
The Impact of Breeding Yellow-Legged Gulls on Vegetation Cover and Plant Composition of Grey Dune Habitats




Abstract
1. Introduction
2. Materials and Methods
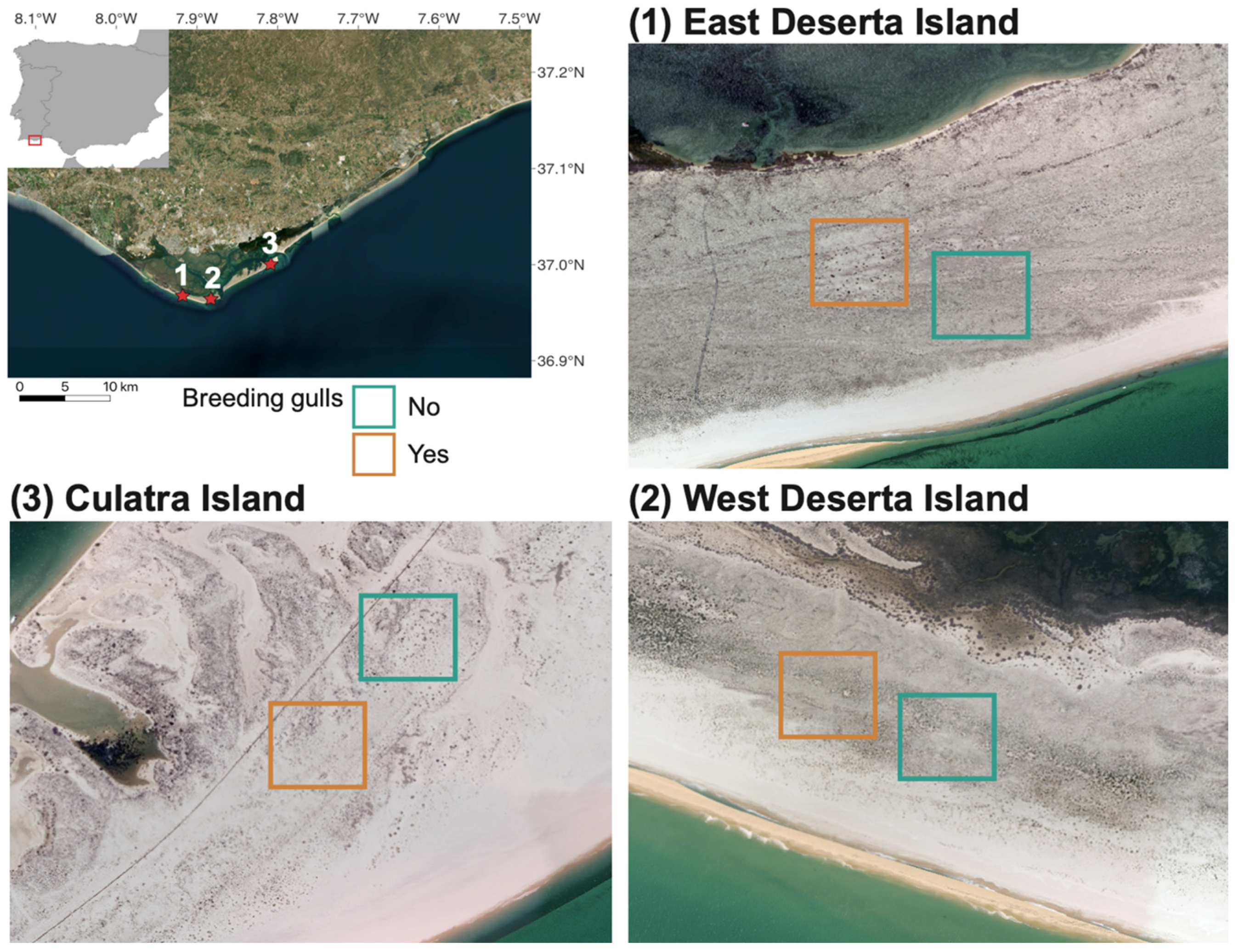
2.1. Study Area
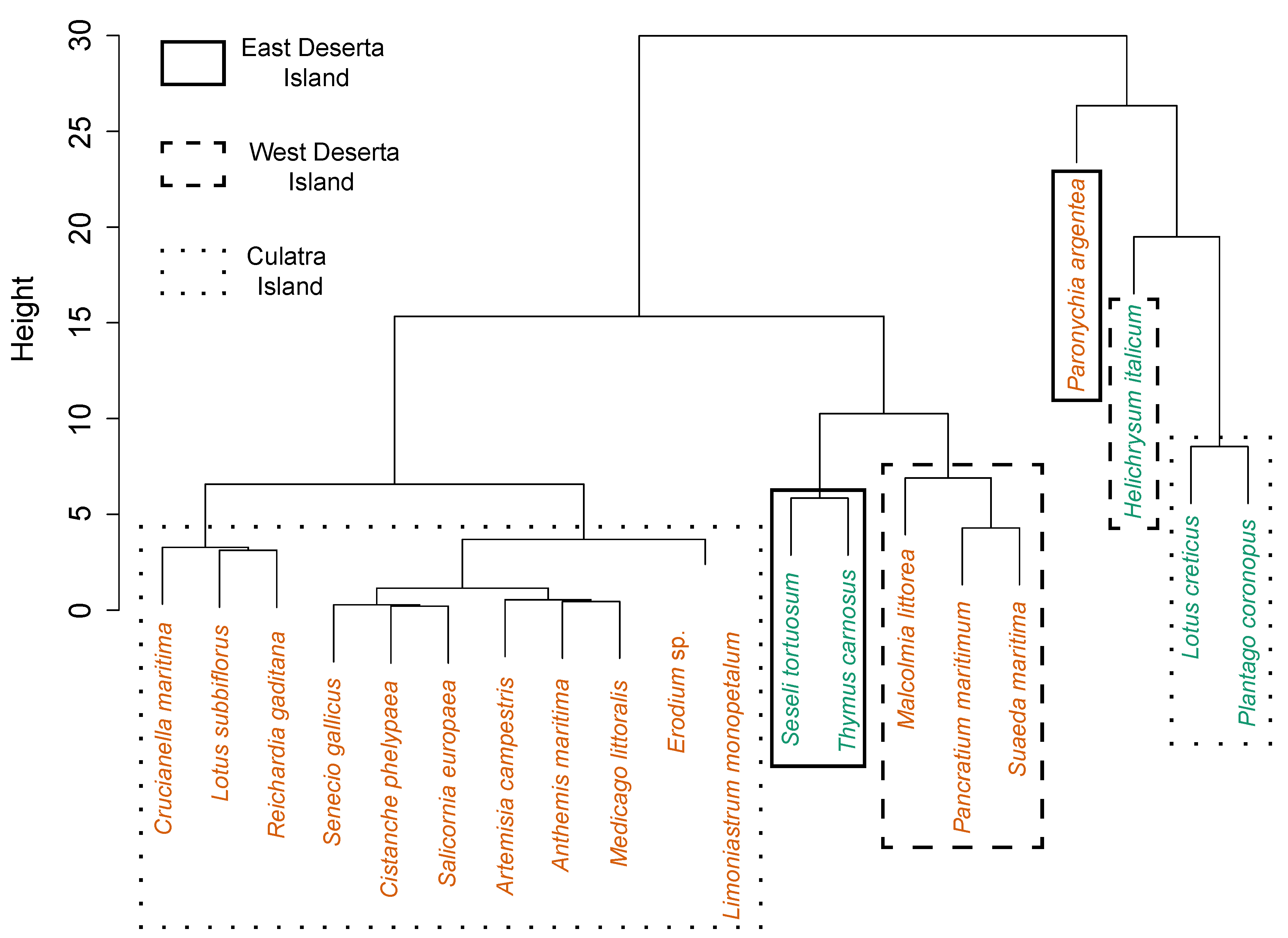
2.2. Vegetation Cover and Plant Species Composition
2.3. Statistical Analysis
3. Results
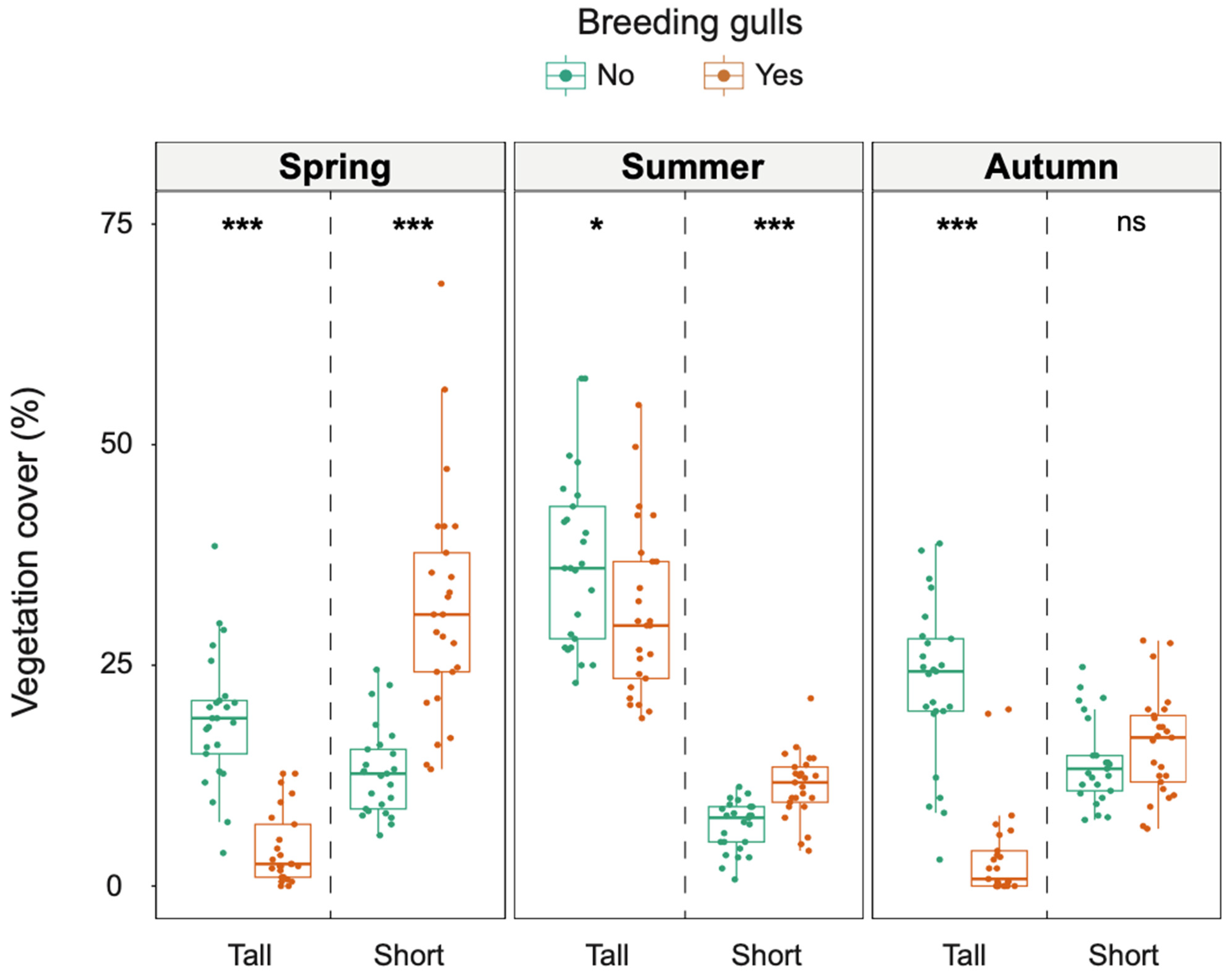
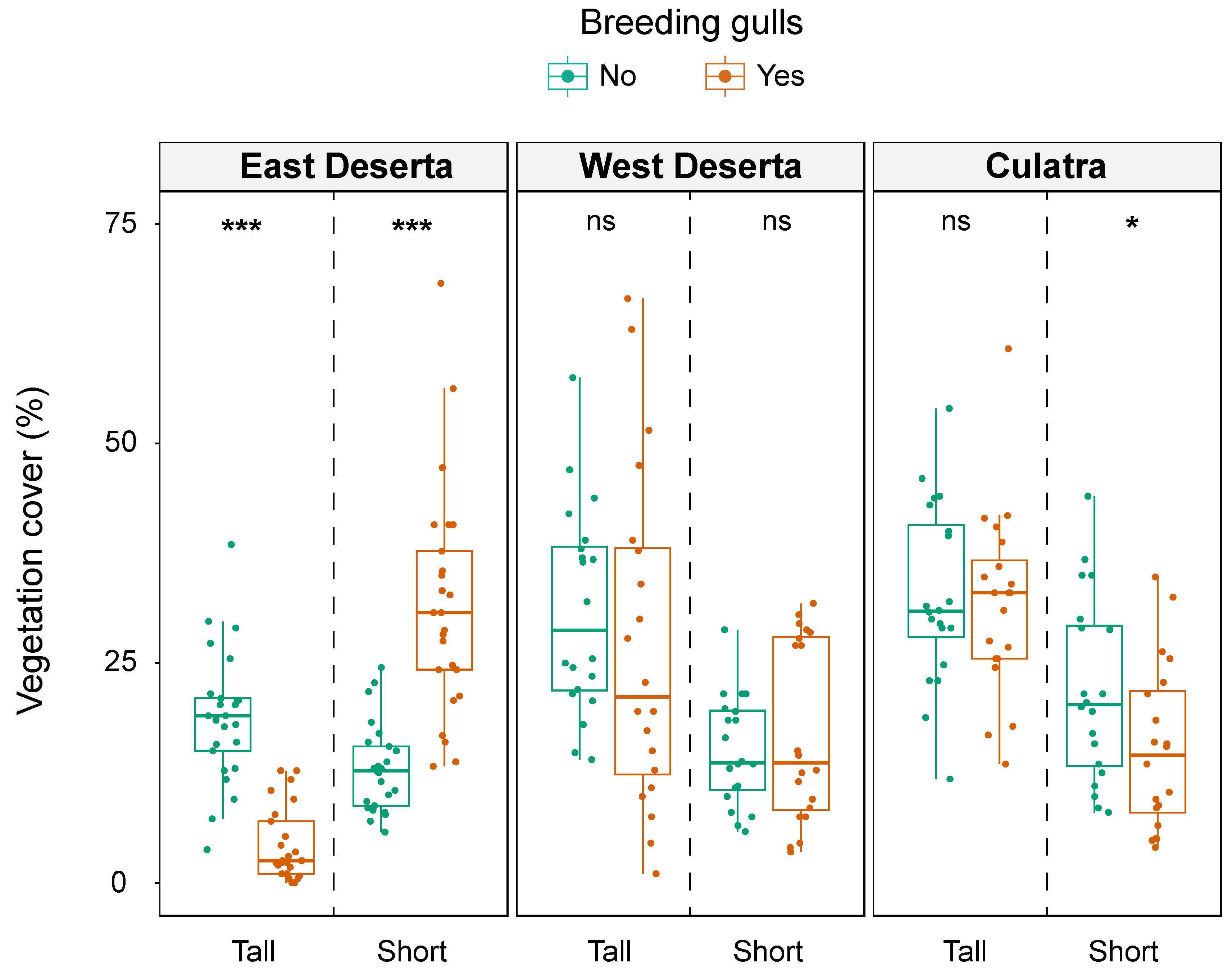
3.1. Vegetation Cover in Old and New Colony Sites
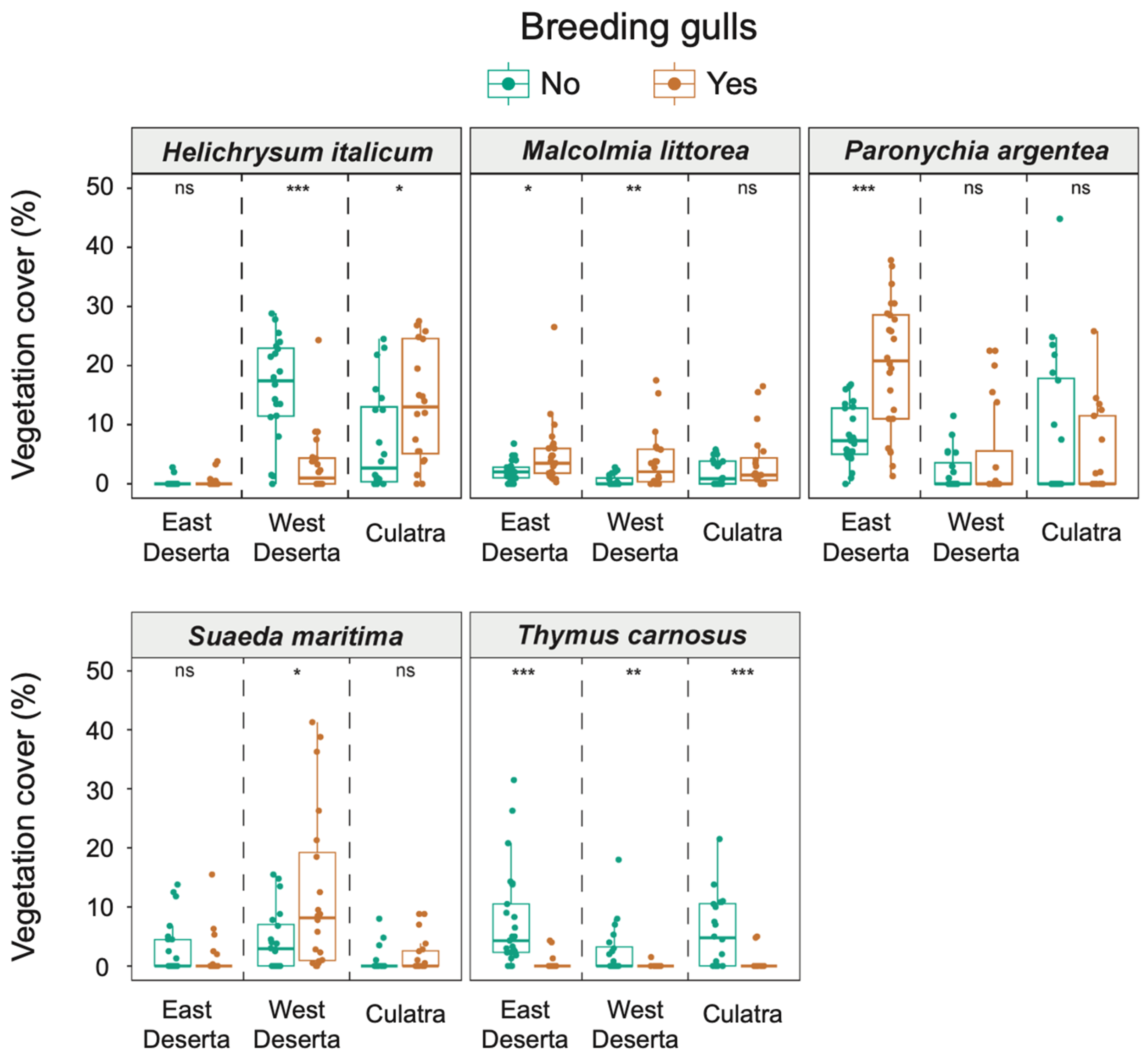
3.2. Cover of Plant Species in Old and New Colony Sites
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evans, K.L.; Gaston, K.J.; Sharp, S.P.; McGowan, A.; Simeoni, M.; Hatchwell, B.J. Effects of Urbanisation on Disease Prevalence and Age Structure in Blackbird Turdus merula Populations. Oikos 2009, 118, 774–782. [Google Scholar] [CrossRef]
- Fuirst, M.; Veit, R.R.; Hahn, M.; Dheilly, N.; Thorne, L.H. Effects of Urbanization on the Foraging Ecology and Microbiota of the Generalist Seabird Larus argentatus. PLoS ONE 2018, 13, e0209200. [Google Scholar] [CrossRef]
- Samia, D.S.M.; Nakagawa, S.; Nomura, F.; Rangel, T.F.; Blumstein, D.T. Increased Tolerance to Humans among Disturbed Wildlife. Nat. Commun. 2015, 6, 8877. [Google Scholar] [CrossRef]
- de Faria, J.P.; Paiva, V.H.; Veríssimo, S.; Gonçalves, A.M.M.; Ramos, J.A. Seasonal Variation in Habitat Use, Daily Routines and Interactions with Humans by Urban-Dwelling Gulls. Urban Ecosyst. 2021, 24, 1101–1115. [Google Scholar] [CrossRef]
- French, S.S.; Webb, A.C.; Hudson, S.B.; Virgin, E.E. Town and Country Reptiles: A Review of Reptilian Responses to Urbanization. Integr. Comp. Biol. 2018, 58, 948–966. [Google Scholar] [CrossRef]
- Mikula, P.; Hromada, M.; Albrecht, T.; Tryjanowski, P. Nest Site Selection and Breeding Success in Three Turdus Thrush Species Coexisting in an Urban Environment. Acta Ornithol. 2014, 49, 83–92. [Google Scholar] [CrossRef]
- Spelt, A.; Williamson, C.; Shamoun-Baranes, J.; Shepard, E.; Rock, P.; Windsor, S. Habitat Use of Urban-Nesting Lesser Black-Backed Gulls during the Breeding Season. Sci. Rep. 2019, 9, 10527. [Google Scholar] [CrossRef] [PubMed]
