


Deep Aggregations of the Polychaete Amage adspersa (Grube, 1863) in the Ionian Sea (Central Mediterranean Sea) as Revealed via ROV Observations

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
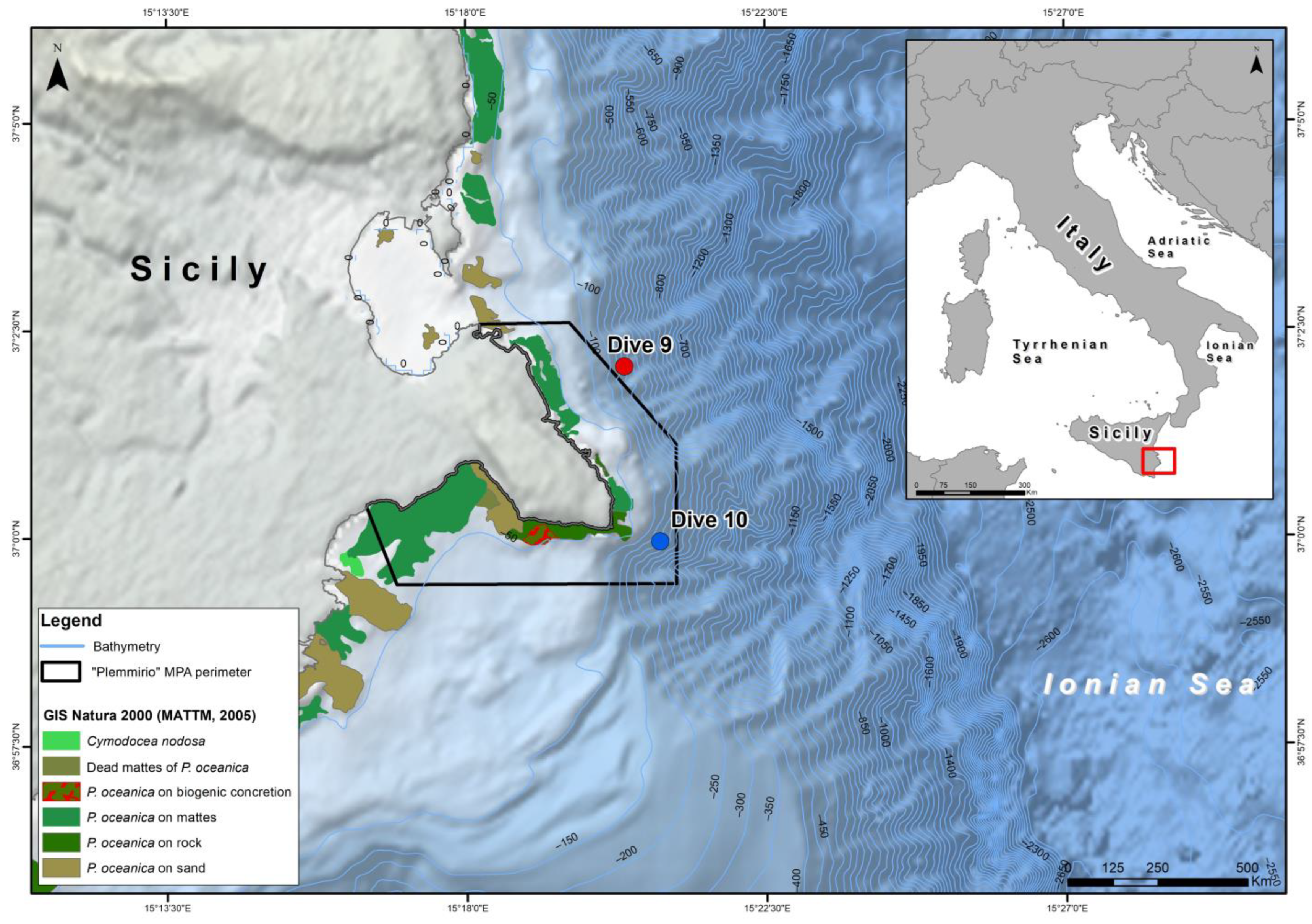
2.1. Study Area
2.2. ROV Survey
2.3. Data Analysis
2.4. Taxonomic Analysis
3. Results
3.1. Taxonomy
- Class POLYCHAETA Grube, 1850
- Order TEREBELLIDA sensu Rouse & Fauchald, 1997
- Family AMPHARETIDAE Malmgren, 1866
- Amage Malmgren, 1866
- Amage adspersa (Grube, 1863)
3.1.1. Material Examined
3.1.2. Comparative Material Examined
3.1.3. Description
3.1.4. Tubes
3.1.5. Methyl green
3.1.6. Distribution
3.1.7. Remarks
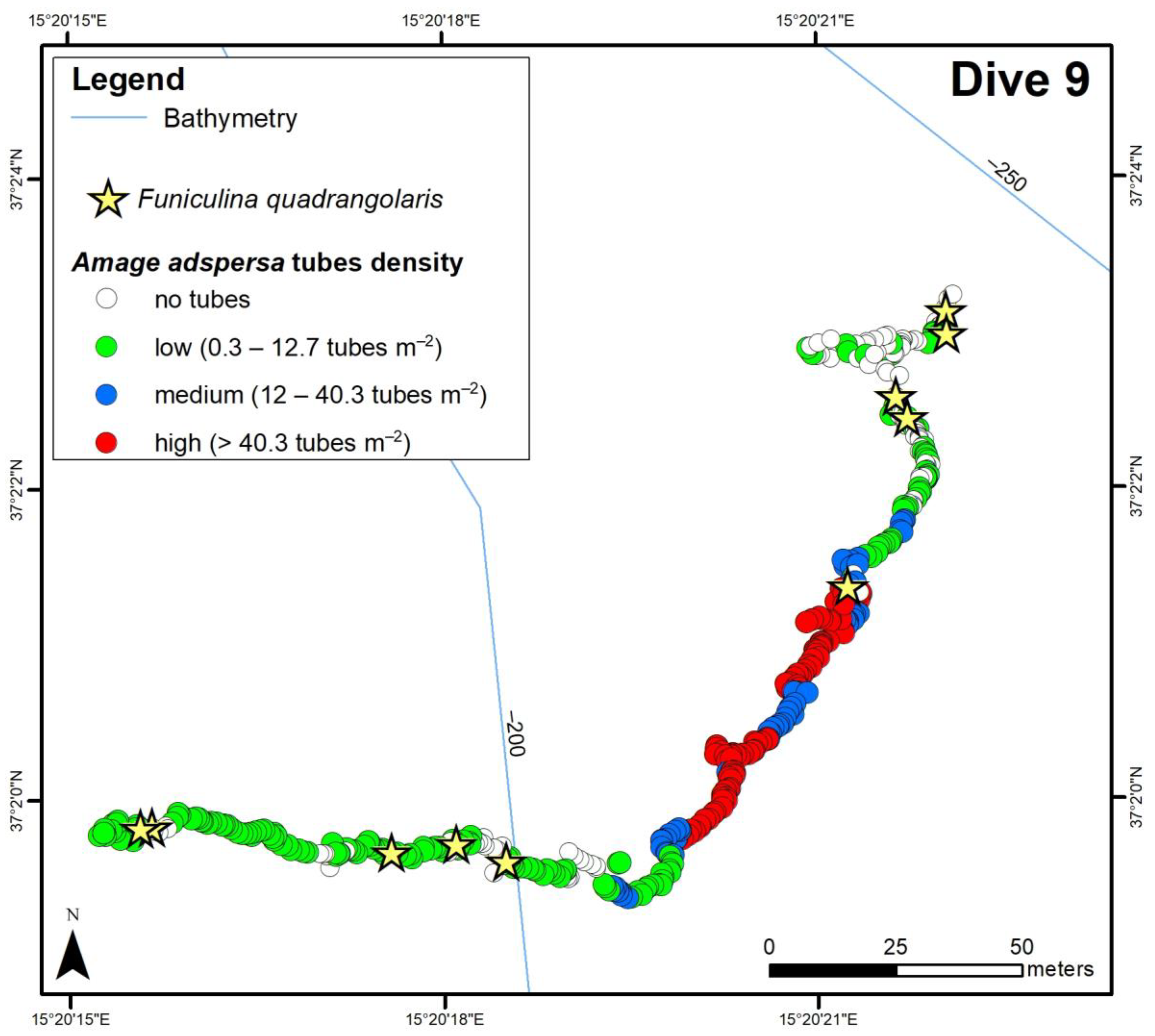
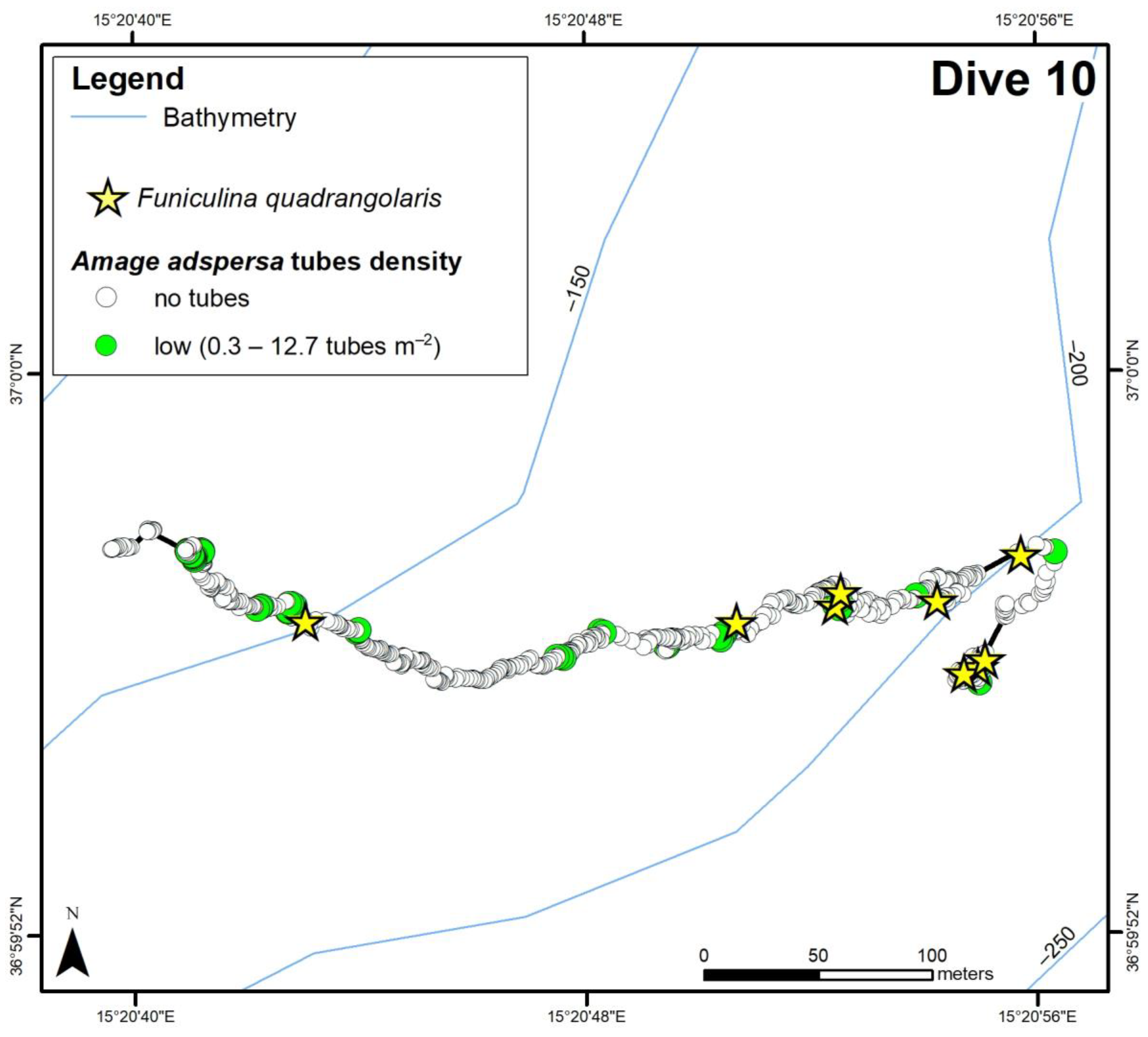
3.2. ROV Analysis
3.2.1. Amage adspersa aggregations
3.2.2. Megafauna
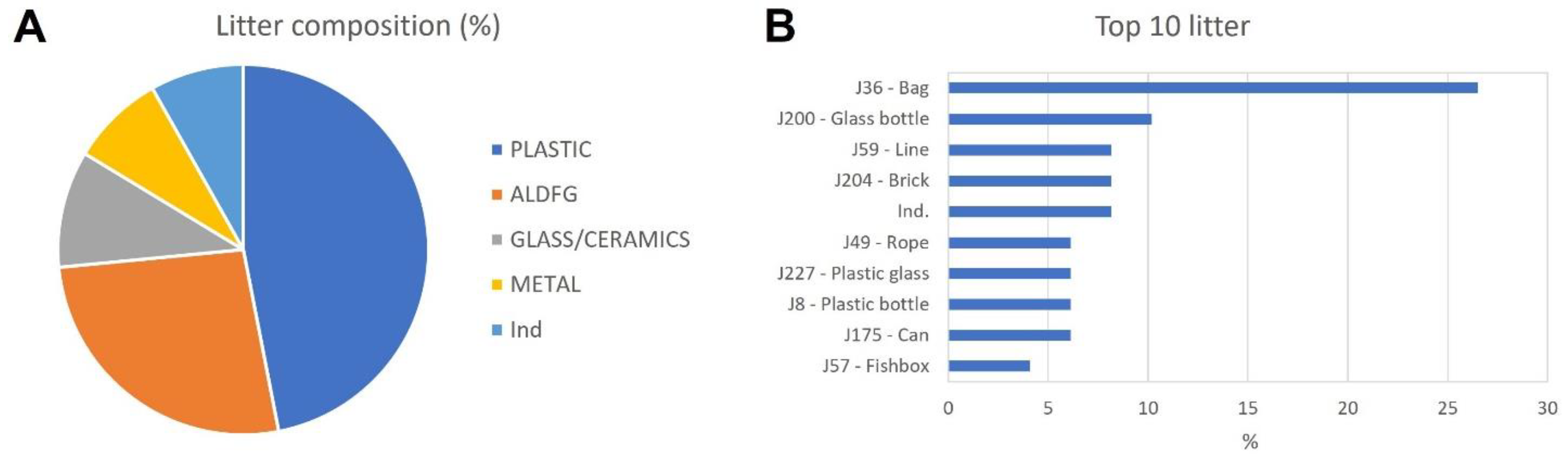
3.2.3. Anthropogenic Impact
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Wright, J.P.; Jones, C.G. The concept of organisms as ecosystem engineers ten years on: Progress, limitations, and challenges. BioScience 2006, 56, 203–209. [Google Scholar] [CrossRef]
- Buhl-Mortensen, L.; Vanreusel, A.; Gooday, A.J.; Levin, L.A.; Priede, I.G.; Buhl-Mortensen, P.; Gheerardyn, H.; King, N.J.; Raes, M. Biological structures as a source of habitat heterogeneity and biodiversity on the deep ocean margins. Mar. Ecol. 2010, 31, 21–50. [Google Scholar] [CrossRef]
- Eckman, J.E.; Nowell, A.R.; Jumars, P.A. Sediment destabilization by animal tubes. J. Mar. Res. 1981, 39, 361–374. [Google Scholar]
- Andersen, F.Ø.; Kristensen, E. Effects of burrowing macrofauna on organic matter decomposition in coastal marine sediments. Symp. Zool. Soc. Lond. 1991, 63, 69–88. [Google Scholar]
- Meadows, P.S.; Meadows, A. The geotechnical and geochemical implications of bioturbation in marine sedimentary ecosystems. In Symposium of the Zoolgical Society of London; Clarendon Press: Oxford, UK, 1991; Volume 63, pp. 157–181. [Google Scholar]
