
Wild Plant Diversity and Soil Characteristics of Desert Roadside Vegetation in the Eastern Desert


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
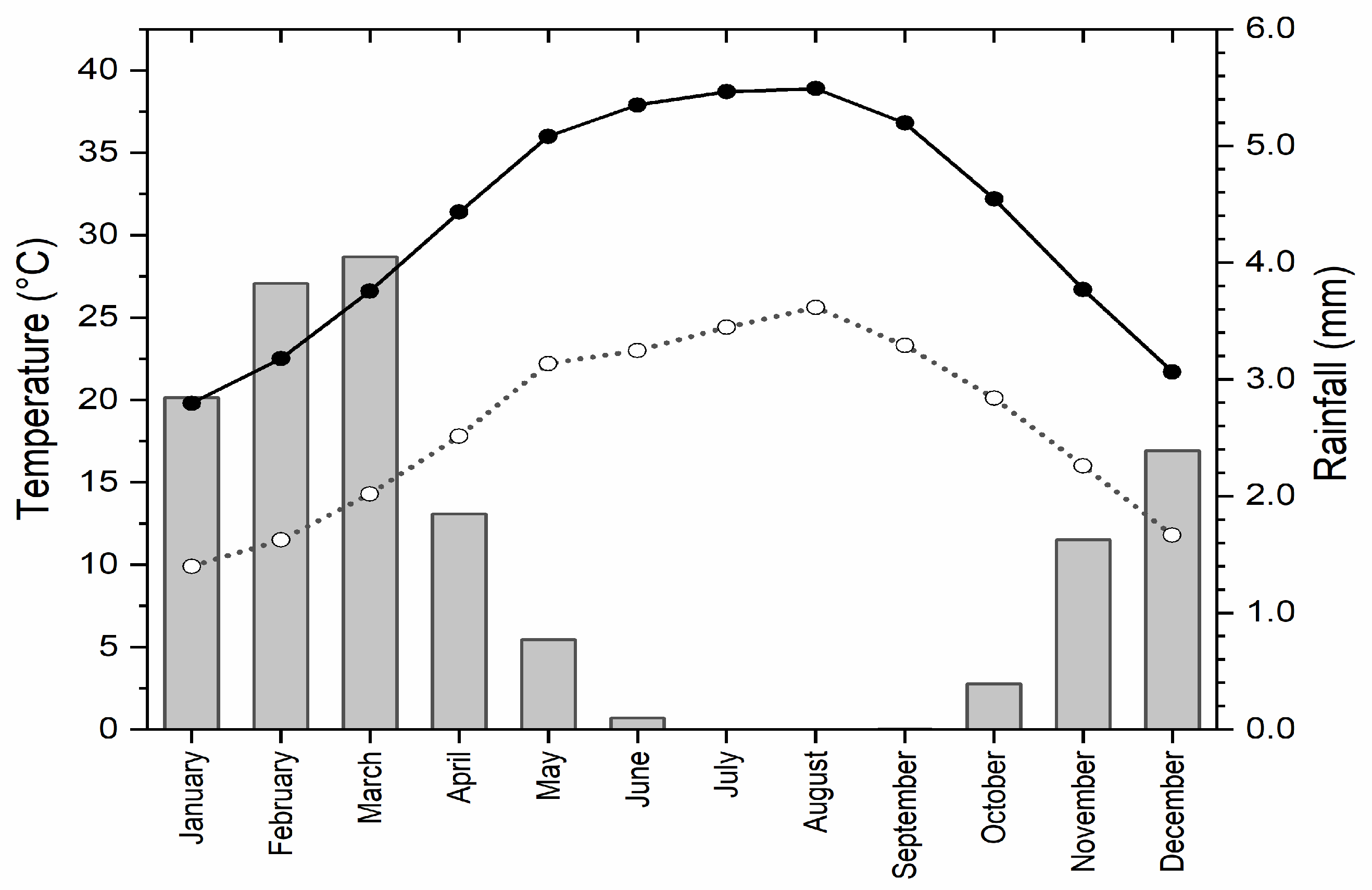
2.1. Site Description
2.2. Vegetation Sampling and Species Identification
2.3. Soil Sampling and Analyses
2.4. Data Analysis
3. Results
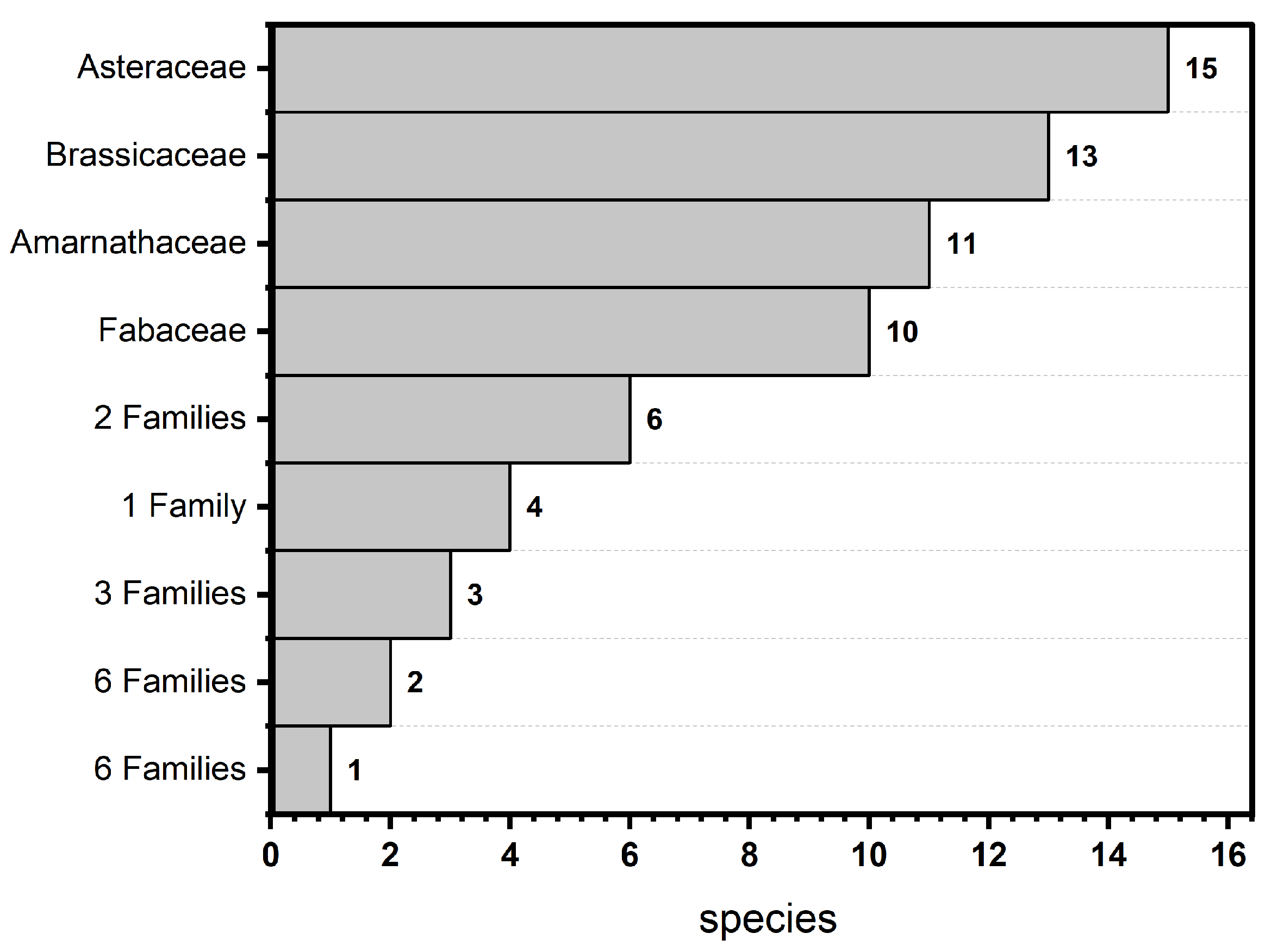
3.1. Floristic Composition
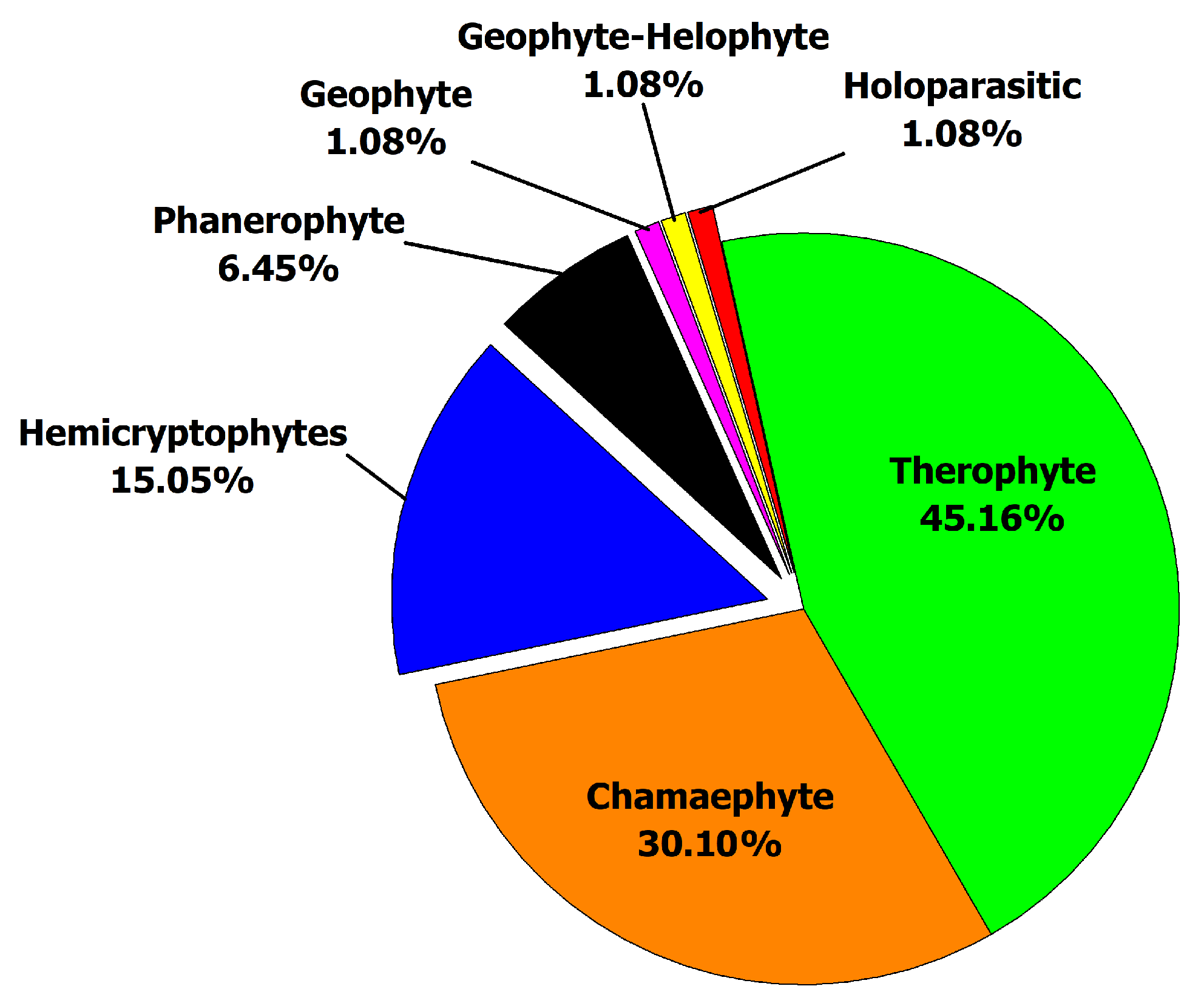
3.2. Life-Form Spectra
3.3. Chorological Affinities
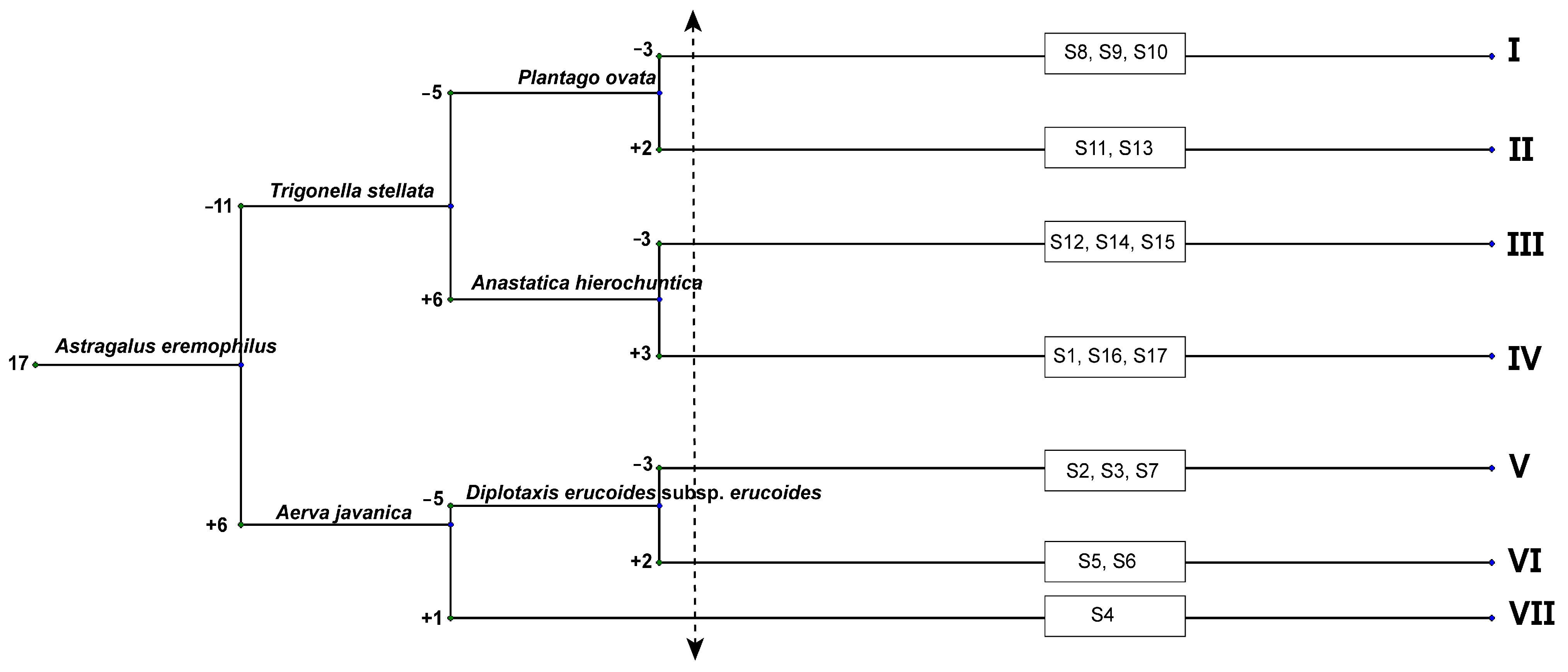
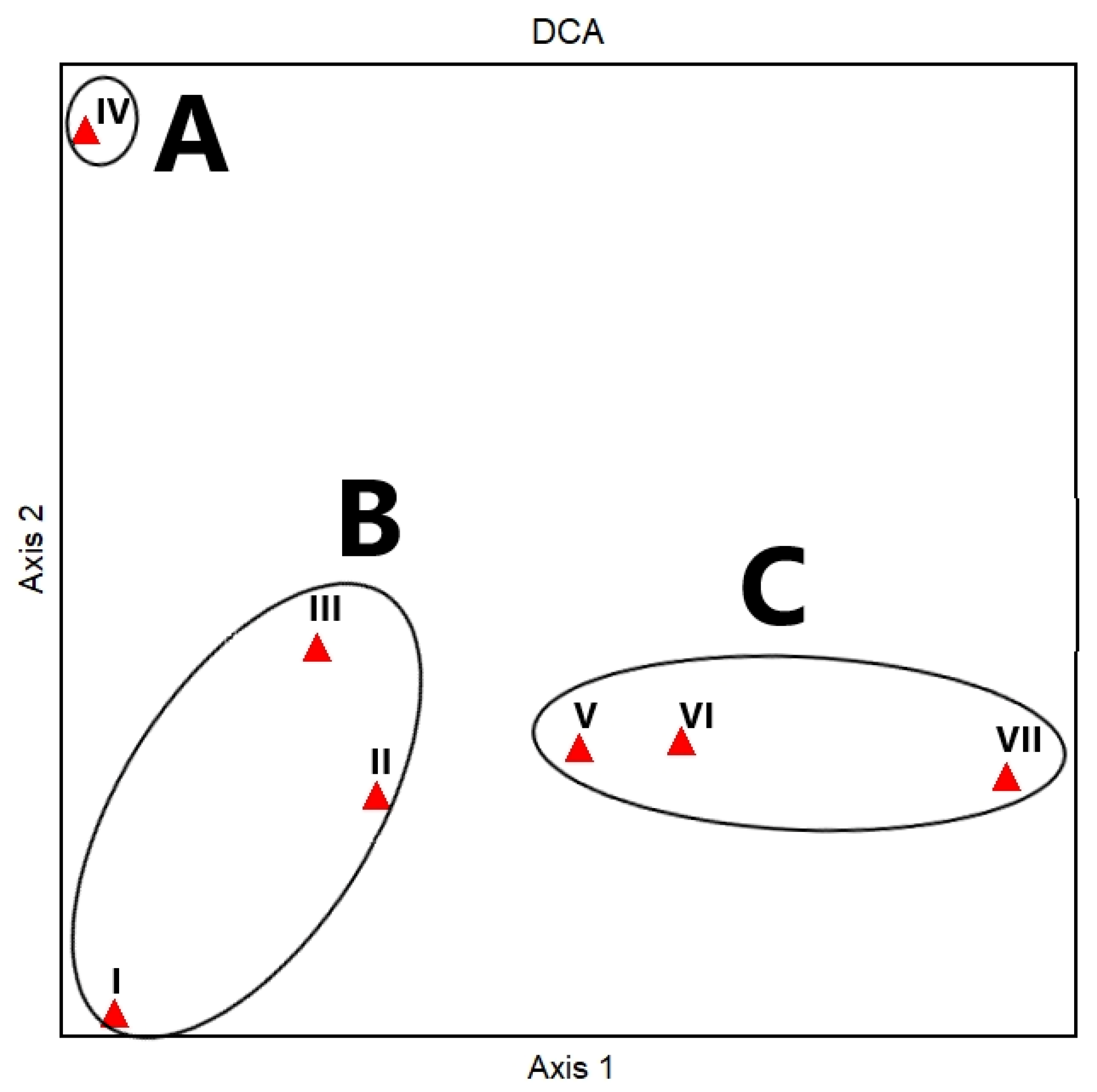
3.4. Vegetation Analysis
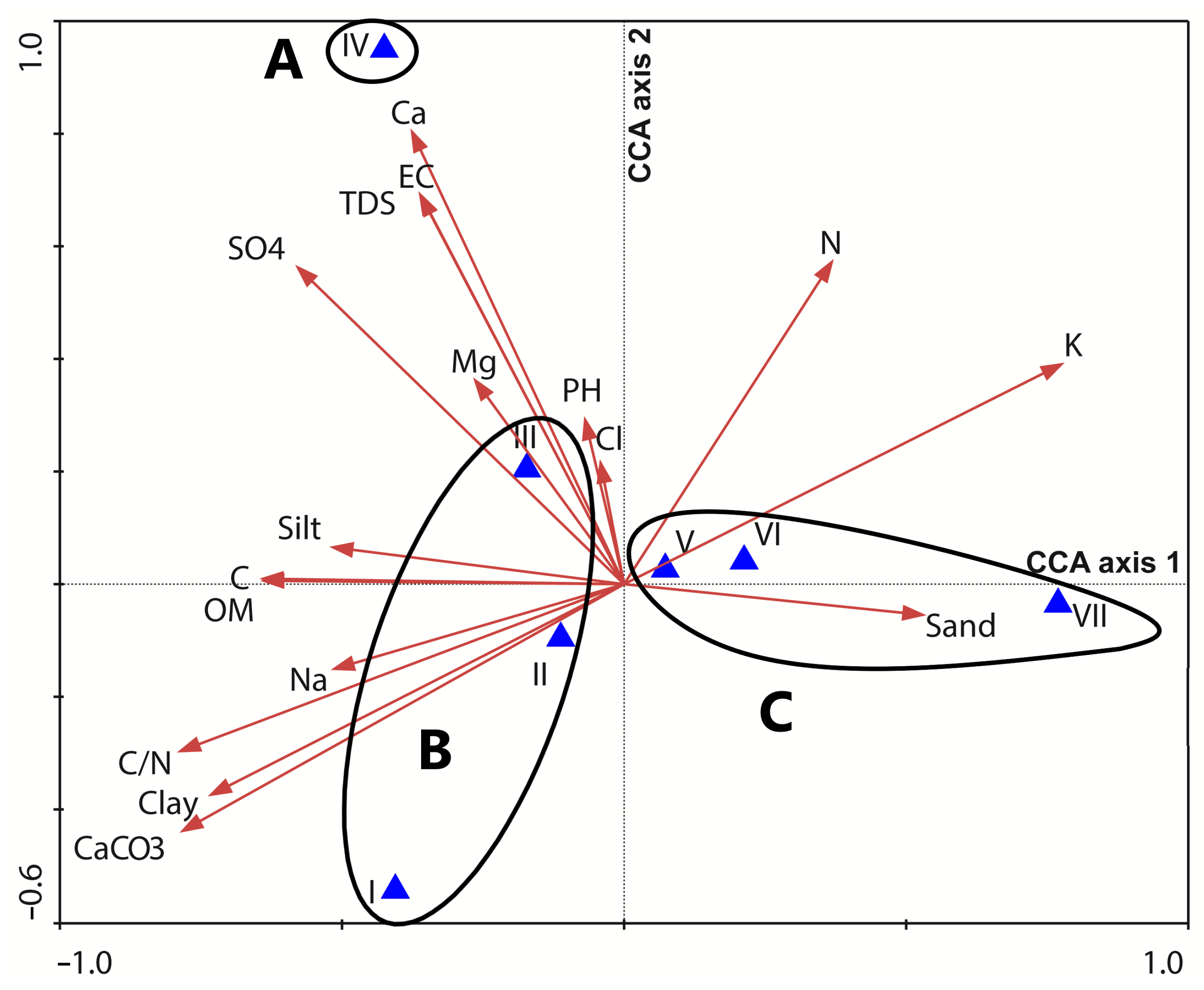
3.5. Plant Community and Soil Correlation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Taxa | Growth Form | Life Span | Life Form | Chorotype |
|---|---|---|---|---|---|
| Acanthaceae | Blepharis ciliaris (L.) B.L.Burtt | Subshrub | Per. | Cham. | IT + SS + SZ |
| Amaranthaceae | Aerva javanica (Burm.f.) Juss. ex Schult. | Subshrub | Per. | Cham. | PAL |
| Anabasis setifera Moq. | Subshrub | Per. | Cham. | IT + SS + SZ | |
| Atriplex turcomanica (Moq.) Boiss. | Subshrub | Per. | Cham. | IT + ME+ SS + SZ | |
| Bassia eriophora (Schrad.) AsCham. | Herb | Ann. | Ther. | IT + SS | |
| Caroxylon imbricatum (Forssk.) Moq. | Shrub | Per. | Cham. | IT + ME + SS + SZ | |
| Caroxylon volkensii (Schweinf. & AsCham.) Akhani & Roalson | Herb | Ann. | Ther. | IT + SS | |
| Cornulaca aucheri Moq. | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Cornulaca monacantha Delile | Subshrub | Per. | Cham. | IT + ME + SS + SZ | |
| Halocnemum strobilaceum (Pall.) M.Bieb. | Subshrub | Per. | Cham. | ES + IT + ME + SS + SZ | |
| Suaeda pruinosa Lange | Shrub | Per. | Cham. | IT + ME + SS | |
| Suaeda vera Forssk. ex J.F.Gmel. | Shrub | Per. | Cham. | ES + ME + SS | |
| Aerva javanica (Burm.f.) Juss. ex Schult. | Subshrub | Per. | Cham. | PAL | |
| Apocynaceae | Cynanchum acutum L. | Climbing subshrub | Per. | Cham. | ES + IT + ME + SS + SJ |
| Asteraceae | Artemisia judaica L. | Shrub | Per. | Cham. | IT + ME + SS + SZ |
| Brocchia cinerea (Delile) Vis. | Herb | Ann. | Ther. | IT + ME + SS | |