- De Faria, J.P.; Paiva, V.H.; Veríssimo, S.N.; Lopes, C.S.; Soares, R.; Oliveira, J.; dos Santos, I.; Norte, A.C.; Ramos, J.A. Plenty of Rooftops with Few Neighbours Occupied by Young Breeding Yellow-Legged Gulls (Larus michahellis): Does This Occur at the Expense of Their Health Condition? IBIS 2023, 165, 312–321. [Google Scholar] [CrossRef]
- de Faria, J.P.; Lopes, C.S.; Louise, E.K.; Blight, K.; Nager, R.G. Urban Gulls with Humans. In Seabird Biodiversity and Human Activities; Ramos, J.A., Pereira, L., Eds.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Oxfordshire, UK, 2022; pp. 90–105. ISBN 9781003047520. [Google Scholar]
- Spelt, A.; Soutar, O.; Williamson, C.; Memmott, J.; Shamoun-Baranes, J.; Rock, P.; Windsor, S. Urban Gulls Adapt Foraging Schedule to Human-Activity Patterns. IBIS 2021, 163, 274–282. [Google Scholar] [CrossRef]
- Vidal, E.; Medail, F.; Tatoni, T. Is the Yellow-Legged Gull a Superabundant Bird Species in the Mediterranean? Impact on Fauna and Flora, Conservation Measures and Research Priorities. Biodivers. Conserv. 1998, 7, 1013–1026. [Google Scholar] [CrossRef]
- Migura-Garcia, L.; Ramos, R.; Cerdà-Cuéllar, M. Antimicrobial Resistance of Salmonella serovars and Campylobacter Spp. Isolated from an Opportunistic Gull Species, Yellow-Legged Gull (Larus michahellis). J. Wildl. Dis. 2017, 53, 148–152. [Google Scholar] [CrossRef]
- Arcos, J.M.; Oro, D.; Sol, D. Competition between the Yellow-Legged Gull Larus cachinnans and Audouin’s Gull Larus audouinii Associated with Commercial Fishing Vessels: The Influence of Season and Fishing Fleet. Mar. Biol. 2001, 139, 807–816. [Google Scholar] [CrossRef]
- Belant, J.L. Gulls in Urban Environments: Landscape-Level Management to Reduce Conflict. Landsc. Urban Plan. 1997, 38, 245–258. [Google Scholar] [CrossRef]
- Baumberger, T.; Affre, L.; Torre, F.; Vidal, E.; Dumas, P.J.; Tatoni, T. Plant Community Changes as Ecological Indicator of Seabird Colonies’ Impacts on Mediterranean Islands. Ecol. Indic. 2012, 15, 76–84. [Google Scholar] [CrossRef]
