
Two New Uromunna Species (Isopoda: Asellota: Munnidae) from the Korean Peninsula and Their Phylogenetic Position within Munnoid Groups †




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
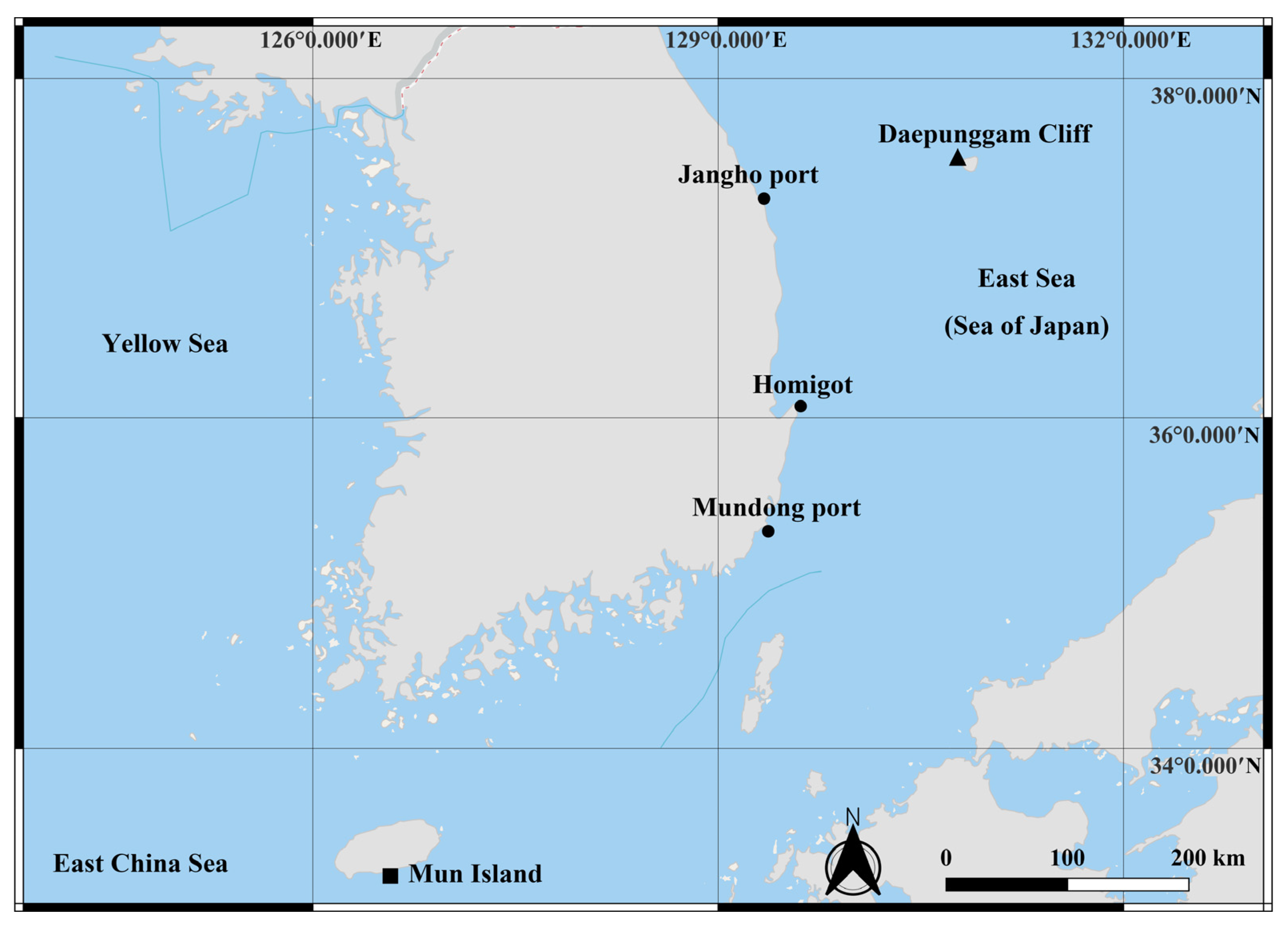
2. Material and Methods
2.1. Sample Collection and Morphological Examination
2.2. DNA Amplification and Sequencing
2.3. Sequence Alignment and Phylogenetic Analysis
3. Results
3.1. Taxonomy
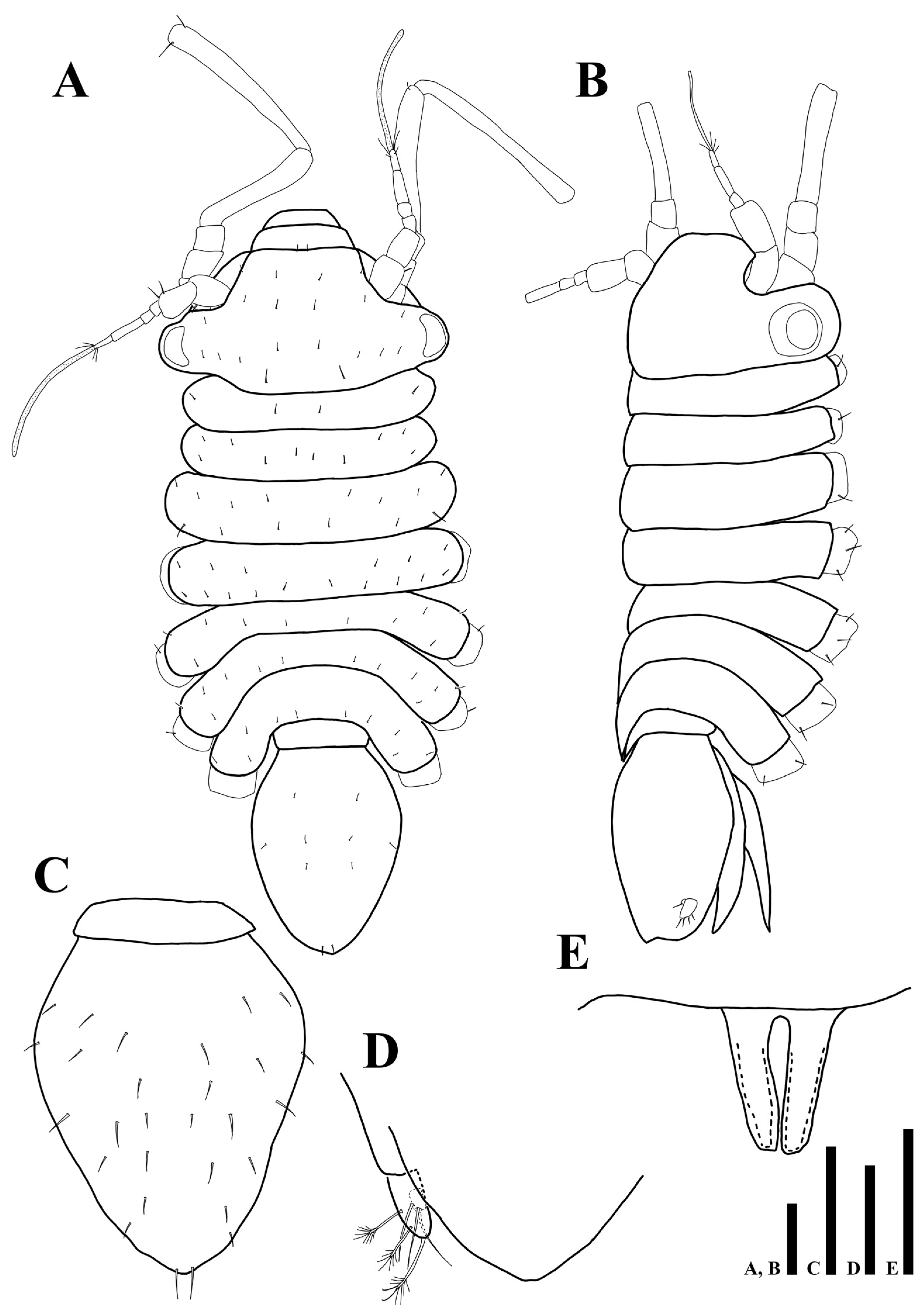
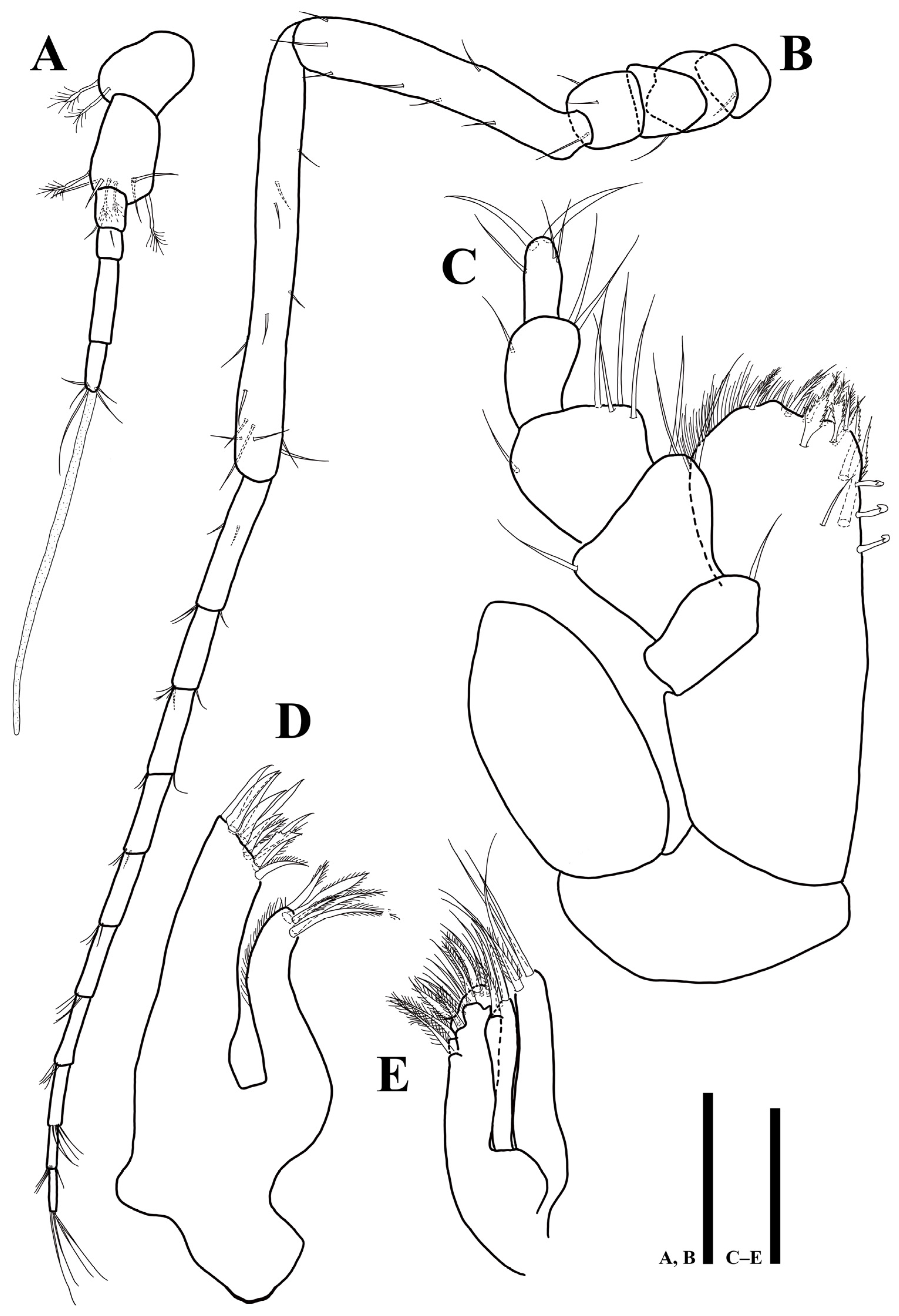
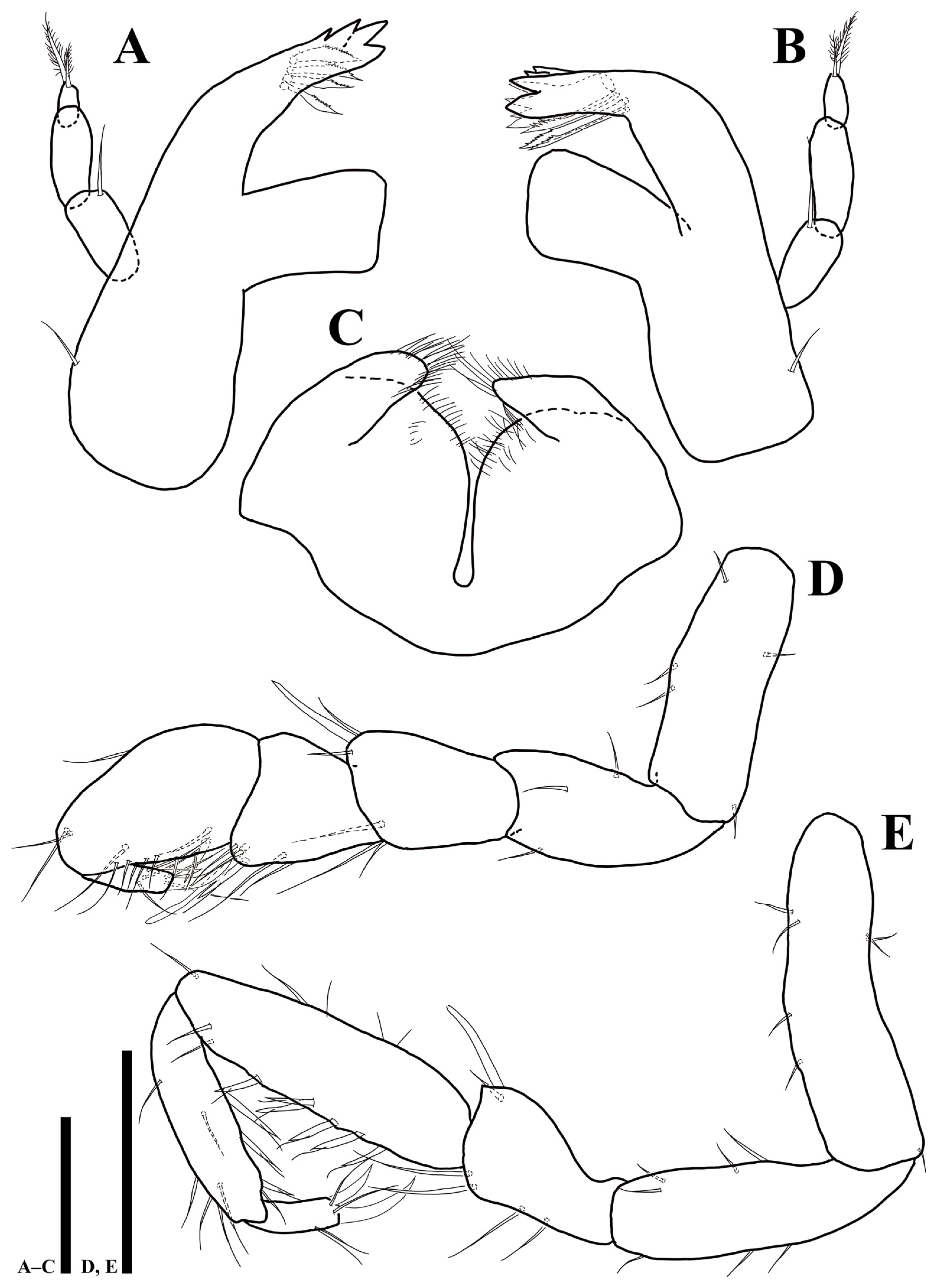
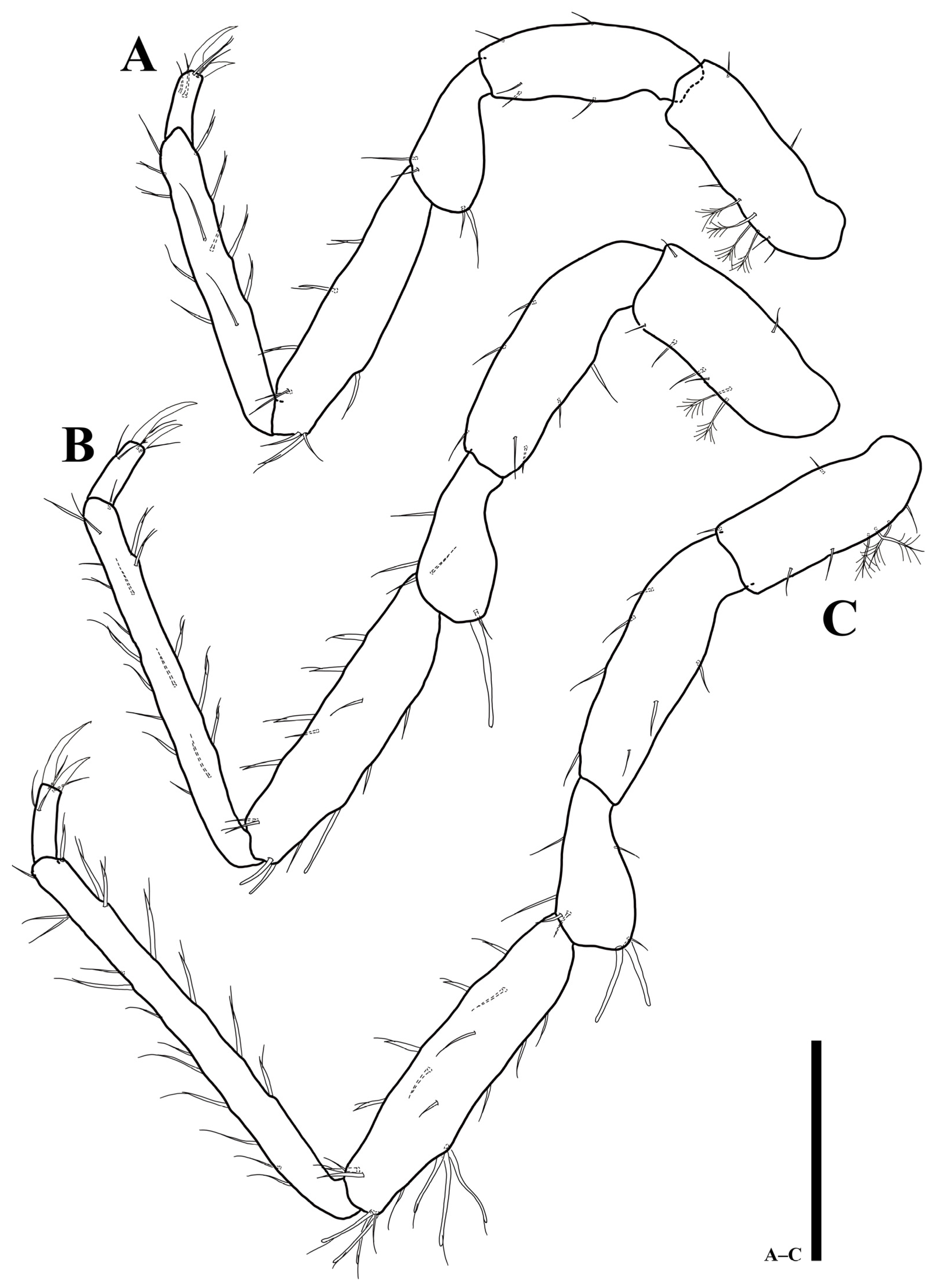
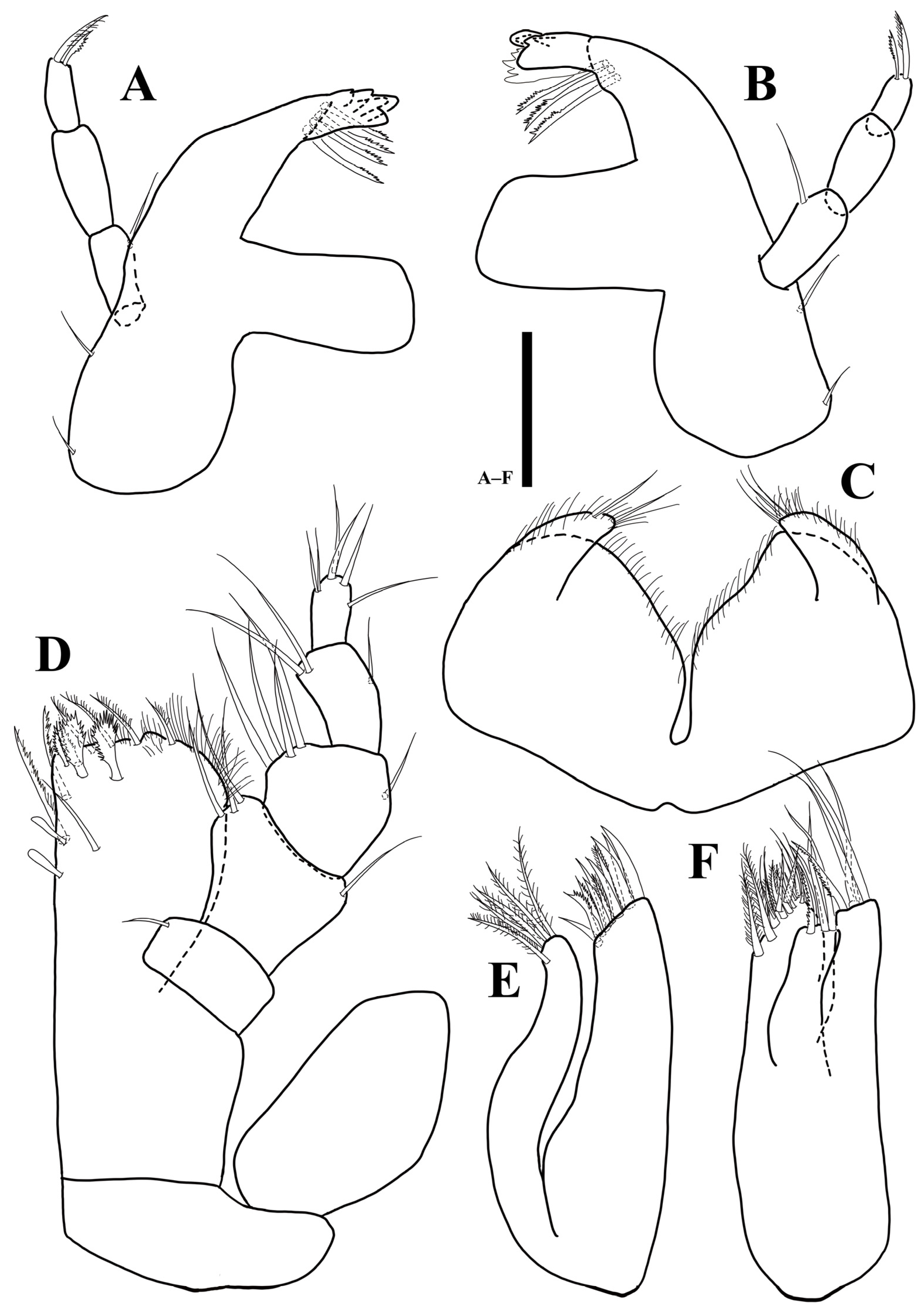
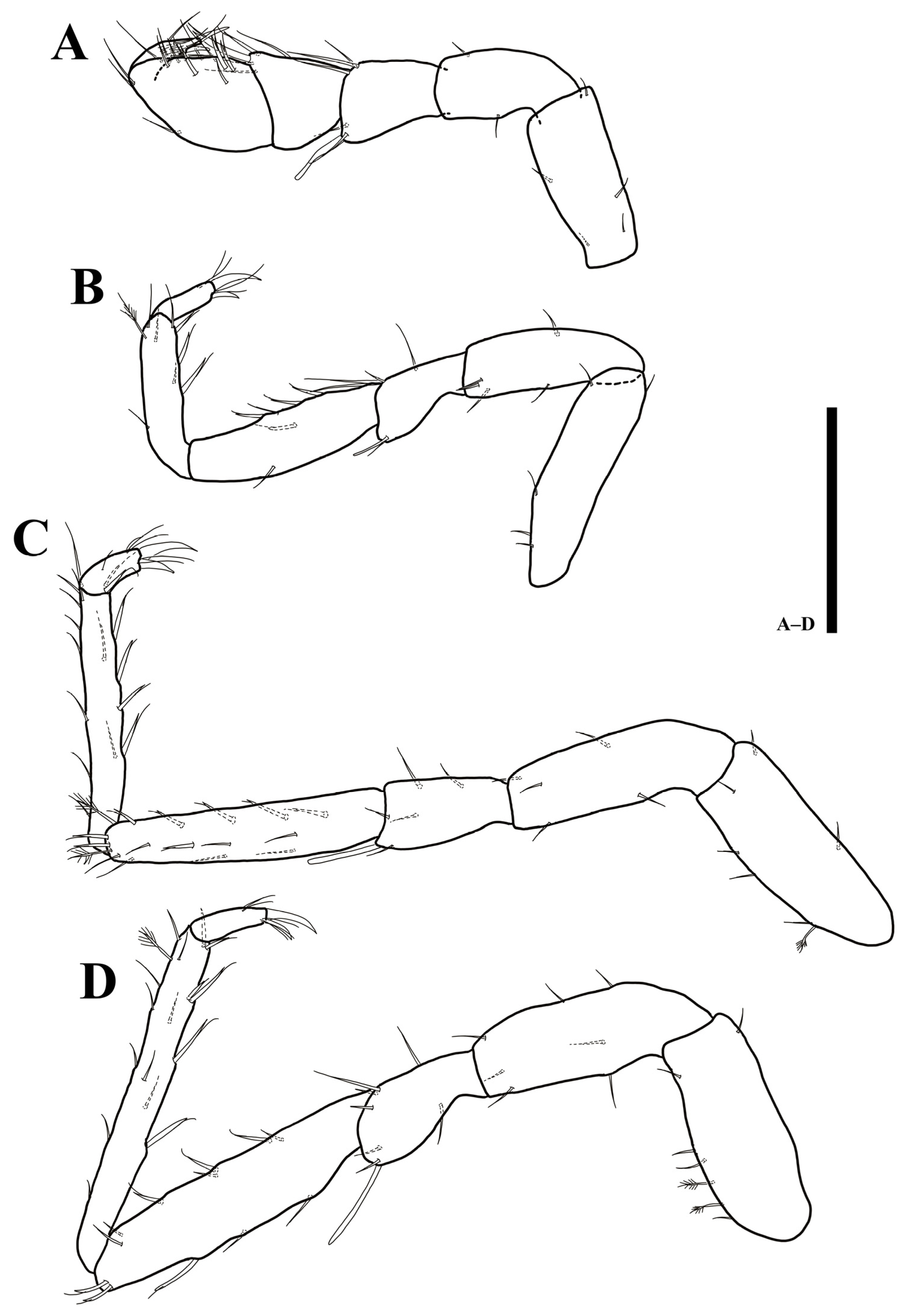
3.2. Uromunna mundongensis sp. n.
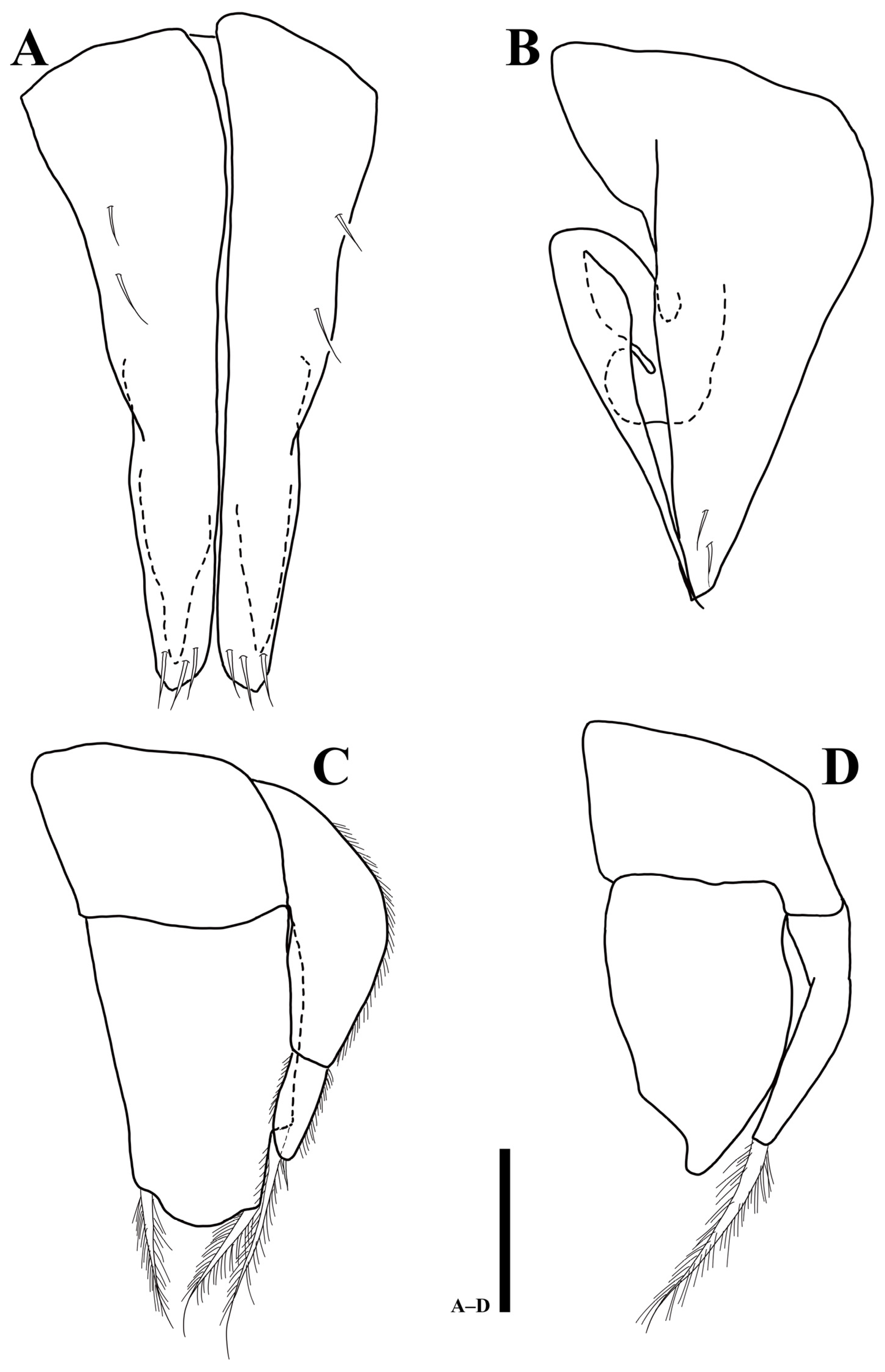
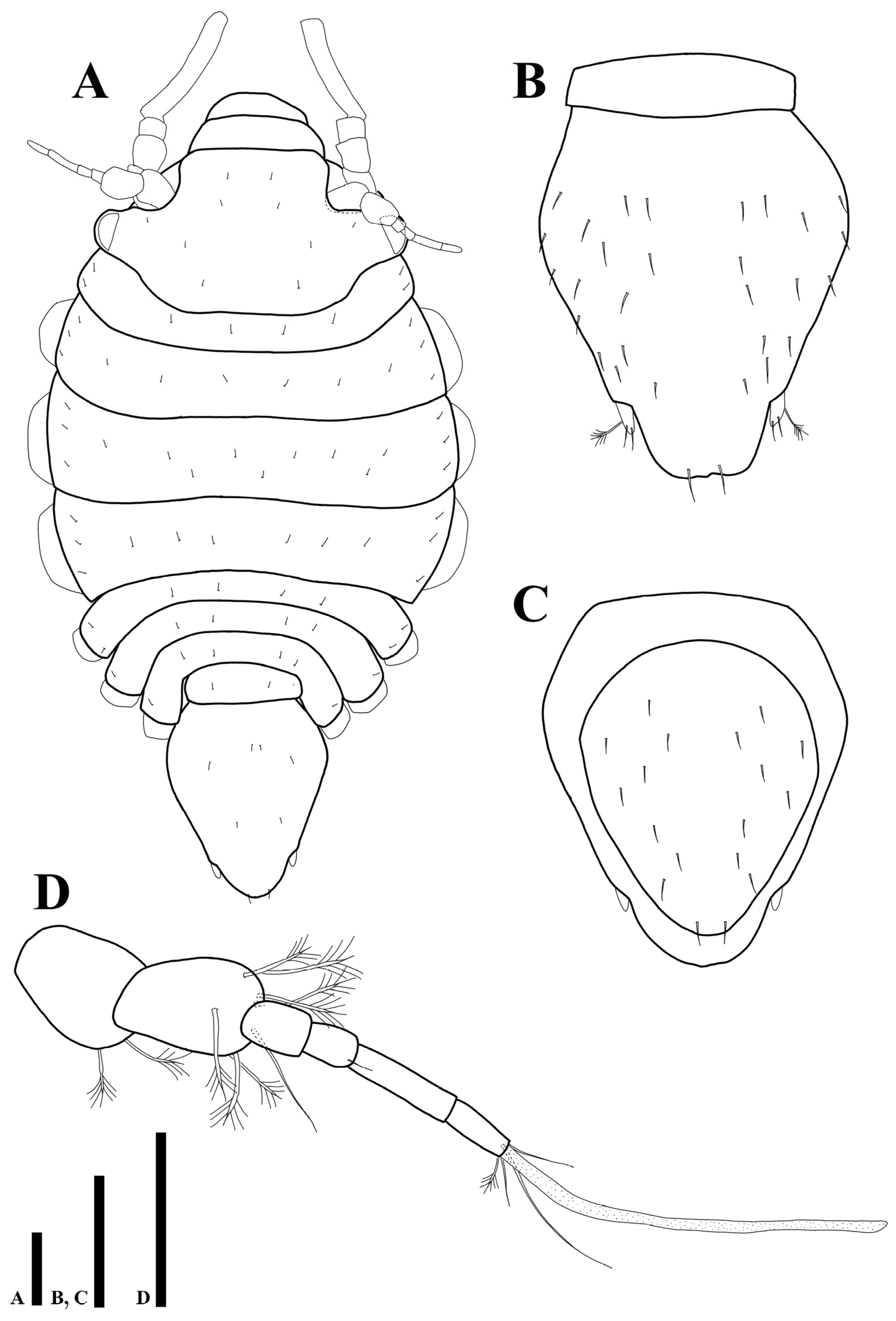
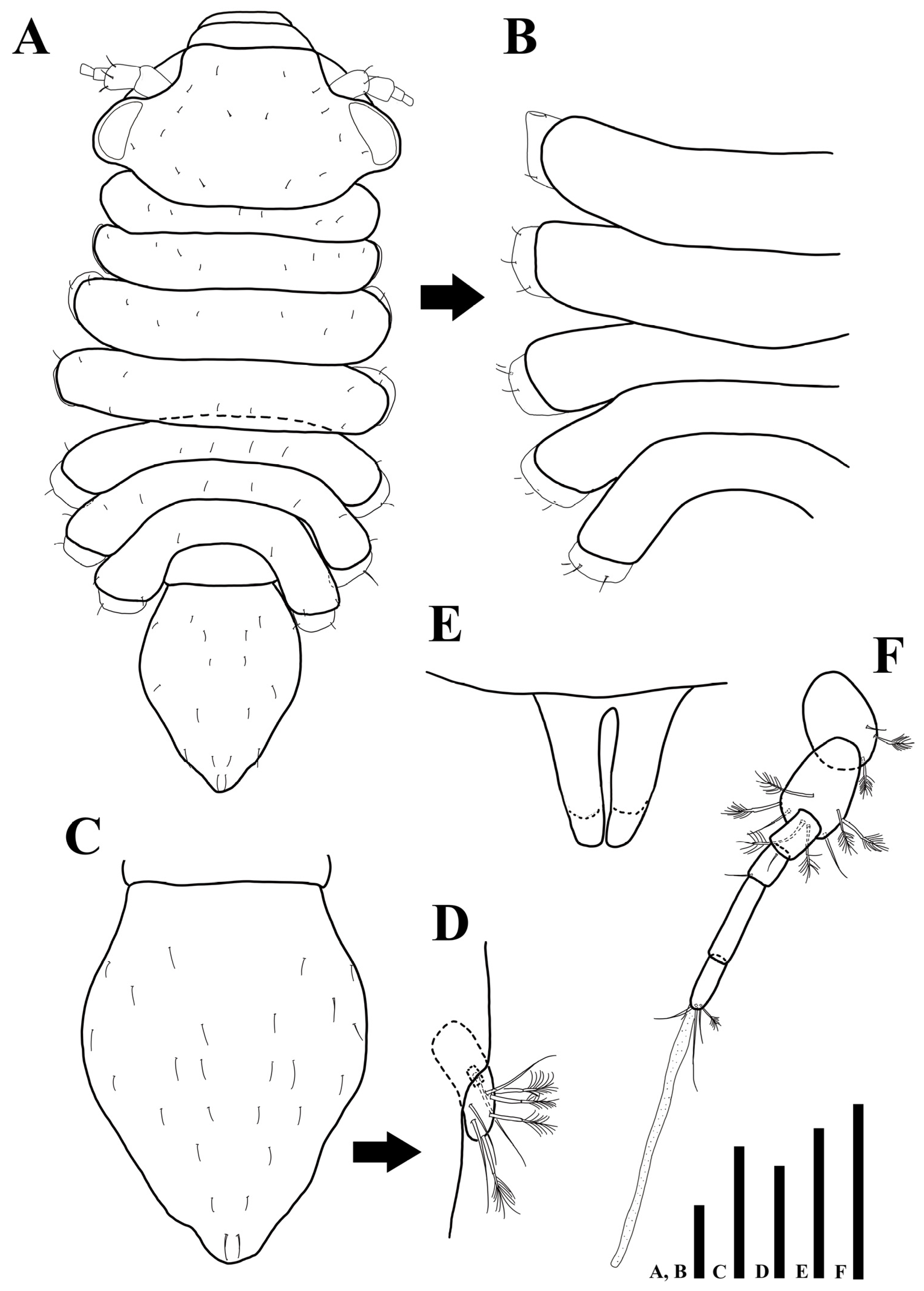
3.2.1. Description of the Male Holotype
3.2.2. Sexual Dimorphism
3.2.3. Remarks
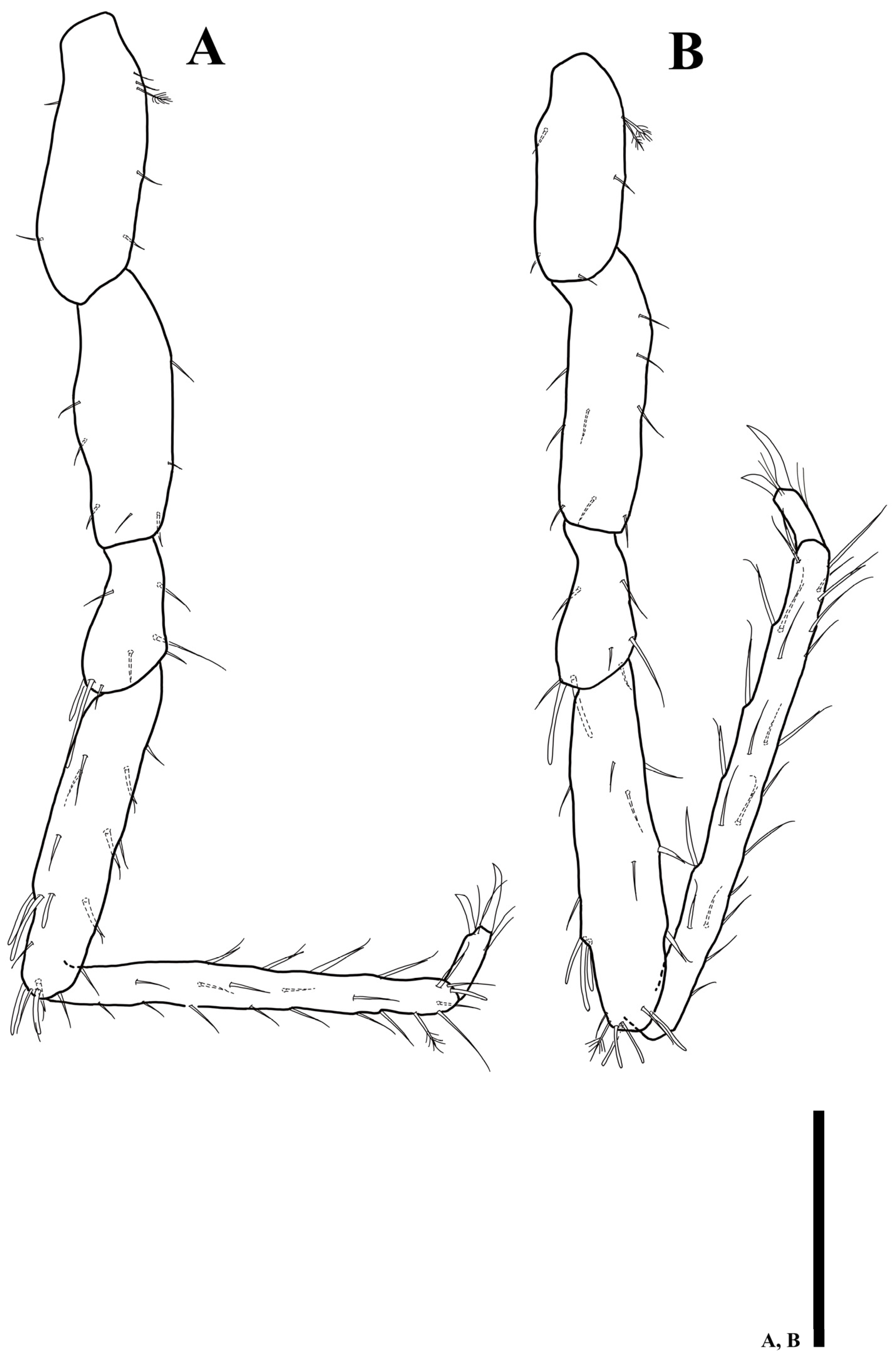
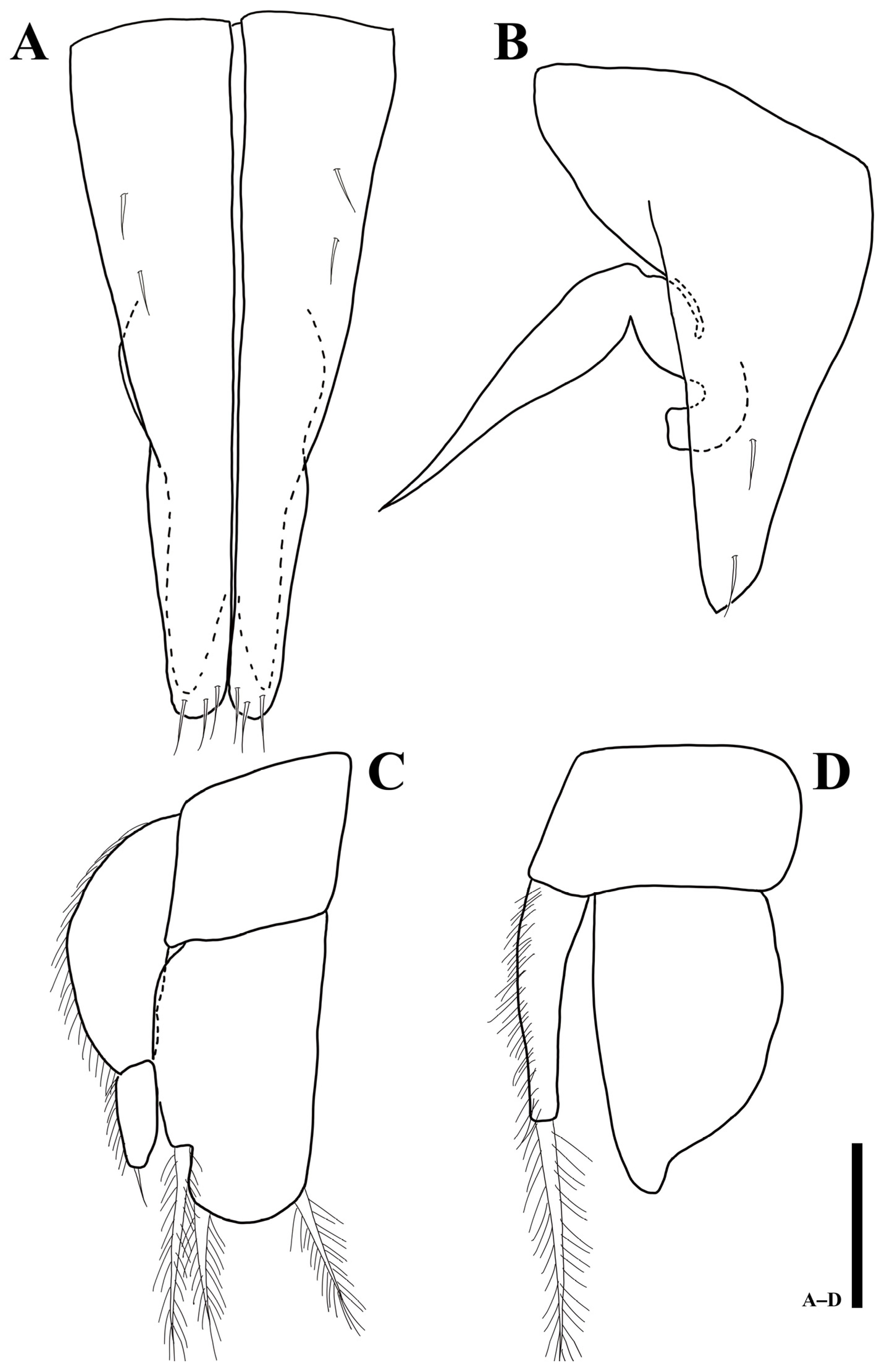
3.3. Uromunna jejuensis sp. n.
3.3.1. Description of the Male Holotype
3.3.2. Sexual Dimorphism
3.3.3. Remarks
3.4. Key to the East Asian species of Uromunna
- 1.
- Antennula second article with six setae; maxillipedal palp first article without setae; lateral margin of pleotelson serrated ............................................................ U. serricauda
- −
- Antennula second article with eight setae; maxillipedal palp first article with one seta; lateral margin of pleotelson smooth …………………………..…………...……………. 2
