

The Anodontini of Vietnam (Mollusca: Bivalvia: Unionidae: Unioninae) with the Description of a New Species

Abstract
1. Introduction
2. Materials and Methods
2.1. Field and Tissue Sampling
2.2. DNA Extraction, PCR, and Sequencing
2.3. Phylogenetic Analyses
2.4. Species Delimitation
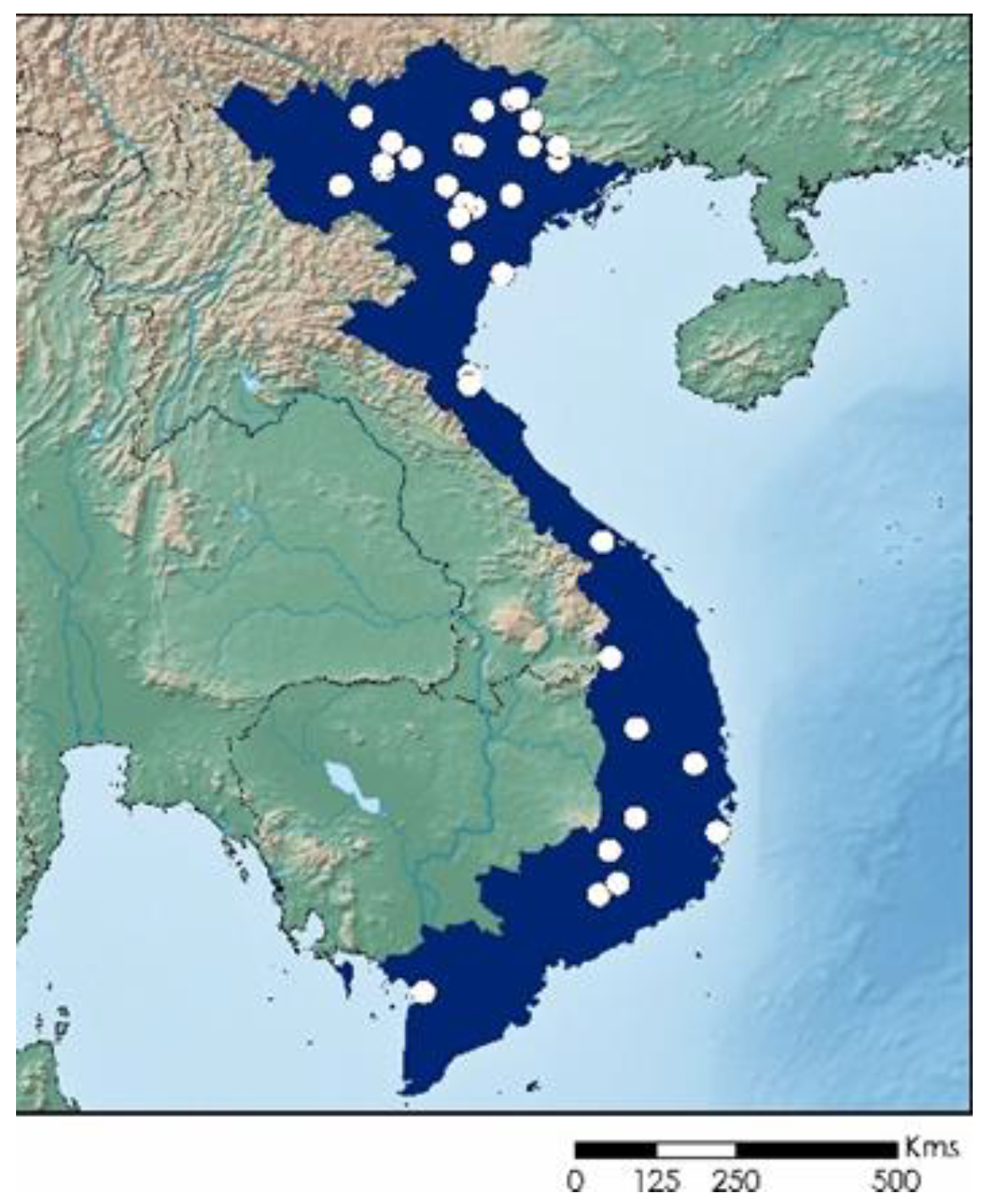
2.5. Distribution Maps
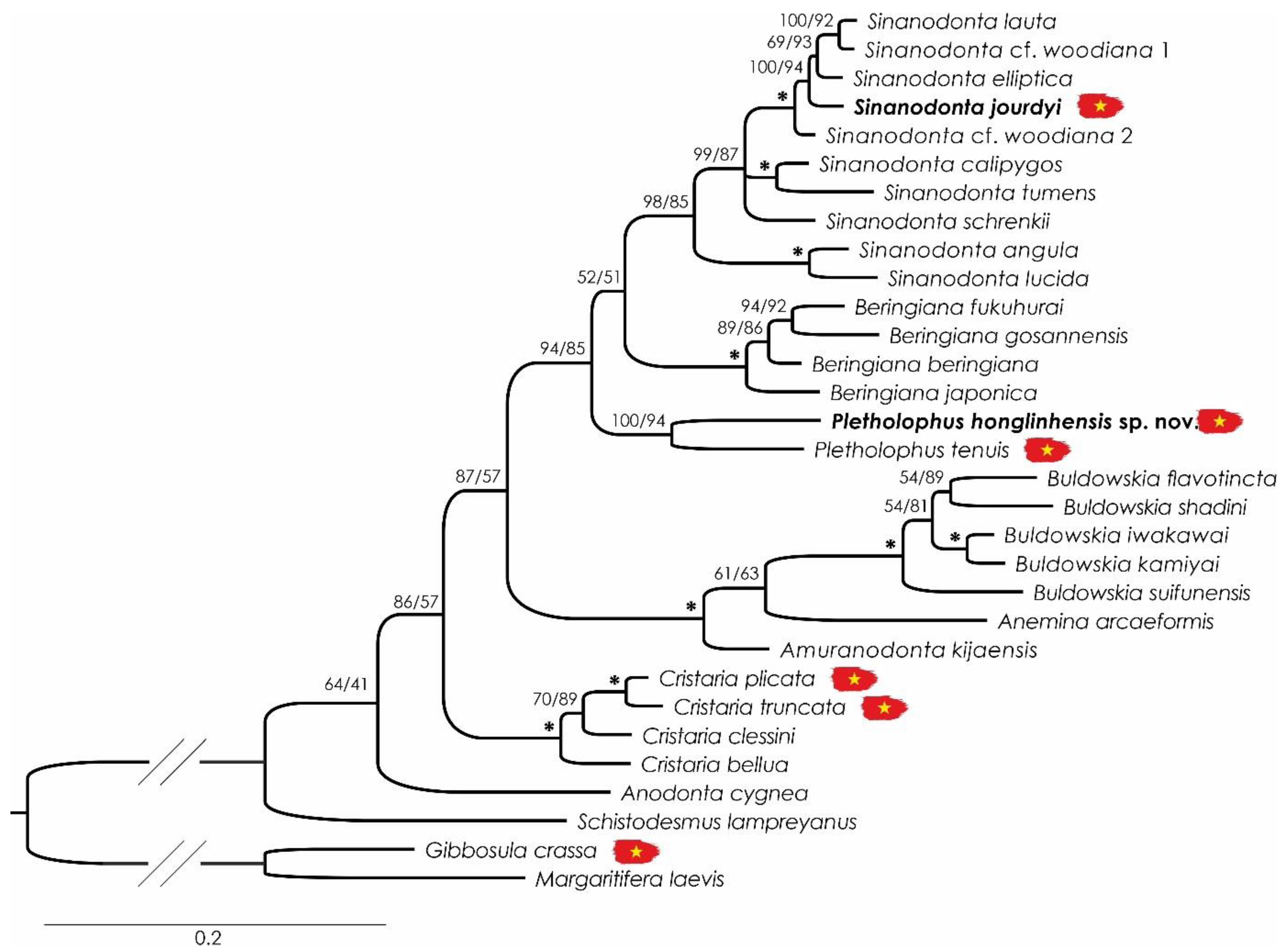
3. Results and Discussion
3.1. Taxonomy of Sinanodonta, Cristaria, and Pletholophus
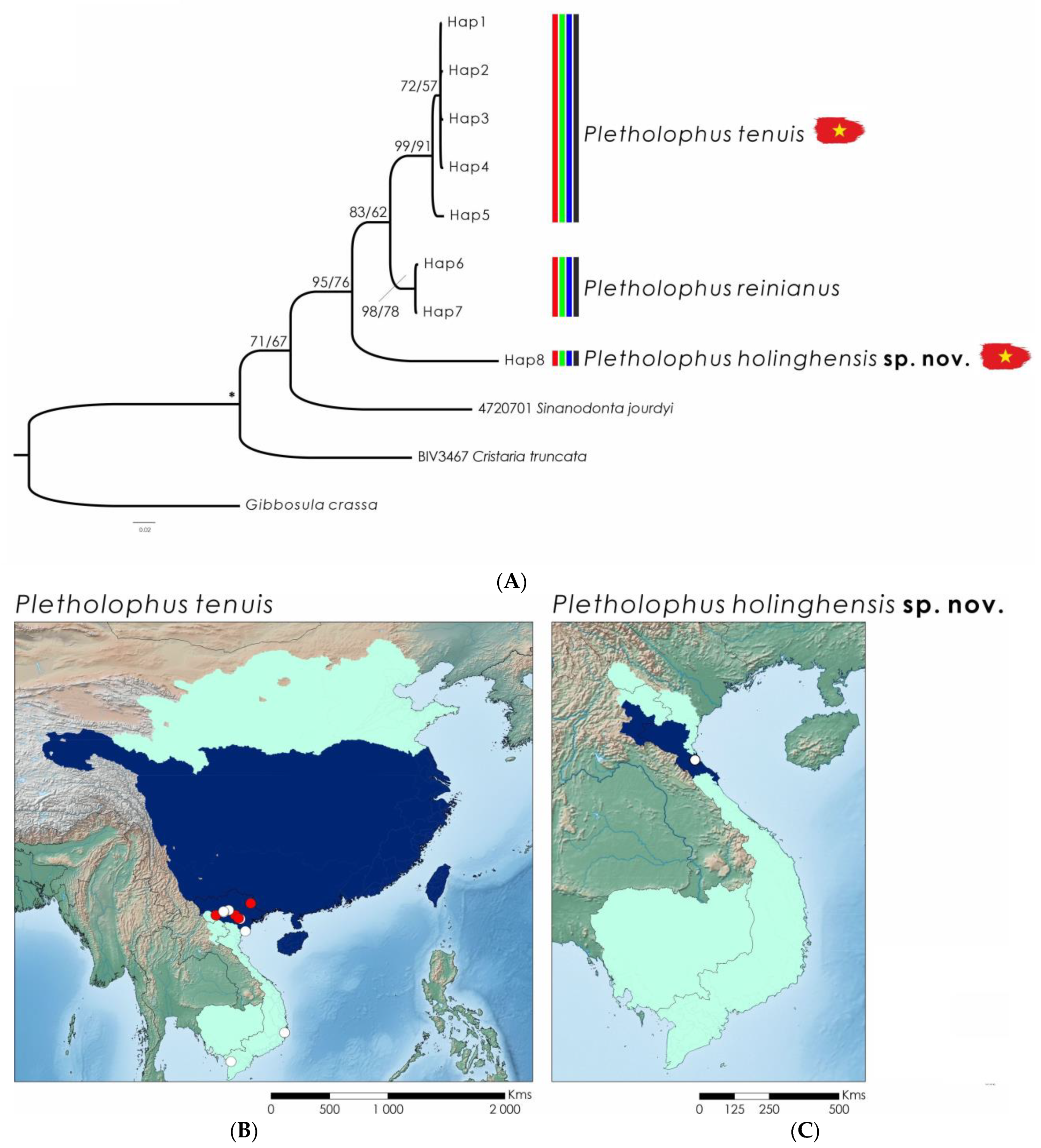
3.2. Systematics
3.3. Description
3.4. Pletholophus tenuis
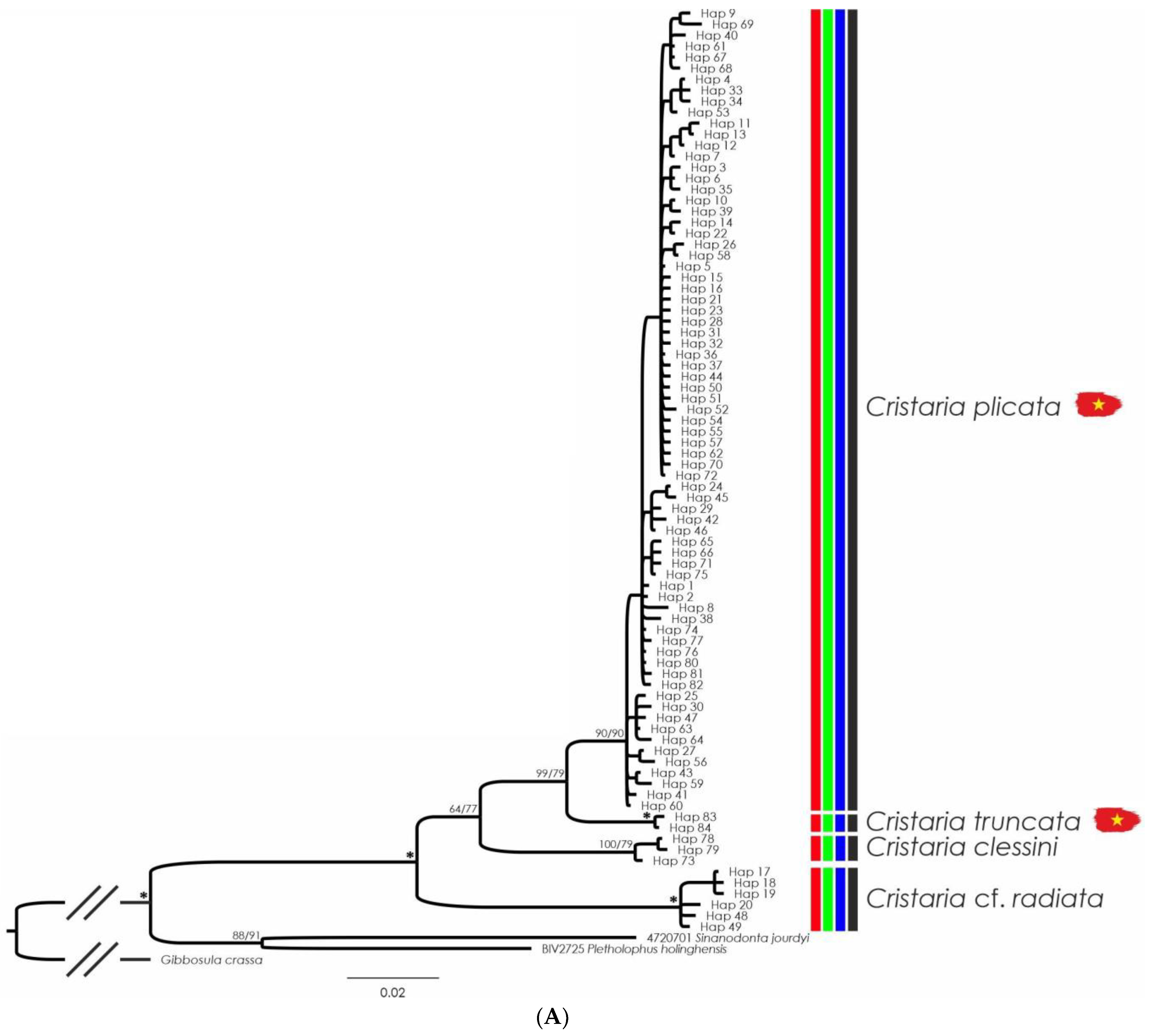
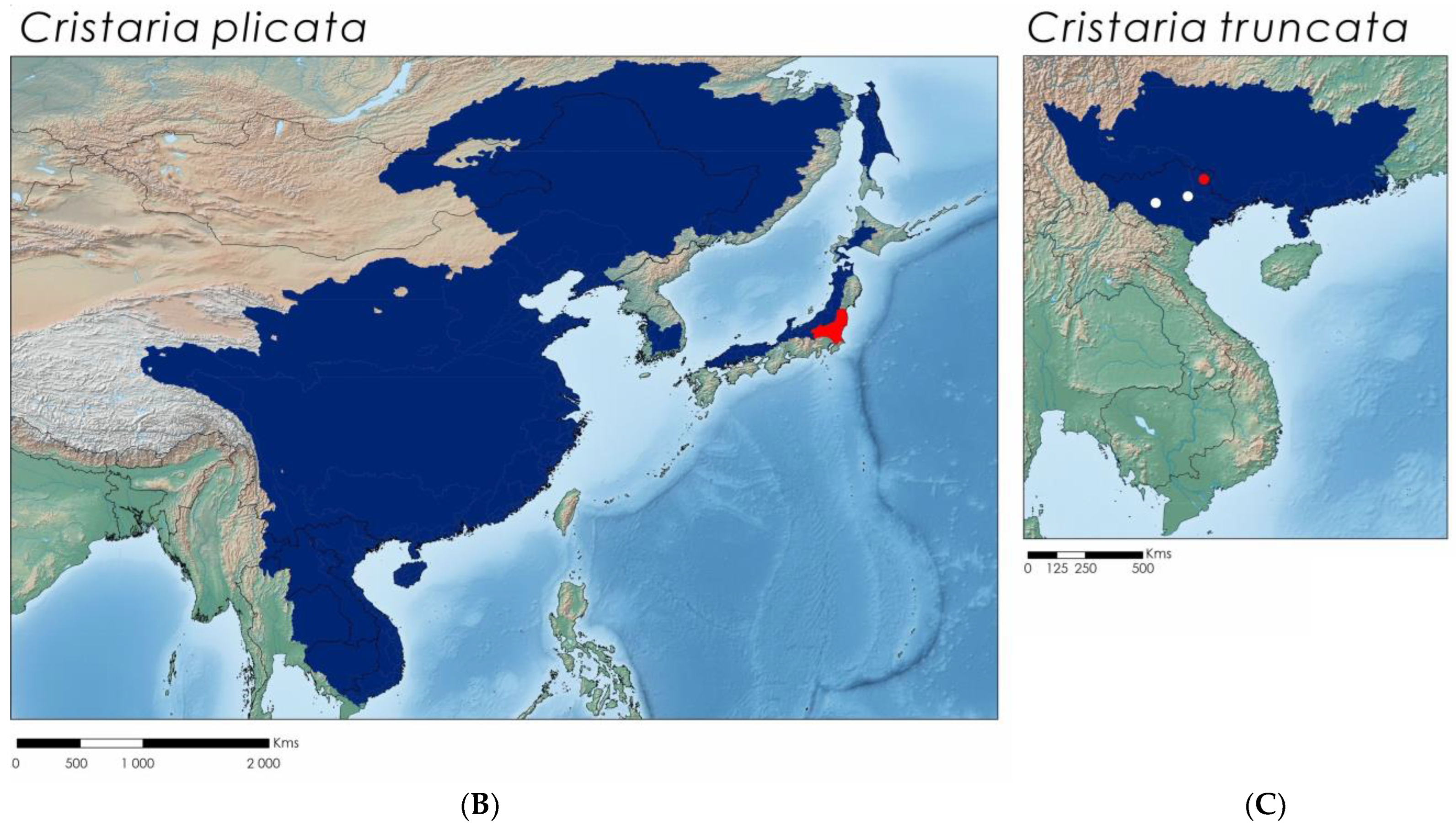
3.5. Cristaria plicata
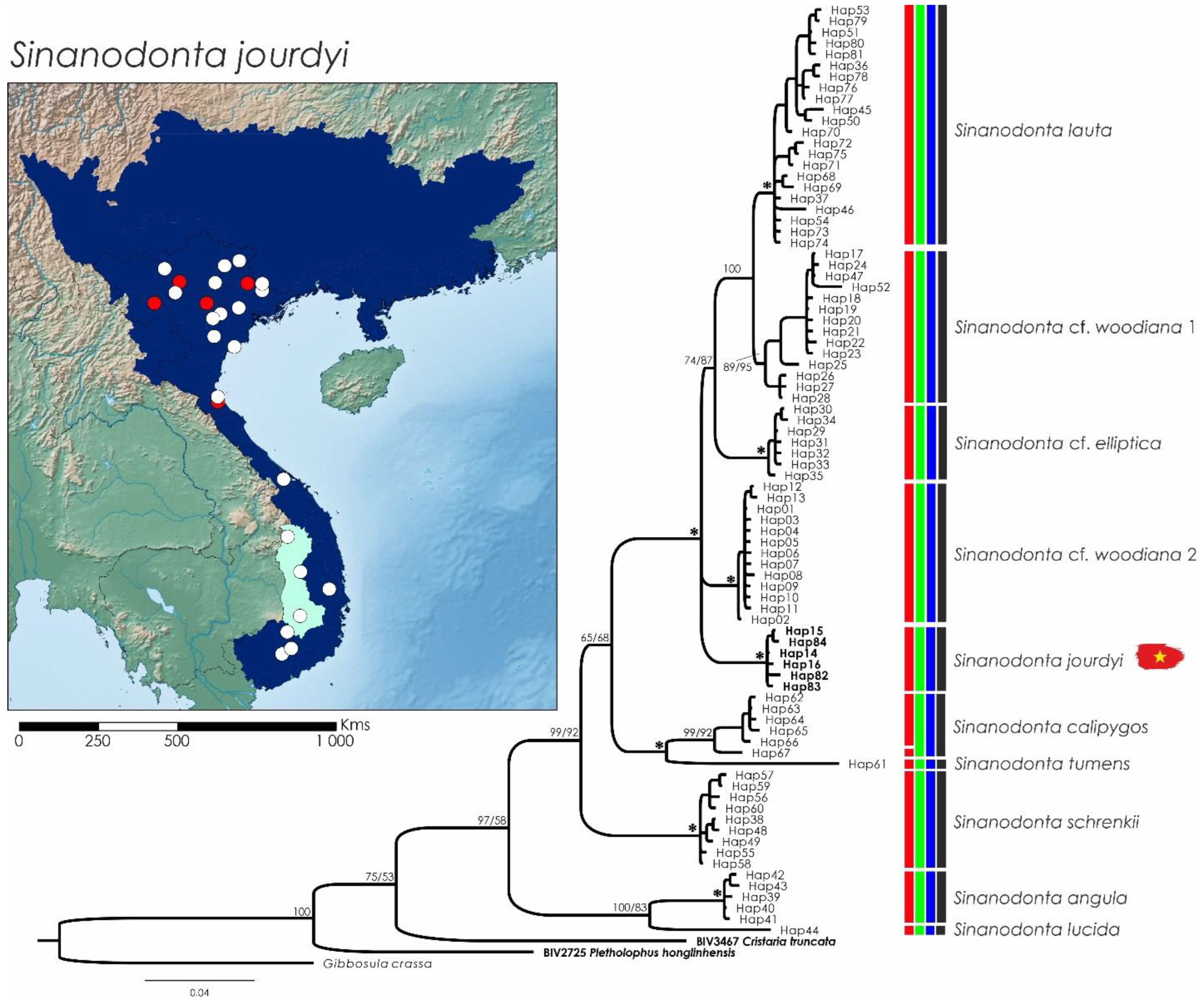
3.6. Sinanodonta jourdyi
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Modesto, V.; Ilarri, M.; Sopuza, A.T.; Lopes-Lima, M.; Douda, K.; Clavero, M.; Sousa, R. Fish and mussels: Importance of fish for freshwater mussel conservation. Fish Fish. 2018, 19, 244–259. [Google Scholar] [CrossRef]
- Guerra, D.; Lopes-Lima, M.; Froufe, E.; Gan, H.M.; Ondina, P.; Amaro, R.; Klunzinger, M.W.; Callil, C.; Prié, V.; Bogan, A.E.; et al. Variability of mitochondrial ORFans hints at possible differences in the system of doubly uniparental inheritance of mitochondria among families of freshwater mussels (Bivalvia: Unionida). BMC Evol. Biol. 2019, 19, 229. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, C.C. Ecosystem services provided by freshwater mussels. Hydrobiologia 2018, 810, 15–27. [Google Scholar] [CrossRef]
- Zieritz, A.; Froufe, E.; Bolotov, I.; Gonçalves, D.V.; Aldridge, D.C.; Bogan, A.E.; Gan, H.M.; Gomes-dos-Santos, A.; Sousa, R.; Teixeira, A.; et al. Mitogenomic phylogeny and fossil-calibrated mutation rates for all F- and M-type mtDNA genes of the largest freshwater mussel family (Bivalvia: Unionidae). Zool. J. Linn. Soc. 2021, 193, 1088–1107. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Bolotov, I.N.; Do, V.T.; Aldridge, D.C.; Fonseca, M.F.; Gan, H.M.; Gofarov, M.Y.; Kondakov, A.V.; Prié, V.; Sousa, R.; et al. Expansion and systematics redefinition of the most threatened freshwater mussel family, the Margaritiferidae. Mol. Phylogenet. Evol. 2018, 127, 98–118. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Teixeira, A.; Froufe, E.; Lopes, A.; Varandas, S.; Sousa, R. Biology and conservation of freshwater bivalves: Past, present and future perspectives. Hydrobiologia 2014, 735, 1–13. [Google Scholar] [CrossRef]
- Böhm, M.; Dewhurst-Richman, N.I.; Seddon, M.; Ledger, S.E.H.; Albrecht, C.; Allen, D.; Bogan, A.E.; Cordeiro, J.; Cummings, K.S.; Cuttelod, A.; et al. The conservation status of the world’s freshwater molluscs. Hydrobiologia 2021, 848, 3231–3254. [Google Scholar] [CrossRef]
