CrossRef]
- Anjum, T.; Bajwa, R. Importance of germination indices in interpretation of allelochemical effects. Int. J. Agric. Biol. 2005, 7, 1560–8530. [Google Scholar]
- Wardle, D.; Ahmed, M.; Nicholson, K. Allelopathic influence of nodding thistle (Carduus nutans L.) seeds on germination and radicle growth of pasture plants. N. Z. J. Agric. Res. 1991, 34, 185–191. [Google Scholar] [CrossRef]
- Scott, S.J.; Jones, R.; Williams, W. Review of data analysis methods for seed germination 1. Crop Sci. 1984, 24, 1192–1199. [Google Scholar] [CrossRef]
- Kader, M. A comparison of seed germination calculation formulae and the associated interpretation of resulting data. J. Proceeding R. Soc. New South Wales 2005, 138, 65–75. [Google Scholar] [CrossRef]
- Kader, M.; Omari, M.; Hattar, B. Maximizing germination percentage and speed of four Australian indigenous tree species. Dirasat Agric Sci 1998, 25, 157–169. [Google Scholar]
- Dhindwal, A.; Lather, B.; Singh, J. Efficacy of seed treatments on germination. Seedling emergence and vigour of cotton (Gossypium hirsutum) genotypes. Seed Res. 1991, 19, 59. [Google Scholar]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Rodrigues, C.I.; Marta, L.; Maia, R.; Miranda, M.; Ribeirinho, M.; Máguas, C. Application of solid-phase extraction to brewed coffee caffeine and organic acid determination by UV/HPLC. J. Food Compos. Anal. 2007, 20, 440–448. [Google Scholar] [CrossRef]
- Voegele, A.; Graeber, K.; Oracz, K.; Tarkowská, D.; Jacquemoud, D.; Turečková, V.; Urbanová, T.; Strnad, M.; Leubner-Metzger, G. Embryo growth, testa permeability, and endosperm weakening are major targets for the environmentally regulated inhibition of Lepidium sativum seed germination by myrigalone A. J. Exp. Bot. 2012, 63, 5337–5350. [Google Scholar] [CrossRef]
- da Silva Junior, V.A.; Junior, A.S.N. Topical effect of thiamethoxam on adults of Rhynchophorus palmarum L.(Coleoptera: Curculionidae) in laboratory. BioAssay 2022, 17, ba17001. [Google Scholar] [CrossRef]
- Zaid, N.; Khaled, M.; Eman, K.; Nouha, F. Phytochemical screening and heavy metals contents of Nicotiana glauca plant. Ijppr Hum. 2015, 4, 82–91. [Google Scholar]
- Mushtaq, W.; Siddiqui, M.B. Allelopathy in Solanaceae plants. J. Plant Prot. Res. 2018, 58, 1. [Google Scholar]
- Bachheti, A.; Sharma, A.; Bachheti, R.; Husen, A.; Pandey, D. Plant allelochemicals and their various applications. In Co-Evolution of Secondary Metabolites. Reference Series in Phytochemistry; Springer: Cham, Switzerland, 2020; pp. 441–465. [Google Scholar]