- 2.
- Antennula first article with two setae; antennula fourth article without setae; three maxillipedal coupling hooks; one seta on basal margin of mandible ……………………………………………………………………… U. mundongensis sp. n.
- −
- Antennula first article with three setae; antennula fourth article with one seta; two maxillipedal coupling hooks; two setae on basal margin of mandible …………………………………………………………………………… U. jejuensis sp. n.
- 1.
- Lateral margin of pleotelson smooth............................................ U. mundongensis sp. n.
- −
- Lateral margin of pleotelson serrated ……………...……………………...……………. 2
- 2.
- Operculum as long as broad with eight simple setae on dorsal surface …………………………………………………………………………………. U. serricauda
- −
- Operculum longer than wide with 18 simple setae on dorsal surface …………………………………………………………………………… U. jejuensis sp. n.
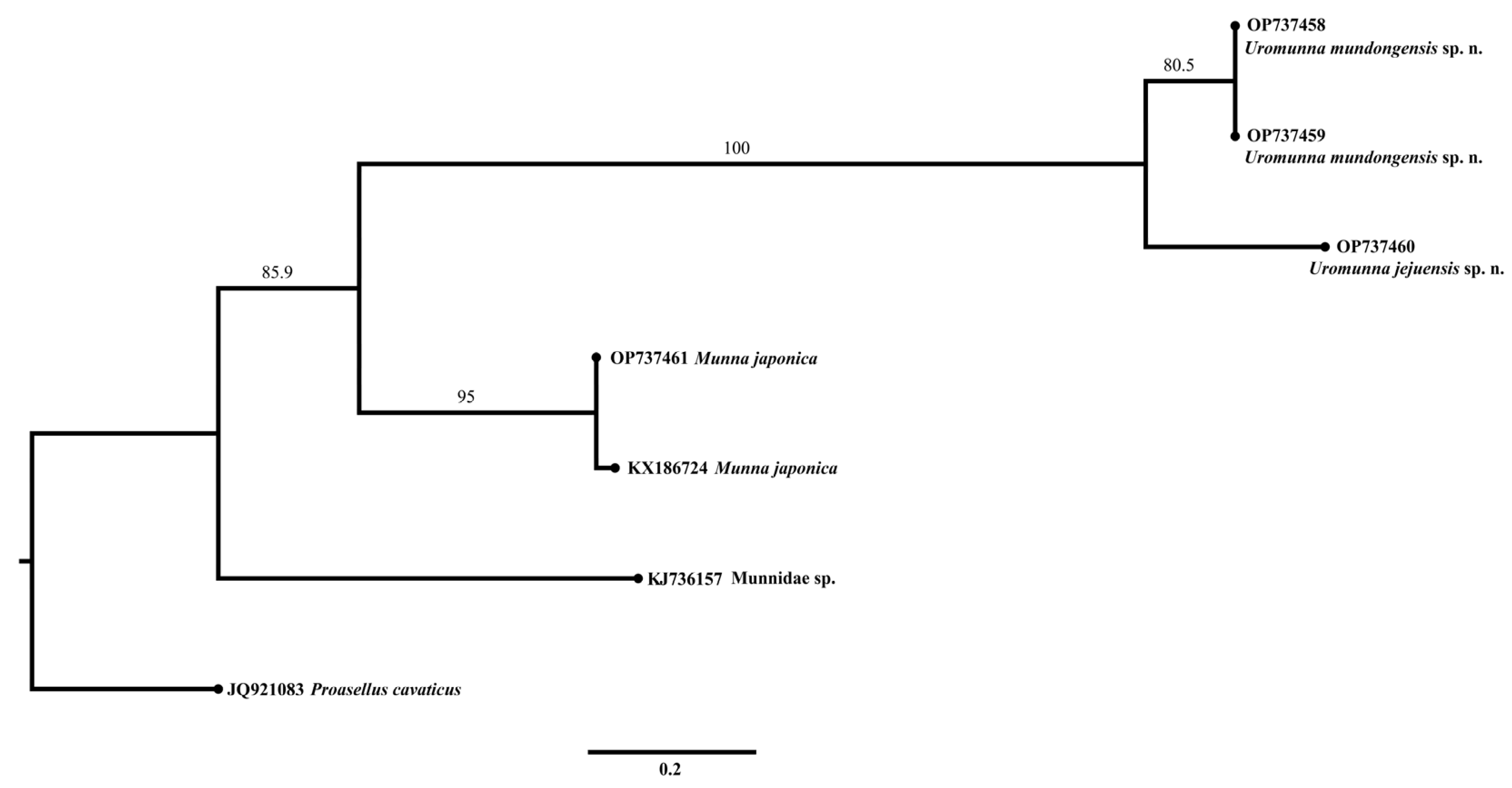
3.5. Phylogeny
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bock, W.J. Species: The concept, category and taxon. J. Zoolog. Syst. Evol. Res. 2004, 42, 178–190. [Google Scholar] [CrossRef]
- Hey, J. On the failure of modern species concepts. Trends Ecol. Evol. 2006, 21, 447–450. [Google Scholar] [CrossRef]
- De Queiroz, K. Branches in the lines of descent: Charles Darwin and the evolution of the species concept. Biol. J. Linn. Soc. Lond. 2011, 103, 19–35. [Google Scholar] [CrossRef]