- Zieritz, A.; Bogan, A.E.; Froufe, E.; Klishko, A.; Kondo, T.; Kovitvadhi, U.; Kovitvadhi, S.; Lee, J.H.; Lopes-Lima, M.; Pfeiffer, J.M.; et al. Diversity, biogeography, and conservation of freshwater mussels (Bivalvia: Unionida) in East and Southeast Asia. Hydrobiologia 2018, 810, 29–44. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Vikhrev, I.V.; Kondakov, A.V.; Konopleva, E.S.; Gofarov, M.Y.; Aksenova, O.V.; Tumpeesuwan, S. New taxa of freshwater mussels (Unionidae) from a species-rich but overlooked evolutionary hotspot in Southeast Asia. Sci. Rep. 2017, 7, 11573. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Pfeiffer, J.M.; Konopleva, E.S.; Vikhrev, I.V.; Kondakov, A.V.; Aksenova, O.V.; Gofarov, M.Y.; Tumpeesuwan, S.; Win, T. A new genus and tribe of freshwater mussel (Unionidae) from Southeast Asia. Sci. Rep. 2018, 8, 10030. [Google Scholar] [CrossRef]
- Régnier, C.; Fontaine, B.; Bouchet, P. Not knowing, not recording, not listing: Numerous unnoticed mollusk extinctions. Conser. Biol. 2009, 23, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Régnier, C.; Achaz, G.; Lambert, A.; Cowie, R.H.; Bouchet, P.; Fontaine, B. Mass extinction in poorly known taxa. Proc. Nat. Acad. Sci. USA 2015, 112, 7761–7766. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, P.A.; Bigorne, R.; Bogan, A.E.; Giam, X.; Jezequel, C.; Hugueny, B. Estimating how many undescribed species have gone extinct. Conser. Biol. 2014, 28, 1360–1370. [Google Scholar] [CrossRef] [PubMed]
- Bogan, A.E.; Do, V.T. An overlooked new species of freshwater bivalve from Northern Vietnam (Mollusca: Bivalvia: Unionidae). Raffles Bull. Zool. 2018, 66, 878–889. [Google Scholar]
- Bolotov, I.N.; Konopleva, E.S.; Vikhrev, I.V.; Gofarov, M.Y.; Lopes-Lima, M.; Bogan, A.E.; Lunn, Z.; Chan, N.; Win, T.; Aksenova, O.V.; et al. New freshwater mussel taxa discoveries clarify biogeographic division of Southeast Asia. Sci. Rep. 2020, 10, 6616. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Kondakov, A.V.; Konopleva, E.S.; Vikhrev, I.V. A new genus of ultra-elongate freshwater mussels from Vietnam and eastern China (Bivalvia: Unionidae). Ecol. Montenegrina 2021, 39, 1–6. Available online: https://www.biotaxa.org/em/article/view/em.2021.39.1 (accessed on 20 March 2020). [CrossRef]
- Huang, X.C.; Su, J.H.; Ouyang, J.X.; Ouyang, S.; Zhou, S.H.; Wu, X.P. Towards a global phylogeny of freshwater mussels (Bivalvia: Unionida): Species delimitation of Chinese taxa, mitochondrial phylogenomics, and diversification patterns. Mol. Phylogenet. Evol. 2019, 130, 45–59. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Hattori, A.; Kondo, T.; Lee, J.H.; Kim, S.K.; Shirai, A.; Hayashi, H.; Usui, T.; Sakuma, K.; Toriya, T.; et al. Freshwater mussels (Bivalvia: Unionidae) from the Rising Sun (Far East Asia): Phylogeny, systematics, and distribution. Mol. Phylogenet. Evol. 2020, 146, 106755. [Google Scholar] [CrossRef]
