

Brachiopod Fauna from the Deep Mediterranean Sea: Distribution Patterns and Ecological Preferences

 ,
, 
 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Morphology of the Target Species
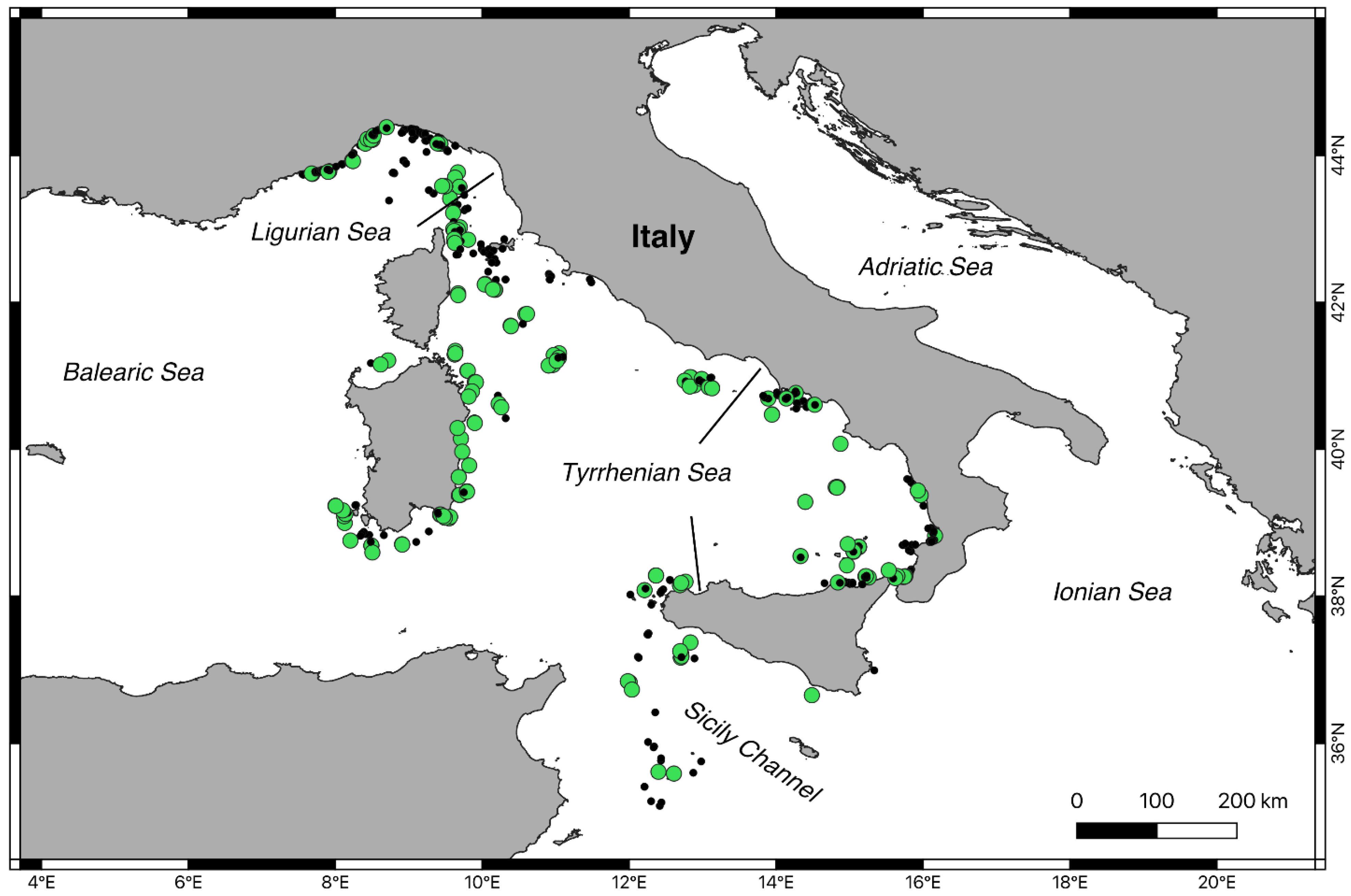
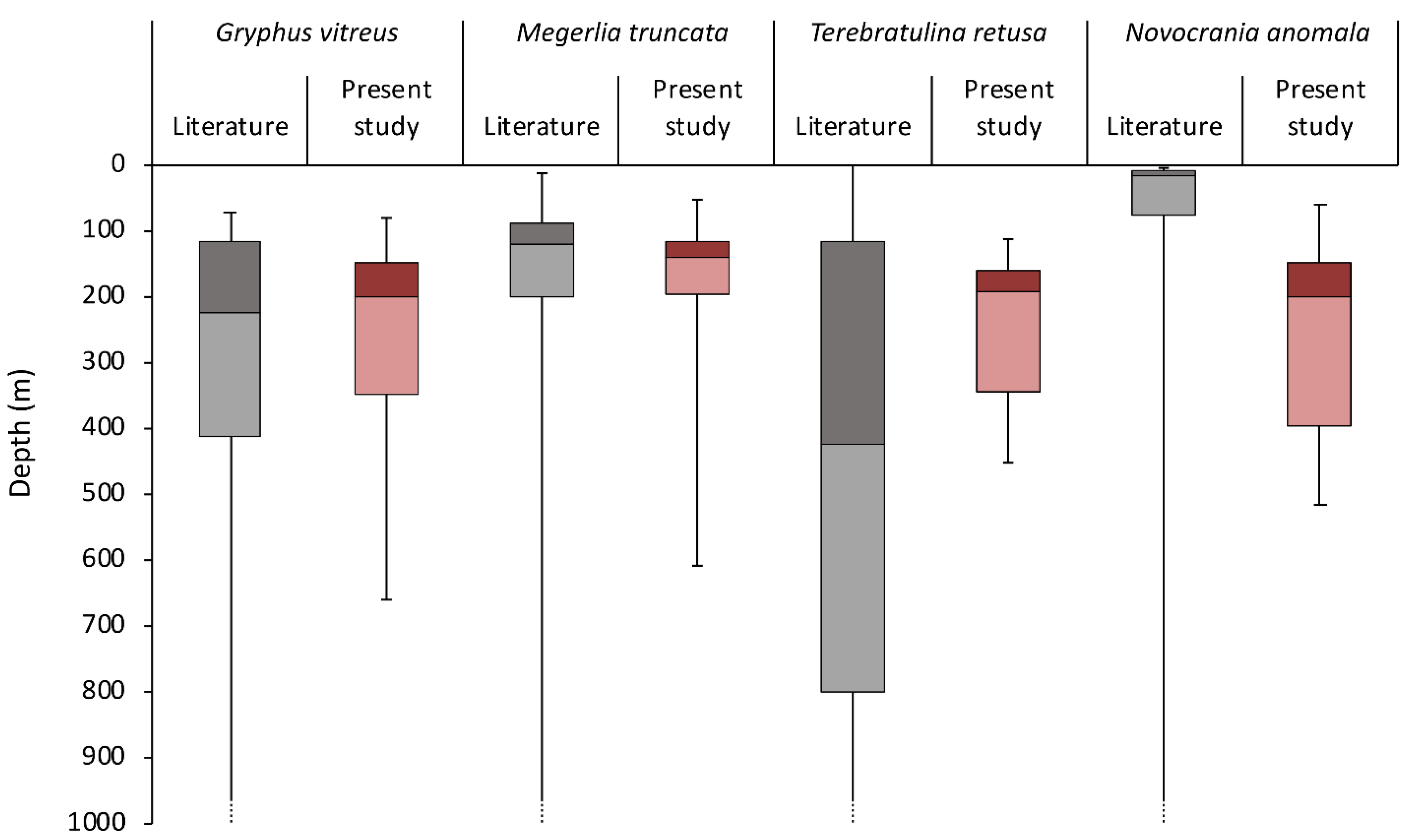
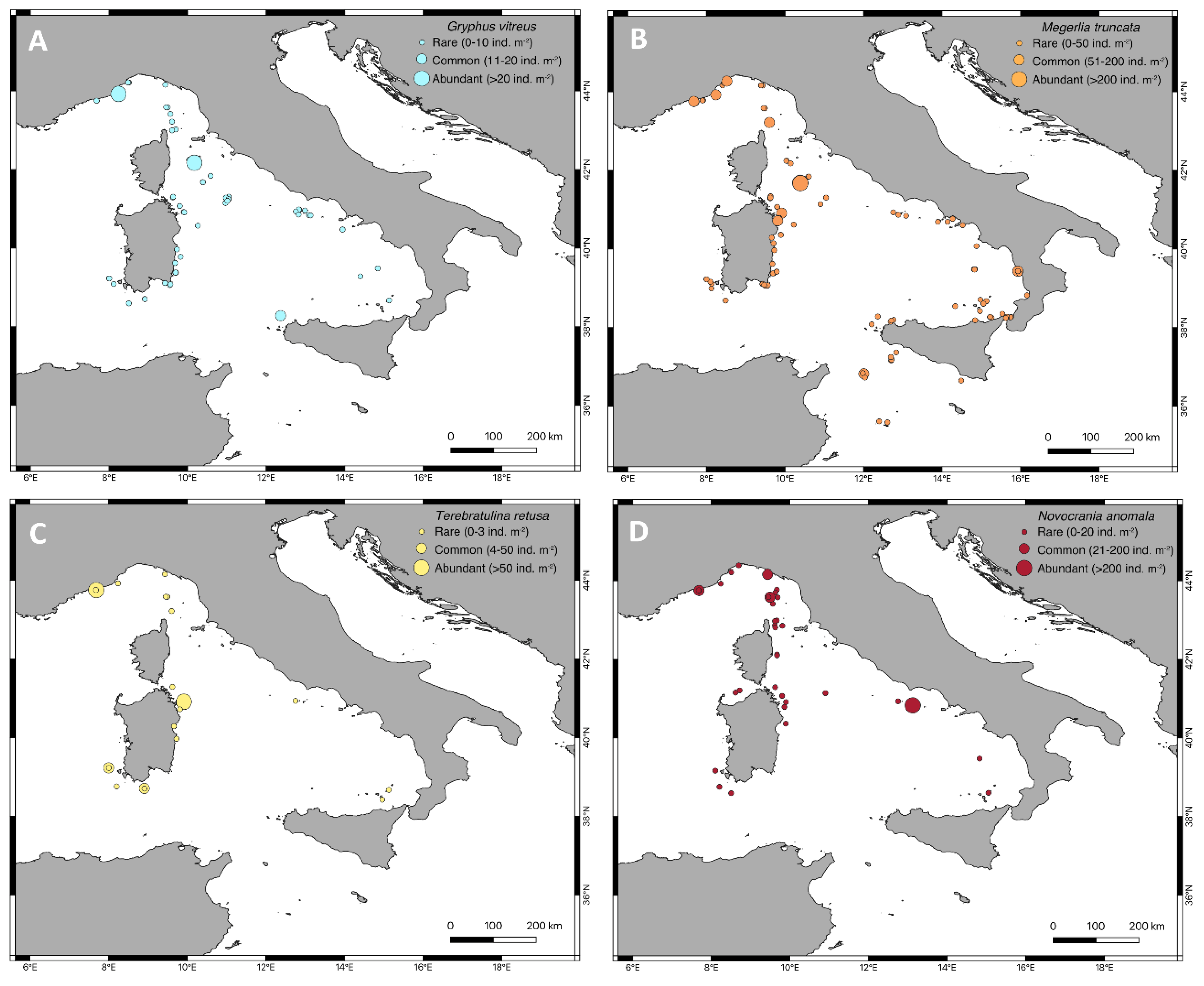
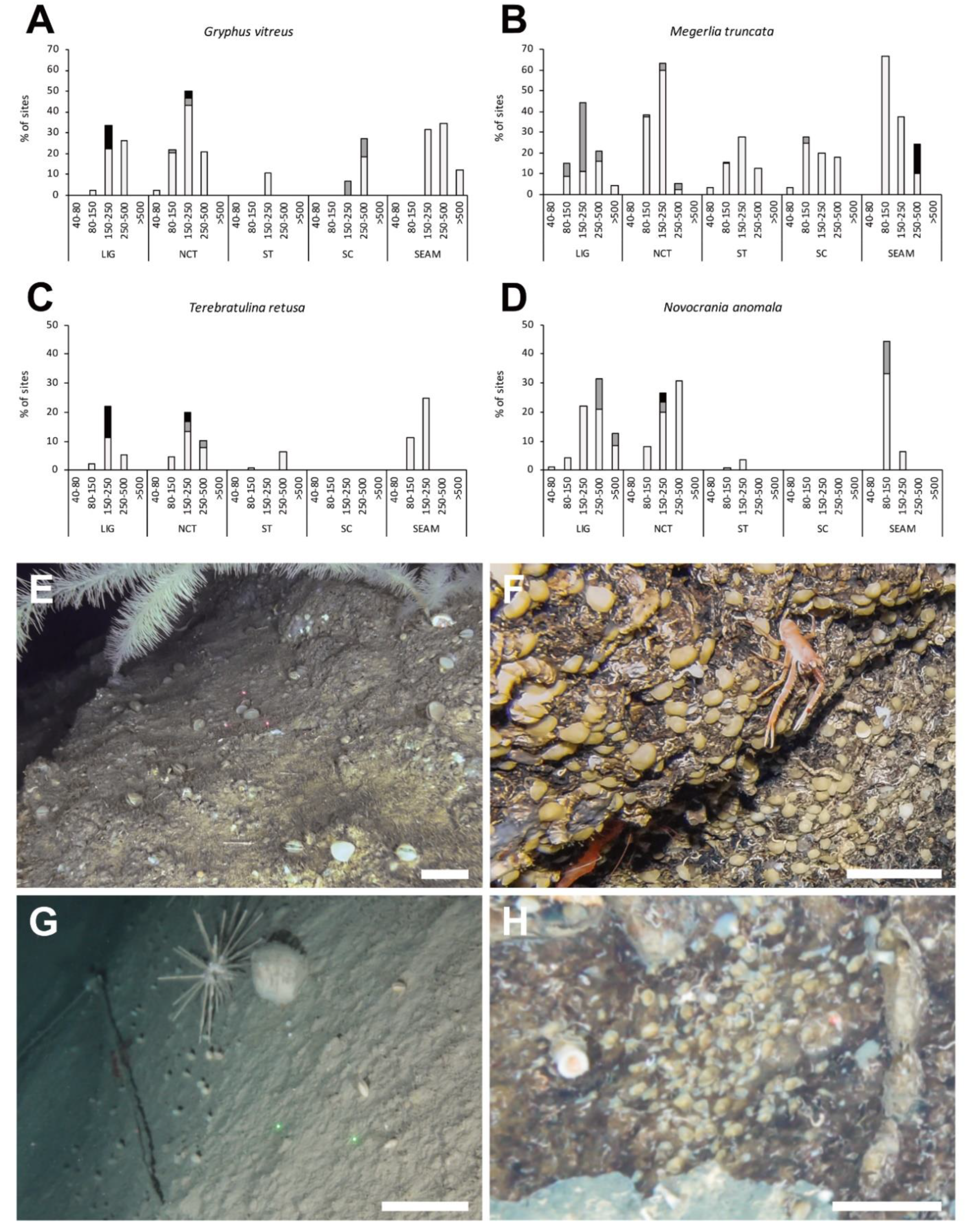
3.2. Geographic and Bathymetric Distribution
3.3. Population Density
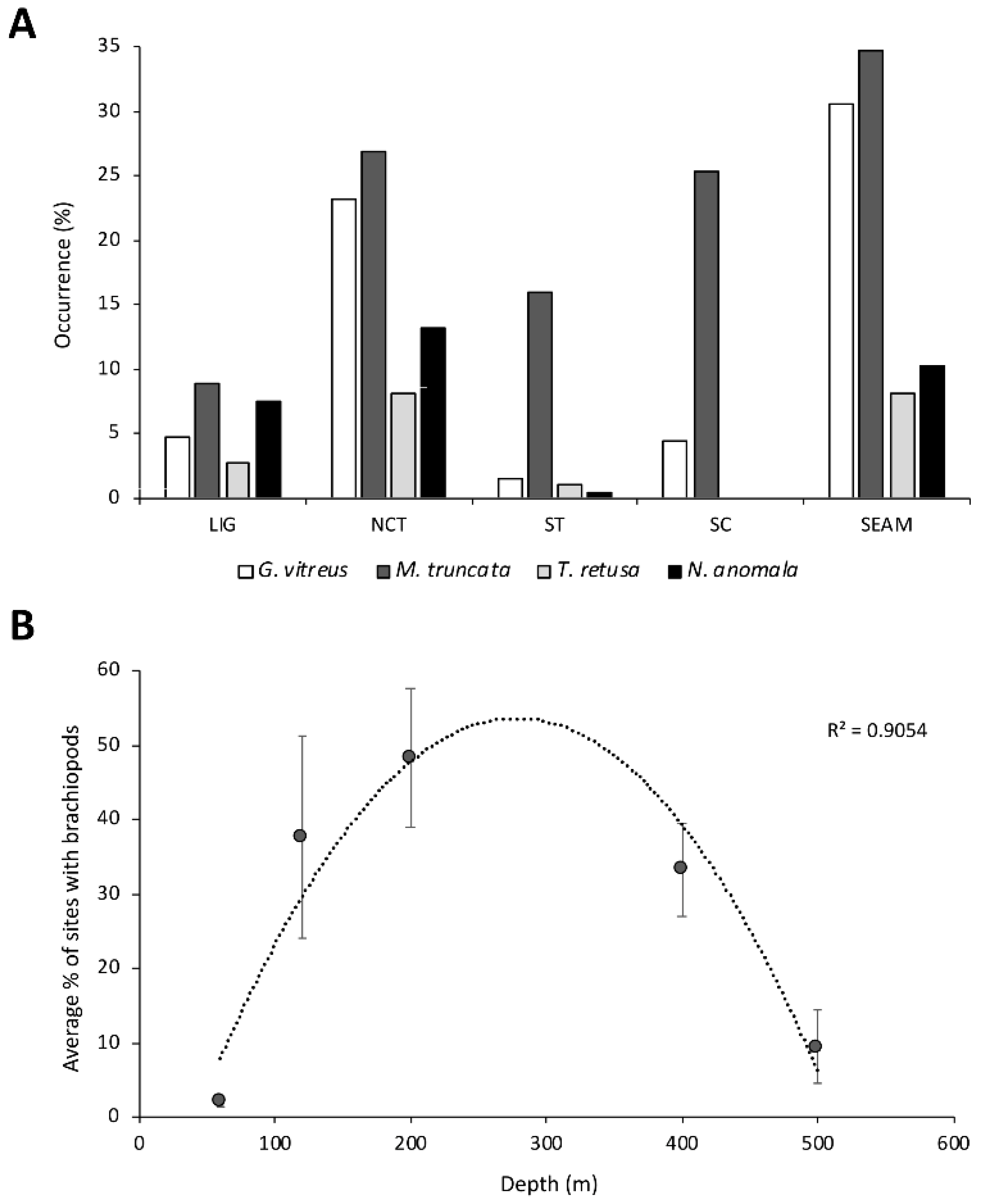
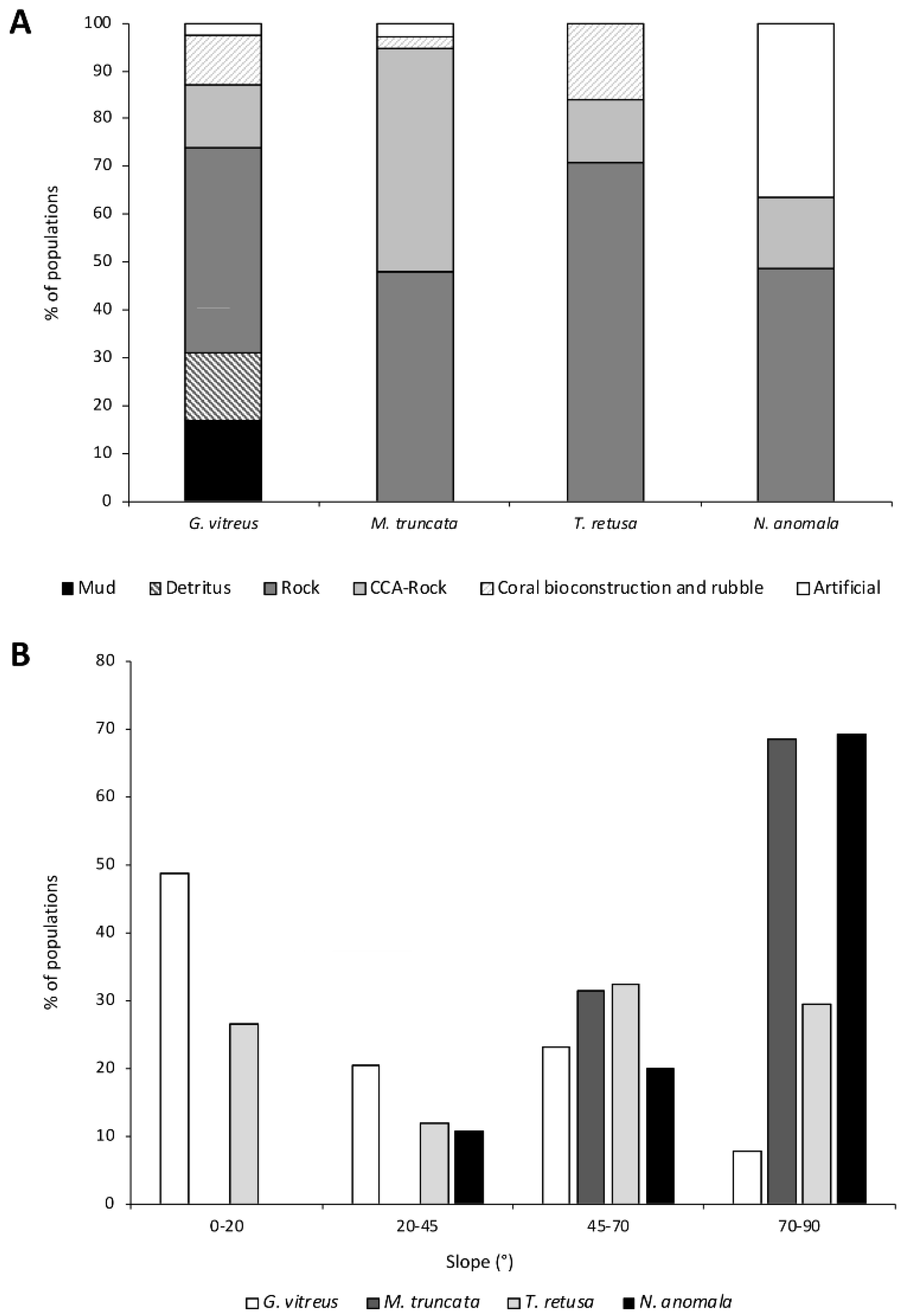
3.4. Ecological Preferences and Associated Fauna
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giribet, G.; Edgecombe, G.D. The Invertebrate Tree of Life; Princeton University Press: Princeton, NJ, USA, 2020; p. 54. [Google Scholar]
- Emig, C.C.; Bitner, M.A.; Álvarez, F. Phylum Brachiopoda. Animal Biodiversity: An Outline of Higher-level Classification and Survey of Taxonomic Richness (Addenda 2013). Zootaxa 2013, 3703, 75–78. [Google Scholar] [CrossRef]
- Zezina, O.N. Global surface-water circulation and the main features of brachiopod biogeography. In Brachiopods Past and Present; Brunton, C.H.C., Cocks, L.R.M., Long, S.L., Eds.; The Systematics Association Special Volume Series; Taylor & Francis: Oxfordshire, UK, 2001; Volume 63, pp. 102–107. [Google Scholar]