- Hausdorf, B. Progress toward a general species concept. Evol. Int. J. Org. Evol. 2011, 65, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Sites, J.W., Jr.; Marshall, J.C. Delimiting species: A Renaissance issue in systematic biology. Trends Ecol. Evol. 2003, 18, 462–470. [Google Scholar] [CrossRef]
- De Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef]
- Yeates, D.K.; Seago, A.; Nelson, L.; Cameron, S.L.; Joseph, L.E.O.; Trueman, J.W. Integrative taxonomy, or iterative taxonomy? Syst. Entomol. 2011, 36, 209–217. [Google Scholar] [CrossRef]
- Padial, J.M.; Miralles, A.; De la Riva, I.; Vences, M. The integrative future of taxonomy. Front. Zool. 2010, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- De Queiroz, K. A unified concept of species and its consequences for the future of taxonomy. In Proceedings of the California Academy of Sciences; Leviton, A.E., Williams, G.C., Aldrich, M.L., Eds.; California Academy of Sciences: San Francisco, CA, USA, 2005; Volume 54, pp. 196–215. [Google Scholar]
- Fišer, C.; Robinson, C.T.; Malard, F. Cryptic species as a window into the paradigm shift of the species concept. Mol. Ecol. 2018, 27, 613–635. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Dayrat, B.; Gosliner, T.M. Species names and metaphyly: A case study in Discodorididae (Mollusca, Gastropoda, Euthyneura, Nudibranchia, Doridina). Zool. Scr. 2005, 34, 199–224. [Google Scholar] [CrossRef]
- Schlick-Steiner, B.C.; Steiner, F.M.; Seifert, B.; Stauffer, C.; Christian, E.; Crozier, R.H. Integrative taxonomy: A multisource approach to exploring biodiversity. Annu. Rev. Entomol. 