- De La Peña-Lastra, S.; Torre, F.; Carballeira, R.; Santiso, M.J.; Pérez-Alberti, A.; Otero, X.L. The Rapid Effects of Yellow-Legged Gull (Larus michahellis) Colony on Dune Habitats and Plant Landscape in the Atlantic Islands National Park (NW Spain). Land 2022, 11, 258. [Google Scholar] [CrossRef]
- Thomas, G.J. A Review of Gull Damage and Management Methods at Nature Reserves. Biol. Conserv. 1972, 4, 117–127. [Google Scholar] [CrossRef]
- Vidal, E.; Médail, F.; Tatoni, T.; Bonnet, V. Seabirds Drive Plant Species Turnover on Small Mediterranean Islands at the Expense of Native Taxa. Oecologia 2000, 122, 427–434. [Google Scholar] [CrossRef]
- Otero, X.L.; de la Peña-Lastra, S.; Romero, D.; Nobrega, G.N.; Ferreira, T.O.; Pérez-Alberti, A. Trace Elements in Biomaterials and Soils from a Yellow-Legged Gull (Larus michahellis) Colony in the Atlantic Islands of Galicia National Park (NW Spain). Mar. Pollut. Bull. 2018, 133, 144–149. [Google Scholar] [CrossRef]
- Otero, X.L.; Fernández-Balado, C.; Ferreira, T.O.; Pérez-Alberti, A.; Revilla, G. Soil Eutrophication in Seabird Colonies Affects Cell Wall Composition: Implications for the Conservation of Rare Plant Species. Mar. Pollut. Bull. 2021, 168, 112469. [Google Scholar] [CrossRef]
- Otero Pérez, X.L. Effects of Nesting Yellow-Legged Gulls (Larus cachinnans Pallas) on the Heavy Metal Content of Soils in the Cies Islands (Galicia, North-West Spain). Mar. Pollut. Bull. 1998, 36, 267–272. [Google Scholar] [CrossRef]
- Otero, X.L.; Tejada, O.; Martín-Pastor, M.; De La Peña, S.; Ferreira, T.O.; Pérez-Alberti, A. Phosphorus in Seagull Colonies and the Effect on the Habitats. The Case of Yellow-Legged Gulls (Larus michahellis) in the Atlantic Islands National Park (Galicia-NW Spain). Sci. Total Environ. 2015, 532, 383–397. [Google Scholar] [CrossRef]
- Battisti, C. Heterogeneous composition of anthropogenic litter recorded in nests of Yellow-legged gull (Larus michahellis) from a small Mediterranean island. Mar. Pollut. Bull. 2020, 150, 110682. [Google Scholar] [CrossRef]
