

Phylogenetic Relationships in the Group Caespitosa of Paspalum L. (Poaceae, Panicoideae, Paspaleae)

 , and
, and 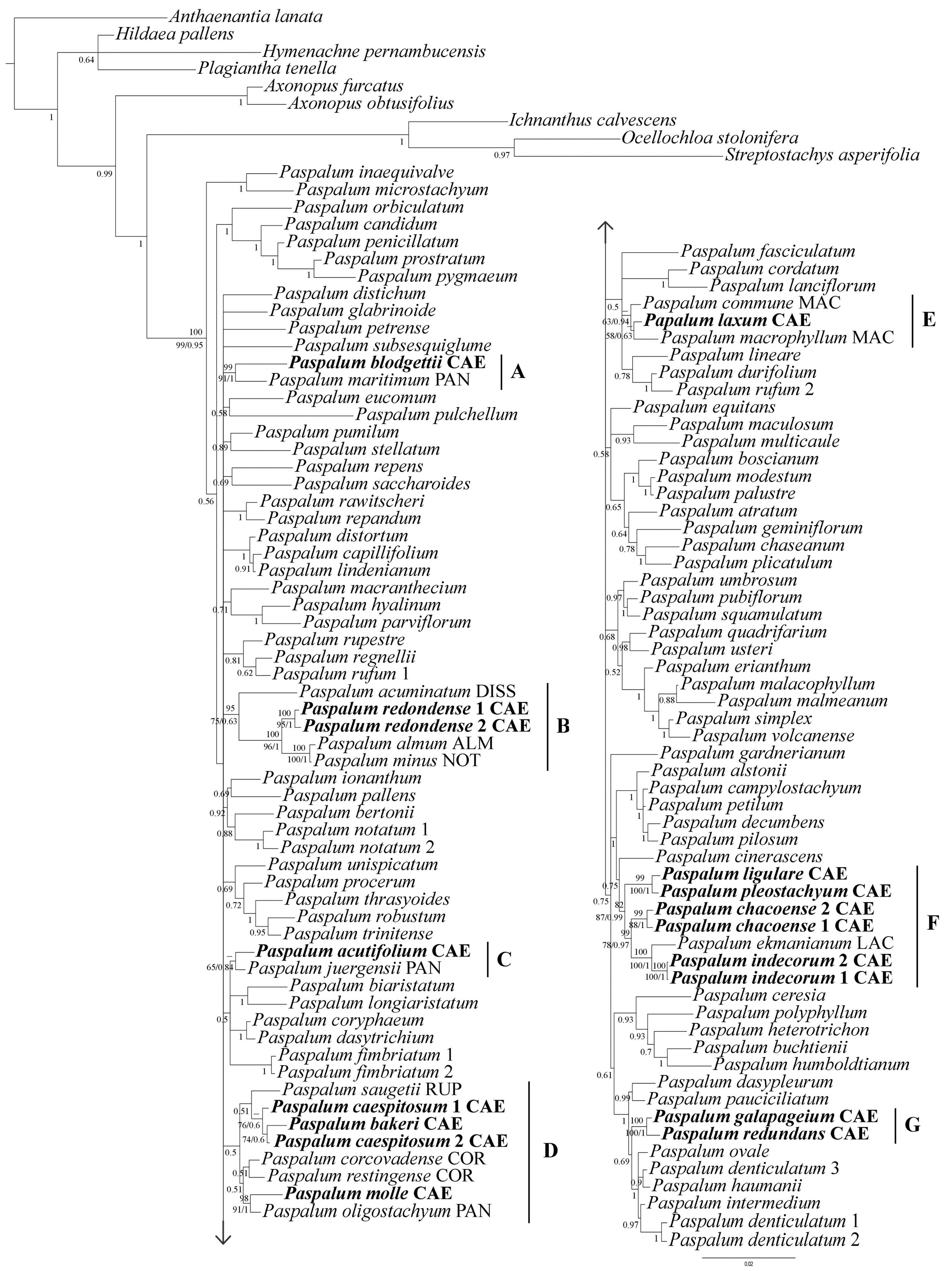{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Amplification and Sequencing
2.3. Phylogenetic Analyses
3. Results
4. Discussion
4.1. Major Results and Comparison with Previous Molecular Studies
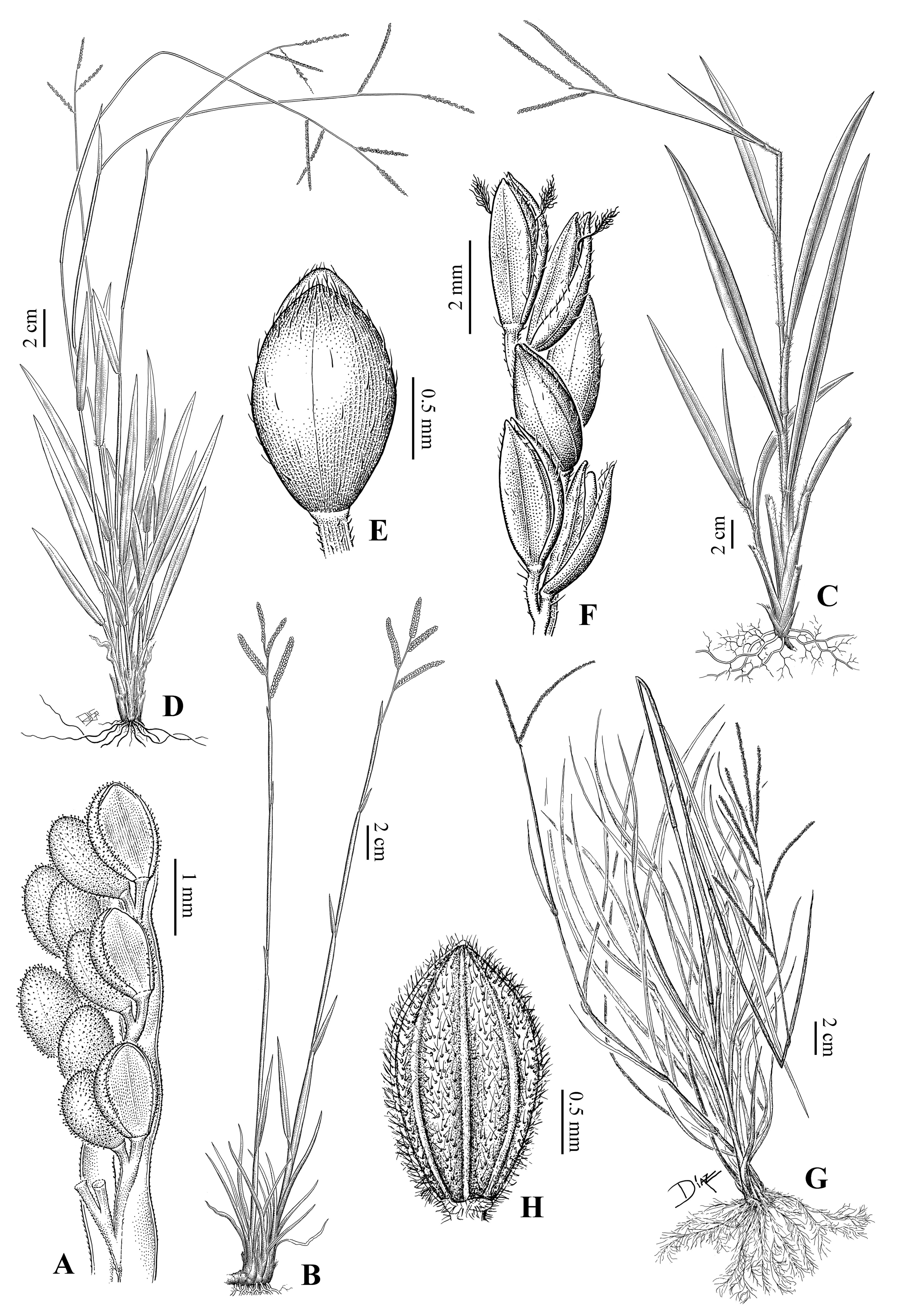
4.2. Relationships of Caespitosa Taxa


4.3. Needs for Future Molecular Studies in Paspalum
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuloaga, F.O.; Morrone, O. Revisión de las especies de Paspalum para América del Sur Austral (Argentina, Bolivia, sur de Brasil, Chile, Paraguay y Uruguay). Monogr. Syst. Bot. Mo. Bot. Gard. 2005, 102, 304. [Google Scholar] [CrossRef]
- Bennett, H.W. Dallisgrass, bahiagrass, and vaseygrass. In Forages: The Science of Grassland Agriculture; Hudges, H.D., Heath, M.E., Metcalfe, D.S., Eds.; The Iowa State University Press: Ames, IA, USA, 1962; pp. 281–285. [Google Scholar]
- Filguerias, T.S. Gramíneas forrageiras nativas do Distrito Federal, Brasil. Pesqui. Agropecu. Bras. 1992, 27, 1103–1111. [Google Scholar]
- Rua, G.H.; Arakaki, P.R.; Speranza, M.; Vaio, M. A phylogenetic analysis of the genus Paspalum (Poaceae) based on cpDNA and morphology. Plant Syst. Evol. 2010, 288, 227–243. [Google Scholar] [CrossRef]
- BFG (The Brazil Flora Group). Brazilian Flora 2020: Leveraging the power of a collaborative scientific network. Taxon 2021, 71, 178–198. [Google Scholar] [CrossRef]
- Nees von Esenbeck, C.G.D. III. Paspalus. In Flora Brasiliensis Seu Enumeratio Plantarum; Martius, C.F.P., Ed.; Sumptibus JG Cottage: Tuebingen, Germany, 1829; Volume 2, pp. 18–83. [Google Scholar]
- Döll, J.C. Gramineae. In Flora Brasiliensis; Martius, C.F.P., Eichler, A.W., Eds.; Frid. Fleischer: Germany, 1877; Volume 2, pp. 1–358. [Google Scholar]
- Chase, A. The North American species of Paspalum. Contr. U.S. Natl. Herb. 1929, 28, 1–310. [Google Scholar]
- Chase, A. Paspalum of South America; Smithsonian Institution, Hitchcock and Chase Library: Washington, DC, USA, 1939; unpublsihed manuscript. [Google Scholar]
