


Factors Influencing Activity and Detection of Species in a Cross Timbers Snake Assemblage



Abstract
1. Introduction
2. Materials and Methods
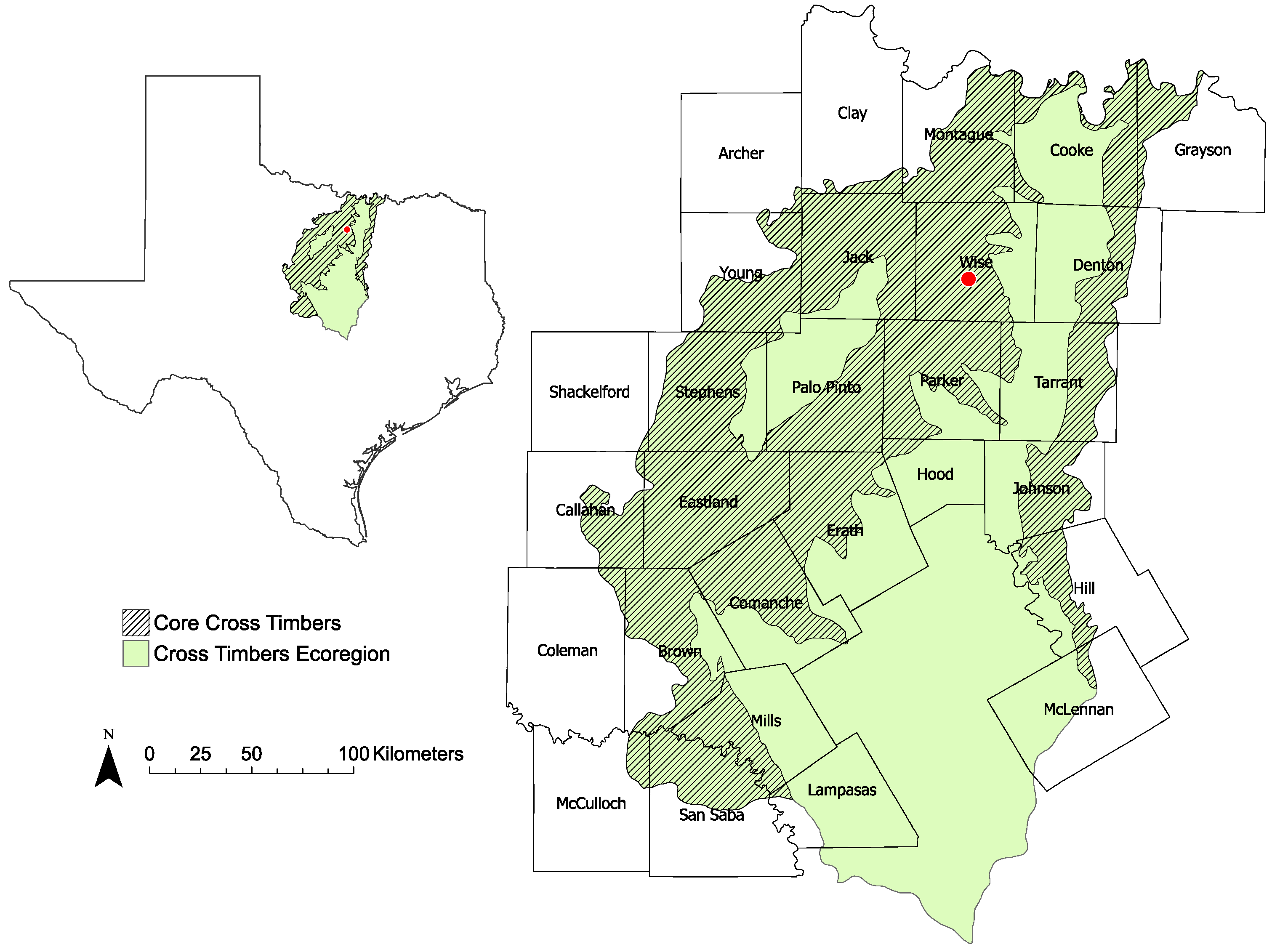
2.1. Study Area and Protocols
2.2. Statistical Analyses
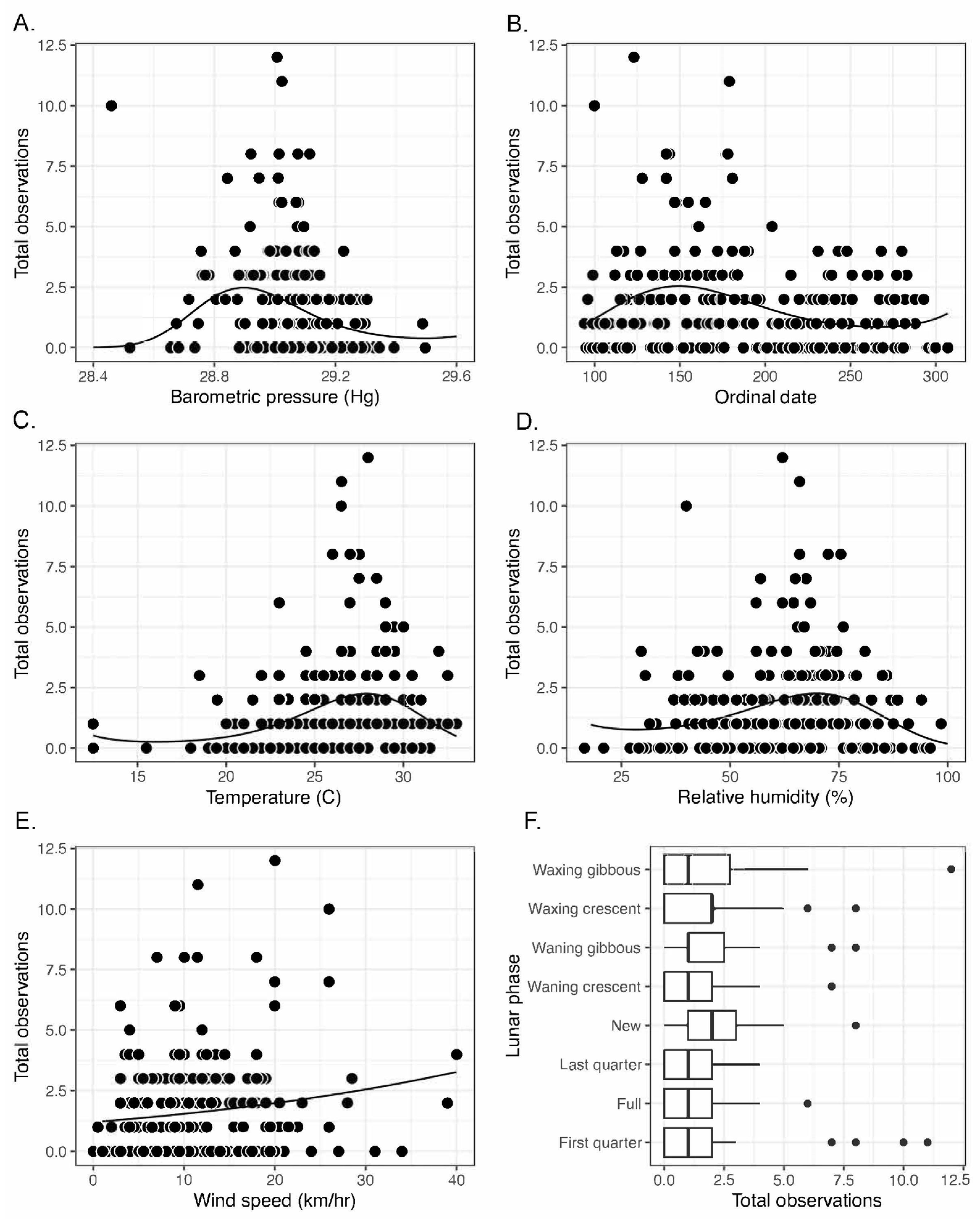
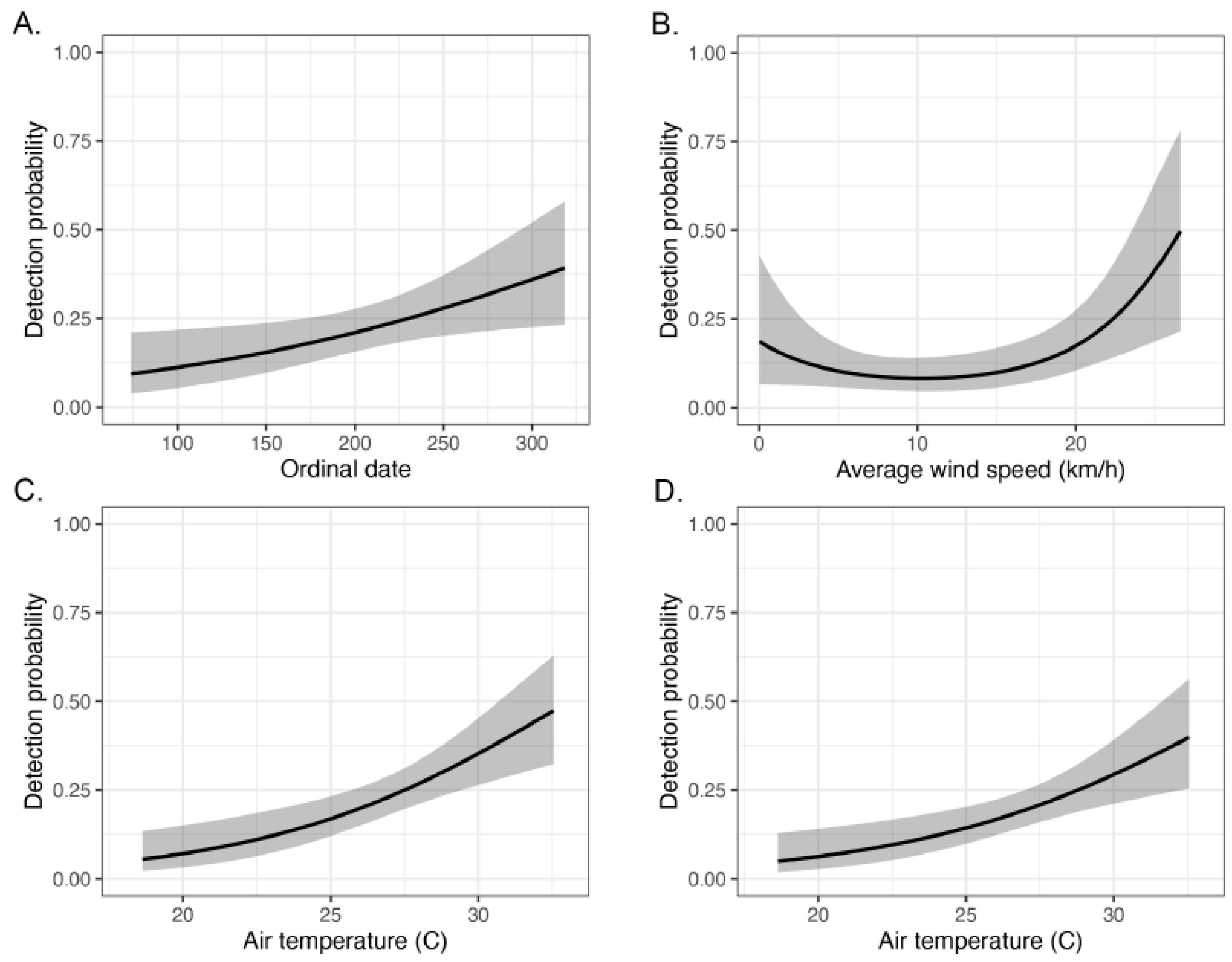
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Francaviglia, R.V. The Cast Iron Forest: A Natural and Cultural History of the North American Cross Timbers; University of Texas Press: Austin, TX, USA, 1998. [Google Scholar]
- Hoagland, B.W.; Butler, I.H.; Johnson, F.L.; Glenn, S. The Cross Timbers. In Savannas, Barrens, and Rock Outcrop Plant Communities of North