- Ng, T.H.; Jeratthitikul, E.; Sutcharit, C.; Chhuoy, S.; Pin, K.; Pholyotha, A.; Siriwut, W.; Srisonchai, R.; Hogan, Z.S.; Ngor, P.B. Annotated checklist of freshwater molluscs from the largest freshwater lake in Southeast Asia. ZooKeys 2020, 958, 107–141. [Google Scholar] [CrossRef]
- Sano, I.; Saito, T.; Miyazaki, J.I.; Shirai, A.; Uechi, T.; Kondo, T.; Chiba, S. Evolutionary History and Diversity of Unionoid Mussels (Mollusca: Bivalvia) in the Japanese Archipelago. Plankton Benthos Res. 2020, 15, 97–111. [Google Scholar] [CrossRef]
- Konopleva, E.S.; Bolotov, I.N.; Pfeifer, J.M.; Vikhrev, I.V.; Kondakov, A.V.; Gofarov, M.Y.; Tomilova, A.A.; Tanmuangpak, K.; Tumpeesuwan, S. New freshwater mussels from two Southeast Asian genera Bineurus and Thaiconcha (Pseudodontini, Gonideinae, Unionidae). Sci. Rep. 2021, 11, 8244. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, J.M.; Graf, D.L.; Cummings, K.S.; Page, L.M. Taxonomic revision of a radiation of South-east Asian freshwater mussels (Unionidae: Gonideinae: Contradentini+Rectidentini). Invertebr. Systemat. 2021, 35, 394–470. [Google Scholar] [CrossRef]
- Sahidin, A.; Muhammad, G.; Hasan, Z.; Arief, M.C.W.; Marwoto, R.M.; Komaru, A. Indonesian freshwater bivalves: A meta-analysis of endemicity, ecoregion distributions, and conservation status. AACL Aquac. Aquar. Conserv. Legis. Bioflux 2021, 14, 3750–3775. [Google Scholar]
- Wu, R.W.; Liu, X.J.; Wang, S.; Roe, K.J.; Ouyang, S.; Wu, X.P. Analysis of mitochondrial genomes resolves the phylogenetic position of Chinese freshwater mussels (Bivalvia, Unionidae). ZooKeys 2019, 812, 23–46. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.P.; Dai, Y.T.; Yin, N.; Shu, F.Y.; Chen, Z.G.; Guo, L.; Zhou, C.H.; Ouyang, S.; Huang, X.C. Mitogenomic phylogeny resolves Cuneopsis (Bivalvia: Unionidae) as polyphyletic: The description of two new genera and a new species. Zool. Scr. 2022, 51, 173–184. [Google Scholar] [CrossRef]
- Do, V.T.; Le, Q.T.; Bogan, A.E. Freshwater mussels (Bivalvia: Unionoida) of Vietnam: Diversity, distribution, and conservation status. Freshw. Mollusk Biol. Conserv. 2018, 21, 1–18. [Google Scholar] [CrossRef]
- Naimo, T.J.; Damschen, E.D.; Rada, R.G.; Monroe, E.M. Nonlethal evaluation of the physiological health of unionid mussels: Methods for biopsy and glycogen analysis. J. N. Am. Benthol. Soc. 1998, 17, 121–128. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Harbor Spring Press: New York, NY, USA, 1989; p. 545. [Google Scholar]