2010, 55, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Andújar, C.; Arribas, P.; Ruiz, C.; Serrano, J.; Gómez--Zurita, J. Integration of conflict into integrative taxonomy: Fitting hybridization in species delimitation of Mesocarabus (Coleoptera: Carabidae). Mol. Ecol. 2014, 23, 4344–4361. [Google Scholar] [CrossRef]
- Pante, E.; Schoelinck, C.; Puillandre, N. From integrative taxonomy to species description: One step beyond. Syst. Biol. 2015, 64, 152–160. [Google Scholar] [CrossRef]
- Karanovic, T.; Djurakic, M.; Eberhard, S.M. Cryptic species or inadequate taxonomy? Implementation of 2D geometric morphometrics based on integumental organs as landmarks for delimitation and description of copepod taxa. Syst. Biol. 2016, 65, 304–327. [Google Scholar] [CrossRef]
- Poore, G.C.B.; Bruce, N.L. Global Diversity of Marine Isopods (Except Asellota and Crustacean Symbionts). PLoS ONE 2012, 7, e43529. [Google Scholar] [CrossRef]
- Held, C.; Wägele, J.W. Cryptic speciation in the giant Antarctic isopod Glyptonotus antarcticus (Isopoda, Valvifera, Chaetiliidae). Sci. Mar. 2005, 69, 175–181. [Google Scholar] [CrossRef]
- Raupach, M.J.; Wägele, J.W. Distinguishing cryptic species in Antarctic Asellota (Crustacea: Isopoda)—A preliminary study of mitochondrial DNA in Acanthaspidia drygalskii. Antarct. Sci. 2006, 18, 191–198. [Google Scholar] [CrossRef]
- Leese, F.; Kop, A.; Wägele, J.W.; Held, C. Cryptic speciation in a benthic isopod from Patagonian and Falkland Island waters and the impact of glaciations on its population structure. Front. Zool. 2008, 5, 19. [Google Scholar] [CrossRef]