- Gili, J.-M.; Coma, R. Benthic suspension feeders: Their paramount role in littoral marine food webs. Trends Ecol. Evol. 1998, 13, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Enrichetti, F.; Baldrighi, E.; Bavestrello, G.; Betti, F.; Canese, S.; Costa, A.; del Pasqua, M.; Giangrande, A.; Langeneck, J.; Misic, C.; et al. Ecological role and phylogenetic position of a new habitat-forming species (Canalipalpata, Sabellidae) from the Mediterranean mesophotic soft bottoms. Estuar. Coast. Shelf Sci. 2022, 265, 107737. [Google Scholar] [CrossRef]
- Jackson, J.B.C. Distribution and ecology of clonal and aclonal benthic invertebrates. In Population Biology and Evolution of Clonal Organisms; Jackson, J.B.C., Buss, W., Cook, R.E., Ashmun, J.V., Eds.; Yale University Press: London, UK, 1985; pp. 297–355. [Google Scholar]
- Wood, R. Reef Evolution; Oxford University Press: Oxford, UK; New York, NY, USA, 1999; ISBN 978-0-19-857784-3. [Google Scholar]
- Rossi, S.; Bramanti, L.; Gori, A.; Orejas, C. Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Gutiérrez, J.L.; Jones, C.G.; Strayer, D.L.; Iribarne, O.O. Mollusks as ecosystem engineers: The role of shell production in aquatic habitats. Oikos 2003, 101, 79–90. [Google Scholar] [CrossRef]
- Cardone, F.; Corriero, G.; Longo, C.; Mercurio, M.; Onen Tarantini, S.; Gravina, M.F.; Lisco, S.; Moretti, M.; De Giosa, F.; Giangrande, A.; et al. Massive bioconstructions built by Neopycnodonte cochlear (Mollusca, Bivalvia) in a mesophotic environment in the central Mediterranean Sea. Sci. Rep. 2020, 10, 6337. [Google Scholar] [CrossRef]
- Giangrande, A.; Gambi, M.C.; Gravina, M.F. Polychaetes as habitat former: Structure and function. In Perspectives on the Marine Animal Forests of the World; Rossi, S., Bramanti, L., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 219–237. ISBN 978-3-030-57054-5. [Google Scholar]
- Levinton, J. Bioturbators as ecosystem engineers: Control of the sediment fabric, inter-individual interactions, and material fluxes. In Linking Species & Ecosystems; Jones, C.G., Lawton, J.H., Eds.; Springer: Boston, MA, USA, 1995; pp. 29–36. ISBN 978-1-4615-1773-3. [Google Scholar]