- Pilger, R.K.F. Bemerkungen zur Systematik der Gattung Paspalum L. Repert. Spec. Nov. Regni Veg. 1929, 26, 228–231. [Google Scholar]
- Pilger, R.K.F. Gramineae III. Unterfamilie Panicoideae. In Die Natürlichen Pflanzenfamilien; Engler, A., Prantl, K., Eds.; W Engelmann: Leipzig, Germany, 1940; Volume 14e, pp. 01–208. [Google Scholar]
- Clayton, W.D.; Renvoize, S.A. Genera graminum. Grasses of the World. Kew Bull. Addit. Ser. 1986, 13, 1–389. [Google Scholar]
- Rodríguez, H. Pectinata chase ex Rodríguez, nueva sección en el género Paspalum L. (Gramineae). Ernstia 1992, 2, 21–23. [Google Scholar]
- Rodríguez, H. El subgénero Ceresia (Pers.) Reichenb. del género Paspalum L. (Gramineae) en Venezuela. Ernstia 1998, 8, 7–50. [Google Scholar]
- Morrone, O.; Denham, S.S.; Aliscioni, S.S.; Zuloaga, F.O. Revisión de las especies de Paspalum (Panicoideae: Paniceae), subgénero Anachyris. Candollea 2000, 55, 105–155. [Google Scholar]
- Denham, S.S.; Zuloaga, F.O.; Morrone, O. Systematic revision and phylogeny of Paspalum subgenus Ceresia (Poaceae: Panicoideae: Paniceae). Ann. Mo. Bot. Gard. 2002, 89, 337–399. [Google Scholar] [CrossRef]
- Rua, G.H.; Aliscioni, S.S. A morphology-based cladistic analysis of Paspalum sect. Pectinata (Poaceae). Syst. Bot. 2002, 27, 489–501. [Google Scholar] [CrossRef]
- Zuloaga, F.O.; Pensiero, J.; Morrone, O. Systematics of Paspalum grupo Notata (Poaceae, Panicoideae, Paniceae). Syst. Bot. Monogr. 2004, 71, 1–75. [Google Scholar] [CrossRef]
- Denham, S.S. Revisión sistemática del subgénero Harpostachys de Paspalum (Poaceae: Panicoideae: Paniceae). Ann. Mo. Bot. Gard. 2005, 92, 463–532. [Google Scholar]
- Delfini, C.; Souza, V.C.; Zuloaga, F.O. Taxonomic revision and nomenclatural update of Paspalum sect. Pectinata (Poaceae, Panicoideae, Paspaleae). Phytotaxa 2017, 323, 1–26. [Google Scholar] [CrossRef]
- Scataglini, M.A.; Zuloaga, F.O.; Giussani, L.M.; Denham, S.S.; Morrone, O. Phylogeny of new world Paspalum (Poaceae, Panicoideae, Paspaleae) based on plastid and nuclear markers. Plant Syst. Evol. 2014, 300, 1051–1070. [Google Scholar] [CrossRef]
- Cialdella, A.M.; Morrone, O.; Zuloaga, F.O. Revisión de las especies del género Paspalum (Poaceae: Panicoideae: Paniceae) grupo Bonplandiana. Darwiniana 1995, 33, 67–95. [Google Scholar]