- Brix, S.; Riehl, T.; Leese, F. First genetic data for species of the genus Haploniscus Richardson, 1908 (Isopoda: Asellota: Haploniscidae) from neighbouring deep-sea basins in the South Atlantic. Zootaxa 2011, 2838, 79. [Google Scholar] [CrossRef]
- Kim, J.; Kim, J.; Lee, W.; Karanovic, I. The first insight into the patterns of size and shape variation of a microcerberid isopod. Water 2021, 13, 515. [Google Scholar] [CrossRef]
- Schnurr, S.; Brandt, A.; Brix, S.; Fiorentino, D.; Malyutina, M.; Svavarsson, J. Composition and distribution of selected munnopsid genera (Crustacea, Isopoda, Asellota) in Icelandic waters. Deep-Sea Res. I Oceanogr. Res. Pap. 2014, 84, 142–155. [Google Scholar] [CrossRef]
- Kaiser, S.; Brix, S.; Kihara, T.C.; Janssen, A.; Jennings, R.M. Integrative species delimitation in the deep-sea genus Thaumastosoma Hessler, 1970 (Isopoda, Asellota, Nannoniscidae) reveals a new genus and species from the Atlantic and central Pacific abyss. Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 148, 151–179. [Google Scholar] [CrossRef]
- Riehl, T.; Lins, L.; Brandt, A. The effects of depth, distance, and the Mid-Atlantic Ridge on genetic differentiation of abyssal and hadal isopods (Macrostylidae). Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 148, 74–90. [Google Scholar] [CrossRef]
