

Impacts of Diffuse Land-Use on Plant Diversity Patterns in the Miombo Woodlands of Western Zambia


Abstract
1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Study Design and Data Collection

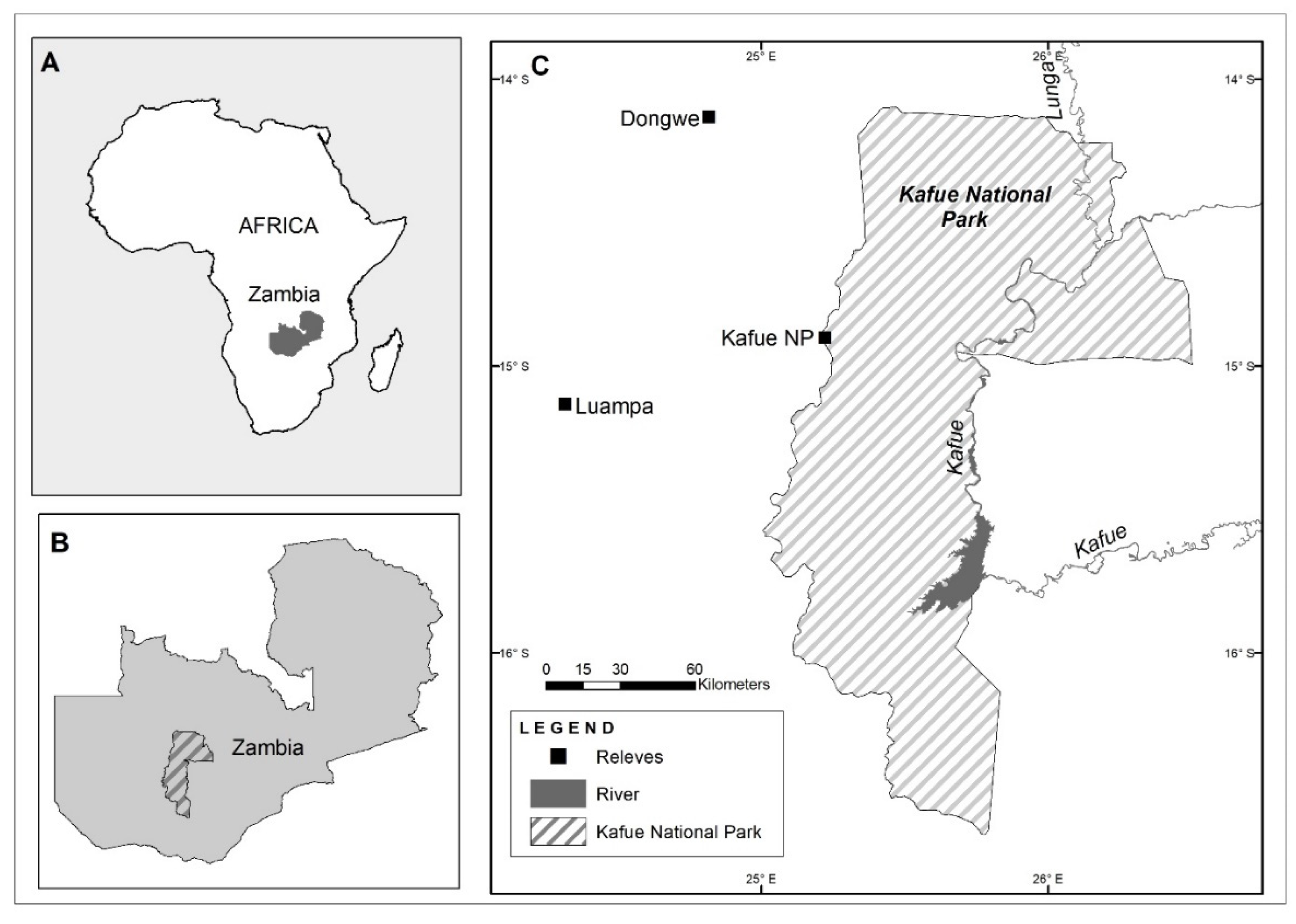{kind=link}
{kind=link}
| Site Name | Mean Annual Rainfall [mm/yr] | Mean Annual Temperature [°C] | Land-Use Type | Land-Use Intensity | GPS Coordinates (North West Corner) | Elevation Min–Max [masl] |
|---|---|---|---|---|---|---|
| Luampa | 875.5 | 22.22 | Forest Reserve | High | S15.13782, E24.48778 | 1152–1158 |
| Dongwe | 990.6 | 22.34 | Community Forest | Medium | S14.09577, E24.01520 | 1066–1145 |
| Kafue NP | 897.4 | 21.98 | National Park | Low | S14.89830, E25.43676 | 1091–1210 |
2.3. Data Analysis
3. Results
3.1. Indicators for Land-Use Intensities at the Three Sites
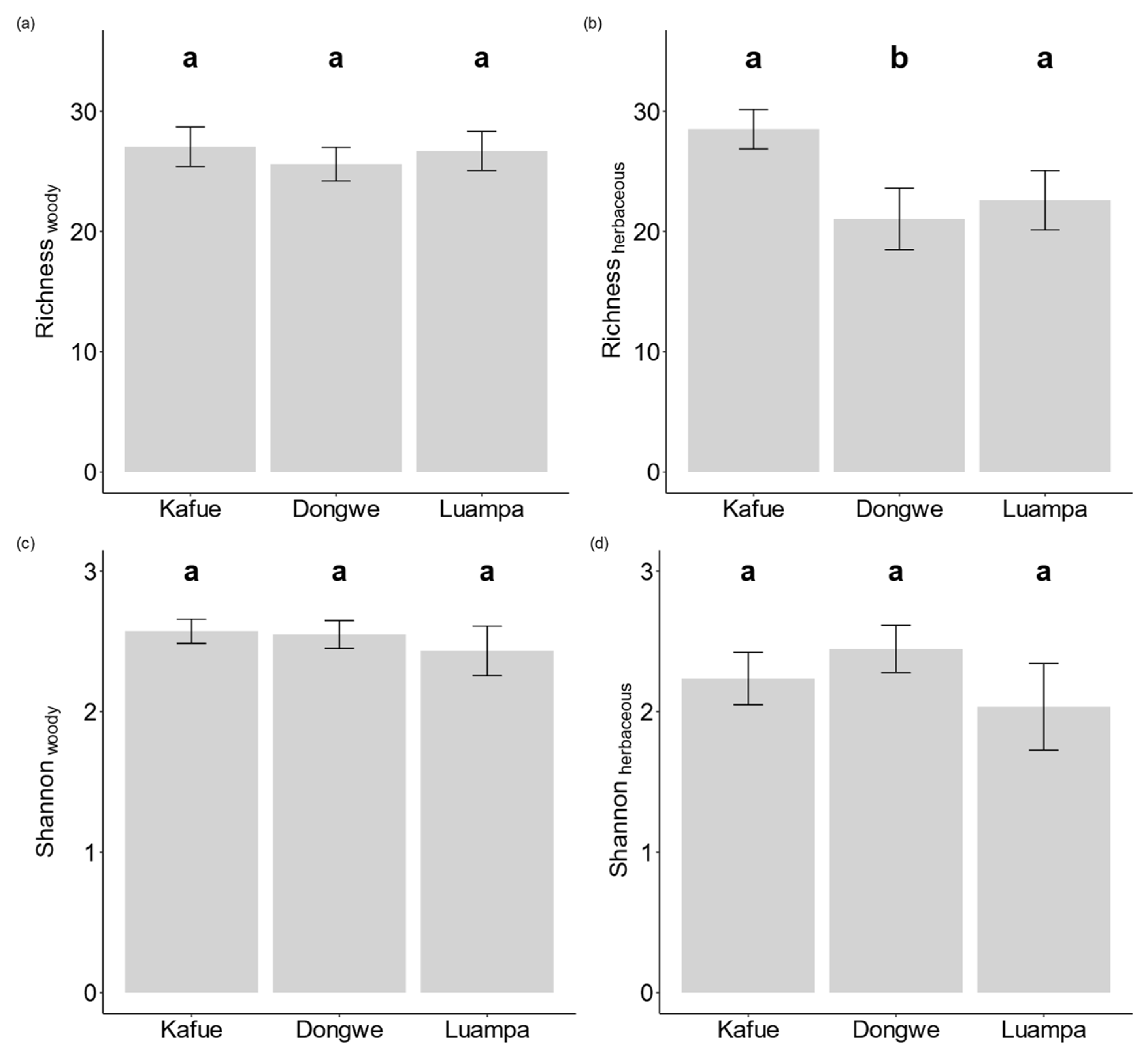
3.2. Diversity Patterns at the Different Land-Use Intensities
3.3. Environmental Drivers of Diversity at Plot Level
4. Discussion
4.1. Effect of Land Use on Plant Species Diversity
4.2. Drivers of Diversity at Plot Level
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Species Name | Life Form | Abundance | Uses | Voucher Number | Location |
|---|---|---|---|---|---|
| Acanthaceae Duosperma quadrangulare (Klotzsch) Brummitt Hypoestes forskaolii (Vahl) R.Br. | |||||
| Herbaceous | rare | FO | 138076 | K | |
| Herbaceous | TX | 132148 | K | ||
| Amaryllidaceae Crinum macowanii Baker | rare | ||||
| Herbaceous | rare | NW | 132141 | L, K | |
| Anacardiaceae | |||||
| Searsia quartiniana (A. Rich.) A.J. Mill. | Woody | Frequent | FO | 131071, 142196 | L, D |
| Sclerocarya birrea (A.Rich.) Hochst. | Woody | rare | FO | 132128 | K |
| Annonaceae | |||||
| Friesodielsia obovata (Benth.) Verdc. | Woody | occasional | FO, FD | 142610 | K |
| Uvariastrum hexaloboides (R.E.Fr.) | Woody | occasional | FO | 132107 | L, K |
| Xylopia odoratissima Welw. ex Oiv. | Woody | common | NW | 131181 | L, D |
| Apocynaceae | |||||
| Diplorhynchus condylocarpon (Müll. Arg.) Pichon | Woody | common | NW | 140121 | D, L, K |
| Landolphia parvifolia K. Schum. | Woody | frequent | FO | 142505 | D, L |
| Strophanthus welwitschii (Baill.) K. Schum. | Woody | occasional | 142649 | D | |
| Asparagaceae | |||||
| Asparagus racemosus Willd. | Herbaceous | occasional | 142788 | K | |
| Asphodelaceae | |||||
| Bulbine abyssinica A.Rich. | Herbaceous | rare | NW | 131355, 132214 | D, K |
| Asteraceae | |||||
| Conyza gouanii (L.) Willd. | Herbaceous | occasional | NW | 132207 | K |
| Crassocephalum rubens (Jacq.) S. Moore | Herbaceous | rare | NW | 131301 | D |
| Dicoma anomala Sond. | Herbaceous | occasional | 138099 | K | |
| Elephantopus scaber L. | Herbaceous | frequent | 131119 | D, L, K | |
| Erythrocephalum zambesianum Oliv. & Hiern | Herbaceous | frequent | NW | 142503 | L, K |
| Felicia welwitschii (Hiern) Grau | Herbaceous | rare | 142698 | D | |
| Macledium poggei (O.Hoffm.) S.Ortiz | Herbaceous | rare | 142660 | K | |
| Pleiotaxis eximia O. Hoffm. Vernonia glabra ssp. laxa (Seetz) Vatke | Herbaceous | occasional | NW | 142665 | L, K L, K |
| Vernonia melleri Oliv. & Hiern | Herbaceous | occasional | 131401 | L | |
| Vernonia petersii Oliv. & Hiern ex Oliv. | Herbaceous | frequent | FD | 142678 | D, K |
| Vernonia poskeana Vatke & Hildebr. | Herbaceous | rare | 131157 | L | |
| Capparaceae | |||||
| Capparis tomentosa Lam. | Herbaceous | frequent | NW, FO, AE | 138093 | K |
| Cleome hirta (Klotzsch) Oliv. | Herbaceous | frequent | 138076 | K | |
| Maerua triphylla ssp. pubescens A. Rich. (Klotzsch) DeWolf | Herbaceous | frequent | 131358 | K | |
| Chrysobalanaceae | |||||
| Parinari capensis Harv. | Woody | frequent | NW, FO | L | |
| Parinari curatellifolia Planch. ex Benth. | Woody | frequent | NW, FO, WO | 131213 | L, K |