- Woodin, S.A.; Wethey, D.S.; Volkenborn, N. Infaunal hydraulic ecosystem engineers: Cast of characters and impacts. Integr. Comp. Biol. 2010, 50, 176–187. [Google Scholar] [CrossRef]
- Bruschetti, M. Role of reef-building, ecosystem engineering Polychaetes in shallow water ecosystems. Diversity 2019, 11, 168. [Google Scholar] [CrossRef]
- Fager, E.W. Marine sediments: Effects of a tube-building Polychaete. Science 1964, 143, 356–359. [Google Scholar] [CrossRef]
- Carey, D.A. Sedimentological effects and palaeoecological implications of the tube-building Polychaete Lanice Conchilega Pallas. Sedimentology 1987, 34, 49–66. [Google Scholar] [CrossRef]
- Rabaut, M.; Vincx, M.; Degraer, S. Do Lanice conchilega (Sandmason) aggregations classify as reefs? quantifying habitat modifying effects. Helgol. Mar. Res. 2009, 63, 37–46. [Google Scholar] [CrossRef]
- Callaway, R.; Desroy, N.; Dubois, S.F.; Fournier, J.; Frost, M.; Godet, L.; Hendrick, V.J.; Rabaut, M. Ephemeral bio-engineers or reef-building Polychaetes: How stable are aggregations of the tube worm Lanice conchilega (Pallas, 1766)? Integr. Comp. Biol. 2010, 50, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Sanders, H.L.; Goudsmit, E.M.; Mills, E.L.; Hampson, G.E. A study of the intertidal fauna of Barnstable Harbor, Massachusetts. Limnol. Oceanogr. 1962, 7, 63–79. [Google Scholar] [CrossRef]
- Surugiu, V. Systematics and ecology of species of the Polydora-complex (Polychaeta: Spionidae) of the Black Sea. Zootaxa 2012, 3518, 45–65. [Google Scholar] [CrossRef]
- Bolam, S.G.; Fernandes, T.F. Dense aggregations of Pygospio elegans (Claparede): Effect on macrofaunal community structure and sediments. J. Sea Res. 2003, 49, 171–185. [Google Scholar] [CrossRef]
- Gallagher, E.D.; Jumars, P.A.; Trueblood, D.D. Facilitation of soft-bottom benthic succession by tube builders. Ecology 1983, 64, 1200–1216. [Google Scholar] [CrossRef]
- Bell, S.S. Habitat complexity of Polychaete tube-caps: Influence of architecture on dynamics of a meioepibenthic assemblage. J. Mar. Res. 1985, 43, 647–671. [Google Scholar] [CrossRef]