- Zezina, O.N. Biogeography of the recent brachiopods. Paléontol. J. 2008, 42, 830–858. [Google Scholar] [CrossRef]
- Rosso, A.; Sanfilippo, R.; Taddei Ruggiero, E.; Di Martino, E. Faunas and ecological groups of Serpuloidea, Bryozoa and Brachiopoda from submarine caves in Sicily (Mediterranean Sea). Boll. Della Soc. Paleontol. Ital. 2013, 52, 167–176. [Google Scholar]
- Rosso, A.; Sanfilippo, R.; Guido, A.; Gerovasileiou, V.; Taddei Ruggiero, E.; Belmonte, G. Colonisers of the dark: Biostalac-tite-associated metazoans from “lu Lampiùne” submarine cave (Apulia, Mediterranean Sea). Mar. Ecol. 2021, 42, e12634. [Google Scholar] [CrossRef]
- Radolović, M.; Bakran-Petricioli, T.; Petricioli, D.; Surić, M.; Perica, D. Biological response to geochemical and hydrological processes in a shallow submarine cave. Mediterr. Mar. Sci. 2015, 16, 305. [Google Scholar] [CrossRef]
- Bergamin, L.; Ruggiero, E.T.; Pierfranceschi, G.; Andres, B.; Constantino, R.; Crovato, C.; D’Ambrosi, A.; Marassich, A.; Romano, E. Benthic foraminifera and brachiopods from a marine cave in Spain: Environmental significance. Mediterr. Mar. Sci. 2020, 21, 506–518. [Google Scholar] [CrossRef]