| Combretaceae | |||||
| Combretum collinum Fresen. | Woody | occasional | NW, TI | 131113 | D, L, K |
| Combretum elaeagnoides Klotzsch | Woody | occasional | 131403 | D, L | |
| Combretum molle R.Br. ex G.Don | Woody | frequent | NW, TI | 142578 | D, L, K |
| Combretum psidioides Welw. | Woody | occasional | NW | 131402 | D |
| Combretum zeyheri Sond. | Woody | common | NW, WO | 131183 | L, K |
| Pteleopsis anisoptera (Welw. ex M.A.Lawson) Engl. & Diels | Woody | occasional | NW, TI, WO | 1320102 | D |
| Terminalia brachystemma Welw. ex Hiern | Woody | occasional | NW, FO, WO | 131165 | L, K |
| Commelinaceae | |||||
| Cyanotis longifolia Benth. | Herbaceous | frequent | 131016 | D, L, K | |
| Dipterocarpaceae | |||||
| Marquesia macroura Gilg | Woody | occasional | 140121 | K | |
| Gilg | Woody | occasional | 132222 | K | |
| Monotes glaber Sprague | Woody | occasional | NW, WO | 142639, 141189 | L |
| Ebenaceae | |||||
| Diospyros batocana Hiern | Woody | common | NW, FO, PU | 140159 | D, L, K |
| Diospyros mespiliformis Hochst. ex A.DC. | Woody | frequent | NW, FO | K | |
| Diospyros virgata (Gürke) Brenan | Woody | common | NW | 131054, 131191, 140143 | D, L, K |
| Ericaceae | |||||
| Cleistanthus polystachyus Hook. f. ex Planch. | Herbaceous | rare | 132122 | L | |
| Erythroxylaceae | |||||
| Erythroxylum emarginatum Thonn. | Woody | occasional | 142649 | K | |
| Euphorbiaceae | |||||
| Acalypha ornata Hochst. ex A. Rich. | Herbaceous | occasional | 132101 | L | |
| Flueggea virosa (Roxb. ex Willd.) Voigt | Woody | occasional | NW, FO | 142566 | L, K |
| Hymenocardia acida Tul. | Woody | common | NW | 142526 | D, L, K |
| Maprounea africana Müll.Arg. | Woody | frequent | NW | L | |
| Oldfieldia dactylophylla (Welw. ex Oliv.) Léonard | Woody | frequent | 142693 | D | |
| Pseudolachnostylis maprouneifolia Pax | Woody | common | NW, FO | 140131, 131047 | D, L, K |
| Sclerocroton oblongifolius (Müll. Arg.) Kruijt & Roebers | Herbaceous | rare | NW | 131056, 142172, 135679 | L, K |
| Uapaca kirkiana Müll. Arg. | Woody | occasional | NW, FO | 142817 | D, K |
| Uapaca nitida ssp. nitida | Woody | occasional | NW, FO | L, K | |
| Fabaceae | |||||
| Afzelia quanzensis Welw. | Woody | frequent | NW, FO TI, WO | 131163 | L, K |
| Albizia antunesiana Harms | Woody | frequent | NW, WO | 142766 | D, L, K |
| Albizia versicolor Welw. ex Oliv. | Woody | common | NW, WO | 132156 | D, K |
| Anisophyllea boehmii Engl. | Woody | occasional | NW, FO | 132216 | K |
| Baphia massaiensis var. obovata Taub. | Woody | common | NW, FO, FD | 131007 | D, L, K |
| Bauhinia petersiana Bolle | Woody | frequent | NW, FO, FD | 142520 | D, L, K |
| Bobgunnia madagascariensis (Desv.) J.H.Kirkbr. & Wiersema | Woody | common | NW, FO, TI | 131191 | L, K |
| Brachystegia boehmii Taub. | Woody | common | TI, WO, PO, NW | D, L, K | |
| Brachystegia spiciformis Benth. | Woody | common | TI, WO, PO, NW | 131146 | D, L, K |
| Burkea africana Hook. | Woody | common | WO, PO, NW | D, L, K | |
| Cassia abbreviata Oliv. | Woody | occasional | NW | 142622 | K |
| Chamaecrista mimosoides (L.) Greene | Herbaceous | occasional | NW | 131025, 142559 | D, K |
| Copaifera baumiana Harms | Woody | common | NW | 131085 | L, K |
| Crotalaria alexandri Baker f. | Herbaceous | occasional | 142576 | L, D | |
| Crotalaria anisophylla (Hiern) Welw. ex Baker f. | Herbaceous | occasional | 142774 | L | |
| Crotalaria caudata Welw. ex Baker | Herbaceous | occasional | 131176 | D, L | |
| Crotalaria cephalotes Steud. ex A.Rich. | Herbaceous | occasional | NW | 132180 | K |
| Crotalaria laburnifolia L. | Herbaceous | common | 142570 | D | |
| Crotalaria microcarpa Hochst. ex Benth. | Herbaceous | rare | 131099 | D, L | |