- Teske, P.R.; Papadopoulos, I.; Zardi, G.I.; McQuaid, C.D.; Edkins, M.T.; Griffiths, C.L.; Barker, N.P. Implications of life history for genetic structure and migration rates of southern African coastal invertebrates: Planktonic, abbreviated and direct development. Mar. Biol. 2007, 152, 697–711. [Google Scholar] [CrossRef]
- Raupach, M.J.; Malyutina, M.; Brandt, A.; Wägele, J.W. Molecular data reveal a highly diverse species flock within the munnopsoid deep-sea isopod Betamorpha fusiformis (Barnard, 1920) (Crustacea: Isopoda: Asellota) in the Southern Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 1820–1830. [Google Scholar] [CrossRef]
- Brandt, A.; Brix, S.; Held, C.; Kihara, T.C. Molecular differentiation in sympatry despite morphological stasis: Deep-sea Atlantoserolis Wägele, 1994 and Glabroserolis Menzies, 1962 from the south-west Atlantic (Crustacea: Isopoda: Serolidae). Zool. J. Linn. Soc. 2014, 172, 318–359. [Google Scholar] [CrossRef][Green Version]
- Kim, J.; Malyutina, M.; Lee, W.; Karanovic, I. Incongruence between morphological and molecular diversity in Coxicerberus fukudai (Ito, 1974) (Isopoda: Microcerberidea) from East Asia. J. Crustac. Biol. 2018, 38, 315–328. [Google Scholar] [CrossRef]
- Wilson, G.D.F. New insights into the colonization of the deep sea: Systematics and zoogeography of the Munnidae and the Pleurogoniidae comb. nov. (Isopoda; Janiroidea). J. Nat. Hist. 1980, 14, 215–236. [Google Scholar] [CrossRef]
- Hessler, R.R.; Strömberg, J.O. Behavior of janiroidean isopods (Asellota), with special reference to deep-sea genera. Sarsia 1989, 74, 45–159. [Google Scholar] [CrossRef]