- Froufe, E.; Sobral, C.; Teixeira, A.; Sousa, R.; Varandas, S.C.; Aldridge, D.; Lopes-Lima, M. Genetic diversity of the pan-European freshwater mussel Anodonta anatina (Bivalvia: Unionoida) based on CO1: New phylogenetic insights and implications for conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 24, 561–574. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Sela, I.; Ashkenazy, H.; Katoh, K.; Pupko, T. GUIDANCE2: Accurate detection of unreliable alignment regions accounting for the uncertainty of multiple parameters. Nucleic Acids Res. 2015, 43, W7–W14. [Google Scholar] [CrossRef]
- Fonseca, M.M.; Lopes-Lima, M.; Eackles, M.S.; King, T.L.; Froufe, E. The female and male mitochondrial genomes of Unio delphinus and the phylogeny of freshwater mussels (Bivalvia: Unionida). Mitochondrial DNA Part B 2016, 1, 954–957. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating Maximum-Likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A.; Susko, E. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Xia, X. DAMBE6: New tools for microbial genomics, phylogenetics and molecular evolution. J. Hered. 2017, 108, 431–437. [Google Scholar] [CrossRef]
- Konopleva, E.S.; Bolotov, I.N.; Vikhrev, I.V.; Inkhavilay, K.; Gofarov, M.Y.; Kondakov, A.V.; Tomilova, A.A.; Chapurina, Y.E.; Do, V.T.; Pfeiffer, J.M., III; et al. A freshwater mussel species reflects a Miocene stream capture between the Mekong Basin and East Asian rivers. Zoosyst. Evol. 2023, 99, 29–42. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Froufe, E.; Do, V.T.; Ghamizi, M.; Mock, K.E.; Kebapçi, U.; Klishko, O.; Kovitvadhi, S.; Kovitvadhi, U.; Paul, O.S.; et al. Phylogeny of the most species rich freshwater bivalve family (Bivalvia: Unionida: Unionidae): Defining modern subfamilies and tribes. Mol. Phylogenet. Evol. 2017, 106, 174–191. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-based registry for all animal species: The Barcode Index Number (BIN) system. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef]
- Puillandre, N.; Modica, M.V.; Zhang, Y.; Sirovich, L.; Boisselier, M.-C.; Cruaud, C.; Holford, M.; Samadi, S. Large-scale species delimitation method for hyperdiverse groups. Mol. Ecol. 2012, 21, 2671–2691. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Đặng, N.T.; Thái, T.B.; Phạm, V.M. Identification of freshwater invertebrates of North Vietnam. In Định Loại Động Vật Không Xương Sống Nước Ngọt Bắc Việt Nam; Sciences and Technology Publishing Co.: Ha Noi, Vietnam, 1980; pp. 491–573. (In Vietnamese) [Google Scholar]
- Đặng, N.T.; Hồ, T.H. Trai Ốc Nước Ngọt Nội Địa Việt Nam (Mollusca: Gastropoda, Bivalvia); [Inland Freshwater Molluscs (Mollusca: Gastropoda, Bivalvia) of Vietnam] Ðộng Vật Chí Việt Nam [Fauna of Vietnam]; Science and Technology Publishing House: Hanoi, Vietnam, 2017; 360p, ISBN 978-604-913-677-1. (In Vietnamese, English Abstract). [Google Scholar]
- International Union for the Conservation of Nature (IUCN). The IUCN Red List of Threatened Species. 2023. Available online: https://www.iucnredlist.org/ (accessed on 1 January 2023).