- Morrone, O.; Zuloaga, F.O.; Carbonó, E. Revisión del grupo Racemosa del género Paspalum (Poaceae: Panicoideae: Paspaleae). Ann. Mo. Bot. Gard. 1995, 82, 82–116. [Google Scholar] [CrossRef]
- Morrone, O.; Vega, A.S.; Zuloaga, F.O. Revisión de las especies del género Paspalum L. (Poaceae: Panicoideae: Paniceae), grupo Dissecta (s. str.). Candollea 1996, 51, 2–34. [Google Scholar]
- Morrone, O.; Denham, S.S.; Zuloaga, F.O. Revisión taxonómica del género Paspalum grupo Eriantha (Poaceae, Panicoideae, Paniceae). Ann. Mo. Bot. Gard. 2004, 91, 225–246. [Google Scholar]
- Oliveira, R.C.; Valls, J.F.M. Taxonomia de Paspalum L., grupo Linearia (Gramineae–Paniceae) do Brasil. Rev. Bras. Bot. 2002, 25, 371–389. [Google Scholar] [CrossRef]
- Denham, S.S.; Morrone, O.; Zuloaga, F.O. Estudios en el género Paspalum (Poaceae, Panicoideae, Paniceae). Paspalum denticulatum y especies afines. Ann. Mo. Bot. Gard. 2010, 97, 11–33. [Google Scholar] [CrossRef]
- Pozzobon, M.T.; Paganella, M.B.; Santos, S.; Valls, J.F.M. Aspectos citológicos e reprodutivos no grupo Caespitosa do gênero Paspalum. Cienc. Rural 2013, 43, 2004–2010. [Google Scholar] [CrossRef]
- Delfini, C.; Souza, V.C.; Zuloaga, F.O. Notas Nomenclaturales en Paspalum grupo Caespitosa (Poaceae, Panicoideae, Paspaleae). Novon 2019, 27, 156–161. [Google Scholar] [CrossRef]
- Zuloaga, F.O.; Morrone, O.; Davidse, G.; Filgueiras, T.S.; Peterson, P.M.; Soreng, R.J.; Judziewicz, E.J. Catalogue of New World Grasses (Poaceae): III. Subfamilies Panicoideae, Aristidoideae, Arundinoideae, and Danthonioideae. Contr. U.S. Natl. Herb. 2003, 46, 1–662. [Google Scholar]
- Delfini, C.; Souza, V.C.; Zuloaga, F.O. Taxonomic Revision of Paspalum group Caespitosa (Poaceae, Panicoideae, Paspaleae). Ann. Mo. Bot. Gard. 2023, 108, 1–50. [Google Scholar] [CrossRef]
- Nash, G.V. Poaceae. In North American Flora; Britton, N.L., Murrill, W.A., Barnhart, J.H., Eds.; The New York Botanical Garden: New York, NY, USA, 1912; Volume 17, pp. 99–196. [Google Scholar]