| Centaurea aegyptiaca L. | Subshrub | Per. | Cham. | SS + SZ | |
| Centaurea scoparia Sieber ex Spreng. | Subshrub | Per. | Cham. | ME + SS | |
| Ifloga spicata (Forssk.) SCham.Bip. | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Launaea capitata (Spreng.) Dandy | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Launaea mucronata subsp. cassiniana (Jaub. & Spach) N.Kilian | Subshrub | Per. | Hemicr. | IT + ME + SS + SZ | |
| Launaea mucronata (Forssk.) Muschl. subsp. mucronata | Subshrub | Per. | Hemicr. | IT + ME + SS + SZ | |
| Launaea nudicaulis (L.) Hook.f. | Subshrub | Per. | Hemicr. | IT + ME + SS + SZ | |
| Pluchea dioscoridis (L.) DC. | Shrub | Per. | Phan. | IT + ME + SS + SZ | |
| Pulicaria incisa (Lam.) DC. | Herb | Per. | Hemicr. | SS + SZ | |
| Pulicaria undulata (L.) C.A.Mey. | Herb | Per. | Cham. | IT + ME + SS + SZ | |
| Reichardia tingitana (L.) Roth | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Senecio glaucus subsp. coronopifolius (Maire) C.Alexander | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Sonchus oleraceus L. | Herb | Ann. | Ther. | ES + IT + ME + SS + SZ | |
| Zoegea purpurea Fresen. | Herb | Ann. | Ther. | IT + ME + SS | |
| Boraginaceae | Gastrocotyle hispida (Forssk.) Bunge | Herb | Ann. | Ther. | IT + ME + SS + SZ |
| Trichodesma africanum (L.) Sm. | Herb | Per. | Cham. | CAP + GC + IT + ME + SS + SZ | |
| Brassicaceae | Anastatica hierochuntica L. | Herb | Ann. | Ther. | IT + ME + SS + SZ |
| Diplotaxis acris (Forssk.) Boiss. | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Diplotaxis erucoides subsp. erucoides | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Diplotaxis harra (Forssk.) Boiss. | Herb | Per. | Hemicr. | IT + ME + SS + SZ | |
| Eremobium aegyptiacum (Spreng.) AsCham. ex Boiss. | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Farsetia aegyptia Turra | Herb | Per. | Cham. | IT + ME + SS + SZ | |
| Matthiola arabica Boiss. | Herb | Per. | Hemicr. | SS | |
| Matthiola longipetala (Vent.) DC. | Herb | Ann. | Ther. | IT + ME + SS | |
| Morettia philaeana (Delile) DC. | Subshrub | Per. | Hemicr. | SS + SZ | |