- Luckenbach, M.W. Effects of adult infauna on new recruits: Implications for the role of biogenic refuges. J. Exp. Mar. Biol. Ecol. 1987, 105, 197–206. [Google Scholar] [CrossRef]
- Fauchald, K.; Jumars, P.A. The diet of worms: A study of Polychaete feeding guilds. Oceanogr. Mar. Biol. Annu. Rev. 1979, 17, 193–284. [Google Scholar]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of worms emended: An update of Polychaete feeding guilds. Annu. Rev. Mar. Sci. 2015, 7, 497–520. [Google Scholar] [CrossRef] [PubMed]
- Hilbig, B. Family Ampharetidae Malmgren, 1867. In Taxonomic Atlas of the benthic fauna of the santa maria basin and western Santa Barbara Channel. Volume 7. The Annelida Part 4. Polychaeta: Flabelligeridae to Sternaspidae; Blake, J.A., Hilbig, B., Scott, P.V., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 2000; pp. 169–230. [Google Scholar]
- Ebbe, G.; Purschke, G. Ampharetidae Malmgren, 1866. In Pleistoannelida, Sedentaria III and Errantia I; Purschke, G., Westheide, W., Böggemann, G., Eds.; De Gruyter: Berlin, Germany, 2020; ISBN 978-3-11-029170-4. [Google Scholar]
- Warwick, R.M.; Davies, J.R. The Distribution of sublittoral macrofauna communities in the Bristol Channel in relation to the substrate. Estuar. Coast. Mar. Sci. 1977, 5, 267–288. [Google Scholar] [CrossRef]
- Cacabelos, E.; Moreira, J.; Lourido, A.; Troncoso, J.S. Ecological features of Terebellida fauna (Annelida, Polychaeta) from Ensenada de San Simón (NW Spain). Anim. Biodivers. Conserv. 2011, 34, 141–150. [Google Scholar] [CrossRef]
- Hilbig, B. Faunistic and zoogeographical characterization of the benthic infauna on the Carolina Continental Slope. Deep Sea Res. Part II Top. Stud. Oceanogr. 1994, 41, 929–950. [Google Scholar] [CrossRef]
- Schüller, M.; Ebbe, B. Global distributional patterns of selected deep-sea Polychaeta (Annelida) from the Southern Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 1737–1751. [Google Scholar] [CrossRef]
- GBIF—The Global Biodiversity Information Facility. What is GBIF? 2023. Available online: https://www.gbif.org/what-is-gbif (accessed on 13 March 2020).