- Hiratsuka, J.; Shimada, H.; Whittier, R.; Ishibashi, T.; Sakamoto, M.; Mori, M.; Kondo, C.; Honji, Y.; Sun, C.-R.; Meng, B.-Y.; et al. The complete sequence of the rice (Oryza sativa) chloroplast genome: Intermolecular recombination between distinct tRNA genes accounts for a major plastid DNA inversion during the evolution of the cereals. Mol. Genet. Genom. 1989, 217, 185–194. [Google Scholar] [CrossRef]
- Chang, C.-C.; Lin, H.-C.; Lin, I.-P.; Chow, T.-Y.; Chen, H.-H.; Chen, W.-H.; Cheng, C.-H.; Lin, C.-Y.; Liu, S.-M.; Chang, C.-C.; et al. The chloroplast genome of Phalaenopsis aphrodite (Orchidaceae): Comparative analysis of evolutionary rate with that of grasses and its phylogenetic implications. Mol. Biol. Evol. 2006, 23, 279–291. [Google Scholar] [CrossRef]
- Acosta, J.M.; Zuloaga, F.O.; Reinheimer, R. Nuclear phylogeny and hypothesized allopolyploidization events in the subtribe Otachyriinae (Paspaleae, Poaceae). Syst. Biodivers. 2019, 17, 277–294. [Google Scholar] [CrossRef]
- Acosta, J.M.; Scataglini, M.A.; Reinheimer, R.; Zuloaga, F.O. A phylogenetic study of subtribe Otachyriinae (Poaceae, Panicoideae, Paspaleae). Plant Syst. Evol. 2014, 300, 2155–2166. [Google Scholar] [CrossRef]
- Delfini, C.; Acosta, J.M.; Souza, V.C.; Zuloaga, F.O. Molecular phylogeny of Axonopus (Poaceae, Panicoideae, Paspaleae): Monophyly, synapomorphies and taxonomic implications for infrageneric classification and species complexes. Ann. Mo. Bot. Gard. 2020, 105, 459–480. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. Bot. Soc. Am. 1987, 19, 11–15. [Google Scholar]
- Baldwin, B.G.; Markos, S. Phylogenetic utility of the external transcribed spacers (ETS) of 18S-26S rDNA: Congruence of ETS and ITS trees of Calycadenia (Compositae). Mol. Phylogenet. Evol. 1998, 10, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.; Lickey, E.; Beck, J.T.; Farmer, S.B.; Liu, W.; Miller, J.; Siripun, K.C.; Winder, C.T.; Schilling, E.E.; Small, R.L. The tortoise and the hare II: Relative utility of 21 noncoding chloroplast DNA sequences for phylogenetic analysis. Am. J. Bot. 2005, 92, 142–166. [Google Scholar] [CrossRef] [PubMed]
- Kelchner, S.; Clark, L.G. Molecular evolution and phylogenetic utility of the chloroplast rpl16 intron in Chusquea and the Bambusoideae (Poaceae). Mol. Phylogenet. Evol. 1997, 8, 385–397. [Google Scholar] [CrossRef]