| Cryptosepalum exfoliatum ssp. pseudotaxus De Wild. | Woody | common | TI, NW, PO, | 142504 | D |
| Dichrostachys cinerea (L.) Wight & Arn. | Woody | common | NW, WO | D, L, K | |
| Erythrophleum africanum | Woody | common | WO, PO, NW | 131124 | D, L, K |
| Guibourtia coleosperma (Benth.) J.Leonard | Woody | common | TI, NW, | 140151 | D, L |
| Indigofera demissa Taub. | Herbaceous | occasional | 131174 | D, L | |
| Indigofera flavicans Baker | Herbaceous | common | 131130 | L, K | |
| Isoberlinia angolensis (Benth.) Hoyle & Brenan | Woody | occasional | TI, NW, AE | K | |
| Julbernardia paniculata (Benth.) Troupin | Woody | common | TI, WO, PO, NW, FD | 140155 | D, L, K |
| Keetia venosa (Oliv.) Bridson | Woody | occasional | 142561 | K | |
| Lannea edulis (Sond.) Engl. | Woody | frequent | NW | 131178 | L, K |
| Mucuna poggei Taub. | Woody | occasional | NW | 142564 | K |
| Pericopsis angolensis (Baker) Meeuwen | Woody | occasional | TI, WO, NW, FD | 138072 | L, K |
| Piliostigma thonningii (Schumach.) Milne-Redh. | Woody | occasional | K | ||
| Pterocarpus angolensis DC. | Woody | common | TI, WO, NW | D, L, K | |
| Rhynchosia caribaea (Jacq.) DC. | Woody | occasional | NW | 131195 | L, K |
| Flacourtiaceae | |||||
| Flacourtia indica (Burm.f.) Merr. | Woody | frequent | NW, FO | 131198 | K |
| Hypericaceae | |||||
| Psorospermum baumii Engl. | Woody | occasional | NW, FO | 131162 | D |
| Lamiaceae | |||||
| Ocimum africanum Lour. | Herbaceous | rare | K | ||
| Tinnea vestita Baker | Herbaceous | occasional | NW, FO | 131021, 142739 | L, K |
| Vitex doniana Sweet | Woody | occasional | TI, FO, NW, FD | 140154 | L |
| Vitex madiensis Oliv. subsp. milanjiensis (Britten) F. White | Woody | rare | 142638 | L | |
| Lauraceae | |||||
| Cassytha pondoensis ssp. Pondoensis Engl. | Woody | occasional | NW | 131134 | D, L |
| Malvaceae | |||||
| Abutilon angulatum (Guill. & Perr.) Mast. | Woody | occasional | NW | 142504 | D |
| Pavonia senegalensis (Cav.) Leistner | Woody | rare | 131268 | D, L | |
| Meliaceae | |||||
| Bersama abyssinica Fresen. | Woody | rare | NW, AE | 142874 | K |
| Trichilia emetica Vahl | Woody | rare | NW, AE | K | |
| Myrtaceae | |||||
| Syzygium guineense (Willd.) DC. | Woody | occasional | NW, FO | 142200 | L |
| Ochna pulchra Hook. | Woody | common | NW | 131059 | D, L, K |
| Olacaceae | |||||
| Olax obtusifolia De Wild. | Woody | occasional | 142598 | L | |
| Ximenia americana L. | Woody | frequent | NW, FO | 142523 | L, K |
| Ximenia caffra Sond. | Woody | frequent | NW, FO | 131047 | K |
| Oleaceae | |||||
| Olea capensis L. | Woody | occasional | 138065 | K | |
| Schrebera trichoclada Welw. | Woody | rare | 144524 | L, K | |
| Orobanchaceae | |||||
| Striga asiatica (L.) Kuntze | Herbaceous | occasional | NW | 138077 | L, D |
| Oxalidaceae | |||||
| Biophytum abyssinicum Steud. Ex A. Rich. | Herbaceous | occasional | NW | D | |
| Biophytum umbraculum Welw. | Herbaceous | occasional | 131098 | D, L | |
| Passifloraceae | |||||
| Paropsia brazzeana Baill. | Woody | frequent | NW | 131080 | D, L, K |
| Polygalaceae | |||||
| Securidaca longepedunculata Fresen. | Woody | occasional | NW | 131204 | L |
| Proteaceae | |||||
| Protea angolensis Welw. | Woody | rare | NW | 142795 | K |
| Protea gaguedi J.F. Gmel. | Woody | frequent | NW | K | |
| Ranunculaceae | |||||
| Clematis chrysocarpa Welw. ex Oliv. | Herbaceous | occasional | 131351, 142755, 142609 | L, K | |
| Rhamnaceae | |||||
| Ziziphus mucronata Willd. | Woody | occasional | FO, NW | K | |
| Rubiaceae | |||||
| Agathisanthemum bojeri Klotzsch | Woody | occasional | NW | 142781 | L, K |
| Fadogia cienkowskii Schweinf. | Woody | occasional | 142191 | L | |
| Gardenia ternifolia Schumach. & Thonn. | Woody | occasional | NW, FO | 132132 | D |