- Fauvel, F.L.A. Première Note Préliminaire sur les Polychètes Provenant des Campagnes de l’Hirondelle et de La Princesse-Alice, ou Deposées Dans le Musée Océanographique de Monaco; Bulletin de l’Institut Océanographique de Monaco: Monaco, 1907. [Google Scholar]
- Fauvel, P. Polychètes Sédentaires; Faune de France: Paris, France, 1927. [Google Scholar]
- Fauvel, P. Contribution a la faune des Annélides Polychètes du Maroc; Mémoires de la Société Des Sciences Naturelles du Maroc; Institut Scientifique Chérifien: Rabat, Morocco, 1936. [Google Scholar]
- Spärck, R. The benthonic animal communities of the coastal waters. Zool. Icel. 1937, 6, 1–45. [Google Scholar]
- Clark, R.B. New records of sub-littoral Polychætes from the Clyde Sea area, with a description of a new species. Proc. R. Soc. Edinb. Sect. B Biol. Sci. 1953, 65, 1–26. [Google Scholar] [CrossRef]
- Langerhans, P. Die wurmfauna von Madeira, IV. Z. Für Wiss. Zool. 1884, 40, 247–285. [Google Scholar]
- Intes, A.; Le Loeuff, P. Les annelides Polychetes de Cote d’Ivoire. II: Polychetes Sedentaires—Compte rendu systematique. Cah. ORSTOM Série Océanographie 1977, 15, 215–249. [Google Scholar]
- Lo Bianco, S. Gli Anellidi Tubicoli trovati nel Golfo di Napoli (Con 3 Tavole). R. Accad. Delle Sci. Fis. Mat. 1893, 5, 1–13. [Google Scholar]
- Wolff, T. Utilization of seagrass in the deep sea. Aquat. Bot. 1976, 2, 161–174. [Google Scholar] [CrossRef]
- Dimech, M.; Borg, J.A.; Schembri, P.J. Motile macroinvertebrate assemblages associated with submerged Posidonia oceanica litter accumulations. Biol. Mar. Mediterr. 2006, 13, 130–133. [Google Scholar]
- Maggiore, F.; Berthon, J.; Boudouresque, C.; Lawrence, J. Données préliminaires sur les relations entre Paracentrotus lividus, Arbacia lixula et le phytobenthos dans la Baie de Port-Cros (Var, France, Méditerranée). In Proceedings of the colloque international sur Paracentrotus lividus et les Oursins comestibles; GIS Posidonie: Marseille, France, 1987; pp. 65–82. [Google Scholar]
- Boudouresque, C.F.; Verlaque, M. Paracentrotus lividus. In Sea urchins: Biology and ecology; Lawrence, J.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 297–327. ISBN 978-0-12-396491-5. [Google Scholar]
- Tunesi, L.; Casazza, G.; Dalù, M.; Giorgi, G.; Silvestri, C. The implementation of the Marine Strategy Framework Directive in Italy: Knowledge to support the management. Biol. Mar. Mediterr. 2013, 20, 35–52. [Google Scholar]
- Danovaro, R.; Fanelli, E.; Canals, M.; Ciuffardi, T.; Fabri, M.-C.; Taviani, M.; Argyrou, M.; Azzurro, E.; Bianchelli, S.; Cantafaro, A.; et al. Towards a Marine Strategy for the deep Mediterranean Sea: Analysis of current ecological status. Mar. Policy 2020, 112, 103781. [Google Scholar] [CrossRef]
- Corriero, G.; Pierri, C.; Mercurio, M.; Nonnis Marzano, C.; Onen Tarantini, S.; Gravina, M.F.; Lisco, S.; Moretti, M.; De Giosa, F.; Valenzano, E.; et al. A Mediterranean mesophotic coral reef built by non-symbiotic scleractinians. Sci. Rep. 2019, 9, 3601. [Google Scholar] [CrossRef]
- Bo, M.; Al Mabruk, S.A.A.; Balistreri, P.; Bariche, M.; Batjakas, I.E.; Betti, F.; Bilan, M.; Canese, S.; Cattaneo-vietti, R.; Corsini-foka, M.; et al. New records of rare species in the Mediterranean Sea (October 2020). Mediterr. Mar. Sci. 2020, 21, 608–630. [Google Scholar] [CrossRef]
- Toma, M.; Enrichetti, F.; Bavestrello, G.; Canese, S.; Cau, A.; Andaloro, F.; Angiolillo, M.; Greco, S.; Bo, M. Brachiopod fauna from the deep Mediterranean Sea: Distribution patterns and ecological preferences. Diversity 2022, 14, 753. [Google Scholar] [CrossRef]
- Bonanno, A.; Goncharov, S.; Mazzola, S.; Popov, S.; Cuttitta, A.; Patti, B.; Basilone, G.; Di Nieri, A.; Patti, C.; Aronica, S.; et al. Acoustic evaluation of anchovy larvae distribution in relation to oceanography in the Cape Passero area (Strait of Sicily). Chem. Ecol. 2006, 22, S265–S273. [Google Scholar] [CrossRef]
- Bergamasco, A.; Malanotte-Rizzoli, P. The circulation of the Mediterranean Sea: A historical review of experimental investigations. Adv. Oceanogr. Limnol. 2010, 1, 11–28. [Google Scholar] [CrossRef]
- Celentano, P.; Falco, P.; Zambianchi, E. Surface connection between the Ionian Sea and different areas of the Mediterranean derived from drifter data. Deep Sea Res. Part I Oceanogr. Res. Pap. 2020, 166, 103431. [Google Scholar] [CrossRef]