America; Anderson, R.C., Fralish, J.S., Baskin, J.M., Eds.; Cambridge University Press: Cambridge, UK, 1999; pp. 231–245. [Google Scholar]
- Therrell, M.D.; Stahle, D.W. A predictive model to locate ancient forests in the Cross Timbers of Osage County, Oklahoma. J. Biogeogr. 1998, 25, 847–854. [Google Scholar] [CrossRef]
- Ryberg, W.A.; Fitzgerald, L.A. Herpetofaunal inventory of Fort Wolters in north-central Texas. Southwest. Nat. 2005, 50, 267–272. [Google Scholar] [CrossRef]
- Dixon, J.R. Amphibians and Reptiles of Texas: With Keys, Taxonomic Synopses, Bibliography, and Distribution Maps, 3rd ed.; Texas A&M University Press: College Station, TX, USA, 2013. [Google Scholar]
- Sullivan, B.K. Long-term shifts in snake populations: A California site revisited. Biol. Conserv. 2000, 94, 321–325. [Google Scholar] [CrossRef]
- Owen, J.D.; Meik, J.M.; Schwertner, T.W. Preliminary nocturnal road survey of snakes in northeastern Swaziland: Effects of agriculture on relative abundance. Herpetol. Rev. 2015, 46, 12–15. [Google Scholar]
- Kéry, M.; Royle, J.A. Prelude and Static Models. In Applied Hierarchical Modeling in Ecology: Analysis of Distribution, Abundance and Species Richness in R and BUGS; Elsevier Academic Press: London, UK, 2016; Volume 1. [Google Scholar]
- U.S. Climate Data. Available online: usclimatedata.com (accessed on 1 April 2023).
- Daltry, J.C.; Ross, T.; Thorpe, R.S.; Wüsterm, W. Evidence that humidity influences snake activity patterns: A field study of the Malayan pit viper Calloselasma rhodostoma. Ecography 1998, 21, 25–34. [Google Scholar] [CrossRef]
- Eskew, E.A.; Todd, B.D. Too cold, too wet, too bright, or just right? Environmental predictors of snake movement and activity. Copeia 2017, 105, 584–591. [Google Scholar] [CrossRef]
- Schalk, C.M.; Weng, Y.H.; Adams, C.S.; Saenz, D. Spatiotemporal patterns of snake captures and activity in upland pine forests. Am. Midl. Nat. 2022, 187, 195–209. [Google Scholar] [CrossRef]
- Lazaridis, E. Lunar; Version 0.2-01; Lunar Phase and Distance, Seasons and Other Environmental Factors; 2022. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 31 October 2022).