| Pavetta schumanniana F. Hoffm ex K. Schum | Woody | frequent | 142749 | D, K | |
| Rothmannia engleriana (K.Schum.) Keay | Woody | frequent | NW | 131196 | D, L |
| Spermacoce pusilla Wall. | Woody | occasional | 131100 | D, L | |
| Tricalysia longituba De Wild. | Woody | occasional | 131255 | L, K | |
| Vangueriopsis lanciflora (Hiern) Robyns | Woody | frequent | NW, WO | 131180 | D, L |
| Sapindaceae | |||||
| Zanha africana (Radlk.) Exell | Woody | occasional | WO | 142751 | K |
| Sapotaceae | |||||
| Englerophytum magalismontanum (Sond.) T.D.Penn. | Herbaceous | occasional | NW | 131079 | D, L |
| Solanaceae | |||||
| Solanum mauritianum Scop. | Herbaceous | occasional | K | ||
| Strychnaceae | |||||
| Strychnos cocculoides Baker | Woody | frequent | NW, FO, AE, PO | 131083 | D, L, K |
| Strychnos pungens Soler. | Woody | frequent | NW, FO | 140149 | D, L, K |
| Thelypteridaceae | |||||
| Christella chaseana (Schelpe) Holttum | Herbaceous | occasional | D | ||
| Tiliaceae | |||||
| Grewia flavescens Juss. | Herbaceous | occasional | NW, FO | 131013 | D, L, K |
| Triumfetta annua L. | Herbaceous | occasional | 131109 | L, K | |
| Verbenaceae | |||||
| Endostemon obtusifolius (E. Mey. ex Benth.) N.E.Br. | Herbaceous | occasional | D, L | ||
| Lantana angolensis Moldenke | Herbaceous | occasional | NW, FO, FD | 142644 | L, K |
| Vitaceae | |||||
| Cyphostemma junceum Wild & R.B. Drumm. | Herbaceous | frequent | NW, FO | 132169 | D |
| Cyphostemma princeae Wild & R.B. Drumm | Herbaceous | frequent | NW, FO | 131382 | D, L, K |
| Zingiberaceae | occasional | ||||
| Aframomum alboviolaceum (Ridl.) K.Schum. | Herbaceous | NW, FO | D |
| Analysis Variable | Method and Reference | Unit |
|---|---|---|
| pH | using the CaCl2 mixture in H2O [75] | |
| Nitrogen (N) | Kjeldahl method [76] | percentage per total weight |
| Phosphorus (P) | Bray I extractant [77] | parts per million |
| Organic Carbon (Org C) | Walkley Black technique [78] | percentage per total weight |
| Calcium (Ca) | Ammonia acetate extraction [79] | parts per million |
| Magnesium (Mg) | Ammonia acetate extraction [79] | parts per million |
| Sodium (Na) | Ammonia acetate extraction [79] | parts per million |
| Potassium (K) | Ammonia acetate extraction [79] | parts per million |
| Zinc (Zn) | DPTA method [80] | parts per million |
| Manganese (Mn) | DPTA method [80] | parts per million |
| Iron (Fe) | DPTA method [80] | parts per million |
| Cation Electronic Exchange (CEC) | Conductivity method [81] | Milli-equivalents |
| Variables | Mean and Covariance | Luampa | Dongwe | Kafue National Park |
|---|---|---|---|---|
| pH | 4.4 | 3.9 | 4.9 | |
| CV | 0.04 | 0.07 | 0.08 | |
| N | 0.016 | 0.011 | 0.018 | |
| CV | 0.5 | 0.64 | 0.66 | |
| P | 5.0381 | 3.01 | 7.73 | |
| CV | 0.42 | 0.39 | 0.83 | |
| Org C | 0.25 | 0.21 | 0.26 | |
| CV | 0.43 | 0.37 | 0.5 | |
| Ca | 40 | 32 | 120 | |
| CV | 0.71 | 1.61 | 0.49 | |
| Mg | 10.23 | 11 | 24.31 | |
| CV | 0.58 | 0.14 | 0.57 | |
| Na | 2.23 | 3.75 | 6.53 | |
| CV | 1.49 | 0.64 | 0.65 | |
| K | 10.91 | 10.35 | 42.05 | |
| CV | 1.28 | 1.14 | 0.48 | |
| Zn | 0.27 | 0.05 | 0.15 | |
| CV | 2.19 | 1.43 | 1.4 | |
| Mn | 14.42 | 12.1 | 82.05 | |
| CV | 1.15 | 2.6 | 0.58 | |
| Fe | 11.23 | 11.95 | 28.47 | |
| CV | 0.49 | 0.84 | 0.51 | |
| CEC | 3.45 | 2.4 | 3.83 | |
| CV | 0.35 | 0.58 | 0.32 |
References
- Campbell, B.M.; Angelsen, A.; Cunningham, A.B.; Katerere, Y.; Sitoe, A.; Wunder, S. Miombo Woodlands—Opportunities and Barriers to Sustainable Forest Management; Centre for International Forest Research: Bogor, Indonesia, 2008. [Google Scholar]
- Frost, P. The ecology of miombo woodlands. In The Miombo in Transition: Woodlands and Welfare in Africa; Campbell, B., Ed.; Centre for International Forestry Research: Bogor, Indonesia, 1996; pp. 11–57. [Google Scholar]