- Battisti, C.; Fanelli, G. Alien-dominated plant communities’ syntopic with seabird’s nests: Evidence and possible implication from a Mediterranean insular ecosystem. Ethol. Ecol. Evol. 2021, 33, 543–552. [Google Scholar] [CrossRef]
- Hogg, E.H.; Morton, J.K. The Effects of Nesting Gulls on the Vegetation and Soil of Islands in the Great Lakes. Can. J. Bot. 1983, 61, 3240–3254. [Google Scholar] [CrossRef]
- Tozato, H.C. Gestão Da Biodiversidade Na União Europeia: O Programa Natura 2000 Como Instrumento Para o Alcance Da Meta 11 de Aichi. Rev. Gestão Políticas Públicas 2016, 6, 163–186. [Google Scholar] [CrossRef]
- Ceia, F.R.; Patrício, J.; Marques, J.C.; Dias, J.A. Coastal Vulnerability in Barrier Islands: The High Risk Areas of the Ria Formosa (Portugal) System. Ocean. Coast. Manag. 2010, 53, 478–486. [Google Scholar] [CrossRef]
- Matos, D.M.; Ramos, J.A.; Calado, J.G.; Ceia, F.R.; Hey, J.; Paiva, V.H. How fishing intensity affects the spatial and trophic ecology of two gull species breeding in sympatry. ICES J. Mar. Sci. 2018, 75, 1949–1964. [Google Scholar] [CrossRef]
- Calado, J.G.; Verissimo, S.N.; Paiva, V.H.; Ramos, R.; Vaz, P.T.; Matos, D.M.; Pereira, J.M.; Lopes, C.; Oliveira, N.; Quaresma, A.; et al. Influence of fisheries on the spatio-temporal feeding ecology of gulls along the western Iberian coast. Mar. Ecol. Prog. Ser. 2021, 661, 187–201. [Google Scholar] [CrossRef]
- Ben-Shachar, M.; Lüdecke, D.; Makowski, D. Effectsize: Estimation of Effect Size Indices and Standardized Parameters. J. Open Source Softw. 2020, 5, 2815. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions, R package version 2.1.4; R Foundation: Indianapolis, IN, USA, 2022. [Google Scholar]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- García, L.V.; Marañón, T.; Ojeda, F.; Clemente, L.; Redondo, R. Seagull Influence on Soil Properties, Chenopod Shrub Distribution, and Leaf Nutrient Status in Semi-Arid Mediterranean Islands. Oikos 2002, 98, 75–86. [Google Scholar] [CrossRef]