- Bitner, M.A.; Gerovasileiou, V. Taxonomic composition and assemblage structure of brachiopods from two submarine caves in the Aegean Sea, Eastern Mediterranean. Eur. Zool. J. 2021, 88, 316–327. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Bianchi, C.N. Mediterranean Marine Caves: A Synthesis of Current Knowledge. Oceanogr. Mar. Biol. Annu. Rev. 2021, 59, 1–87. [Google Scholar] [CrossRef]
- Asgaard, U.; Bromley, R.G. Population dynamics and autecology of ‘Rhynchonella’triangularis, a Late. Brachiopods Through Time 1991, 247. [Google Scholar]
- Taddei Ruggiero, E. Brachiopods from bio-and thanatocoenoses of the Isca submarine cave (Sorrento Peninsula). Boll. Soc. Paleontol. Ital. 1994, 33, 313–324. [Google Scholar]
- Taddei Ruggiero, E. Functional morphology with reference to substrate connections in Neocrania and to feeding mecha-nisms in Anisactinella and Amphitomella (Brachiopoda). Autecology Sel. Foss. Org. Achiev. Probl. 1996, 223. [Google Scholar]
- Ye, F.; Shi, G.R.; Bitner, M.A. Global biogeography of living brachiopods: Bioregionalization patterns and possible controls. PLoS ONE 2021, 16, e0259004. [Google Scholar] [CrossRef] [PubMed]