- Kalaba, F.K.; Quinn, C.H.; Dougill, A.J.; Vinya, R. Floristic composition, species diversity and carbon storage in charcoal and agriculture fallows and management implications in Miombo woodlands of Zambia. For. Ecol. Manag. 2013, 304, 99–109. [Google Scholar] [CrossRef]
- Chidumayo, E.N.; Gumbo, D.J. The Dry Forests and Woodlands of Africa: Managing for Products and Services; Chidumayo, E.N., Gumbo, D.J., Eds.; Centre for International Forest Research: London, UK, 2010. [Google Scholar]
- Chapin, F.S., 3rd; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Wessels, K.J.; Reyers, B.; Van Jaarsveld, A.S. Incorporating land cover information into regional biodiversity assessments in South Africa. Anim. Conserv. 2000, 3, 67–79. [Google Scholar] [CrossRef]
- Walters, D.J.J.; Kotze, D.C.; O’connor, T.G. Impact of land use on vegetation composition, diversity, and selected soil properties of wetlands in the southern Drakensberg mountains, South Africa. Wetl. Ecol. Manag. 2006, 14, 329–348. [Google Scholar] [CrossRef]
- Vinya, R.; Syampungani, S.; Kasumu, C.E.; Monde, C.; Kasubika, R. Preliminary Study on the Drivers of Deforestation and Potential for REDD+ in Zambia. A Consultancy Report Prepared for Forestry Department and FAO under the National UN-REDD+ Programme; Ministry of Lands & Natural Resources: Lusaka, Zambia, 2011. [Google Scholar]
- Hawthorne, W.D.; Marshall, C.A.M.; Abu-Juam, M.; Agyeman, V.K. The Impact of Logging Damage on Tropical Rain-Forests, Their Recovery and Regeneration: An Annotated Bibliography; Oxford Forestry Institute: Oxford, UK, 2011; 123p. [Google Scholar]
- Newbold, T.; Tittensor, D.P.; Harfoot, M.B.J.; Scharlemann, J.P.W.; Purves, D.W. Nonlinear changes in modelled terrestrial ecosystems subjected to perturbations. Sci. Rep. 2020, 10, a14051. [Google Scholar] [CrossRef]
- White, P.S.; Jentsch, A. The Search for Generality in Studies of Disturbance and Ecosystem Dynamics. Botany 2001, 62, 399–450. [Google Scholar] [CrossRef]
- Revermann, R.; Wallenfang, J.; Oldeland, J.; Finckh, M. Species richness and evenness respond to diverging land-use patterns—A cross-border study of dry tropical woodlands in southern Africa. Afr. J. Ecol. 2016, 55, 152–161. [Google Scholar] [CrossRef]
- Barlow, J.; Lennox, G.D.; Ferreira, J.; Berenguer, E.; Lees, A.C.; Mac Nally, R.; Thomson, J.R.; Ferraz, S.F.D.B.; Louzada, J.; Oliveira, V.H.F.; et al. Anthropogenic disturbance in tropical forests can double biodiversity loss from deforestation. Nature 2016, 535, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Marcial, N.; González-Espinosa, M.; Williams-Linera, G. Anthropogenic disturbance and tree diversity in Montane Rain Forests in Chiapas, Mexico. For. Ecol. Manag. 2001, 154, 311–326. [Google Scholar] [CrossRef]
- Vallan, D.; Andreone, F.; Raherisoa, V.H.; Dolch, R. Does selective wood exploitation affect amphibian diversity? The case of An’Ala, a tropical rainforest in eastern Madagascar. Oryx 2004, 38, 410–417. [Google Scholar] [CrossRef]
- Savadogo, P.; Tiveau, D.; Sawadogo, L.; Tigabu, M. Herbaceous species responses to long-term effects of prescribed fire, grazing and selective tree cutting in the savanna-woodlands of West Africa. Perspect. Plant Ecol. Evol. Syst. 2008, 10, 179–195. [Google Scholar] [CrossRef]
- Kalema, V.N.; Witkowski, E.T. Land-use impacts on woody plant density and diversity in an African savanna charcoal production region. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2012, 8, 231–247. [Google Scholar] [CrossRef]
- Chidumayo, E.N. Species structure in Zambian miombo woodland. J. Trop. Ecol. 1987, 3, 109–118. [Google Scholar] [CrossRef]
- Chidumayo, E.N. Changes in miombo woodland structure under different land tenure and use systems in central Zambia. J. Biogeogr. 2002, 29, 1619–1626. [Google Scholar] [CrossRef]