- Duda, M.P.; Glew, J.R.; Michelutti, N.; Robertson, G.J.; Montevecchi, W.A.; Kissinger, J.A.; Eickmeyer, D.C.; Blais, J.M.; Smol, J.P. Long-Term Changes in Terrestrial Vegetation Linked to Shifts in a Colonial Seabird Population. Ecosystems 2020, 23, 1643–1656. [Google Scholar] [CrossRef]
- Mochalova, O.A.; Khoreva, M.G. Changes in the Vegetation Cover of Cape Ostrovnoy (Gulf of Tauisk, the Sea of Okhotsk) under the Influence of Colonial Seabirds. Contemp. Probl. Ecol. 2013, 6, 57–64. [Google Scholar] [CrossRef]
- Costa, J.C.; Lousã, M.; Espírito-Santo, M.D. A Vegetação Do Parque Natural Da Ria Formosa (Algarve, Portugal). Stud. Bot. 1996, 15, 69–157. [Google Scholar]
- Stevens, C.J.; Dise, N.B.; Mountford, J.O.; Gowing, D.J. Impact of Nitrogen Deposition on the Species Richness of Grasslands. Science 2004, 303, 1876–1879. [Google Scholar] [CrossRef]
- Ellis, J.C. Marine Birds on Land: A Review of Plant Biomass, Species Richness, and Community Composition in Seabird Colonies. Plant Ecol. 2005, 181, 227–241. [Google Scholar] [CrossRef]
- Talavera, L.; Costas, S.; Ferreira, Ó. A New Index to Assess the State of Dune Vegetation Derived from True Colour Images. Ecol. Indic. 2022, 137, 108770. [Google Scholar] [CrossRef]
- Martín-Vélez, V.; Montalvo, T.; Afán, I.; Sánchez-Márquez, A.; Aymí, R.; Figuerola, J.; Lovas-Kiss, Á.; Navarro, J. Gulls Living in Cities as Overlooked Seed Dispersers within and Outside Urban Environments. Sci. Total Environ. 2022, 823, 153535. [Google Scholar] [CrossRef]
- Signa, G.; Mazzola, A.; Vizzini, S. Effects of a Small Seagull Colony on Trophic Status and Primary Production in a Mediterranean Coastal System (Marinello Ponds, Italy). Estuar. Coast. Shelf Sci. 2012, 111, 27–34. [Google Scholar] [CrossRef]
- Sigurdsson, B.D.; Magnusson, B. Effects of Seagulls on Ecosystem Respiration, Soil Nitrogen and Vegetation Cover on a Pristine Volcanic Island, Surtsey, Iceland. Biogeosciences 2010, 7, 883–891. [Google Scholar] [CrossRef]
- Bukacinski, D.; Rutkowska, A.; Bukacinska, M. The Effect of Nesting Black-Headed Gulls (Larus ridibundus L.) on the Soil and Vegetation of a Vistual River Island, Poland. Ann. Bot. Fenn. 1994, 31, 233–243. [Google Scholar]