- Fürsich, F.T.; Hurst, J.M. Environmental factors determining the distribution of brachiopods. Palaeontology 1974, 17, 879–900. [Google Scholar]
- Emig, C.C. Les Brachiopodes actuels sont-ils des indicateurs (paléo) bathymétriques? Géologie Méditerranéenne 1988, 15, 65–71. [Google Scholar] [CrossRef]
- Emig, C.C. Distributional patterns along the mediterranean continental margin (upper bathyal) using gryphus vitreus (Brachiopoda) densities. Palaeogeogr. Palaeoclim. Palaeoecol. 1989, 71, 253–256. [Google Scholar] [CrossRef]
- Tunnicliffe, V.; Wilson, K. Brachiopod populations: Distribution in fjords of British Columbia (Canada) and tolerance of low oxygen concentrations. Mar. Ecol. Prog. Ser. 1988, 47, 117–128. [Google Scholar] [CrossRef]
- Gordillo, S.; Muñoz, D.F.; Bayer, M.S.; Malvé, M.E. How physical and biotic factors affect brachiopods from the Patagonian Continental Shelf. J. Mar. Syst. 2018, 187, 223–234. [Google Scholar] [CrossRef]
- Logan, A. The Recent Brachiopoda of the Mediterranean Sea. Bull. De L’institut Océanographique Monaco 1979, 72, 1–112. [Google Scholar]