| Savignya parviflora (Delile) Webb | Herb | Ann. | Ther. | IT + ME + SS | |
| Schouwia purpurea (Forssk.) Schweinf. | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Sisymbrium irio L. | Herb | Ann. | Ther. | ES + IT + ME + SJ + SS + SZ | |
| Zilla spinosa (L.) Prantl | Subshrub | Per. | Cham. | IT + ME + SS + SZ | |
| Caryophyllaceae | Polycarpaea repens (Forssk.) AsCham. & Schweinf. | Subshrub | Per. | Hemicr. | IT + ME + SS + SZ |
| Pteranthus dichotomus Forssk. | Herb | Ann. | Ther. | IT + ME + SS | |
| Spergularia diandra (Guss.) Heldr. | Herb | Ann. | Ther. | ES + IT + ME + SJ + SS + SZ | |
| Cleomaceae | Cleome africana BotsCham. | Herb | Ann. | Ther. | IT + ME + SS |
| Cleome amblyocarpa Barratte & Murb. Barratte & Murb. | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Convolvulaceae | Convolvulus hystrix Vahl | Shrub | Per. | Cham. | SS + SZ |
| Convolvulus pilosellifolius Desr. | Subshrub | Per. | Cham. | IT + ME + SS + SZ | |
| Euphorbiaceae | Euphorbia retusa Forssk. | Herb | Ann. | Ther. | IT + ME + SS |
| Fabaceae | Astragalus arpilobus subsp. hauarensis (Boiss.) Podlech | Herb | Ann. | Ther. | IT + ME + SS + SZ |
| Astragalus eremophilus Boiss. | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Astragalus spinosus (Forssk.) Muschl. | Subshrub | Per. | Cham. | IT + ME + SS | |
| Astragalus trigonus DC. | Subshrub | Per. | Cham. | IT + ME + SS | |
| Astragalus vogelii (Webb) Bornm. | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Lotus arabicus Sol. ex L. | Herb | Ann. | Ther. | GC + SS + SZ | |
| Lotus halophilus Boiss. & Spruner | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Lotus hebranicus Hochst. ex Brand | Subshrub | Per. | Hemicr. | SS + SZ | |
| Taverniera aegyptiaca Boiss. | Subshrub | Per. | Cham. | SS + SZ | |
| Trigonella stellata Forssk. | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Geraniaceae | Erodium arborescens (Desf.) Willd. | Subshrub | Per. | Hemicr. | ME + SS |
| Erodium oxyrhinchum subsp. bryoniifolium (Boiss.) Schönb.-Tem. | Herb | Per. | Hemicr. | IT + ME + SS | |
| Monsonia nivea (Decne.) Webb | Herb | Per. | Hemicr. | IT + ME + SS + SZ | |