- Sanchez-Pinero, F.; Polis, G.A. Bottom-up Dynamics of Allochthonous Input: Direct and Indirect Effects of Seabirds on Islands. Ecology 2000, 81, 3117–3132. [Google Scholar] [CrossRef]
- De la Peña-Lastra, S.; Affre, L.; Otero, X.L. Soil Nutrient Dynamics in Colonies of the Yellow-Legged Seagull (Larus michahellis) in Different Biogeographical Zones. Geoderma 2020, 361, 114109. [Google Scholar] [CrossRef]
- Kim, S.Y.; Monaghan, P. Effects of Vegetation on Nest Microclimate and Breeding Performance of Lesser Black-Backed Gulls (Larus fuscus). J. Ornithol. 2005, 146, 176–183. [Google Scholar] [CrossRef]
- Parsons, K.C.; Chao, J. Nest Cover and Chick Survival in Herring Gulls (Larus argentatus). Colonial Waterbirds 1983, 6, 154. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Presence of Breeding Gulls | Mann–Whitney U Test | Effect Size | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Vegetation Cover | Yes | No | Z-Value | p-Value | Effect | Index | CI 95% | ||
| Season | Spring | Tall | 4.3 ± 4.2 | 18.9 ± 7.5 | 5.61 | <0.001 | Yes < No | 0.795 | 0.70–0.85 |
| Short | 31.6 ± 13.0 | 13.0 ± 5.0 | 5.35 | <0.001 | Yes > No | 0.758 | 0.63–0.83 | ||
| Summer | Tall | 31.1 ± 9.7 | 37.0 ± 9.8 | 2.08 | 0.037 | Yes < No | 0.295 | 0.04–0.53 | |
| Short | 11.4 ± 3.4 | 6.8 ± 2.8 | 4.25 | <0.001 | Yes > No | 0.603 | 0.40–0.78 | ||
| Autumn | Tall | 3.5 ± 5.5 | 22.9 ± 9.2 | 5.66 | <0.001 | Yes < No | 0.802 | 0.72–0.86 | |
| Short | 16.1 ± 5.9 | 14.1 ± 4.8 | 1.08 | 0.281 | 0.154 | 0.01–0.44 | |||
| Site | East Deserta Island | Tall | 4.3 ± 4.2 | 18.9 ± 7.5 | 5.61 | <0.001 | Yes < No | 0.795 | 0.70–0.85 |
| Short | 31.6 ± 13.0 | 13.0 ± 5.0 | 5.35 | <0.001 | Yes > No | 0.758 | 0.64–0.83 | ||
| West Deserta Island | Tall | 26.9 ± 19.0 | 31.0 ± 11.7 | 1.22 | 0.224 | 0.195 | 0.01–0.49 | ||
| Short | 17.1 ± 10.4 | 15.0 ± 6.2 | 0.30 | 0.766 | 0.050 | 0.01–0.41 | |||
| Culatra Island | Tall | 31.8 ± 10.6 | 32.7 ± 10.3 | 0.16 | 0.871 | 0.195 | 0.02–0.50 | ||
| Short | 15.3 ± 9.4 | 21.9 ± 10.5 | 2.03 | 0.042 | Yes < No | 0.050 | 0.01–0.39 | ||
| Species | East Deserta Island | Paronychia argentea | 21.4 ± 12.7 | 8.1 ± 4.9 | 3.78 | <0.001 | Yes > No | 0.537 | 0.29–0.73 |