- Esquete, P.; Moreira, J.; Troncoso, J.S. Peracarid assemblages of Zostera meadows in an estuarine ecosystem (O Grove inlet, NW Iberian Peninsula): Spatial distribution and seasonal variation. Helgol. Mar. Res. 2011, 65, 445–455. [Google Scholar] [CrossRef][Green Version]
- Esquete, P.; Wilson, G.D.F.; Troncoso, J.S. Ecology and systematics of a new species of Uromunna (Crustacea: Isopoda) from Spanish eelgrass beds. Helgol. Mar. Res. 2014, 68, 329–339. [Google Scholar] [CrossRef][Green Version]
- Boyko, C.B.; Bruce, N.L.; Hadfield, K.A.; Merrin, K.L.; Ota, Y.; Poore, G.C.B.; Taiti, S. World Marine, Freshwater and Terrestrial Isopod Crustaceans database. 2008 onwards. Available online: https://www.marinespecies.org/isopoda (accessed on 14 November 2022). [CrossRef]
- Menzies, R.J. The zoogeography, ecology and systematics of the Chilean marine isopods. Reports of the Lund University Chile Expedition 1948–49. Lunds Univ. Arsskrift 1962, 42, 1–162. [Google Scholar]
- Kussakin, O.G. On the fauna of Munnidae (Isopoda: Asellota) from the Far-Eastern seas of the USSR. Tr. Zool. Inst. Akad. Nauk. USSR 1962, 30, 66–109. [Google Scholar]
- Frankenberg, D.; Menzies, R.J. 1966. A new species of asellote marine isopod, Munna (Uromunna) reynoldsi (Crustacea: Isopoda). Bull. Mar. Sci. 1966, 16, 200–208. [Google Scholar]
- Kensley, B.F. A new species of Munna Kroyer from Nigeria (Crustacea: Isopoda: Asellota). Proc. Biol. Soc. Wash. 1980, 93, 136–140. Available online: https://repository.si.edu/handle/10088/10269 (accessed on 11 November 2022).