- Zhang, W. Phylogeny of the grass family (Poaceae) from rpl16 intron sequence data. Mol. Phylogenet. Evol. 2000, 15, 385–397. [Google Scholar] [CrossRef]
- Cialdella, A.M.; Giussani, L.M.; Aagesen, L.; Zuloaga, F.O.; Morrone, O. A phylogeny of Piptochaethium based on a combined analysis including trnL-F, rpl16 and morphology. Syst. Bot. 2007, 32, 545–559. [Google Scholar] [CrossRef]
- Jordan, W.C.; Courtney, W.M.; Neigel, E.J. Low levels of intraspecific genetic variation at a rapidly evolving chloroplast DNA locus in North American duckwoods (Lemnaceae). Am. J. Bot. 1996, 83, 430–439. [Google Scholar] [CrossRef]
- Giussani, L.M.; Zuloaga, F.O.; Quarín, C.L.; Cota-Sánchez, J.H.; Ubayasena, K.; Morrone, O. Phylogenetic relationships in the genus Paspalum (Poaceae: Panicoideae: Paniceae): An assessment of the Quadrifaria and Virgata informal groups. Syst. Bot. 2009, 34, 32–43. [Google Scholar] [CrossRef]
- Olmstead, R.G.; Sweere, J.A. Combining data in phylogenetic systematics: An empirical approach using three molecular data sets in the Solanaceae. Syst. Biol. 1994, 43, 467–481. [Google Scholar] [CrossRef]
- Aliscioni, S.S.; Giussani, L.M.; Zuloaga, F.O.; Kellogg, E.A. A molecular phylogeny of Panicum (Poaceae: Paniceae). Test of monophyly and phylogenetic placement within the Panicoideae. Am. J. Bot. 2003, 90, 796–821. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Fitch, W.M. Toward defining the course of evolution: Minimal change for a specific tree topology. Syst. Zool. 1971, 20, 406–416. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Crandall, K.A. Phylogeny estimation and hypothesis testing using maximum likelihood. Annu. Rev. Ecol. Evol. Syst. 1997, 28, 437–466. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Larget, B.; Miller, R.E.; Ronquist, F. Potential applications and pitfalls of Bayesian inference of phylogeny. Syst. Biol. 2002, 51, 673–688. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Nixon, K.C.; Carpenter, J.M. On simultaneous analysis. Cladistics 1996, 12, 221–241. [Google Scholar] [CrossRef]