- Logan, A.; Bianchi, C.N.; Morri, C.; Zibrowius, H.; Bitar, G. New records of Recent brachiopods from the eastern Mediter-ranean Sea. Ann. Del Mus. Civ. Di Stor. Nat. “G. Doria” 2002, 94, 407–418. [Google Scholar]
- Logan, A.; Bianchi, C.N.; Morri, C.; Zibrowius, H. The present-day Mediterranean brachiopod fauna: Diversity, life habits, biogeography and paleobiogeography. Sci. Mar. 2004, 68, 163–170. [Google Scholar] [CrossRef][Green Version]
- Emig, C.C. Novocrania turbinata synonyme de N. anomala. Carnets De Géologie 2014, 14, 159–171. [Google Scholar] [CrossRef]
- Simon, E.; Willems, G. Gwynia capsula (Jeffreys, 1859) and other Recent brachiopods from submarine caves in Croatia. Bulle-Tin De L’institut R. Des Sci. Nat. De Belg. Biol. 1999, 69, 15–21. [Google Scholar]
- Logan, A.; Zibrowius, H. A New Genus and Species of Rhynchonellid (Brachiopoda, Recent) from Submarine Caves in the Mediterranean Sea. Mar. Ecol. 1994, 15, 77–88. [Google Scholar] [CrossRef]
- Emig, C.C. Brachiopodes récoltés lors de campagnes (1976-2014) dans l’étage Bathyal des côtes françaises méditerranéennes. Redéfinition des limites du système phytal dans le domaine marin benthique. Carnets De Géologie 2018, CG2018_B01, 1–100. [Google Scholar]