- Mangiafico, S.S. Rcompanion; Version 2.4.30; Functions to Support Extension Education Program Evaluation; 2023. [Google Scholar]
- Mazerolle, M.J. AICcmodavg; Version 2.1-1; Model Selection and Multimodal Inference Based on (Q)AIC(c); 2017. [Google Scholar]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Tyre, A.J.; Tenhumberg, B.; Field, S.A.; Niejalke, D.; Parris, K.; Possingham, H.P. Improving precision and reducing bias in biological surveys: Estimating false-negative error rates. Ecol. Appl. 2003, 13, 1790–1801. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Fiske, I.; Chandler, R. unmarked: An R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Soft. 2011, 43, 1–23. [Google Scholar] [CrossRef]
- Werler, J.E.; Dixon, J.R. Texas Snakes: Identification, Distribution, and Natural History; University of Texas Press: Austin, TX, USA, 2000. [Google Scholar]
- Ernst, C.H.; Ernst, E.M. Snakes of the United States and Canada; Smithsonian Books: Washington, DC, USA, 2003. [Google Scholar]
- Bernardino, F.S.; Dalrymple, G.H. Seasonal activity and road mortality of the snakes of the Pa-hay-okee westlands of the Everglades National Park, USA. Biol. Conserv. 1992, 62, 71–75. [Google Scholar] [CrossRef]
- Marques, O.A.V.; Eterovic, A.; Endo, W. Seasonal activity of snakes in the Atlantic Forest in southeastern Brazil. Amphibia-Reptilia 2000, 22, 103–111. [Google Scholar] [CrossRef]
- McDonald, P.J. Snakes on roads: An arid Australian perspective. J. Arid Environ. 2012, 79, 116–119. [Google Scholar] [CrossRef]
- Sun, L.; Shine, R.; Debi, Z.; Zhengren, T. Biotic and abiotic influences on activity patterns of insular pit-vipers (Gloydius shedaoensis, Viperidae) from north-eastern China. Biol. Conserv. 2001, 97, 387–398. [Google Scholar] [CrossRef]
- Houston, D.; Shine, R. Movements and activity patterns of Arafura filesnakes (Serpentes: Acrochordidae) in tropical Australia. Herpetologica 1994, 50, 349–357. [Google Scholar]
- Sperry, J.H.; Ward, M.P.; Weatherhead, P.J. Effects of temperature, moon phase, and prey on nocturnal activity in ratsnakes: An automated telemetry study. J. Herpetol. 2013, 47, 105–111. [Google Scholar] [CrossRef]
- Brown, G.P.; Shine, R. Influence of weather conditions on activity of tropical snakes. Aust. Ecol. 2002, 57, 596–605. [Google Scholar] [CrossRef]
- Platt, J.R. Stong inference. Science 1964, 146, 347–353. [Google Scholar] [CrossRef]
- Laurent, E.J.; Kingsbury, B.A. Habitat separation among three species of water snakes in northwestern Kentucky. J. Herpetol. 2003, 37, 229–235. [Google Scholar] [CrossRef]
- Fitch, H.S. A Kansas Snake Community: Composition and Changes Over 50 Years; Krieger Publishing Company: Malabar, FL, USA, 1999. [Google Scholar]
- Rosen, P.C.; Lowe, C.H. Highway mortality of snakes in the Sonoran Desert of southern Arizona. Biol. Conserv. 1994, 68, 143–148. [Google Scholar] [CrossRef]
- Jochimsen, D.M.; Peterson, C.R.; Harmon, L.J. Influence of ecology and landscape on snake road mortality in a sagebrush-steppe ecosystem. Anim. Conserv. 2014, 17, 583–592. [Google Scholar] [CrossRef]
- Mccardle, L.D.; Fontenot, C.L., Jr.; Lutterschmidt, W.I. Demographic patterns of activity and road mortality from a long-term study of a wetland snake assemblage. Herpetol. Conserv. Biol. 2022, 17, 378–397. [Google Scholar]