- Farris, J.S.; KAllersjö, M.; Kluge, A.G.; Bult, C. Constructing a significance test for incongruence. Syst. Bot. Monogr. 1995, 44, 570–572. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic analysis using parsimony (and other methods), version 4.0 b10. Evolution 2002, 56, 1776–1788. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihoodbased phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web-servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Adamowski, E.V.; Pagliarini, M.S.; Batista, L.A.R. Chromosome number and microsporogenesis in Paspalum maritimum (Caespitosa group; Gramineae). Braz. Arch. Biol. Technol. 2000, 43, 301–305. [Google Scholar] [CrossRef]
- Maciel, J.R.; Oliveira, R.C.; Alves, M. Paspalum L. (Poaceae: Panicoideae: Paniceae) no estado de Pernambuco, Brasil. Acta Bot. Bras. 2009, 23, 1145–1161. [Google Scholar] [CrossRef]
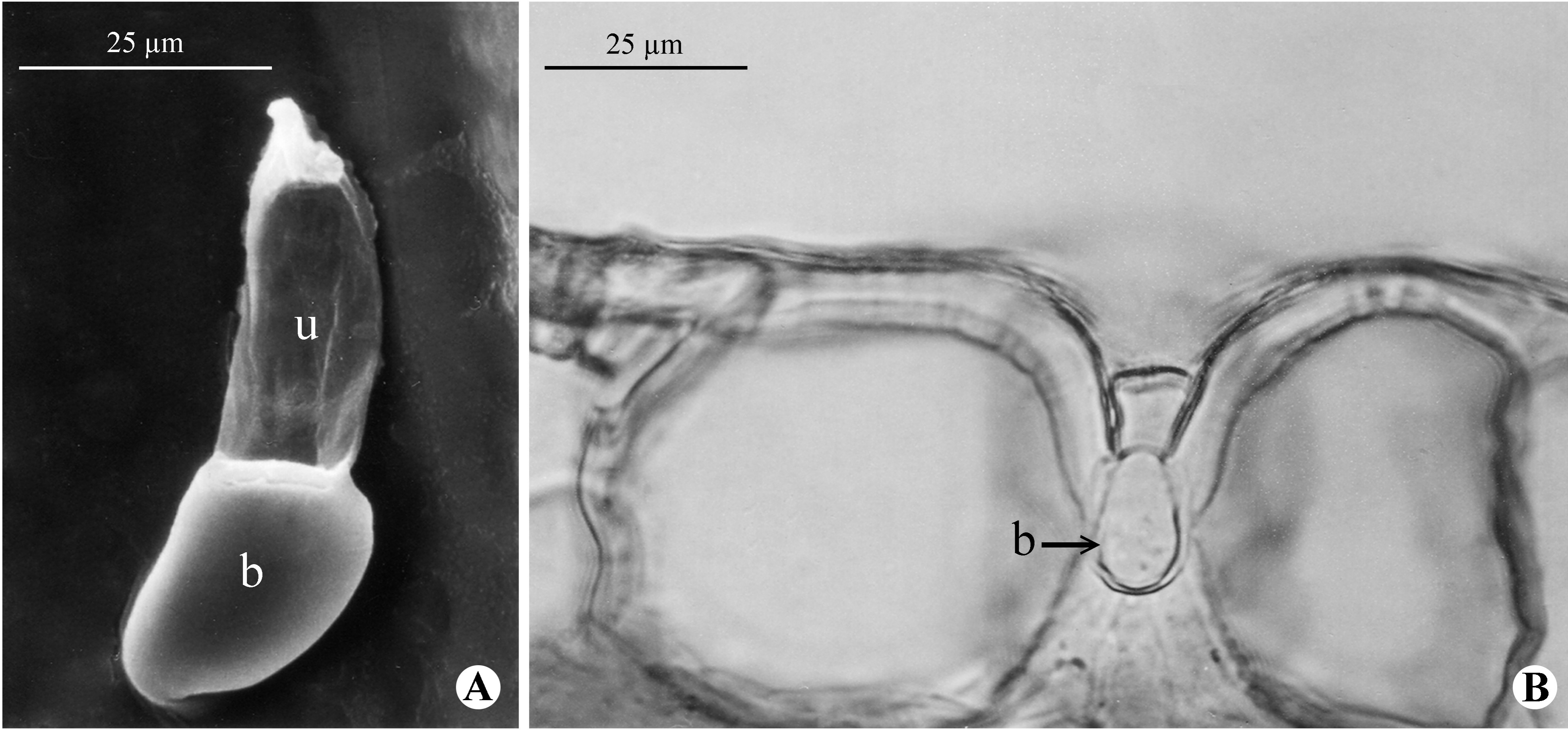
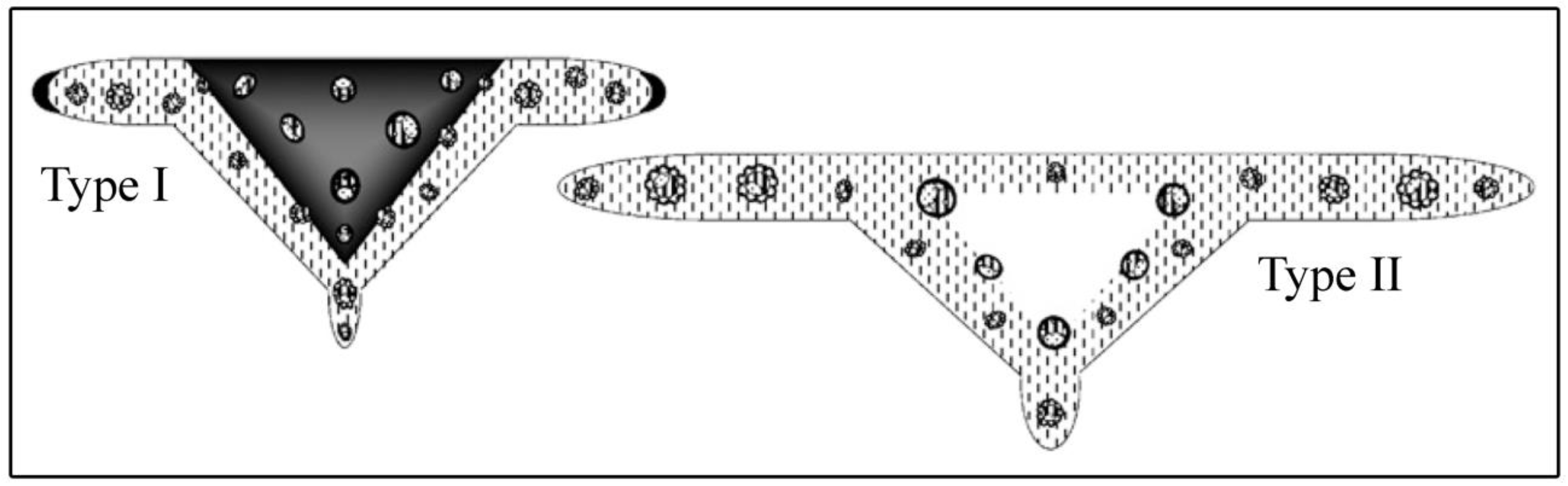
- Aliscioni, S.S. Anatomía ecológica de algunas especies del género Paspalum (Poaceae, Panicoideae, Paniceae). Darwiniana 2000, 38, 187–207. [Google Scholar] [CrossRef]
- Liphschitz, Ν.; Waisel, Y. Adaptation of plants to saline environments: Salt excretion and glandular structure. In Contributions of the Ecology of Halophytcs. Tasks for Vegetation Science; Sen, D.N., Rajpurohit, K.S., Eds.; Springer: Dordrecht, Netherlands, 1982; Volume 2, pp. 187–214. [Google Scholar]
- Aliscioni, S.S.; Denham, S.S. Rachis of the genus Paspalum L. (Poaceae: Panicoideae: Paniceae): Anatomy and taxonomic significance of the primary branches of the inflorescences. Flora 2008, 203, 60–76. [Google Scholar] [CrossRef]
- Aliscioni, S.S. Estudio Histofoliar Comparado de Especies Americanas del Género Paspalum L. (Poaceae: Panicoideae: Paniceae). PhD Thesis, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata,, La Plata, Argentina, 1999. [Google Scholar]
- POWO (Plants of the World Online). Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 8 July 2022).
- Hunziker, J.H.; Zuloaga, F.O.; Morrone, O.; Escobar, A. Estudios cromosómicos en Paniceae sudamericanas (Poaceae: Panicoideae). Darwiniana 1998, 35, 29–36. [Google Scholar]
- Morrone, O.; Escobar, A.; Zuloaga, F.O. Chromosome studies in American Panicoideae (Poaceae). Ann. Mo. Bot. Gard. 2006, 93, 647–657. [Google Scholar] [CrossRef]