- Gačić, M.; Borzelli, G.L.E.; Civitarese, G.; Cardin, V.; Yari, S. Can internal processes sustain reversals of the Ocean Upper Circulation? The Ionian Sea example. Geophys. Res. Lett. 2010, 37, L09608. [Google Scholar] [CrossRef]
- Gačić, M.; Civitarese, G.; Eusebi Borzelli, G.L.; Kovačević, V.; Poulain, P.-M.; Theocharis, A.; Menna, M.; Catucci, A.; Zarokanellos, N. On the relationship between the Decadal Oscillations of the northern Ionian Sea and the salinity distributions in the Eastern Mediterranean. J. Geophys. Res. Ocean. 2011, 116, C12002. [Google Scholar] [CrossRef]
- Ministero Dell’ambiente e della Tutela del Territorio e del Mare. Cartografia delle praterie di Posidonia oceanica (L.) Délile lungo le coste italiane; Ministero Dell’ambiente e della Tutela del Territorio e del Mare: Roma, Italy, 2005. [Google Scholar]
- Fleet, D.; Vlachogianni, T.; Hanke, G. Joint List of Litter Categories for Marine Macro-Litter Monitoring: Manual for the Application of the Classification System; EUR 30348 EN; Publications Office of the European Union: Luxembourg, 2021; ISBN 978-92-76-21445-8. [Google Scholar]
- Angiolillo, M.; Gérigny, O.; Valente, T.; Fabri, M.-C.; Tambute, E.; Rouanet, E.; Claro, F.; Tunesi, L.; Vissio, A.; Daniel, B.; et al. Distribution of seafloor litter and its interaction with benthic organisms in deep waters of the Ligurian Sea (Northwestern Mediterranean). Sci. Total Environ. 2021, 788, 147745. [Google Scholar] [CrossRef]
- Grube, A.E. Beschreibung neuer oder wenig bekannter Anneliden. Sechster Beitrag. Arch. Für Naturgeschichte Berl. 1863, 29, 37–69. [Google Scholar] [CrossRef]
- Drago, N.; Albertelli, N.; Cattaneo, G. Osservazioni faunistiche sul benthos dell’Isola di Capraia. Ann. Mus. Civ. Stor. Nat. Genova 1978, 82, 72–77. [Google Scholar]
- Chimenz Gusso, C.; Gravina, M.; Maggiore, F. Temporal variations in soft bottom benthic communities in central Tyrrhenian Sea (Italy). Archo Oceanogr. Limnol. 2001, 22, 175–182. [Google Scholar]
- Gourret, O. Documents sur les Térébellacees et les Ampharetiens du Golfe de Marseille. Mémoires Société Zool. Fr. 1901, 14, 373–387. [Google Scholar]
- Tsikopoulou, I.; Nikolopoulou, S.; Digitation of les fonds de peche pres d’Alexandrie. XI Annelides Polychetes. Fauvel, P. Notes et Memoires No 19; Direction des Recherches des Pecheries, 1937, Egypt. 2015. Available online: http://ipt.medobis.eu/resource?r=egyptexpeditionpolychaeta (accessed on 16 April 2023).
- Cantone, G.; Fassari, G.; Mollica, E. Benthic Polychaetous Annelids of Ognina Bay (Eastern Sicily, Italy). Atti Della Soc. Toscana Sci. Nat. Ser. B 1993, 100, 39–48. [Google Scholar]
- Dean, H.K. The use of Polychaetes (Annelida) as indicator species of marine pollution: A review. Rev. Biol. Trop. 2008, 56, 11–38. [Google Scholar] [CrossRef]
- Pedel, L.; Fabri, M. Description et Cartographie des Biocénoses Benthiques Rencontrées au Cours de la Campagne MEDSEACAN. 2011, p. 109. Available online: https://archimer.ifremer.fr/doc/00044/15513/ (accessed on 4 June 2023).
- Calizza, E.; Costantini, M.L.; Carlino, P.; Bentivoglio, F.; Orlandi, L.; Rossi, L. Posidonia oceanica Habitat loss and changes in litter-associated biodiversity organization: A stable isotope-based preliminary study. Estuar. Coast. Shelf Sci. 2013, 135, 137–145. [Google Scholar] [CrossRef]
- Como, S.; Magni, P.; Baroli, M.; Casu, D.; De Falco, G.; Floris, A. Comparative analysis of macrofaunal species richness and composition in Posidonia oceanica, Cymodocea nodosa and leaf litter beds. Mar. Biol 2008, 153, 1087–1101. [Google Scholar] [CrossRef]
- Gallmetzer, I.; Pflugfelder, B.; Zekely, J.; Ott, J.A. Macrofauna diversity in Posidonia oceanica detritus: Distribution and diversity of mobile macrofauna in shallow sublittoral accumulations of Posidonia oceanica Detritus. Mar. Biol. 2005, 147, 517–523. [Google Scholar] [CrossRef]
- Remy, F.; Michel, L.N.; Mascart, T.; De Troch, M.; Lepoint, G. Trophic Ecology of macrofauna inhabiting seagrass litter accumulations is related to the pulses of dead leaves. Estuar. Coast. Shelf Sci. 2021, 252, 107300. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Pergent, G.; Pergent-Martini, C.; Ruitton, S.; Thibaut, T.; Verlaque, M. The necromass of the Posidonia oceanica seagrass meadow: Fate, role, ecosystem services and vulnerability. Hydrobiologia 2016, 781, 25–42. [Google Scholar] [CrossRef]