| Malvaceae | Malva parviflora L. | Herb | Ann. | Ther. | ES + IT + ME + SS + SZ |
| Orobanchaceae | Cistanche phelypaea (L.) Cout. | Herb | Per. | Holopar. | IT + ME + SS + SZ |
| Plantaginaceae | Plantago ciliata Desf. | Herb | Ann. | Ther. | IT + ME + SS + SZ |
| Plantago ovata Forssk. | Herb | Ann. | Ther. | IT + ME + SS + SZ | |
| Poaceae | Cynodon dactylon (L.) Pers. | Herb | Per. | Geo. | COSM |
| Polypogon monspeliensis (L.) Desf. | Herb | Ann. | Ther. | ES + IT + ME + SJ + SS + SZ | |
| Rostraria cristata (L.) Tzvelev | Herb | Ann. | Ther. | ES + IT + ME + SS + SZ | |
| Schismus barbatus (L.) Thell. | Herb | Ann. | Ther. | CAP + ES + IT + ME + SJ + SS + SZ | |
| Stipagrostis ciliata (Desf.) De Winter | Herb | Per. | Hemicr. | CAP + IT + ME + SS + SZ | |
| Phragmites australis (Cav.) Trin. ex Steud. | Reed | Per. | Geo.–Hel. | COSM | |
| Polygonaceae | Calligonum comosum L’Hér. | Shrub | Per. | Phan. | IT + ME + SS + SZ |
| Rumex vesicarius L. | Herb | Ann. | Ther. | IT + ME + SJ + SS + SZ | |
| Resedaceae | Ochradenus baccatus Delile | Shrub | Per. | Phan. | IT + ME + SS + SZ |
| Reseda muricata C.Presl | Subshrub | Per. | Cham. | ME + SS + SZ | |
| Reseda pruinosa Delile | Herb | Ann. | Ther. | ME + SS + SZ | |
| Reseda urnigera Webb | Herb | Ann. | Ther. | ME + SS | |
| Solanaceae | Hyoscyamus desertorum (AsCham. ex Boiss.) Täckh. | Herb | Ann. | Ther. | IT + ME + SS |
| Hyoscyamus muticus L. | Subshrub | Per. | Cham. | IT + ME + SS + SZ | |
| Tamaricaceae | Tamarix aphylla (L.) H.Karst. | Tree | Per. | Phan. | IT + ME + SS + SZ |
| Tamarix nilotica (Ehrenb.) Bunge | Tree | Per. | Phan. | IT + ME + SS + SZ | |
| Tamarix passerinoides Delile ex Decne. | Shrub | Per. | Phan. | IT + ME + SS + SZ | |
| Urticaceae | Forsskaolea tenacissima L. | Herb | Per. | Hemicr. | IT + ME + SS + SZ |
| Zygophyllaceae | Tribulus macropterus Boiss. | Herb | Ann. | Ther. | IT + ME + SS + SZ |
| Zygophyllum arabicum (L.) Christenh. & Byng | Subshrub | Per. | Cham. | IT + ME + SS + SZ | |
| Zygophyllum coccineum L. | Subshrub | Per. | Cham. | IT + ME + SS + SZ | |