- Zisadza-Gandiwa, P.; Mabika, C.T.; Kupika, O.L.; Gandiwa, E.; Murungweni, C. Vegetation Structure and Composition across Different Land Uses in a Semiarid Savanna of Southern Zimbabwe. Int. J. Biodivers. 2013, 2013, 692564. [Google Scholar] [CrossRef]
- Dewees, P.A.; Campbell, B.M.; Katerere, Y.; Sitoe, A.; Cunningham, A.B.; Angelsen, A.; Wunder, S. Managing the Miombo woodlands of Southern Africa: Policies, incentives and options for the rural poor. J. Nat. Resour. Policy Res. 2010, 2, 57–73. [Google Scholar] [CrossRef]
- Trapnell, C.G. Ecological Results of Woodland and Burning Experiments in Northern Rhodisia. J. Ecol. 1959, 47, 129. [Google Scholar] [CrossRef]
- Werger, M.J.A.; Coetzee, B.J. The Sudano-Zambezian region. In Biogeography and Ecology of Southern Africa; Monographiae Biologicae; Springer: Dordrecht, The Netherlands, 1978. [Google Scholar] [CrossRef]
- Phiri, P. A Checklist of Zambian Vascular Plants; Southern African Botanical Diversity Network: Pretoria, South Africa, 2005. [Google Scholar]
- Desanker, P.V.; Frost, P.G.H.; Frost, C.O.; Justice, C.O.; Scholes, R.J. (Eds.) The Miombo Network: Framework for a Terrestrial Transect Study of Land-Use and Land-Cover Change in the Miombo Ecosystems of Central Africa; IGBP Report 41; The International Geosphere-Biosphere Programme (IGBP): Stockholm, Sweden, 1997; 109p. [Google Scholar]
- Japan Association for International Collaboration of Agriculture and Forestry. Agriculture and Forestry in Zambia: Present Situation and Issues for Future Development; Japan Association for International Collaboration of Agriculture and Forestry: Tokyo, Japan, 2008. [Google Scholar]
- Survey Department. Soil map of Zambia; Survey Department: Lusaka, Zambia, 1983. [Google Scholar]
- White, F. The savannah woodlands of the Zambesian and Sudanian domain: An ecological and phytogeographical comparison. Syst. Assn. Publ. 1965, 4, 71–103. [Google Scholar]
- White, F. The Vegetation of Africa: A Descriptive; United Nations Educational Scientific and Cultural Organisation: Paris, France, 1983. [Google Scholar]
- Hofiço, N.D.S.A.; Fleig, F.D. Diversity and Structure of Miombo Woodlands in Mozambique Using a Range of Sampling Sizes. J. Agric. Sci. Technol. B 2015, 5, 679–690. [Google Scholar] [CrossRef]
- Furley, P.A.; Rees, R.M.; Ryan, C.M.; Saiz, G. Savanna burning and the assessment of long-term fire experiments with particular reference to Zimbabwe. Prog. Phys. Geogr. Earth Environ. 2008, 32, 611–634. [Google Scholar] [CrossRef]
- Chidumayo, E.N. Miombo Ecology and Management: An Introduction; IT Publications in association with the Stockholm Environment Institute: London, UK, 1997; 166p. [Google Scholar]
- Environmental Council of Zambia. State of Environment Report in Zambia 2000; Environmental Council of Zambia: Lusaka, Zambia, 2001. [Google Scholar]
- Google Earth 6.0–Version. Western Province, Zambia. 14040′51.22″ S, 24043′14.54″ E Eye Alt 225.46 mi. Digital Globe 2012. 2014. Available online: http://www.earth.google.com (accessed on 17 April 2014).
- GLDAS. Global Land Data Assimilation System. GLDAS NOAA R0.25 deg 3H 1948–2010. 2014. Available online: https://developers.google.com/earth-engine/datasets/catalog/NASA_GLDAS_V021_NOAH_G025_T3H (accessed on 12 May 2015).
- Ministry of Tourism Environment and Natural Resources. Current Protected Area Categories and the Institutions Responsible for Their Management in Zambia; Ministry of Tourism Environment and Natural Resources: Lusaka, Zambia, 2004. [Google Scholar]
- Petruzzello, M. Herbaceous Plant. Encyclopedia Britannica, 17 January 2023. 2023. Available online: https://www.britannica.com/plant/herbaceous-plant (accessed on 19 April 2023).