- Montefalcone, M.; Tunesi, L.; Ouerghi, A. A review of the classification systems for marine benthic habitats and the new updated Barcelona Convention classification for the Mediterranean. Mar. Environ. Res. 2021, 169, 105387. [Google Scholar] [CrossRef]
- García-Ramos, D.A.; Ćorić, S.; Joachimski, M.M.; Zuschin, M. The environmental factors limiting the distribution of shal-low-water terebratulid brachiopods. Paleobiology 2020, 46, 193–217. [Google Scholar] [CrossRef]
- Cocks, L.R.M.; Popov, L.E. The identity and significance of the high-latitude Early Ordovician Mediterranean brachiopod Province. Geol. Mag. 2021, 158, 2187–2208. [Google Scholar] [CrossRef]
- Dulai, A. New data on the late Miocene brachiopod fauna of Tetti Borelli (Piedmont, N Italy). Riv. Ital. Di Paleontol. E Stratigr. 2019, 125. [Google Scholar]
- Emig, C.C. Offshore brachiopods investigated by submersible. J. Exp. Mar. Biol. Ecol. 1987, 108, 261–273. [Google Scholar] [CrossRef]
- Emig, C.C.; García-Carrascosa, M.A. Distribution of Gryphus vitreus (Born, 1778) (Brachiopoda) on transect P2 (Continental margin, French Mediterranean coast) investigated by submersible. Sci. Mar. 1991, 55, 385–388. [Google Scholar]