- Khoshkharam, M.; Shahrajabian, M.; Sun, W.; Cheng, Q. Survey the allelopathic effects of tobacco (Nicotiana tabacum L.) on corn (Zea mays L.) growth and germination. Cercet. Agron. Mold. 2019, 4, 332–340. [Google Scholar] [CrossRef]
- Florentine, S.; Westbrooke, M. Invasion of the noxious weed Nicotiana glauca R. Graham after an episodic flooding event in the arid zone of Australia. J. Arid. Environ. 2005, 60, 531–545. [Google Scholar] [CrossRef][Green Version]
- Nekonam, M.S.; Razmjoo, J.; Kraimmojeni, H.; Sharifnabi, B.; Amini, H.; Bahrami, F. Assessment of some medicinal plants for their allelopathic potential against redroot pigweed (Amaranthus retroflexus). J. Plant Prot. Res. 2014, 54, 90–95. [Google Scholar] [CrossRef]
- Li, J.; Chen, L.; Chen, Q.; Miao, Y.; Peng, Z.; Huang, B.; Guo, L.; Liu, D.; Du, H. Allelopathic effect of Artemisia argyi on the germination and growth of various weeds. Sci. Rep. 2021, 11, 4303. [Google Scholar] [CrossRef] [PubMed]
- Bieser, J.M.; Al-Zayat, M.; Murtada, J.; Thomas, S.C. Biochar mitigation of allelopathic effects in three invasive plants: Evidence from seed germination trials. J. Soil Sci. 2022, 102, 213–224. [Google Scholar] [CrossRef]
- ALHaithloul, H.A.S.; Khan, M.I.; Musa, A.; Ghoneim, M.M.; ALrashidi, A.A.; Khan, I.; Azab, E.; Gobouri, A.A.; Sofy, M.R.; El-Sherbiny, M.J.P. Phytotoxic effects of Acacia saligna dry leachates on germination, seedling growth, photosynthetic performance, and gene expression of economically important crops. PeerJ 2022, 10, e13623. [Google Scholar] [CrossRef]
- Qin, F.; Liu, S.; Yu, S. Effects of allelopathy and competition for water and nutrients on survival and growth of tree species in Eucalyptus urophylla plantations. For. Ecol. Manag. 2018, 424, 387–395. [Google Scholar] [CrossRef]
- Ladhari, A.; Gaaliche, B.; Zarrelli, A.; Ghannem, M.; Ben Mimoun, M. Allelopathic potential and phenolic allelochemicals discrepancies in Ficus carica L. cultivars. S. Afr. J. Bot. 2020, 130, 30–44. [Google Scholar] [CrossRef]
- Šoln, K.; Dolenc Koce, J. Allelopathic root inhibition and its mechanisms. Allelopath. J. 2021, 52, 181–198. [Google Scholar] [CrossRef]
- Sodaeizadeh, H.; Rafieiolhossaini, M.; Havlík, J.; Van Damme, P. Allelopathic activity of different plant parts of Peganum harmala L. and identification of their growth inhibitors substances. Plant Growth Regul. 2009, 59, 227–236. [Google Scholar] [CrossRef]
- El-Masry, R.; Ahmed, S.; El-Rokiek, K.G.; Messiha, N.K.; Mohamed, S.A. Allelopathic activity of the leaf powder of Ficus nitida on the growth and yield of Vicia faba and associated weeds. Bull. Natl. Res. Cent. 2019, 43, 67. [Google Scholar] [CrossRef]