| Zygophyllum indicum (Burm.f.) Christenh. & Byng | Subshrub | Per. | Cham. | IT + ME + SS + SZ | |
| Zygophyllum molle (Delile) Christenh. & Byng | Subshrub | Per. | Cham. | IT + ME + SS | |
| Zygophyllum simplex L. | Herb | Ann. | Ther. | CAP + GC + IT + ME + SS + SZ |
References
- Said, R. The Geology of Egypt; Routledge: Abingdon, UK, 2017. [Google Scholar]
- Zahran, M.A.; Willis, A.J. The Vegetation of Egypt; Springer: Berlin/Heidelberg, Germany, 2009; Volume 2. [Google Scholar]
- El Hadidi, M.N. Vegetation of the Nubian Desert (Nabta Region). In Prehistory of the Eastern Sahara; Wendorf, F., Schild, R., Eds.; Academic Press: London, UK, 1980; pp. 345–351. [Google Scholar]
- Abd El-Ghani, M.M. Environmental Correlates of Species Distribution in Arid Desert Ecosystems of Eastern Egypt. J. Arid Environ. 1998, 38, 297–313. [Google Scholar] [CrossRef]
- Fossati, J.; Pautou, G.; Peltier, J.P. Wadi Vegetation of the North-Eastern Desert of Egypt. Feddes Repert. 1998, 109, 313–327. [Google Scholar] [CrossRef]
- El-Ghani, M.A.; Salama, F.; Salem, B.; El-Hadidy, A.; Abdel-Aleem, M. Biogeographical Relations of a Hyperarid Desert Flora in Eastern Egypt. Afr. J. Ecol. 2014, 52, 173–191. [Google Scholar] [CrossRef]
- Abdelaal, M. Current Status of the Floristic Composition in Wadi Hagul, Northwest Suez Gulf, Egypt. Rend. Lincei 2017, 28, 81–92. [Google Scholar] [CrossRef]
- Ward, D. The Biology of Deserts; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Abd El-Wahab, R.H. Plant Assemblage and Diversity Variation with Human Disturbances in Coastal Habitats of the Western Arabian Gulf. J. Arid Land 2016, 8, 787–798. [Google Scholar] [CrossRef]
- Solbrig, O.T. The Origin and Function of Biodiversity. Environment 1991, 33, 16–38. [Google Scholar] [CrossRef]
- Moustafa, A.A.; Abd El-Wahab, R.H.; Zaghloul, M.S. Conservation and Sustainable Use of Medicinal Plants in Arid and Semi-Arid Ecosystems of Egypt; Final Report; 1999. Available online: https://www.researchgate.net/publication/281438983_Conservation_and_Sustainable_use_of_Medicinal_Plants_in_Arid_and_Semi-arid_Ecosystems_of_Sinai_Egypt (accessed on 17 July 2023).