- Massé, C.; Garabetian, F.; Deflandre, B.; Maire, O.; Costes, L.; Mesmer-Dudons, N.; Duchêne, J.-C.; Bernard, G.; Grémare, A.; Ciutat, A. Feeding ethology and surface sediment reworking by the ampharetid Polychaete Melinna palmata Grube, 1870: Effects on sediment characteristics and aerobic bacterial community composition. J. Exp. Mar. Biol. Ecol. 2019, 512, 63–77. [Google Scholar] [CrossRef]
- Salvati, E.; Bo, M.; Rondinini, C.; Battistoni, A.; Teofili, C. Lista Rossa IUCN dei Coralli Italiani; Comitato Italiano IUCN e Ministero dell’Ambiente e Della Tutela del Territorio e del Mare: Roma, Italy, 2014. [Google Scholar]
- Bo, M.; Numa, C.; Del Mar Otero, M.; Orejas, C.; Garrabou, J.; Cerrano, C.; Kružic, P.; Antoniadou, C.; Aguilar, R.; Kipson, S.; et al. Overview of the Conservation Status of Mediterranean Anthozoa; IUCN International Union for Conservation of Nature: Gland, Switzerland, 2017; ISBN 978-2-8317-1845-3. [Google Scholar]
- FAO. International Guidelines for the Management of Deep-Sea Fisheries in the High Seas; FAO: Rome, Italy, 2009; ISBN 978-92-5-006258-7. [Google Scholar]
- Mačić, V.; Đorđević, N.; Đurović, M.; Petović, S.; Russo, T. Improving knowledge of Funiculina quadrangularis and Vulnerable Marine Ecosystems in the South Adriatic. Mediterr. Mar. Sci. 2022, 23, 805–816. [Google Scholar] [CrossRef]
- Pierdomenico, M.; Russo, T.; Ambroso, S.; Gori, A.; Martorelli, E.; D’Andrea, L.; Gili, J.-M.; Chiocci, F.L. Effects of trawling activity on the bamboo-coral Isidella elongata and the sea pen Funiculina quadrangularis along the Gioia Canyon (Western Mediterranean, Southern Tyrrhenian Sea). Prog. Oceanogr. 2018, 169, 214–226. [Google Scholar] [CrossRef]
- Salvalaggio, V.; Fabi, G.; Punzo, E.; Santelli, A.; Tassetti, N.; Strafella, P.; Cvitkovic, I.; Despalatović, M.; Raicevich, S.; Scarcella, G. Distribution of the sea pens Virgularia mirabilis and Funiculina quadrangularis (Cnidaria, Anthozoa) in the northern and central Adriatic Sea. In Proceedings of the CIESM International Commission for Scientific Exploration of the Mediterranean Sea, Kiel, Germany, 14 September 2016. [Google Scholar]
- Available online: Globalfishingwatch.org (accessed on 13 April 2023).
- Available online: Marinetraffic.com (accessed on 13 April 2023).
- Tunesi, L.; Diviacco, G.; Mo, G. Observations by submersible on the biocoenosis of the deep-sea corals off Portofino Promontory (Northwestern Mediterranean Sea). In Proceedings of the First International Symposium on Deep-Sea Corals; Willison, J.H.M., Hall, J., Eds.; Ecology Action Centre and Nova Scotia Museum: Halifax, NS, Canada, 2001; pp. 76–87. [Google Scholar]
- Lauria, V.; Garofalo, G.; Fiorentino, F.; Massi, D.; Milisenda, G.; Piraino, S.; Russo, T.; Gristina, M. Species distribution models of two critically endangered deep-sea octocorals reveal fishing impacts on vulnerable marine ecosystems in central Mediterranean Sea. Sci. Rep. 2017, 7, 8049. [Google Scholar] [CrossRef] [PubMed]
- Bastari, A.; Pica, D.; Ferretti, F.; Micheli, F.; Cerrano, C. Sea pens in the Mediterranean Sea: Habitat suitability and opportunities for ecosystem recovery. ICES J. Mar. Sci. 2018, 75, 1722–1732. [Google Scholar] [CrossRef]
- De Clippele, L.H.; Buhl-Mortensen, P.; Buhl-Mortensen, L. Fauna associated with cold water gorgonians and sea pens. Cont. Shelf Res. 2015, 105, 67–78. [Google Scholar] [CrossRef]
- Pierdomenico, M.; Casalbore, D.; Chiocci, F.L. Massive benthic litter funnelled to deep sea by flash-flood generated hyperpycnal flows. Sci. Rep. 2019, 9, 5330. [Google Scholar] [CrossRef] [PubMed]
- Tubau, X.; Canals, M.; Lastras, G.; Rayo, X.; Rivera, J.; Amblas, D. Marine litter on the floor of deep submarine canyons of the northwestern Mediterranean Sea: The role of hydrodynamic processes. Prog. Oceanogr. 2015, 134, 379–403. [Google Scholar] [CrossRef]
- Angiolillo, M.; Fortibuoni, T. Impacts of marine litter on Mediterranean reef systems: From shallow to deep waters. Front. Mar. Sci. 2020, 7, 581966. [Google Scholar] [CrossRef]
- Canals, M.; Pham, C.K.; Bergmann, M.; Gutow, L.; Hanke, G.; van Sebille, E.; Angiolillo, M.; Buhl-Mortensen, L.; Cau, A.; Ioakeimidis, C.; et al. The quest for seafloor macrolitter: A critical review of background knowledge, current methods and future prospects. Environ. Res. Lett. 2021, 16, 023001. [Google Scholar] [CrossRef]