| Malcolmia littorea | 4.9 ± 5.4 | 2.1 ± 1.7 | 2.38 | 0.017 | Yes > No | 0.338 | 0.08–0.56 | ||
| Helichrysum italicum | 0.3 ± 1.0 | 0.2 ± 0.7 | 0.84 | 0.399 | 0.122 | 0.01–0.36 | |||
| Thymus carnosus | 0.4 ± 1.2 | 7.6 ± 8.4 | 5.21 | <0.001 | Yes < No | 0.738 | 0.56–0.88 | ||
| Suaeda maritima | 1.3 ± 3.4 | 2.5 ± 4.3 | 1.02 | 0.309 | 0.146 | 0.01–0.41 | |||
| West Deserta Island | Paronychia argentea | 4.9 ± 8.5 | 2.1 ± 3.4 | 0.11 | 0.913 | 0.020 | 0.01–0.35 | ||
| Malcolmia littorea | 3.9 ± 5.0 | 0.6 ± 0.9 | 2.94 | 0.001 | Yes > No | 0.467 | 0.19–0.70 | ||
| Helichrysum italicum | 3.5 ± 5.8 | 16.5 ± 8.7 | 4.06 | <0.001 | Yes < No | 0.644 | 0.42–0.82 | ||
| Thymus carnosus | 0.1 ± 0.3 | 2.5 ± 4.4 | 2.95 | 0.003 | Yes < No | 0.470 | 0.22–0.66 | ||
| Suaeda maritima | 12.2 ± 13.7 | 4.4 ± 5.2 | 1.99 | 0.047 | Yes > No | 0.317 | 0.03–0.56 | ||
| Culatra Island | Paronychia argentea | 5.0 ± 7.4 | 8.7 ± 12.7 | 0.24 | 0.810 | 0.040 | 0.01–0.38 | ||
| Malcolmia littorea | 3.7 ± 5.0 | 2.1 ± 2.2 | 0.85 | 0.393 | 0.137 | 0.01–0.45 | |||
| Helichrysum italicum | 13.5 ± 9.7 | 7.3. ± 8.7 | 2.22 | 0.026 | Yes > No | 0.354 | 0.05–0.61 | ||
| Thymus carnosus | 0.5 ± 1.5 | 5.8 ± 6.1 | 3.61 | <0.001 | Yes < No | 0.573 | 0.33–0.77 | ||
| Suaeda maritima | 1.8 ± 3.0 | 0.9 ± 2.1 | 1.30 | 0.193 | 0.208 | 0.01–0.52 | |||
| East Deserta Island | West Deserta Island | Culatra Island | ||||
|---|---|---|---|---|---|---|
| Species | Presence of Breeding Gulls | Presence of Breeding Gulls | Presence of Breeding Gulls | |||
| Yes | No | Yes | No | Yes | No | |
| Anthemis maritima | - | - | - | - | 0.3 ± 1.2 | 0.5 ± 2.2 |
| Artemisia campestris | 0.4 ± 2.0 | - | - | - | 0.5 ± 1.9 | 0.4 ± 1.5 |
| Cistanche phelypaea | - | - | - | - | - | 0.04 ± 0.2 |
| Crucianella maritima | - | 0.1 ± 0.4 | 0.3 ± 1.1 | 2.6 ± 3.3 | 3.7 ± 4.6 | 1.9 ± 3.3 |
| Erodium sp. | - | - | 4.3 ± 5.9 | 0.1 ± 0.3 | 0.7 ± 2.9 | 0.4 ± 1.1 |
| Helichrysum italicum | 0.3 ± 1.0 | 0.2 ± 0.7 | 3.5 ± 5.6 | 16.2 ± 8.5 | 13.5 ± 9.5 | 7.3 ± 8.4 |
| Limoniastrum monopetalum | - | - | - | - | 1.8 ± 7.0 | - |