- Angeletti, L.; Canese, S.; Cardone, F.; Castellan, G.; Foglini, F.; Taviani, M. A brachiopod biotope associated with rocky bottoms at the shelf break in the central Mediterranean Sea: Geobiological traits and conservation aspects. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 30, 402–411. [Google Scholar] [CrossRef]
- Bo, M.; Canese, S.; Spaggiari, C.; Pusceddu, A.; Bertolino, M.; Angiolillo, M.; Giusti, M.; Loreto, M.F.; Salvati, E.; Greco, S.; et al. Deep Coral Oases in the South Tyrrhenian Sea. PLoS ONE 2012, 7, e49870. [Google Scholar] [CrossRef] [PubMed]
- Bo, M.; Coppari, M.; Betti, F.; Enrichetti, F.; Bertolino, M.; Massa, F.; Bava, S.; Gay, G.; Cattaneo-Vietti, R.; Bavestrello, G. The high biodiversity and vulnerability of two Mediterranean bathyal seamounts support the need for creating offshore pro-tected areas. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 543–566. [Google Scholar] [CrossRef]
- Cau, A.; Follesa, M.C.; Moccia, D.; Alvito, A.; Bo, M.; Angiolillo, M.; Canese, S.; Paliaga, E.M.; Orrù, P.E.; Sacco, F.; et al. Deepwater corals biodiversity along roche du large ecosystems with different habitat complexity along the south Sardinia continental margin (CW Mediterranean Sea). Mar. Biol. 2015, 162, 1865–1878. [Google Scholar] [CrossRef]