- Lehner, B.; Grill, G. Global River hydrography and network routing: Baseline data and new approaches to study the world’s large river systems. Hydrol. Process. 2013, 27, 2171–2186. [Google Scholar] [CrossRef]
- von Martens, E. Herr von Martens gab im Anschluss an den vorhergehenden Vortrag eine Uebersicht über die von den Herren Dr. Fr. Hilgendorf und Dr. W. Dönitz in Japan gesammelten Binnenmollusken. Sitz. Ber. Ges. Nat. Freunde Berl. 1877, 1877, 97–123. [Google Scholar]
- Tchang, S.; Li, S.C.; Liu, Y.Y. Bivalves (Mollusca) of Tung-Ting Lake and its surrounding waters, Hunan Province, China. Acta Zool. Sin. 1965, 17, 197–213. [Google Scholar]
- Heude, P. Conchyliologie Fluviatile de la Province de Nanking et de la Chine Centrale; 10 Fascicules, plates, and un-numbered pages; Librairie F. Savy: Paris, France, 1875. [Google Scholar]
- Haas, F. Superfamilia Unionacea. Das. Tierreich. 1969, 88, 663. (In German) [Google Scholar]
- Graf, D.; Cummings, K. Review of the systematics and global diversity of freshwater mussel species (Bivalvia: Unionoida). J. Molluscan Stud. 2007, 73, 291–314. [Google Scholar] [CrossRef]
- Lea, I. Observations on the naïades; and descriptions of new species of that, and other families. Trans. Am. Philos. Soc. 1834, 5, 23–119. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Bespalaya, Y.V.; Gofarov, M.Y.; Kondakov, A.V.; Konopleva, E.S.; Vikhrev, I.V. Spreading of the Chinese pond mussel, Sinanodonta woodiana, across Wallacea: One or more lineages invade tropical island and Europe. Biochem. Syst. Ecol. 2016, 67, 58–64. [Google Scholar] [CrossRef]
- Morlet, L. Diagnoses Molluscorum novorum Tonkini. J. Conchyliol. 1886, 34, 75–78. [Google Scholar]
- Bespalaya, Y.V.; Bolotov, I.N.; Aksenova, O.V.; Gofarov, M.Y.; Kondakov, A.V.; Vikhrev, I.V.; Vinarski, M.V. DNA barcoding reveals invasion of two cryptic Sinanodonta mussel species (Bivalvia: Unionidae) into the largest Siberian river. Limnologica 2018, 69, 94–102. [Google Scholar] [CrossRef]
- Kondakov, A.V.; Palatov, D.M.; Rajabov, Z.P.; Gofarov, M.Y.; Konopleva, E.S.; Tomilova, A.A.; Vikhrev, I.V.; Bolotov, I.N. DNA analysis of a non-native lineage of Sinanodonta woodiana species complex (Bivalvia: Unionidae) from Middle Asia supports the Chinese origin of the European invaders. Zootaxa 2018, 4462, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Graf, D.; Cummings, K. A ‘big data’ approach to global freshwater mussel diversity (Bivalvia: Unionoida), with an updated checklist of genera and species. J. Molluscan Stud. 2021, 87, 1–36. [Google Scholar] [CrossRef]
- Graf, D.; Cummings, K. Mussel-Project Webpage. 2023. Available online: http://mussel-project.uwsp.edu/ (accessed on 1 February 2023).