- Poore, G.C. Redefinition of Munna and Uromunna (Crustacea: Isopoda: Munnidae), with descriptions of five species from coastal Victoria. Proc. R. Soc. Vict. 1984, 96, 61–81. [Google Scholar]
- Esquete, P.; Wilson, G.D.F. The genus Uromunna (Crustacea: Isopoda: Munnidae) in New South Wales, Australia, with a key for all known species. Mar. Biol. Res. 2016, 12, 488–512. [Google Scholar] [CrossRef]
- Shimomura, M.; Mawatari, S.F. Munnidae from Japan (Crustacea: Isopoda: Asellota). Publ. Seto Mar. Biol. Lab. 2011, 39, 45–73. [Google Scholar] [CrossRef][Green Version]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Raupach, M.J.; Mayer, C.; Malyutina, M.; Wägele, J.W. Multiple origins of deep-sea Asellota (Crustacea: Isopoda) from shallow waters revealed by molecular data. Proc. R. Soc. B Biol. Sci. 2009, 276, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Raupach, M.J.; Held, C.; Wägele, J.W. Multiple colonization of the deep sea by the Asellota (Crustacea: Peracarida: Isopoda). Deep Sea Res. Part II Top. Stud. Oceanogr. 2004, 51, 1787–1795. [Google Scholar] [CrossRef]
- Riehl, T.; Brandt, A. Descriptions of two new species in the genus Macrostylis Sars, 1864 (Isopoda, Asellota, Macrostylidae) from the Weddell Sea (Southern Ocean), with a synonymisation of the genus Desmostylis Brandt, 1992 with Macrostylis. Zookeys 2010, 57, 9. [Google Scholar] [CrossRef] [PubMed]
- Geller, J.; Meyer, C.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all--taxa biotic surveys. Mol. Ecol. Resour. 2013, 13, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Boore, J.L.; Brown, W.M. Mitochondrial genomes of Galathealinum, Helobdella, and Platynereis: Sequence and gene arrangement comparisons indicate that Pogonophora is not a phylum and Annelida and Arthropoda are not sister taxa. Mol. Biol. Evol. 2000, 17, 87–106. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Song, J.H.; Min, G.S. New record of limnoriid and asellote species (Crustacea: Malacostraca: Isopoda) from South Korea. J. Species Res. 2017, 6, 237–240. [Google Scholar] [CrossRef]
- Wàgele, J.W. On the origin of the Microcerberidae (Crustacea: Isopoda). J. Zool. Syst. Evol. Res. 1983, 21, 249–262. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree. Tree Figure Drawing Tool. 2009. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 17 November 2022).
- Janssen, A.; Kaiser, S.; Meissner, K.; Brenke, N.; Menot, L.; Martínez Arbizu, P. A reverse taxonomic approach to assess macrofaunal distribution patterns in abyssal Pacific polymetallic nodule fields. PLoS ONE 2015, 10, e0117790. [Google Scholar] [CrossRef]
- Brix, S.; Bober, S.; Tschesche, C.; Kihara, T.C.; Driskell, A.; Jennings, R.M. Molecular species delimitation and its implications for species descriptions using desmosomatid and nannoniscid isopods from the VEMA fracture zone as example taxa. Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 148, 180–207. [Google Scholar] [CrossRef]
- Schnurr, S.; Osborn, K.J.; Malyutina, M.; Jennings, R.; Brix, S.; Driskell, A.; Svavarsson, J.; Martinez Arbizu, P. Hidden diversity in two species complexes of munnopsid isopods (Crustacea) at the transition between the northernmost North Atlantic and the Nordic Seas. Mar. Biodivers. 2018, 48, 813–843. [Google Scholar] [CrossRef]
- Bober, J.; Brandt, A.; Frutos, I.; Schwentner, M. Diversity and distribution of Ischnomesidae (Crustacea: Isopoda: Asellota) along the Kuril-Kamchatka trench—A genetic perspective. Prog. Oceanogr. 2019, 178, 102174. [Google Scholar] [CrossRef]
- Brix, S.; Held, C.; Kaiser, S.; Jennings, R.M.; Driskell, A.; Brandt, A. Evolution and phylogeny of the deep-sea isopod families Desmosomatidae Sars, 1897 and Nannoniscidae Hansen, 1916 (Isopoda: Asellota). Org. Divers. Evol. 2021, 21, 691–717. [Google Scholar] [CrossRef]
- Brix, S.; Svavarsson, J.; Leese, F. A multi-gene analysis reveals multiple highly divergent lineages of the isopod Chelator insignis (Hansen, 1916) south of Iceland. Pol. Polar Res. 2014, 2, 225–242. [Google Scholar] [CrossRef]
- Sota, T.; Kubota, K. Genital lock--and--key as a selective agent against hybridization. Evolution 1998, 52, 1507–1513. [Google Scholar] [CrossRef] [PubMed]
- Masly, J.P. 170 years of “lock-and-key”: Genital morphology and reproductive isolation. Int. J. Evol. Biol. 2012, 2012, 247352. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.D.F. Functional morphology and evolution of isopod genitalia. In Crustacean Sexual Biology, 1st ed.; Raymond, T.B., Joel, W.M., Eds.; Columbia University Press: New York, NY, USA, 1991; pp. 228–245. [Google Scholar] [CrossRef]
- Brown, W.L.; Wilson, E.O. Character displacement. Syst. Zool. 1956, 5, 49–64. [Google Scholar] [CrossRef]
- Blair, W.F. Character displacement in frogs. Am. Zool. 1974, 14, 1119–1125. [Google Scholar] [CrossRef]
- Melville, J. Competition and character displacement in two species of scincid lizards. Ecol. Lett. 2002, 5, 386–393. [Google Scholar] [CrossRef]
- Adams, D.C. Character displacement via aggressive interference in Appalachian salamanders. Ecology 2004, 85, 2664–2670. [Google Scholar] [CrossRef]
- Tsukagoshi, A. Reproductive character displacement in the ostracod genus Cythere. J. Crustac. Biol. 1988, 8, 563–575. [Google Scholar] [CrossRef]
- Kawano, K. Character displacement in giant rhinoceros beetles. Am. Nat. 2002, 159, 255–271. [Google Scholar] [CrossRef]
- Kawano, K. Character displacement in stag beetles (Coleoptera: Lucanidae). Ann. Entomol. Soc. Am. 2003, 96, 503–511. [Google Scholar] [CrossRef]
- Yamaguchi, S. Biogeographical history and morphological evolution of two closely related ostracod species, Ishizakiella ryukyuensis and I. miurensis. J. Crustac. Biol. 2003, 23, 623–632. [Google Scholar] [CrossRef]
- Wilson, G.D.F. The systematics and evolution of Haplomunna and its relatives (Isopoda, Haplomunnidae, new family). J. Nat. Hist. 1976, 10, 569–580. [Google Scholar] [CrossRef]
- Lins, L.S.; Ho, S.Y.; Wilson, G.D.F.; Lo, N. Evidence for Permo-Triassic colonization of the deep sea by isopods. Biol. Lett. 2012, 8, 979–982. [Google Scholar] [CrossRef] [PubMed]
















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Kim, J.; Lee, W.; Karanovic, I. Two New Uromunna Species (Isopoda: Asellota: Munnidae) from the Korean Peninsula and Their Phylogenetic Position within Munnoid Groups. Diversity 2023, 15, 20. https://doi.org/10.3390/d15010020
Kim J, Kim J, Lee W, Karanovic I. Two New Uromunna Species (Isopoda: Asellota: Munnidae) from the Korean Peninsula and Their Phylogenetic Position within Munnoid Groups. Diversity. 2023; 15(1):20. https://doi.org/10.3390/d15010020
Chicago/Turabian StyleKim, Jeongho, Jaehyun Kim, Wonchoel Lee, and Ivana Karanovic. 2023. "Two New Uromunna Species (Isopoda: Asellota: Munnidae) from the Korean Peninsula and Their Phylogenetic Position within Munnoid Groups" Diversity 15, no. 1: 20. https://doi.org/10.3390/d15010020
APA StyleKim, J., Kim, J., Lee, W., & Karanovic, I. (2023). Two New Uromunna Species (Isopoda: Asellota: Munnidae) from the Korean Peninsula and Their Phylogenetic Position within Munnoid Groups. Diversity, 15(1), 20. https://doi.org/10.3390/d15010020