- Vernon, R. Field Guide to Important Arable Weeds of Zambia; Department of Agriculture: Chilanga, Zambia, 1983; 45p. [Google Scholar]
- Bingham, M.; Willemen, A.; Wursten, B.; Ballings, P.; Hyde, M. Flora of Zambia. Lists of Families. 2011. Available online: http://www.zambiaflora.com/speciesdata/listoffamilies (accessed on 16 March 2016).
- Blaser, W.J.; Shanungu, G.K.; Edwards, P.J.; Venterink, H.O. Woody encroachment reduces nutrient limitation and promotes soil carbon sequestration. Ecol. Evol. 2014, 4, 1423–1438. [Google Scholar] [CrossRef] [PubMed]
- Siachoono, S.M.; Syampungani, S.; Mundike, J. Invasive Alien Species in Zambia. Invasive Alien Species 2021, 1, 313–329. [Google Scholar] [CrossRef]
- Nieman, W.A.; Van Wilgen, B.W.; Leslie, A.J. A review of fire management practices in African savanna-protected areas. Koedoe 2021, 63, 13. [Google Scholar] [CrossRef]
- Storrs, A. Know Your Trees—Some of the Common Trees Found in Zambia; Forest Department: Kitwe, Zambia, 1979. [Google Scholar]
- Klassen, E.S.; Craven, P. Checklist of grasses in Namibia. Southern African Botanical Diversity Network Report No. 20; SABONET: Windhoek, Namibia, 2003. [Google Scholar]
- Le Roux, P.; Mueller, M. Le Roux and Müller’s Field Guide to the Trees and Shrubs of Namibia; Macmillan Education: Windhoek, Namibia, 2009. [Google Scholar]
- van Wyk, B.; van Wyk, P. Field Guide to Trees of Southern Africa: An African Perspective; Struik Publishers: Cape Town, South Africa, 2013. [Google Scholar]
- JSTOR. Global Plants Database. Available online: https://plants.jstor.org/ (accessed on 16 March 2016).
- The Plant List. 2013. Available online: http://www.theplantlist.org/ (accessed on 23 April 2016).
- Scheiner, S.M. Biological Diversity: Frontiers in Measurement and Assessment. Edited by Anne E. Magurran and Brian J. McGill. Oxford and New York: Oxford University Press. $135.00 (hardcover); $72.50 (paper). xvii + 345 p.; ill.; index. ISBN: 978-0-19-958066-8 (hc); 978-0-19-958067-5 (pb). 2011. Q. Rev. Biol. 2012, 87, 254. [Google Scholar] [CrossRef]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Calcagno, V.; Mazancourt, C. de glmulti: An R Package for Easy Automated Model Selection with (Generalized) Linear Models. J. Stat. Softw. 2010, 34, 1–29. [Google Scholar] [CrossRef]
- Long, S.J. Regression Models for Categorical and Limited Dependent Variables; Sage Publications: Thousand Oaks, CA, USA, 1997. [Google Scholar]
- Jackman, S.; Tahk, A.; Zeileis, A.; Maimone, C.; Fearon, J.; Meers, Z. Package pscl: Classes and Methods for R Developed in the Political Science Computational Laboratory; CRAN: Stanford, CA, USA, 2017; pp. 1–97. [Google Scholar]
- Ng’andwe, P.; Mwitwa, J.; Muimba-Kankolongo, A. Forest Policy, Economics, and Markets in Zambia; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1–166. [Google Scholar]
- De Cauwer, V.; Muys, B.; Revermann, R.; Trabucco, A. Potential, realised, future distribution and environmental suitability for Pterocarpus angolensis DC in southern Africa. For. Ecol. Manag. 2014, 315, 211–226. [Google Scholar] [CrossRef]
- Jew, E.K.; Dougill, A.J.; Sallu, S.M.; O’connell, J.; Benton, T.G. Miombo woodland under threat: Consequences for tree diversity and carbon storage. For. Ecol. Manag. 2016, 361, 144–153. [Google Scholar] [CrossRef]
- Nacoulma, B.M.I.; Schumann, K.; Traoré, S.; Bernhardt-Römermann, M.; Hahn, K.; Wittig, R.; Thiombiano, A. Impacts of land-use on West African savanna vegetation: A comparison between protected and communal area in Burkina Faso. Biodivers. Conserv. 2011, 20, 3341–3362. [Google Scholar] [CrossRef]
- Smet, M.; Ward, D. A comparison of the effects of different rangeland management systems on plant species composition, diversity and vegetation structure in a semi-arid savanna. Afr. J. Range Forage Sci. 2005, 22, 59–71. [Google Scholar] [CrossRef]
- Giliba, R.A.; Boon, E.K.; Kayombo, C.J.; Musamba, E.B.; Kashindye, A.M.; Shayo, P.F. Species Composition, Richness and Diversity in Miombo Woodland of Bereku Forest Reserve, Tanzania. J. Biodivers. 2011, 2, 1–7. [Google Scholar] [CrossRef]
- Hanke, W.; Boehner, J.; Dreber, N.; Juergens, N.; Schmiedel, U.; Wesuls, D.; Dengler, J. The impact of livestock grazing on plant diversity: An analysis across dryland ecosystems and scales in southern Africa. Ecol. Appl. 2014, 24, 1188–1203. [Google Scholar] [CrossRef]
- Odadi, W.O.; Fargione, J.; Rubenstein, D.I. Vegetation, Wildlife, and Livestock Responses to Planned Grazing Management in an African Pastoral Landscape. Land Degrad. Dev. 2017, 28, 2030–2038. [Google Scholar] [CrossRef]
- Zambia Wildlife Authority. Kafue National Park General Management Plan 2011–2020; Zambia Wildlife Authority: Lusaka, Zambia, 2010. [Google Scholar]
- Asefa, A.; Yosef, M.; Mengesha, G.; Mamo, Y.; Shimelis, A. Woody plant diversity along disturbance gradients in the Northern Afro-Montane Forests of the Bale Mountains, Ethiopia. Int. J. Dev. Res. 2015, 5, 3745–3754. [Google Scholar]