- Adla, K.; Dejan, K.; Neira, D.; Dragana, Š. Degradation of Ecosystems and Loss of Ecosystem Services. In One Health: Integrated Approach to 21st Century Challenges to Health; Academic Press: Cambridge, MA, USA, 2022; pp. 281–327. [Google Scholar]
- Gazoulis, I.; Antonopoulos, N.; Kanatas, P.; Karavas, N.; Bertoncelj, I.; Travlos, I. Invasive Alien Plant Species—Raising Awareness of a Threat to Biodiversity and Ecological Connectivity (EC) in the Adriatic-Ionian Region. Diversity 2022, 14, 387. [Google Scholar] [CrossRef]
- Salama, F.M.; Ahmed, M.K.; El-Tayeh, N.A.; Hammad, S.A. Vegetation Analysis, Phenological Patterns and Chorological Affinities in Wadi Qena, Eastern Desert, Egypt. Afr. J. Ecol. 2012, 50, 193–204. [Google Scholar] [CrossRef]
- Nakahama, N.; Hirasawa, Y.; Minato, T.; Hasegawa, M.; Isagi, Y.; Shiga, T. Recovery of Genetic Diversity in Threatened Plants through Use of Germinated Seeds from Herbarium Specimens. Plant Ecol. 2015, 216, 1635–1647. [Google Scholar] [CrossRef]
- Zohary, M. Man and Vegetation in the Middle East. In Man’s Impact on Vegetation; Holzner, W., Werger, M., Ikusima, I., Eds.; Junk: The Hague, The Netherlands, 1983; pp. 287–295. [Google Scholar]
- Holzapfel, C.; Schmidt, W. Roadside Vegetation along Transects in the Judean Desert. Isr. J. Bot. 1990, 39, 263–270. [Google Scholar]
- El-Ghani, M.M.A. Vegetation along a Transect in the Hijaz Mountains (Saudi Arabia). Feddes Repert. 1994, 105, 517–530. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.; Salama, F.; Salem, B.; El-Hadidy, A.; Abdel-Aleem, M. Phytogeography of the Eastern Desert Flora of Egypt. Wulfenia 2017, 24, 97–120. [Google Scholar]
- El-Amier, Y.A.; Abdulkader, O.M. Vegetation and Species Diversity in the Northern Sector of Eastern Desert, Egypt. West African J. Appl. Ecol. 2015, 23, 75–95. [Google Scholar]
- Shaltout, K.H.; El-Din, A.S. Habitat Types and Plant Communities along a Transect in the Nile Delta Region. Feddes Repert. 1988, 99, 153–162. [Google Scholar] [CrossRef]
- Ayyad, M.A.; Ghabbour, S.I.; Goodall, D.W. Hot Deserts of Egypt and the Sudan. Ecosyst. World 1986, 12, 149–202. [Google Scholar]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley and Sons: New York, NY, USA, 1974. [Google Scholar]
- Boulos, L. Flora of Egypt, Volume 1: Azollaceae—Oxalidaceae; Al Hadara Publishing: Cairo, Egypt, 1999. [Google Scholar]
- Boulos, L. Flora of Egypt, Volume 2: Geraniaceae–Boraginaceae; Al Hadara Publishing: Cairo, Egypt, 2000. [Google Scholar]
- Boulos, L. Flora of Egypt, Volume 3: Verbenaceae-Compositae; Al Hadara Publishing: Cairo, Egypt, 2002. [Google Scholar]
- Boulos, L. Flora of Egypt. Monocotyledons (Alimataceae-Orchidaceae); Al Hadara: Cairo, Egypt, 2005; Volume 4. [Google Scholar]
- Täckholm, V. Students’ Flora of Egypt, 2nd ed.; Cairo University: Cairo, Egypt, 1974. [Google Scholar]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet. Available online: http://powo.science.kew.org/ (accessed on 30 April 2023).
- Hassib, M. Distribution of Plant Communities in Egypt. Bull. Fac. Sci. Univ. Fouad 1 1951, 29, 59–261. [Google Scholar]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- White, F.; Leonard, J.; Léonard, J. Phytogeographical Links between Africa and Southwest Asia. Flora Veg. Mundi 1991, IX, 229–246. [Google Scholar]
- Kilmer, V.J.; Alexander, L.T. Methods of Making Mechanical Analyses of Soils. Soil Sci. 1949, 68, 15–24. [Google Scholar] [CrossRef]
- Sparks, D.L.; Helmke, P.A.; Page, A.L. Methods of Soil Analysis: Chemical Methods; SSSA. 1996. Available online: https://www.waterboards.ca.gov/waterrights/water_issues/programs/bay_delta/california_waterfix/exhibits/docs/Islands/II_41.pdf (accessed on 17 July 2023).
- Jackson ML Soil Chemical Analysis-Advanced Course, 2nd ed.; Department of Soil Science, University of Madison: Madison, WI, USA, 1969.
- Williams, V.; Twine, S. Flame Photometric Method for Sodium, Potassium and Calcium. Mod. Methods Plant Anal. 1960, 5, 3–5. [Google Scholar]
- Johnson, C.M.; Ulrich, A. 2. Analytical Methods for Use in Plant Analysis. Bull. Calif. Agric. Exp. Stn. 1959, 766, 27–78. [Google Scholar]
- Allen, S.E. Chemical Analysis. In Methods in Plant Ecology; Moore, P.D., Chapman, S.B., Eds.; Blackwell: Oxford, UK, 1986; pp. 285–344. [Google Scholar]
- Black, C.A.; Evans, D.D.; Dinauer, R.C. Methods of Soil Analysis; American Society of Agronomy: Madison, WI, USA, 1965; Volume 9. [Google Scholar]
- APHA, A.-W. Standard Methods for the Examination of Water; Ignatius Press: San Francisco, CA, USA, 1998; Volume 21, pp. 3–37. [Google Scholar]
- Bremner, J.M. Nitrogen-Total. In Methods of Soil Analysis, Part 3: Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2018; pp. 1085–1121. [Google Scholar]
- Henderson, P.A.; Seaby, R.M.H. Community Analysis Package (CAP), Version 1.2; Pisces Conservation Ltd. IRC House: Lymington, UK, 1999.
- Frosini, B.V. Descriptive Measures of Ecological Diversity; Università Cattolica del Sacro Cuore, Istituto di Statistica: Milan, Italy, 2003. [Google Scholar]
- Eilertsen, O.; Okland, R.H.; Okland, T.; Pedersen, O. Data Manipulation and Gradient Length Estimation in DCA Ordination. J. Veg. Sci. 1990, 1, 261–270. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002; Available online: www.canoco.com (accessed on 11 April 2023).