- Andrews, K.M.; Gibbons, J.W.; Jochimsen, D.M. Ecological effects of roads on amphibians and reptiles: A literature review. In Urban Herpetology; Mitchell, J.C., Jung Brown, E., Bartholomew, B., Eds.; Society for the Study of Amphibians and Reptiles: Salt Lake City, UT, USA, 2008; Volume 3, pp. 121–143. [Google Scholar]
- Jones, T.R.; Babb, R.D.; Hensley, F.R.; LiWanPo, C.; Sullivan, B.K. Sonoran Desert snake communities at two sites: Concordance and effects of increased road traffic. Herpetol. Conserv. Biol. 2011, 6, 61–71. [Google Scholar]
- Croshaw, D.A.; Cassani, J.R.; Bacher, E.V.; Hancock, T.L.; Everham, E.M., III. Changes in snake abundance after 21 years in southwest Florida, USA. Herpetol. Conserv. Biol. 2019, 14, 31–40. [Google Scholar]
- Dodd, C.K., Jr.; Enge, K.M.; Stuart, J.M. Reptiles on highways in north-central Alabama, USA. J. Herpetol. 1989, 23, 197–200. [Google Scholar] [CrossRef]
- Paterson, J.E.; Baxter-Gilbert, J.; Beaudry, F.; Carstairs, S.; Chow-Fraser, P.; Edge, C.B.; Lentini, A.M.; Litzgus, J.D.; Markle, C.E.; McKeown, K.; et al. Road avoidance and its energetic consequences for reptiles. Ecol. Evol. 2019, 9, 9794–9803. [Google Scholar] [CrossRef]
- Wagner, R.B.; Brune, C.R.; Popescu, V.D. Snakes on a lane: Road type and edge habitat predict hotspots of snake road mortality. J. Nat. Conserv. 2021, 61, 125978. [Google Scholar] [CrossRef]
- Rudolph, D.C.; Burgdorf, S.J. Timber rattlesnakes and Louisiana pine snakes of the west gulf coastal plain: Hypotheses of decline. Texas J. Sci. 1997, 49, 111–122. [Google Scholar]
- Steen, D.A.; Smith, L.L.; Conner, L.M.; Brock, J.C.; Hoss, S.K. Habitat use of sympatric rattlesnake species within the Gulf Coast Plain. J. Wild. Manag. 2007, 71, 759–764. [Google Scholar] [CrossRef]
- Steen, D.A.; McClure, C.J.W.; Brock, J.C.; Rudolph, D.C.; Pierce, J.B.; Lee, J.R.; Humphries, W.J.; Gregory, B.B.; Sutton, W.B.; Smith, L.L.; et al. Landscape-level influences of terrestrial snake occupancy within the southeastern United States. Ecol. Appl. 2012, 22, 1084–1097. [Google Scholar] [CrossRef]
- Mohr, J.R.; Duvall, D. A prairie preference of the Oklahoma timber rattlesnake (Crotalus horridus) on the western fringe of its range. In Biology of Rattlesnakes II; Dreslik, M.J., Hayes, W.K., Beaupre, S.J., Eds.; ECO Herpetological Publishing and Distribution: Rodeo, NM, USA, 2017; pp. 136–141. [Google Scholar]
- Parker, W.S.; Brown, W.S. Species composition and population changes in two complexes of snake hibernacula in northern Utah. Herpetologica 1973, 29, 319–326. [Google Scholar]
- Mendelson, J.R., III; Jennings, W.B. Shifts in the relative abundance of snakes in a desert grassland. J. Herpetol. 1992, 26, 38–45. [Google Scholar] [CrossRef]
- Meik, J.M.; Fontenot, B.E.; Franklin, C.J.; King, C. Apparent natural hybridization between the rattlesnakes Crotalus atrox and C. horridus. Southwest. Nat. 2008, 53, 196–200. [Google Scholar] [CrossRef]
- Hasselman, D.J.; Argo, E.E.; McBride, M.C.; Bentzen, P.; Schultz, T.F.; Perez-Umphrey, A.A.; Palkovacs, E.P. Human disturbance causes the formation of a hybrid swarm between two naturally sympatric fish species. Mol. Ecol. 2014, 23, 1137–1152. [Google Scholar] [CrossRef]