- Shirima, D.D.; Pfeifer, M.; Platts, P.; Totland, Ø.; Moe, S.R. Interactions between Canopy Structure and Herbaceous Biomass along Environmental Gradients in Moist Forest and Dry Miombo Woodland of Tanzania. PLoS ONE 2015, 10, e0142784. [Google Scholar] [CrossRef] [PubMed]
- Day, A.D.; Ludeke, K.L. Plant Nutrients in Desert Environments; Springer: Berlin, Germany, 1993. [Google Scholar] [CrossRef]
- Silva, M.M.V.G.; Cabral-Pinto, M.M.S.; Dinis, P. Geochemistry of subtropical arenosols from Kuito region (Angola). Urbanization effects and environmental implications. J. Afr. Earth Sci. 2021, 183, 104307. [Google Scholar] [CrossRef]
- McCauley, A.; Jones, C.; Olson-Rutz, K. Soil pH and organic matter. Nutr. Manag. Modul. 2017, 8, 1–12. [Google Scholar]
- Kamelarczyk, K.B.F. Carbon Stock Assessment and Modelling in Zambia A UN-REDD Programme Study; National Forest Resource and Assessment Programme: Lusaka, Zambia, 2009. [Google Scholar]
- Shackleton, C.; Shackleton, S.E.; Buiten, E.; Bird, N. The importance of dry woodlands and forests in rural livelihoods and poverty alleviation in South Africa. For. Policy Econ. 2007, 9, 558–577. [Google Scholar] [CrossRef]
- Chidumayo, E. Forest degradation and recovery in a miombo woodland landscape in Zambia: 22 years of observations on permanent sample plots. For. Ecol. Manag. 2013, 291, 154–161. [Google Scholar] [CrossRef]
- Fonge, B.A.; Tchetcha, D.J.; Nkembi, L. Diversity, Distribution, and Abundance of Plants in Lewoh-Lebang in the Lebialem Highlands of Southwestern Cameroon. Int. J. Biodivers. 2013, 2013, 642579. [Google Scholar] [CrossRef]
- Marks, E.; Aflakpui, G.K.S.; Nkem, J.; Poch, R.M.; Khouma, M.; Kokou, K.; Sagoe, R.; Sebastià, M.-T. Conservation of soil organic carbon, biodiversity and the provision of other ecosystem services along climatic gradients in West Africa. Biogeosciences 2009, 6, 1825–1838. [Google Scholar] [CrossRef]
- Quaye, A.K.; Doe, E.K.; Attua, E.M.; Yiran, G.; Arthur, A.; Dogbatse, J.A.; Konlan, S.; Nkroma, Y.D.; Addo, D. Geospatial distribution of soil organic carbon and soil pH within the cocoa agroecological zones of Ghana. Geoderma 2021, 386, 114921. [Google Scholar] [CrossRef]
- Mapaure, I. Small-Scale Variations in Species Composition of Miombo Woodland in Sengwa, Zimbabwe: The Influence of Edaphic Factors, Fire and Elephant Herbivory. Syst. Geogr. Plants 2001, 71, 935. [Google Scholar] [CrossRef]
- Adams, F.; Evans, C.E. A Rapid Method for Measuring Lime Requirement of Red-Yellow Podzolic Soils. Soil Sci. Soc. Am. J. 1962, 26, 355–357. [Google Scholar] [CrossRef]
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern. Anal. Bioanal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Chapman, H.D. Total Exchangeable Bases. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; John Wiley: Hoboken, NJ, USA, 1965; pp. 902–904. [Google Scholar] [CrossRef]
- Lindsay, W.L.; Norvell, W.A. Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- Chapman, H.D.; Wilcox, L.V. Soluble salts. In Methods of Soil Analysis; Norman, A.G., Ed.; Soil Science Society of America: Madison, WI, USA, 1965. [Google Scholar] [CrossRef]


| Land-Use | Luampa (High Land-Use Intensity) [Median Values and (in Brackets) Min–Max Values] | Dongwe (Medium Land-Use Intensity) [Median Values and (in Brackets) Min–Max Values] | Kafue (No Land-Use) [Median Values and (in Brackets) Min–Max Values] |
|---|---|---|---|
| Woodcutting | 1 a (0–3) | 1 a (0–3) | 0 b (0–1) |
| Browsing | 0 a (0–1) | 0 a (0–1) | 1 b (0–2) |
| Time since last fire | 0 a (0–2) | 1 a (0–2) | 2 b (1–2) |
| Diversity Model | Distribution and Link | n | Intercept | logMg | SOC | pH | logFe ‡ | Wood Cutting ‡ | pR2 |
|---|---|---|---|---|---|---|---|---|---|
| Swoody | Poisson (log) | 60 | 3.275 *** | 0.00 | |||||
| Sherbs | Poisson (log) | 60 | 2.593 *** | 0.151 ** | 9.432 * | 0.36 | |||
| H’woody | Gaussian (id) | 60 | 11.880 *** | 0.935 | −5.970 | 0.07 | |||
| H’herbs | Gaussian (id) | 60 | 2.773 *** | 2.506 | −12.952 * | −2.141 * | 0.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sichone, P.; Oldeland, J.; Phiri, P.; Jürgens, N.; Schmiedel, U. Impacts of Diffuse Land-Use on Plant Diversity Patterns in the Miombo Woodlands of Western Zambia. Diversity 2023, 15, 739. https://doi.org/10.3390/d15060739
Sichone P, Oldeland J, Phiri P, Jürgens N, Schmiedel U. Impacts of Diffuse Land-Use on Plant Diversity Patterns in the Miombo Woodlands of Western Zambia. Diversity. 2023; 15(6):739. https://doi.org/10.3390/d15060739
Chicago/Turabian StyleSichone, Priscilla, Jens Oldeland, Patrick Phiri, Norbert Jürgens, and Ute Schmiedel. 2023. "Impacts of Diffuse Land-Use on Plant Diversity Patterns in the Miombo Woodlands of Western Zambia" Diversity 15, no. 6: 739. https://doi.org/10.3390/d15060739
APA StyleSichone, P., Oldeland, J., Phiri, P., Jürgens, N., & Schmiedel, U. (2023). Impacts of Diffuse Land-Use on Plant Diversity Patterns in the Miombo Woodlands of Western Zambia. Diversity, 15(6), 739. https://doi.org/10.3390/d15060739