- Coura, R.D.S.; Nardi, N.B. The State of the Art of Adeno-Associated Virus-Based Vectors in Gene Therapy. Virol. J. 2007, 4, 1–299. [Google Scholar] [CrossRef]
- Jongman, R.H.G.; Ter Braak, C.J.F.; Van Tongeren, O.F.R. Data Analysis in Community and Landscape Ecology; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Xu, L.; Liu, H.; Chu, X.; Su, K. Desert Vegetation Patterns at the Northern Foot of Tianshan Mountains: The Role of Soil Conditions. Flora Morphol. Distrib. Funct. Ecol. Plants 2006, 201, 44–50. [Google Scholar] [CrossRef]
- Rietkerk, M.; Ouedraogo, T.; Kumar, L.; Sanou, S.; Van Langevelde, F.; Kiema, A.; Van De Koppel, J.; Van Andel, J.; Hearne, J.; Skidmore, A.K.; et al. Fine-Scale Spatial Distribution of Plants and Resources on a Sandy Soil in the Sahel. Plant Soil 2002, 239, 69–77. [Google Scholar] [CrossRef]
- Walter, H. The Water Supply of Desert Plants. In The Water Relations of Plants; Rutter, A.J., Whitehead, F.H., Eds.; John Wiley & Sons: New York, NY, USA, 1963; pp. 199–205. [Google Scholar]
- Salama, F.; El-Ghani, M.A.; Gadallah, M.; EL NAGGAR, S.; Amro, A. Characteristics of Desert Vegetation along Four Transects in the Aridenvironment of Southern Egypt. Turk. J. Bot. 2016, 40, 59–73. [Google Scholar] [CrossRef]
- Salama, F.; El-Ghani, M.A.; Gadallah, M.; El-Naggar, S.; Amro, A. Variations in Vegetation Structure, Species Dominance and Plant Communities in South of the Eastern Desert-Egypt. Not. Sci. Biol. 2014, 6, 41–58. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.M. Floristics and Environmental Relations in Two Extreme Desertzones of Western Egypt. Glob. Ecol. Biogeogr. 2000, 9, 499–516. [Google Scholar] [CrossRef]
- El-Amier, Y.A.; El-Halawany, E.F.; Abdullah, T.J. Composition and Diversity of Plant Communities in Sand Formations along the Northern Coast of the Nile Delta in Egypt. Res. J. Pharm. Biol. Chem. Sci. 2014, 5, 826–847. [Google Scholar]
- Grime, J. Plant Strategies, Vegetation Processes, and Ecosystem Properties, 2nd ed.; John Wiley & Sons: New York, NY, USA, 2006. [Google Scholar]
- Alatar, A.; El-Sheikh, M.A.; Thomas, J. Vegetation Analysis of Wadi Al-Jufair, a Hyper-Arid Region in Najd, Saudi Arabia. Saudi J. Biol. Sci. 2012, 19, 357–368. [Google Scholar] [CrossRef]
- Salama, F.; Abd El-Ghani, M.; El-Tayeh, N. Vegetation and Soil Relationships in the Inland Wadi Ecosystem of Central Eastern Desert, Egypt. Turk. J. Bot. 2013, 37, 489–498. [Google Scholar] [CrossRef]
- Shaltout, K.H.; Al-Sodany, Y.M. Vegetation Analysis of Burullus Wetland: A RAMSAR Site in Egypt. Wetl. Ecol. Manag. 2008, 16, 421–439. [Google Scholar] [CrossRef]
- Springuel, I.; Sheded, M.; Darius, F.; Bornkamm, R. Vegetation Dynamics in an Extreme Desert Wadi under the Influence of Episodic Rainfall. Polish Bot. Stud. 2006, 22, 459–472. [Google Scholar]
- Danin, A. Plants of Desert Dunes; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- van Etten, E.J.B.; Fox, J.E.D. Vegetation—Environment Relationships of the Hamersley Ranges, a Mountainous Desert of North-West Australia. Folia Geobot. 2017, 52, 161–173. [Google Scholar] [CrossRef]
- Kassas, M.; El-Abyad, M.S. On the Phytosociology of the Desert Vegetation of Egypt. Ann. Arid Zone 1962, 1, 54–83. [Google Scholar]
- Kassas, M.; Imam, M. Habitat and Plant Communities in the Egyptian Desert: IV. The Gravel Desert. J. Ecol. 1959, 47, 289. [Google Scholar] [CrossRef]
- Sheded, M.G.; Hamed, S.T.; Badry, M.O. Vegetation Analysis of Six Riverian Islands in Hyper-Arid Environments at Qena Governorate (Upper Egypt). Acta Bot. Hung. 2014, 56, 409–431. [Google Scholar] [CrossRef]








| Edaphic Factors | I | II | III | IV | V | VI | VII | F Value | p |
|---|---|---|---|---|---|---|---|---|---|
| Organic matter % | 2.19 ± 0.62 | 2.93 ± 0.82 | 2.36 ± 1.08 | 2.25 ± 1.39 | 2.52 ± 2.06 | 1.59 ± 0.35 | 1.38 ± 0.0 | 0.434 | 0.858 |
| C | 1.27 ± 0.36 | 1.70 ± 0.47 | 1.37 ± 0.63 | 1.30 ± 0.81 | 1.46 ± 1.20 | 0.92 ± 0.20 | 0.80 ± 0.0 | 0.434 | 0.858 |
| N | 0.027 ± 0.004 | 0.032 ± 0.005 | 0.031 ± 0.0053 | 0.036 ± 0.015 | 0.037 ± 0.01 | 0.028 ± 0.0 | 0.035 ± 0.0 | 0.433 | 0.859 |
| CaCO3 (mg/g) | 30.62 ± 3.26 | 28.67 ± 1.29 | 23.00 ± 2.18 | 19.78 ± 6.48 | 17.75 ± 3.41 | 12.09 ± 1.53 | 12.50 ± 0.0 | 6.559 * | 0.006 |
| PH | 8.44 ± 0.06 | 8.60 ± 0.11 | 8.61 ± 0.043 | 8.49 ± 0.19 | 8.47 ± 0.23 | 8.53 ± 0.21 | 8.47 ± 0.0 | 0.344 | 0.913 |
| TDS (mg/L) | 218.09 ± 32.08 | 563.00 ± 425.68 | 762.33 ± 525.65 | 867.74 ± 677.17 | 616.36 ± 633.17 | 137.77 ± 35.35 | 258.67 ± 0.0 | 0.647 | 0.711 |
| EC (µs/cm) | 363 ± 53.56 | 937 ± 708.52 | 1263 ± 861.91 | 1435 ± 1113.94 | 1025.67 ± 1053.68 | 228.50 ± 57.27 | 432 ± 0.0 | 0.648 | 0.710 |
| Na+ (mg/g) | 1.51 ± 1.77 | 2.42 ± 1.17 | 1.70 ± 0.25 | 1.05 ± 1.12 | 2.24 ± 3.14 | 0.34 ± 0.20 | 0.36 ± 0.0 | 0.351 | 0.910 |