- Schumacher, C.F. Essai d’un Nouveau Système des Habitations des Vers Testacés Avec XXII Planches; De l’Imprimerie de Mr. le Directeur Schultz: Copenhague, Denmark, 1817; 287p. [Google Scholar]
- Simpson, C.T. Synopsis of the naiades, or pearly fresh-water mussels. Proc. U.S. Natl. Mus. 1900, 22, 501–1044. [Google Scholar] [CrossRef]
- Heard, W.H. Anatomical systematics of Cristaria plicata (Leach) (Pelecypoda: Unionidae, Unioninae). Malacol. Rev. 1977, 10, 59–70. [Google Scholar]
- He, J.; Zhuang, Z. The Freshwater Bivalves of China; Conchbooks: Harxheim, Germany, 2013; 197p. [Google Scholar]
- Moskvicheva, I.M.; Starobogatov, Y.I. About East-Asian Potomida similar Unionidae (Bivalvia). Bull. Mosc. Soc. Nat. Investig. Sect. Biol. 1973, 78, 21–37. (In Russian) [Google Scholar]
- Klishko, O.; Lima-Lopez, M.; Froufe, E.; Bogan, A.E. Are Cristaria herculea (Middendorff, 1847) and Cristaria plicata (Leach, 1815) (Bivalvia, Unionidae, Unioninae, Anodontini) separate species? Zookeys 2014, 438, 1–15. [Google Scholar] [CrossRef]
- Klishko, O.; Lopes-Lima, M.; Froufe, E.; Bogan, A.E.; Abakumova, V.Y. Systematics and distribution of Cristaria plicata (Bivalvia, Unionidae) from the Russian Far East. Zookeys 2016, 580, 13–27. [Google Scholar] [CrossRef]
- Brandt, R.A.M. The non-marine aquatic Mollusca of Thailand. Arch. Molluskenkd. 1974, 105, 1–423. [Google Scholar]
- Kondo, T. Monograph of Unionoida in Japan (Mollusca: Bivalvia); Special publication of the Malacological Society of Japan; Malacological Society of Japan: Ibaraki, Japan, 2008; 69p. [Google Scholar]
- GenBank. Nucleotide Sequences Database for the National Center for Biotechnology Information (NCBI). 2022. Available online: https://www.ncbi.nlm.nih.gov/nuccore/?term=Cristaria+plicata+mitochondrion (accessed on 1 January 2023).
- MolluscaBase. 2023. Available online: http://www.molluscabase.org/ (accessed on 1 January 2023).
- Simpson, C.T. A Descriptive Catalogue of the Naiades, or Pearly Fresh-Water Mussels; Parts I–III; Bryant Walker: Detroit, MI, USA, 1914; 1540p. [Google Scholar]
- Leach, W.E. The Zoological Miscellany; Being Descriptions of New, or Interesting Animals; McMillan: London, UK, 1814; Volume 1, 134p. [Google Scholar]
- Liu, Y.Y.; Zhang, W.; Wang, Y.; Wang, E. Economic Fauna of China. Freshwater Mollusks; Science Press: Beijing, China, 1979; 134p. (In Chinese) [Google Scholar]
- Lea, I. A Synopsis of the Family of Naïades; Cary, Lea and Blanchard, Philadelphia; John Miller: London, UK, 1836; 59p. [Google Scholar]
- Lea, I. A Synopsis of the Family of Naïades, 2nd ed.; Enlarged and Improved; Cary, Lea and Blanchard: Philadelphia, PA, USA, 1838; 44p. [Google Scholar]
- Lea, I. A Synopsis of the Family of Naïades, 3rd ed.; Greatly Enlarged and Improved; Blanchard and Lea: Philadelphia, PA, USA, 1852; 88p. [Google Scholar]
- Lea, I. A Synopsis of the Family Unionidae, 4th ed.; Very Greatly Enlarged and Improved; Henry C. Lea: Philadelphia, PA, USA, 1870; 184p. [Google Scholar]
- Preston, H.B. A catalogue of the Asiatic naiades in the collection of the Indian Museum, Calcutta, with descriptions of new species. Rec. Indian Mus. 1912, 7, 279–308. [Google Scholar] [CrossRef]
- Modell, H. Die Anodontinae, Ortm. Emend. (Najad., Mollusca). Eine studie über die Zusammenhänge von Klimazonen und Entwicklungsgeschichte. (Klimazonentheorie). Jena. Z. Med. Nat. 1945, 78, 58–100. [Google Scholar]
- Petit, R.E.; Coan, E.V. The molluscan taxa made available in the Griffith& Pidgeon (1833–1834) edition of Cuvier, with notes on the editions of Cuvier and on Wood’s Index Testaceologicus. Malacologia 2008, 50, 219–264. [Google Scholar]
- Griffith, E.; Pidgeon, E. The Mollusca and Radiata. In The Animal Kingdom Arranged in Conformity with Its Organization; Griffith, E., Ed.; Whittaker, and Co.: London, UK, 1833; Volume 12, pp. 8–601. [Google Scholar]
- International Commission on Zoological Nomenclature (ICZN). Incorporating Declaration 44, amendments of Article 74.7.3, with effect from 31 December 1999, and the Amendment on e-publication, amendments to Articles 8, 9, 10, 21 and 78, with effect from 1 January 2012. In International Code of Zoological Nomenclature, 4th ed.; International Trust for Zoological Nomenclature: Singapore, 1999; 306p, Available online: http://www.nhm.ac.uk/hosted-sites/iczn/code/ (accessed on 1 January 2023).
- Imai, H. The first record of Cristaria discoidea (Lea, 1834) (Bivalvia: Unionidae) from Ishigakijima Island, the Ryukyu Archipelago, Japan. Biol. Mag. Okinawa 2008, 46, 65–70. [Google Scholar]
- von Martens, E. Cristaria reiniana n. sp. Jahrbücher Dtsch. Malakozool. Ges. 1875, 2, 136. [Google Scholar]
- Habe, T. Shells of Japan; Color Books; Hoikusha Publishing Co. Ltd.: Osaka, Japan, 1971; 139p. [Google Scholar]
- Masuda, O.; Uchiyama, R. Freshwater Mollusks of Japan. Series 2. Freshwater Mollusks of Japan, Including Brackish Water Species; PICES, Ecological Field Guide Series; Pices Publishers, Co. Ltd.: Tokyo, Japan, 2004; 240p. (In Japanese) [Google Scholar]
- Habe, T. Bivalvia and Scaphopoda; Systematics of Mollusca in Japan; Hokuryukan Publisher: Tokyo, Japan, 1977; 372p. [Google Scholar]
- Linnaeus, C. Systema Naturae. Edition X. (Systema Naturae per Regna tria Naturae, Secundum Classes, Ordines, Genera, Species cum Characteribus, Differentiis, Synonymis, Locis. Tomus, I. Edtio Decima, Reformata); Laurentius Salvius: Stockholm, Sweden, 1758; Volume 1, pp. 1–824. [Google Scholar]
- Gray, J.E. A revision of the arrangement of the families of bivalve shells (Conchifera). Ann. Mag. Nat. Hist. 1854, 13, 408–418. [Google Scholar] [CrossRef]
- Rafinesque, C.S. Monographie des coquilles bivalves fluviatiles de la Rivière Ohio, contenant douze genres et soixante-huit espèces. Ann. Générales Des Sci. Phys. À Brux. 1820, 5, 287–322. [Google Scholar]
- Lee, J.H. Systematic Study of Korean Unionids (Bivalvia: Unionidae) Based on Morphological and Molecular Data. Ph.D. Thesis, School of Life Sciences, Kungpook National University, Daegu, Republic of Korea, 2017. [Google Scholar]