- Argal, S.; Bhat, W.M.; Ahanger, M.A.; Agarwal, R. A Note on Phyto Analysis of P. juliflora (Swartz) DC. J. Funct. Environ. Bot. 2016, 6, 58–65. [Google Scholar] [CrossRef]
- Cai, S.; Mu, X. Allelopathic potential of aqueous leaf extracts of Datura stramonium L. on seed germination, seedling growth and root anatomy of Glycine max (L.) Merrill. Allelopath. J. 2012, 30, 235–245. [Google Scholar]
- Yan, Z.; Li, P.; Xiao, Y.; Cao, L.; Yao, L. Phytotoxic effects of allelochemical acacetin on seed germination and seedling growth of selected vegetables and its potential physiological mechanism. Agronomy 2022, 12, 1038. [Google Scholar] [CrossRef]
- Ercoli, L.; Masoni, A.; Pampana, S.; Arduini, I. Allelopathic effects of rye, brown mustard and hairy vetch on redroot pigweed, common lambsquarter and knotweed. Allelopath. J. 2007, 19, 249. [Google Scholar]
- Gniazdowska, A.; Krasuska, U.; Andrzejczak, O.; Soltys, D. Allelopathic compounds as oxidative stress agents: YES or NO. In Reactive Oxygen and Nitrogen Species Signaling and Communication in Plants; Springer: Cham, Switzerland, 2015; pp. 155–176. [Google Scholar]
- Gatti, A.B.; Ferreira, A.G.; Arduin, M.; Perez, S.C.G.d.A. Allelopathic effects of aqueous extracts of Artistolochia esperanzae O. Kuntze on development of Sesamum indicum L. seedlings. Acta Bot. Bras. 2010, 24, 454–461. [Google Scholar] [CrossRef]
- Kasiotis, K.M.; Evergetis, E.; Papachristos, D.; Vangelatou, O.; Antonatos, S.; Milonas, P.; Haroutounian, S.A.; Machera, K. An essay on ecosystem availability of Nicotiana glauca graham alkaloids: The honeybees case study. BMC Ecol. 2020, 20, 57. [Google Scholar] [CrossRef]
- Al-Wakeel, S.A.; Gabr, M.A.; Hamid, A.A.; Abu-El-Soud, W.M. Allelopathic effects of Acacia nilotica leaf residue on Pisum sativum L. Allelopath. J. 2007, 19, 411. [Google Scholar]
- Rinez, A.; Ladhari, A.; Omezzine, F.; Rinez, I.; Haouala, R. Phytotoxicity of Nicotiana glauca Graham aqueous extracts, a Tunisian invasive plant. In Proceedings of the 3rd International Symposium on Weeds and Invasive Plants, Ascona, Switzerland, 2–7 October 2011; pp. 2–7. [Google Scholar]
- Hassan, H.; El-Hameed, T.; Nasr, E. Ecological and phytochemical studies on Nicotiana glauca from Egypt. Egypt. J. Exp. Biol. 2014, 10, 87–95. [Google Scholar]







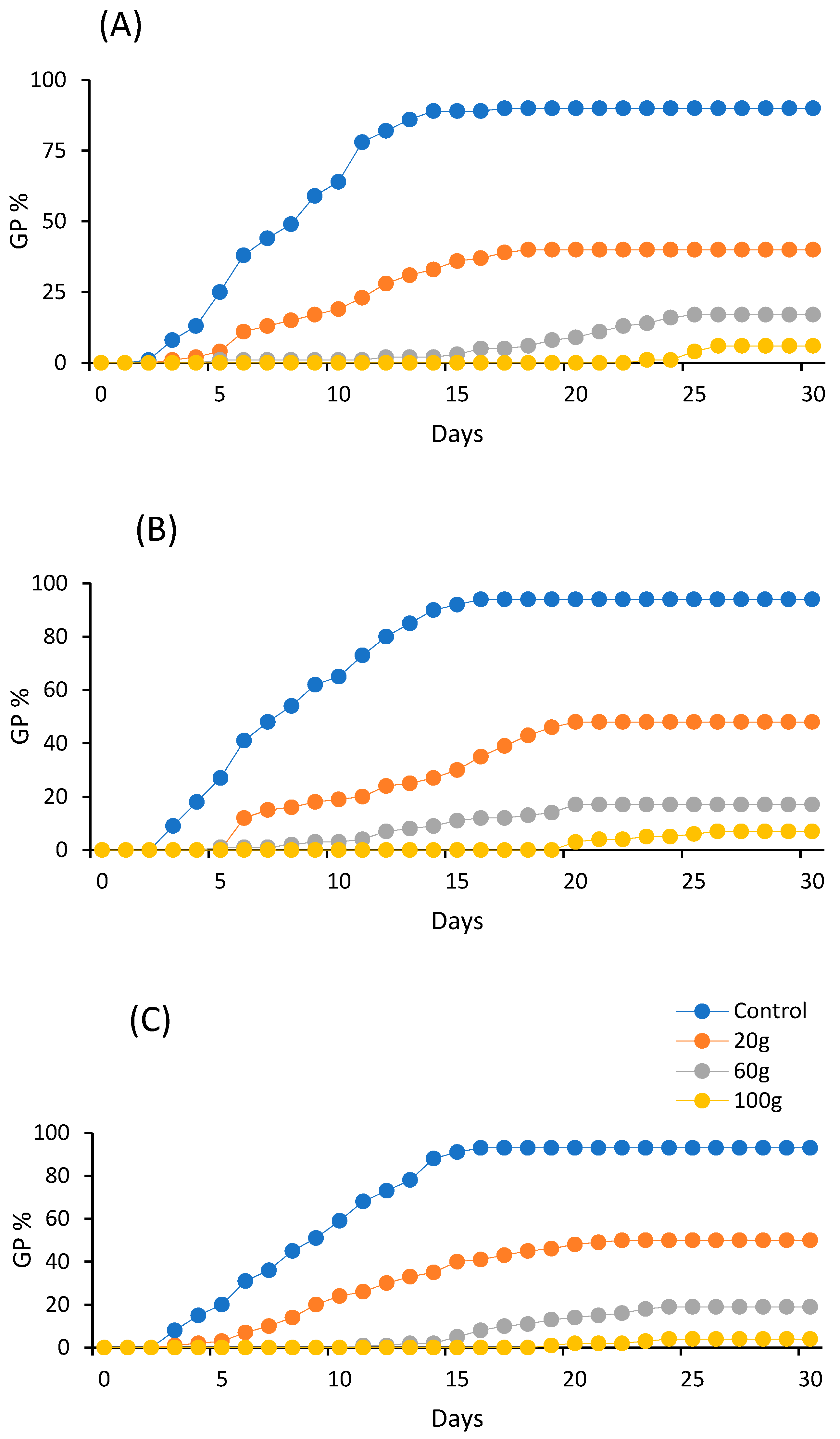{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Part | Concentration (g/L) | Germination Parameters | ||
|---|---|---|---|---|
| MGT (Day) | CVG | GRI (Day) | ||
| Leaves | 0 | 8.06 ± 0.25 d | 12.58 ± 0.25 a | 13.92 ± 0.09 ab |
| 20 | 10.13 ± 1.06 cd | 10.27 ± 1.02 bc | 4.69 ± 0.60 c | |
| 60 | 19.30 ± 1.43 ab | 5.31 ± 0.38 ef | 1.03 ± 0.23 d | |
| 100 | 24.90 ± 0.50 a | 4.14 ± 0.05 fg | 0.23 ± 0.03 d | |