- Sleight, V.; Calafat, A.; Rogers, A.D.; Narayanaswamy, B.E.; Thompson, R.C. The deep sea is a major sink for microplastic debris. R. Soc. Open Sci. 2014, 1, 140317. [Google Scholar] [CrossRef]
- de Carvalho-Souza, G.F.; Llope, M.; Tinôco, M.S.; Medeiros, D.V.; Maia-Nogueira, R.; Sampaio, C.L.S. Marine litter disrupts ecological processes in reef systems. Mar. Pollut. Bull. 2018, 133, 464–471. [Google Scholar] [CrossRef]
- Saldanha, H.J.; Sancho, G.; Santos, M.N.; Puente, E.; Gaspar, M.B.; Bilbao, A.; Monteiro, C.C.; Gomez, E.; Arregi, L. The use of biofouling for ageing lost nets: A case study. Fish. Res. 2003, 64, 141–150. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| D9 | D10 | |
|---|---|---|
| Date | 9 July 2021 | 9 July 2021 |
| Total time | 1:00 | 1:04 |
| Latitude start | 37°01′47.49″ N | 36°59′54.69″ N |
| Longitude start | 15°20′35.14″ E | 15°20′55.14″ E |
| Depth range (m) | 166–236 | 120–196 |
| Track length (m) | 373 | 651 |
| SU (no.) | 15 | 26 |
| Presence of Amage tubes (no. of SU) | 15 | 15 |
| Amage tube (no.) | 1373 | 204 |
| Amage tube density (tubes m−2 ± se) | 28.3 ± 9.7 | 2.5 ± 0.7 |
| Litter items (no.) | 26 | 23 |
| Litter density (items 100 m−2 ± se) | 4.78 ± 1.9 | 3.53 ± 1.2 |
| Taxa | D9 | D10 | |||
|---|---|---|---|---|---|
| no. | % | no. | % | ||
| Porifera | Porifera ind. | 2 | - | - | - |
| Cnidaria | Cerianthus membranaceus (Gmelin, 1791) | 5 | 20 | 33 | 69.2 |
| Funiculina quadrangularis (Pallas, 1766) | 9 | 33 | 10 | 38.5 | |
| Virgularia mirabilis (Müller, 1776) | 11 | 47 | 8 | 23.1 | |
| Annelida | Amage adspersa (Grube, 1863) | >1373 | 100 | 204 | 57.7 |
| Bonellia viridis Rolando, 1822 | 1 | 6.7 | 10 | 11.5 | |
| Polychaeta ind. | 14 | 33 | 18 | ||
| Myxicola sp. | 1 | 6.7 | 1 | 3.8 | |
| Mollusca | Sepia officinalis Linnaeus, 1758 | - | - | 1 | 3.8 |
| Tethys fimbria Linnaeus, 1767 | - | - | 1 | 3.8 | |
| Tonna galea (Linnaeus, 1758) | - | - | 1 | 3.8 | |
| Plesionika sp. | 14 | 47 | - | - | |
| Loligo sp. | 1 | 6.7 | - | - | |
| Echinodermata | Cidaridae | 12 | 53 | 26 | 38.5 |
| Holoturia sp. | - | - | 1 | 3.8 | |
| Parastichopus regalis (Cuvier, 1817) | 1 | 6.7 | 1 | 7.7 | |
| Ophiuroidea | 1 | 6.7 | 1 | 3.8 | |
| Leptometra phalangium (Müller, 1841) | 1 | 6.7 | - | ||
| Chordata | Chlorophthalmus agassizi Bonaparte, 1840 | 14 | 47 | 10 | 26.9 |
| Helicolenus dactylopterus (Delaroche, 1809) | 2 | 13 | 5 | 19.2 | |
| Lepidorhombus whiffiagonis (Walbaum, 1792) | 5 | 33 | 2 | 7.7 | |
| Lepidorhombus boscii (Risso, 1810) | - | - | 1 | 3.8 | |
| Macroramphosus scolopax (Linnaeus, 1758) | 4 | 6.7 | 38 | 46.2 | |
| Ophisurus serpens (Linnaeus, 1758) | 1 | 6.7 | 1 | 3.8 | |
| Peristedion cataphractum (Linnaeus, 1758) | 1 | 6.7 | 2 | 7.7 | |
| Scorpaena elongata (Cadenat, 1943) | 1 | 6.7 | 1 | 3.8 | |
| Serranus cabrilla (Linnaeus, 1758) | - | - | 2 | 3.8 | |
| Litter | Plastic | 22 | 66.6 | 10 | 23.1 |
| Glass/Ceramics | - | - | 9 | 23.1 | |
| Metal | - | - | 4 | 15.3 | |
| Ind. | 4 | 13.3 | - | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angiolillo, M.; Bertasi, F.; Grossi, L.; Loia, M.; Vani, D.; Rende, S.F.; Giusti, M.; Tunesi, L. Deep Aggregations of the Polychaete Amage adspersa (Grube, 1863) in the Ionian Sea (Central Mediterranean Sea) as Revealed via ROV Observations. Diversity 2023, 15, 906. https://doi.org/10.3390/d15080906
Angiolillo M, Bertasi F, Grossi L, Loia M, Vani D, Rende SF, Giusti M, Tunesi L. Deep Aggregations of the Polychaete Amage adspersa (Grube, 1863) in the Ionian Sea (Central Mediterranean Sea) as Revealed via ROV Observations. Diversity. 2023; 15(8):906. https://doi.org/10.3390/d15080906
Chicago/Turabian StyleAngiolillo, Michela, Fabio Bertasi, Laura Grossi, Marco Loia, Danilo Vani, Sante Francesco Rende, Michela Giusti, and Leonardo Tunesi. 2023. "Deep Aggregations of the Polychaete Amage adspersa (Grube, 1863) in the Ionian Sea (Central Mediterranean Sea) as Revealed via ROV Observations" Diversity 15, no. 8: 906. https://doi.org/10.3390/d15080906
APA StyleAngiolillo, M., Bertasi, F., Grossi, L., Loia, M., Vani, D., Rende, S. F., Giusti, M., & Tunesi, L. (2023). Deep Aggregations of the Polychaete Amage adspersa (Grube, 1863) in the Ionian Sea (Central Mediterranean Sea) as Revealed via ROV Observations. Diversity, 15(8), 906. https://doi.org/10.3390/d15080906