| K+ (mg/g) | 1.12 ± 0.29 | 1.56 ± 0.42 | 1.62 ± 0.51 | 1.66 ± 0.91 | 1.55 ± 0.68 | 1.35 ± 0.47 | 2.27 ± 0.0 | 0.458 | 0.842 |
| Ca2+ (mg/g) | 0.46 ± 0.17 | 0.74 ± 0.53 | 2.87 ± 3.13 | 3.71 ± 3.56 | 1.34 ± 1.18 | 0.22 ± 0.0 | 0.62 ± 0.0 | 0.853 | 0.573 |
| Mg2+ (mg/g) | 0.12 ± 0.03 | 0.18 ± 0.14 | 0.85 ± 1.21 | 0.28 ± 0.22 | 0.21 ± 0.13 | 0.13 ± 0.56 | 0.10 ± 0.0 | 0.955 | 0.513 |
| Cl− (mg/g) | 0.56 ± 0.12 | 1.40 ± 1.19 | 1.00 ± 0.07 | 0.91 ± 0.49 | 1.47 ± 1.87 | 0.38 ± 0.0 | 0.77 ± 0.0 | 0.341 | 0.915 |
| SO42− (mg/g) | 0.49 ± 0.05 | 0.67 ± 0.15 | 0.97 ± 0.35 | 0.91 ± 0.62 | 0.60 ± 0.26 | 0.34 ± 0.42 | 0.35 ± 0.0 | 0.966 | 0.507 |
| Clay | 1.60 ± 0.46 | 1.20 ± 0.28 | 1.53 ± 0.85 | 1.20 ± 0.43 | 1.37 ± 0.15 | 1 ± 0.0 | 1 ± 0.0 | 2.857 | 0.073 |
| Silt | 10.53 ± 7.62 | 26.30 ± 8.34 | 24.40 ± 30.50 | 11.10 ± 14.82 | 14.13 ± 18.60 | 4.25 ± 2.75 | 1.90 ± 0.0 | 2.792 | 0.077 |
| Sand | 87.87 ± 7.68 | 72.50 ± 8.06 | 74.07 ± 31.35 | 87.70 ± 15.25 | 84.50 ± 18.63 | 94.75 ± 2.75 | 97.10 ± 0.0 | 2.859 | 0.072 |
| AX1 | AX2 | OM | N | CaCO3 | pH | TDS | EC | Na | K | Ca | Mg | Cl | SO4 | Clay | Silt | Sand | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AX1 | −0.300 | −0.615 | 0.227 | −0.755 * | −0.091 | −0.499 | −0.497 | −0.451 | 0.644 | −0.522 | −0.303 | −0.074 | −0.666 | −0.668 | −0.494 | 0.504 | |
| AX2 | −0.300 | 0.052 | 0.544 | −0.263 | 0.100 | 0.716 | 0.713 | −0.169 | 0.259 | 0.848 * | 0.261 | 0.110 | 0.617 | −0.246 | 0.011 | −0.004 | |
| OM. | −0.615 | 0.052 | 0.138 | 0.733 | 0.423 | 0.617 | 0.619 | 0.952 ** | −0.383 | 0.285 | 0.277 | 0.770 * | 0.616 | 0.527 | 0.886 ** | −0.888 ** | |
| N | 0.227 | 0.544 | 0.138 | −0.354 | −0.154 | 0.590 | 0.592 | 0.160 | 0.652 | 0.461 | −0.006 | 0.646 | 0.285 | −0.254 | 0.001 | 0.005 | |
| CaCO3 | −0.755 * | −0.263 | 0.733 | −0.354 | 0.188 | 0.218 | 0.219 | 0.697 | −0.537 | 0.058 | 0.164 | 0.266 | 0.395 | 0.731 | 0.672 | −0.682 | |
| pH | −0.091 | 0.100 | 0.423 | −0.154 | 0.188 | 0.371 | 0.371 | 0.326 | 0.010 | 0.216 | 0.630 | 0.305 | 0.486 | −0.007 | 0.742 | −0.732 | |
| T.D.S. | −0.499 | 0.716 | 0.617 | 0.590 | 0.218 | 0.371 | 1.000 ** | 0.500 | 0.136 | 0.899 ** | 0.611 | 0.642 | 0.936 ** | 0.293 | 0.617 | −0.617 | |
| EC. | −0.497 | 0.713 | 0.619 | 0.592 | 0.219 | 0.371 | 1.000 ** | 0.503 | 0.137 | 0.897 ** | 0.610 | 0.646 | 0.935 ** | 0.293 | 0.619 | −0.618 | |
| Na | −0.451 | −0.169 | 0.952 ** | 0.160 | 0.697 | 0.326 | 0.500 | 0.503 | −0.330 | 0.130 | 0.254 | 0.824 * | 0.479 | 0.602 | 0.844 * | −0.848 * | |
| K | 0.644 | 0.259 | −0.383 | 0.652 | −0.537 | 0.010 | 0.136 | 0.137 | −0.330 | 1 | 0.148 | 0.021 | 0.189 | −0.047 | −0.539 | −0.230 | 0.240 |
| Ca | −0.522 | 0.848 * | 0.285 | 0.461 | 0.058 | 0.216 | 0.899 ** | 0.897 ** | 0.130 | 0.148 | 1 | 0.647 | 0.256 | 0.898 ** | 0.249 | 0.331 | −0.333 |
| Mg | −0.303 | 0.261 | 0.277 | −0.006 | 0.164 | 0.630 | 0.611 | 0.610 | 0.254 | 0.021 | 0.647 | 1 | 0.195 | 0.772 * | 0.488 | 0.599 | −0.603 |
| Cl | −0.074 | 0.110 | 0.770 * | 0.646 | 0.266 | 0.305 | 0.642 | 0.646 | 0.824 * | 0.189 | 0.256 | 0.195 | 1 | 0.454 | 0.190 | 0.674 | −0.670 |
| SO4 | −0.666 | 0.617 | 0.616 | 0.285 | 0.395 | 0.486 | 0.936 ** | 0.935 ** | 0.479 | −0.047 | 0.898 ** | 0.772 * | 0.454 | 1 | 0.475 | 0.705 | −0.708 |
| Clay | −0.668 | −0.246 | 0.527 | −0.254 | 0.731 | −0.007 | 0.293 | 0.293 | 0.602 | −0.539 | 0.249 | 0.488 | 0.190 | 0.475 | 1 | 0.513 | −0.531 |
| Silt | −0.494 | 0.011 | 0.886 ** | 0.001 | 0.672 | 0.742 | 0.617 | 0.619 | 0.844 * | −0.230 | 0.331 | 0.599 | 0.674 | 0.705 | 0.513 | −1.000 ** | |
| Sand | 0.504 | −0.004 | −0.888 ** | 0.005 | −0.682 | −0.732 | −0.617 | −0.618 | −0.848 * | 0.240 | −0.333 | −0.603 | −0.670 | −0.708 | −0.531 | −1.000 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sallam, H.; Alzain, M.N.; Abuzaid, A.O.; Loutfy, N.; Badry, M.O.; Osman, A.K.; Hammad, S.A. Wild Plant Diversity and Soil Characteristics of Desert Roadside Vegetation in the Eastern Desert. Diversity 2023, 15, 874. https://doi.org/10.3390/d15070874
Sallam H, Alzain MN, Abuzaid AO, Loutfy N, Badry MO, Osman AK, Hammad SA. Wild Plant Diversity and Soil Characteristics of Desert Roadside Vegetation in the Eastern Desert. Diversity. 2023; 15(7):874. https://doi.org/10.3390/d15070874
Chicago/Turabian StyleSallam, Heba, Mashail Nasser Alzain, Amani Omar Abuzaid, Naglaa Loutfy, Mohamed O. Badry, Ahmed K. Osman, and Sabah A. Hammad. 2023. "Wild Plant Diversity and Soil Characteristics of Desert Roadside Vegetation in the Eastern Desert" Diversity 15, no. 7: 874. https://doi.org/10.3390/d15070874
APA StyleSallam, H., Alzain, M. N., Abuzaid, A. O., Loutfy, N., Badry, M. O., Osman, A. K., & Hammad, S. A. (2023). Wild Plant Diversity and Soil Characteristics of Desert Roadside Vegetation in the Eastern Desert. Diversity, 15(7), 874. https://doi.org/10.3390/d15070874