| Lotus creticus | - | 0.8 ± 1.3 | 3.4 ± 3.8 | 4.1 ± 5.1 | 4.6 ± 6.3 | 14.6 ± 14.9 |
| Lotus subbiflorus | - | - | 2.2 ± 5.5 | 3.8 ± 7.03 | 0.9 ± 1.9 | - |
| Malcolmia littorea | 4.9 ± 5.3 | 2.1 ± 1.7 | 3.9 ± 4.9 | 0.6 ± 0.9 | 3.8 ± 4.9 | 2.1 ± 2.2 |
| Medicago littoralis | - | - | 0.1 ± 0.4 | - | - | 0.9 ± 1.7 |
| Pancratium maritimum | - | - | 5.3 ± 8.3 | 5.0 ± 7.1 | 1.2 ± 1.8 | 0.5 ± 1.3 |
| Paronychia argentea | 21.4 ± 12.5 | 8.1 ± 4.8 | 4.9 ± 8.3 | 2.1 ± 3.3 | 5.0 ± 7.2 | 8.4 ± 12.3 |
| Plantago coronopus | 0.6 ± 2.2 | - | 0.3 ± 0.7 | - | 3.3 ± 4.0 | 8.6 ± 9.7 |
| Reichardia gaditana | - | 1.0 ± 3.0 | 0.9 ± 1.5 | 1.5 ± 1.4 | 0.8 ± 1.0 | 1.8 ± 3.1 |
| Salicornia europaea | - | - | - | - | - | 0.2 ± 1.1 |
| Senecio gallicus | - | - | 0.3 ± 0.4 | - | - | - |
| Seseli tortuosum | - | 5.6 ± 6.0 | - | 0.3 ± 0.8 | - | 1.3 ± 3.0 |
| Suaeda maritima | 1.3 ± 3.3 | 2.5 ± 4.2 | 12.2 ± 13.4 | 4.4 ± 5.1 | 1.8 ± 2.9 | 0.9 ± 2.1 |
| Thymus carnosus | 0.4 ± 1.1 | 7.6 ± 8.2 | 0.1 ± 0.3 | 2.5 ± 4.3 | 0.5 ± 1.5 | 5.8 ± 6.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Portela, D.; Pereira, J.M.; Cerveira, L.R.; Paiva, V.H.; Ramos, J.A. The Impact of Breeding Yellow-Legged Gulls on Vegetation Cover and Plant Composition of Grey Dune Habitats. Diversity 2023, 15, 589. https://doi.org/10.3390/d15050589
Portela D, Pereira JM, Cerveira LR, Paiva VH, Ramos JA. The Impact of Breeding Yellow-Legged Gulls on Vegetation Cover and Plant Composition of Grey Dune Habitats. Diversity. 2023; 15(5):589. https://doi.org/10.3390/d15050589
Chicago/Turabian StylePortela, Diogo, Jorge M. Pereira, Lara R. Cerveira, Vitor H. Paiva, and Jaime A. Ramos. 2023. "The Impact of Breeding Yellow-Legged Gulls on Vegetation Cover and Plant Composition of Grey Dune Habitats" Diversity 15, no. 5: 589. https://doi.org/10.3390/d15050589
APA StylePortela, D., Pereira, J. M., Cerveira, L. R., Paiva, V. H., & Ramos, J. A. (2023). The Impact of Breeding Yellow-Legged Gulls on Vegetation Cover and Plant Composition of Grey Dune Habitats. Diversity, 15(5), 589. https://doi.org/10.3390/d15050589