- Grabenstein, K.C.; Taylor, S.A. Breaking barriers: Causes, consequences, and experimental utility of human-mediated hybridization. Trends Ecol. Evol. 2018, 33, 198–212. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Transects | % AOR | Activity | GoF Test | ||
|---|---|---|---|---|---|---|
| A | B | C | ||||
| Agkistrodon laticinctus | 24 | 1 | 30 | 85 | DCN | p = 0.0002 |
| Agkistrodon piscivorus | 0 | 23 | 12 | 69 | DCN | p = 0.0002 |
| Coluber constrictor | 0 | 1 | 0 | 0 | D | |
| Coluber flagellum | 1 | 0 | 2 | 33 | D | |
| Crotalus atrox | 1 | 0 | 5 | 100 | DCN | |
| Crotalus horridus | 7 | 17 | 10 | 56 | DCN | p = 0.092 |
| Haldea striatula | 0 | 3 | 1 | 25 | Semifossorial | |
| Lampropeltis calligaster | 3 | 1 | 1 | 80 | CN | |
| Nerodia erythrogaster | 14 | 17 | 8 | 64 | DCN | p = 0.201 |
| Nerodia rhombifer | 29 | 16 | 17 | 66 | DCN | p = 0.084 |
| Opheodrys aestivus | 0 | 5 | 3 | 0 | D | |
| Pantherophis emoryi | 1 | 0 | 5 | 83 | DCN | |
| Pantherophis obsoletus | 24 | 29 | 21 | 64 | DCN | p = 0.545 |
| Pituophis catenifer | 2 | 0 | 0 | 50 | DC | |
| Storeria dekayi | 2 | 7 | 4 | 69 | Semifossorial | |
| Thamnophis proximus | 24 | 24 | 11 | 56 | DCN | p = 0.060 |
| Transects | |||
|---|---|---|---|
| A | B | C | |
| AOR | 84 [85.5] | 85 [93.3] | 95 [84.2] |
| DOR | 48[46.5] | 59 [50.7] | 36 [45.8] |
| Variable | K | AICc | ΔAICc | Weight | LL | Pseudo-R2 |
|---|---|---|---|---|---|---|
| Barometric pressure (Hg) | 4 | 894.86 | 0.00 | 1 | −443.35 | 0.26 |
| Ordinal date | 4 | 933.58 | 38.72 | 0 | −462.71 | 0.21 |
| Air temperature (°C) | 4 | 933.69 | 38.83 | 0 | −462.76 | 0.21 |
| Humidity (%) | 4 | 943.14 | 48.28 | 0 | −467.49 | 0.17 |
| Wind speed (km/hr) | 2 | 943.64 | 48.78 | 0 | −469.79 | 0.16 |
| Lunar phase | 8 | 973.79 | 78.93 | 0 | −478.60 | 0.10 |
| Species | AICc N | p [95% CI] | AICc G | LRT P | Covar. | Coef. ± SE |
|---|---|---|---|---|---|---|
| A. laticinctus | 215.2 | 0.22 [0.17, 0.28] | 208.7 | 0.002 | ord date | 0.51 ± 0.21 |
| A. piscivorus | 167.1 | 0.16 [0.12, 0.23] | 166.9 | 0.019 | ||
| C. horridus | 183.4 | 0.12 [0.08, 0.16] | 189.6 | 0.120 | ||
| N. erythrogaster | 187.5 | 0.12 [0.09, 0.17] | 183.8 | 0.006 | wind | 0.13 ± 0.21 |
| wind2 | 0.39 ± 0.17 | |||||
| N. rhombifer | 235.4 | 0.21 [0.16, 0.27] | 232.3 | 0.007 | ||
| P. obsoletus | 270.0 | 0.22 [0.18, 0.28] | 261.9 | 0.001 | temp | 0.68 ± 0.19 |
| T. proximus | 245.7 | 0.19 [0.14, 0.24] | 238.4 | 0.002 | temp | 0.63 ± 0.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
King, C.; Meik, J.M. Factors Influencing Activity and Detection of Species in a Cross Timbers Snake Assemblage. Diversity 2023, 15, 952. https://doi.org/10.3390/d15090952
King C, Meik JM. Factors Influencing Activity and Detection of Species in a Cross Timbers Snake Assemblage. Diversity. 2023; 15(9):952. https://doi.org/10.3390/d15090952
Chicago/Turabian StyleKing, Clint, and Jesse M. Meik. 2023. "Factors Influencing Activity and Detection of Species in a Cross Timbers Snake Assemblage" Diversity 15, no. 9: 952. https://doi.org/10.3390/d15090952
APA StyleKing, C., & Meik, J. M. (2023). Factors Influencing Activity and Detection of Species in a Cross Timbers Snake Assemblage. Diversity, 15(9), 952. https://doi.org/10.3390/d15090952