- Metcalfe, C.R. Anatomy of the Monocotyledons, I: Gramineae, 1st ed.; Clarendon Press: Oxford, MS, USA, 1960; 794p. [Google Scholar]
- Ellis, R.P. A procedure for standardizing comparative leaf anatomy in the Poaceae. II. The epidermis as seen in surface view. Bothalia 1979, 12, 641–671. [Google Scholar] [CrossRef]
- Hitchcock, A.S. New species of grasses from the Galapagos and the Revillagigedo Islands. Proc. Calif. Acad. Sci. 1935, 21, 295–300. [Google Scholar]
- Cotton, J.L.; Wysocki, W.P.; Clark, L.G.; Kelchner, S.A.; Pires, J.C.; Edger, P.P.; Mayfield-Jones, D.; Duvall, M.R. Resolving deep relationships of PACMAD grasses: A phylogenomic approach. BMC Plant Biol. 2015, 15, 178. [Google Scholar] [CrossRef]
- Burke, S.V.; Wysocki, W.P.; Zuloaga, F.O.; Craine, J.M.; Pires, J.C.; Edger, P.P.; Mayfield-Jones, D.; Clark, L.G.; Kelchner, S.A.; Duvall, M.R. Evolutionary relationships in Panicoid grasses based on plastome phylogenomics (Panicoideae; Poaceae). BMC Plant Biol. 2016, 16, 140. [Google Scholar] [CrossRef]
- Saarela, J.M.; Burke, S.V.; Wysocki, W.P.; Barrett, M.D.; Clark, L.G.; Craine, J.M.; Peterson, P.M.; Soreng, R.J.; Vorontsova, M.S.; Duvall, M.R. A 250 plastome phylogeny of the grass family (Poaceae): Topological support under different data partitions. PeerJ 2018, 6, e4299. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, L.; Columbus, J.T.; Hu, Y.; Zhao, Y.; Tang, L.; Guo, Z.; Chen, W.; McKain, M.; Bartlett, M.; et al. A well-supported nuclear phylogeny of Poaceae and implications for the evolution of C4 photosynthesis. Mol. Plant 2022, 15, 755–777. [Google Scholar] [CrossRef]
- Estep, M.C.; McKain, M.R.; Diaz, D.V.; Zhong, J.; Hodge, J.G.; Hodkinson, T.R.; Layton, D.J.; Malcomber, S.T.; Pasquet, R.; Kellogg, E.A. Allopolyploidy, diversification, and the Miocene grassland expansion. Proc. Natl. Acad. Sci. USA 2014, 111, 15149–15154. [Google Scholar] [CrossRef]