- Lee, J.H.; Choi, E.H.; Kim, S.K.; Ryu, S.H.; Hwang, U.W. Mitochondrial genome of the cockscomb pearl mussel Cristaria plicata (Bivalvia, Unionoida, Unionidae). Mitogenome Anoun. 2012, 23, 39–41. [Google Scholar]
- Jia, M.J.; Li, J.L.; Niu, D.H.; Bai, Z.Y. Sequence variation of COI gene in ten populations of Cristaria plicata from the Middle and Lower Yangtze River. Chin. J. Zool. 2009, 44, 1–8. [Google Scholar]
- Wang, X.F.; Liu, Z.M.; Wu, W.J. Transcriptome analysis of the freshwater pearl mussel (Cristaria plcaat) mantle unravels genes involved in the formation of shell and pearls. Mol. Genet. Genom. 2017, 292, 343–352. [Google Scholar] [CrossRef]
- Wu, R.W.; Liu, Y.T.; Wang, S.; Liu, X.J.; Zanatta, D.T.; Roe, K.J.; Song, X.L.; An, C.T.; Wu, X.P. Testing the utility of DNA barcodes and a preliminary phylogenetic framework for Chinese freshwater mussels (Bivalvia: Unionidae) from the middle and lower Yangtze River. PloS ONE 2018, 13, e0200956. [Google Scholar] [CrossRef]
- Song, X.L.; Ouyang, S.; Zhou, C.H.; Wu, X.P. Complete maternal mitochondrial genome of freshwater mussel Anodonta lucida (Bivalvia: Unionidae: Anodontinae). Mitochondrial DNA Part A 2016, 27, 549–550. [Google Scholar] [CrossRef] [PubMed]
- Sayenko, E.M.; Soroka, M.; Kholin, S.K. Comparison of the species Sinanodonta amurensis Moskvicheva, 1973 and Sinanodonta primorjensis Bogatov et Zatrawkin, 1988 (Bivalvia: Unionidae: Anodontinae) in view of variability of the mitochondrial DNA cox1 gene and conchological features. Biol. Bull. 2017, 44, 266–276. [Google Scholar] [CrossRef]
- Ng, T.H.; Tan, S.K.; Wong, W.H.; Meier, R.; Chan, S. Molluscs for Sale: Assessment of Freshwater Gastropods and Bivalves in the Ornamental Pet Trade. PLoS ONE 2016, 11, e0161130. [Google Scholar] [CrossRef] [PubMed]
- Stelbrink, B.; Rintelen, T.V.; Albrecht, C.; Clewing, C.; Naga, P.O. Forgotten for decades: Lake Lanao and the genetic assessment of its mollusc diversity. Hydrobiologi 2019, 843, 31–49. [Google Scholar] [CrossRef]
- Zieritz, A.; Lopes-Lima, M.; Bogan, A.E.; Sousa, R.; Walton, S.; Rahim, K.A.; Wilson, J.J.; Ng, P.Y.; Froufe, E.; McGowan, S. Factors driving changes in freshwater mussel (Bivalvia, Unionida) diversity and distribution in Peninsular Malaysia. Sci. Total Environ. 2016, 571, 1069–1078. [Google Scholar] [CrossRef]
- Clusa, L.; Miralles, L.; Basanta, A.; Escot, C.; Garcı, E. eDNA for detection of five highly invasive molluscs. A case study in urban rivers from the Iberian Peninsula. PloS ONE 2017, 12, e0188126. [Google Scholar] [CrossRef]
- Konečný, A.; Popa, O.P.; Bartáková, V.; Douda, K.; Bryja, J.; Smith, C.; Popa, L.O.; Reichard, M. Modelling the invasion history of Sinanodonta woodiana in Europe: Tracking the routes of a sedentary aquatic invader with mobile parasitic larvae. Evol. Appl. 2018, 11, 1975–1989. [Google Scholar] [CrossRef]
- Soroka, M. Genetic variability among freshwater mussel Anodonta woodiana (Lea, 1834) (Bivalvia: Unionidae) populations recently introduced in Poland. Zool. Sci. 2005, 22, 1137–1144. [Google Scholar] [CrossRef]
- Soroka, M. Characteristics of Mitochondrial DNA of Unionid Bivalves (Mollusca: Bivalvia: Unionidae). I. Detection andcharacteristics of doubly uniparental inheritance (DUI) of Unionid Mitochondrial DNA (1). Folia Malacol. 2010, 18, 147–188. [Google Scholar] [CrossRef][Green Version]
- Soroka, M. Characteristics of mitochondrial DNA of unionid bivalves (Mollusca: Bivalvia: Unionidae). II. Detection and characteristics of doubly uniparental inheritance (DUI) of unionid mitochondrial DNA (2). Folia Malacol. 2010, 18, 189–209. [Google Scholar] [CrossRef][Green Version]
- Soroka, M.; Urbańska, M.; Andrzejewsk, W. Chinese pond mussel Sinanodonta woodiana (Lea, 1834) (Bivalvia): Origin of the Polish population and GenBank data. J. Limnol. 2014, 73, 454–458. [Google Scholar] [CrossRef]
- Vikhrev, I.V.; Konopleva, E.S.; Gofarov, M.Y.; Kondakov, A.V.; Chapurina, Y.E.; Bolotov, I.N. A Tropical Biodiversity Hotspot Under the New Threat: Discovery and DNA Barcoding of the Invasive Chinese Pond Mussel Sinanodonta Woodiana in Myanmar. Trop. Conserv. Sci. 2017, 10, 1–11. [Google Scholar] [CrossRef]
- Guarneri, I.; Popa, O.P.; Gola, L.L.; Kamburska, L.; Lauceri, R.; Lopes-Lima, M.; Popa, L.O.; Riccardi, N. A morphometric and genetic comparison of Sinanodonta woodiana (Lea, 1834) populations: Does shape really matter? Aquat. Invasions 2014, 9, 183–194. [Google Scholar] [CrossRef]
- Froufe, E.; Lopes-Lima, M.; Riccardi, N.; Zaccara, S.; Vanetti, I.; Lajtner, J.; Teixeira, A.; Varandas, S.; Prié, V.; Zieritz, A.; et al. Lifting the curtain on the freshwater mussel diversity from the Italian Peninsula and Croatian Adriatic coast. Biodivers. Conserv. 2017, 26, 3255–3274. [Google Scholar] [CrossRef]
- Zhang, P.; Fang, H.Y.; Pan, W.J.; Pan, H.C. The complete mitochondrial genome of Chinese pond mussel Sinanodonta woodiana (Unionoida: Unionidae). Mitochondrial DNA Part A 2016, 27, 1620–1621. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | COI | 28S | Country | Voucher | Studies for COI/28S |
|---|---|---|---|---|---|
| INGROUP | |||||
| Amuranodonta kijaensis | MK574204 | MK574473 | Russia | RMBH Biv_228 | Bolotov et al., 2020 [15] |
| Anemina arcaeformis | MG462936 | MG595463 | China | NCU 16_XPWU_SU026 | Huang et al., 2019 [17] |
| Beringiana beringiana | MT020557 | MT020799 | Japan | CIIMAR BIV2700 | Lopes-Lima et al., 2020 [18] |
| Beringiana japonica | MT020576 | MT020803 | Japan | MHS SJSaitNam4 | Lopes-Lima et al., 2020 [18] |
| Beringiana fukuharai | MT020567 | MT020801 | Japan | OMNH Mo 39101 | Lopes-Lima et al., 2020 [18] |
| Beringiana gosannensis | MT020584 | MT020802 | Japan | OMNH Mo 39089 | Lopes-Lima et al., 2020 [18] |
| Buldowskia flavotincta | MT020537 | MT020804 | South Korea | NNIBR IV29232 | Lopes-Lima et al., 2020 [18] |