| Flowers | 0 | 8.04 ± 0.19 d | 12.39 ± 0.32 a | 14.62 ± 0.48 a |
| 20 | 12.22 ± 1.13 cd | 8.43 ± 0.75 cd | 4.74 ± 0.45 c | |
| 60 | 14.43 ± 1.10 bc | 7.08 ± 0.55 de | 1.37 ± 0.31 d | |
| 100 | 21.80 ± 1.07 a | 4.65 ± 0.18 fg | 0.31 ± 0.05 d | |
| Twigs | 0 | 8.82 ± 0.17 cd | 11.29 ± 0.22 ab | 13.21 ± 0.47 b |
| 20 | 11.33 ± 1.04 cd | 8.96 ± 0.92 cd | 5.19 ± 0.61 c | |
| 60 | 14.18 ± 3.60 bcd | 4.53 ± 1.15 fg | 1.10 ± 0.28 d | |
| 100 | 14.20 ± 5.45 bcd | 2.81 ± 1.16 g | 0.18 ± 0.08 d | |
| Plant Part | Concentration (g/L) | Germination Parameters | ||
|---|---|---|---|---|
| FDG | LDG | TSG | ||
| Leaves | 0 | 2.60 ± 0.24 d | 14.00 ± 0.94 c | 11.40 ± 0.97 abc |
| 20 | 6.00 ± 1.14 cd | 15.20 ± 0.86 c | 9.20 ± 1.49 bcd | |
| 60 | 14.40 ± 2.82 b | 23.40 ± 0.67 ab | 9.00 ± 2.46 bcd | |
| 100 | 24.80 ± 0.48 a | 25.00 ± 0.54 a | 0.20 ± 0.20 e | |
| Flowers | 0 | 3.00 ± 0.00 d | 15.00 ± 0.44 c | 12.00 ± 0.44 ab |
| 20 | 6.00 ± 0.00 cd | 18.20 ± 1.11 bc | 12.20 ± 1.11 ab | |
| 60 | 10.40 ± 1.74 bc | 18.40 ± 0.92 bc | 8.00 ± 2.25 cd | |
| 100 | 21.60 ± 1.02 a | 22.00 ± 1.14 ab | 0.40 ± 0.24 e | |
| Twigs | 0 | 3.00 ± 0.00 d | 15.00 ± 0.44 c | 12.00 ± 0.44 ab |
| 20 | 5.40 ± 0.81 cd | 19.20 ± 1.39 abc | 13.80 ± 1.01 a | |
| 60 | 10.80 ± 2.80 bc | 17.60 ± 4.45 bc | 6.80 ± 1.93 d | |
| 100 | 12.60 ± 5.21 b | 13.20 ± 5.45 c | 0.60 ± 0.60 e | |
| Plant Organ | Compounds | |
|---|---|---|
| Total Phenols | Tannins | |
| Leaves | 4.30 ± 0.394 a | 2.61 ± 0.021 b |
| Flowers | 3.63 ± 0.037 a | 4.17 ± 0.019 a |
| Twigs | 0.61 ± 0.027 b | 0.75 ± 0.013 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelmalik, A.M.; Alshahrani, T.S.; Alqarawi, A.A.; Ahmed, E.M. Allelopathic Potential of Nicotiana glauca Aqueous Extract on Seed Germination and Seedlings of Acacia gerrardii. Diversity 2024, 16, 26. https://doi.org/10.3390/d16010026
Abdelmalik AM, Alshahrani TS, Alqarawi AA, Ahmed EM. Allelopathic Potential of Nicotiana glauca Aqueous Extract on Seed Germination and Seedlings of Acacia gerrardii. Diversity. 2024; 16(1):26. https://doi.org/10.3390/d16010026
Chicago/Turabian StyleAbdelmalik, Abdelmalik M., Thobayet S. Alshahrani, Abdulaziz A. Alqarawi, and Eljaily M. Ahmed. 2024. "Allelopathic Potential of Nicotiana glauca Aqueous Extract on Seed Germination and Seedlings of Acacia gerrardii" Diversity 16, no. 1: 26. https://doi.org/10.3390/d16010026
APA StyleAbdelmalik, A. M., Alshahrani, T. S., Alqarawi, A. A., & Ahmed, E. M. (2024). Allelopathic Potential of Nicotiana glauca Aqueous Extract on Seed Germination and Seedlings of Acacia gerrardii. Diversity, 16(1), 26. https://doi.org/10.3390/d16010026