- Kellogg, E.A. Has the connection between polyploidy and diversification actually been tested? Curr. Opin. Plant Biol. 2016, 30, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Sang, T. Utility of low-copy nuclear gene sequences in plant phylogenetics. Crit. Rev. Biochem. Mol. Biol. 2002, 37, 121–147. [Google Scholar] [CrossRef] [PubMed]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff, New York Botanical Garden’s Virtual Herbarium, continuously updated. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 8 March 2022).
- Morrone, O.; Hunziker, J.H.; Zuloaga, F.O.; Escobar, A. Números cromosómicos en Paniceae sudamericanas (Poaceae: Panicoideae). Darwiniana 1995, 3, 53–60. [Google Scholar]
- Burson, B.L. Cytology of Paspalum chacoense and P. durifolium and their relationship to P. dilatatum. Bot. Gaz. 1985, 146, 124–129. [Google Scholar] [CrossRef]
- Quarín, C.L. Recuentos cromosómicos en gramíneas de Argentina subtropical. Hickenia 1977, 1, 73–78. [Google Scholar]
- Quarín, C.L.; Burson, B.L. Cytogenetic relations among Paspalum notatum var. saurae, P. pumilum, P. indecorum, and P. vaginatum. Bot. Gaz. 1983, 144, 433–438. [Google Scholar]
- Quarín, C.L.; Hanna, W.W.; Fernandez, A. Genetic studies in diploid and tetraploid Paspalum species. Embryo sac development, chromosome behavior, and fertility in P. cromyorhizon, P. laxum and P. proliferum. J. Hered. 1982, 73, 254–256. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delfini, C.; Acosta, J.M.; Aliscioni, S.S.; Souza, V.C.; Zuloaga, F.O. Phylogenetic Relationships in the Group Caespitosa of Paspalum L. (Poaceae, Panicoideae, Paspaleae). Diversity 2023, 15, 134. https://doi.org/10.3390/d15020134
Delfini C, Acosta JM, Aliscioni SS, Souza VC, Zuloaga FO. Phylogenetic Relationships in the Group Caespitosa of Paspalum L. (Poaceae, Panicoideae, Paspaleae). Diversity. 2023; 15(2):134. https://doi.org/10.3390/d15020134
Chicago/Turabian StyleDelfini, Carolina, Juan M. Acosta, Sandra S. Aliscioni, Vinicius Castro Souza, and Fernando O. Zuloaga. 2023. "Phylogenetic Relationships in the Group Caespitosa of Paspalum L. (Poaceae, Panicoideae, Paspaleae)" Diversity 15, no. 2: 134. https://doi.org/10.3390/d15020134
APA StyleDelfini, C., Acosta, J. M., Aliscioni, S. S., Souza, V. C., & Zuloaga, F. O. (2023). Phylogenetic Relationships in the Group Caespitosa of Paspalum L. (Poaceae, Panicoideae, Paspaleae). Diversity, 15(2), 134. https://doi.org/10.3390/d15020134