| Buldowskia suifunica | MK574190 | MK574460 | Russia | RMBH biv225_11 | Bolotov et al., 2020 [15] |
| Buldowskia iwakawai | MT020523 | MT020806 | Japan | MHS AAfk080 | Lopes-Lima et al., 2020 [18] |
| Buldowskia kamiyai | MT020525 | MT020808 | Japan | OMNH Mo 39097 | Lopes-Lima et al., 2020 [18] |
| Buldowskia shadini | MK574197 | MK574467 | Russia | RMBH Biv_228_16 | Bolotov et al., 2020 [15] |
| Cristaria bellua | ON704642 | ON695893 | Laos | RMBH Biv813_1 | Konopleva et al., 2023 [38] |
| Cristaria clessini | MT020592 | MT020810 | Japan | MHS MCp-sigB | Lopes-Lima et al., 2020 [18] |
| Cristaria plicata | MG462956 | MG595484 | China | NCU 16_XPWU_SU047 | Huang et al., 2019 [17] |
| Cristaria truncata | OP491287 | OP499826 | Vietnam | IEBR FM CT01 | Konopleva et al., 2023 [38] |
| Pletholophus honglinhensis sp. nov. | SUBMITTED | SUBMITTED | Vietnam | NCSM 47211.01 | THIS STUDY |
| Pletholophus tenuis | KX822658 | KX822614 | Vietnam | NCSM 84924.01 | Lopes-Lima et al., 2017 [39] |
| Sinanodonta angula | MG463053 | MG595580 | China | 16 NCU XPWU SU143 | Huang et al., 2019 [17] |
| Sinanodonta calipygos | MT020623 | MT020833 | Japan | MHS SC_Shiga_4 | Lopes-Lima et al., 2020 [18] |
| Sinanodonta cf. elliptica | MG463058 | MG595585 | China | 16 NCU XPWU SU148 | Huang et al., 2019 [17] |
| Sinanodonta jourdyi | SUBMITTED | SUBMITTED | Vietnam | NCSM 84916.02 | THIS STUDY |
| Sinanodonta lauta | MT020616 | MT020834 | Japan | MHS SL_Shizuoka_1 | Lopes-Lima et al., 2020 [18] |
| Sinanodonta lucida | MG463066 | MG595589 | China | NCU 16_XPWU_SU152 | Huang et al., 2019 [17] |
| Sinanodonta schrenkii | MT020618 | MT020837 | South Korea | NNIBR IV29271 | Lopes-Lima et al., 2020 [18] |
| Sinanodonta tumens | MT020622 | MT020838 | Japan | MHS fk156 | Lopes-Lima et al., 2020 [18] |
| Sinanodonta cf. woodiana 1 | MG463052 | MG595599 | China | NCU 16_XPWU_SU162 | Huang et al., 2019 [17] |
| Sinanodonta cf. woodiana 2 | MG463080 | MG595608 | China | NCU 16_XPWU_SU171 | Huang et al., 2019 [17] |
| OUTGROUP | |||||
| Anodonta cygnea | KX822633 | KX822589 | Italy | Lopes-Lima et al., 2017 [39] | |
| Schistodesmus lampreyanus | MG463037 | MG595570 | China | NCU 16_XPWU_SU133 | Huang et al., 2019 [17] |
| Margaritifera laevis | MT020707 | MT020823 | Japan | MHS MMt-tc01 | Lopes-Lima et al., 2020 [18] |
| Gibbosula crassa | MH293546 | MH293542 | Vietnam | IEBR FM GC01 | Lopes-Lima et al., 2018 [5] |
| Dataset | Sequences (New) | Haplotypes | Size (nt) | Models | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| COI Codon 1 | COI Codon 2 | COI Codon 3 | 28S | ||||||||
| BI | ML | BI | ML | BI | ML | BI | ML | ||||
| COI + 28S | |||||||||||
| Cristariini | 31 (2) | -- | 1029 | GTR + G | TN + F + G4 | F81 + I | F81 + F | HKY + G | TN + F + R2 | GTR | TPM3u + F |
| COI | |||||||||||
| Cristaria | 130 | 89 | 615 | K2P + I | TNe + G4 | F81 + I | F81 + F + I | HKY | HKY + F | ----- | ----- |
| Pletholophus | 16 (3) | 8 | 658 | HKY + I | TN + F + G4 | F81 + I | F81 + F | HKY | HKY + F | ----- | ----- |
| Sinanodonta | 250 (9) | 84 | 609 | GTR + G | TN + F + G4 | F81 + I | F81 + F | HKY + G | TPM3u + F + G4 | ----- | ----- |
| Cristaria | ||||||||||
| C. bellua | C. clessini | C. plicata | Cristaria sp. | C. truncata | ||||||
| C. bellua | 0.011 | 0.011 | 0.014 | 0.010 | ||||||
| C. clessini | 0.082 | 0.010 | 0.003 | 0.010 | ||||||
| C. plicata | 0.087 | 0.080 | 0.011 | 0.008 | ||||||
| Cristaria sp. | 0.119 | 0.115 | 0.100 | 0.013 | ||||||
| C. truncata | 0.086 | 0.077 | 0.044 | 0.112 | ||||||
| Pletholophus | P. honglinhensis | P. reinianus | P. tenuis | |||||||
| P. honglinhensis | 0.014 | 0.013 | ||||||||
| P. reinianus | 0.105 | 0.009 | ||||||||
| P. tenuis | 0.109 | 0.050 | ||||||||
| Sinanodonta | S. angula | S. calipygos | S. cf. elliptica | S. jourdyi | S. lauta | S. lucida | S. schrenkii | S. tumens | S. woodiana 1 | S. woodiana 2 |
| S. angula | 0.012 | 0.013 | 0.012 | 0.012 | 0.009 | 0.013 | 0.012 | 0.012 | 0.012 | |
| S. calipygos | 0.114 | 0.012 | 0.012 | 0.010 | 0.014 | 0.012 | 0.010 | 0.010 | 0.010 | |
| S. cf. elliptica | 0.137 | 0.087 | 0.009 | 0.008 | 0.013 | 0.012 | 0.010 | 0.008 | 0.008 | |
| S. jourdyi | 0.128 | 0.088 | 0.049 | 0.009 | 0.013 | 0.011 | 0.011 | 0.009 | 0.008 | |
| S. lauta | 0.127 | 0.078 | 0.048 | 0.056 | 0.013 | 0.012 | 0.010 | 0.005 | 0.008 | |
| S. lucida | 0.076 | 0.131 | 0.132 | 0.130 | 0.133 | 0.013 | 0.013 | 0.014 | 0.013 | |
| S. schrenkii | 0.120 | 0.081 | 0.093 | 0.089 | 0.105 | 0.127 | 0.011 | 0.012 | 0.011 | |
| S. tumens | 0.119 | 0.077 | 0.103 | 0.106 | 0.098 | 0.133 | 0.100 | 0.010 | 0.010 | |
| S. woodiana 1 | 0.131 | 0.078 | 0.047 | 0.060 | 0.032 | 0.133 | 0.099 | 0.106 | 0.008 | |
| S. woodiana 2 | 0.117 | 0.080 | 0.045 | 0.046 | 0.049 | 0.123 | 0.087 | 0.094 | 0.051 |
| Types | Shell Length, mm | Shell Height, mm | Shell Width mm |
|---|---|---|---|
| Holotype NCSM 47211 | 74.5 | 41.4 | 26.4 |
| Paratype NCSM 103062.2 | 77.5 | 44.3 | 26.2 |
| Paratype NCSM 103062.3 | 90.6 | 47.6 | 28.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bogan, A.E.; Do, V.T.; Froufe, E.; Lopes-Lima, M. The Anodontini of Vietnam (Mollusca: Bivalvia: Unionidae: Unioninae) with the Description of a New Species. Diversity 2023, 15, 710. https://doi.org/10.3390/d15060710
Bogan AE, Do VT, Froufe E, Lopes-Lima M. The Anodontini of Vietnam (Mollusca: Bivalvia: Unionidae: Unioninae) with the Description of a New Species. Diversity. 2023; 15(6):710. https://doi.org/10.3390/d15060710
Chicago/Turabian StyleBogan, Arthur E., Van Tu Do, Elsa Froufe, and Manuel Lopes-Lima. 2023. "The Anodontini of Vietnam (Mollusca: Bivalvia: Unionidae: Unioninae) with the Description of a New Species" Diversity 15, no. 6: 710. https://doi.org/10.3390/d15060710
APA StyleBogan, A. E., Do, V. T., Froufe, E., & Lopes-Lima, M. (2023). The Anodontini of Vietnam (Mollusca: Bivalvia: Unionidae: Unioninae) with the Description of a New Species. Diversity, 15(6), 710. https://doi.org/10.3390/d15060710