- Enrichetti, F.; Dominguez-Carrió, C.; Toma, M.; Bavestrello, G.; Betti, F.; Canese, S.; Bo, M. Megabenthic communities of the Ligurian deep continental shelf and shelf break (NW Mediterranean Sea). PLoS ONE 2019, 14, e0223949. [Google Scholar] [CrossRef]
- Angiolillo, M.; La Mesa, G.; Giusti, M.; Salvati, E.; Di Lorenzo, B.; Rossi, L.; Canese, S.; Tunesi, L. New records of scleractini-an cold-water coral (CWC) assemblages in the southern Tyrrhenian Sea (western Mediterranean Sea): Human impacts and conservation prospects. Prog. Oceanogr. 2021, 197, 102656. [Google Scholar] [CrossRef]
- Consoli, P.; Altobelli, C.; Perzia, P.; Bo, M.; Rosso, A.; Alongi, G.; Serio, D.; Canese, S.; Romeo, T.; Andaloro, F. Species and habitats of conservation interest in the Ecologically and Biologically Significant Area of the Strait of Sicily: A contribution towards the creation of a Specially Protected Areas of Mediterranean Importance. Mediterr. Mar. Sci. 2021, 22, 297–316. [Google Scholar] [CrossRef]
- Moccia, D.; Cau, A.; Bramanti, L.; Carugati, L.; Canese, S.; Follesa, M.C.; Cannas, R. Spatial distribution and habitat charac-terization of marine animal forest assemblages along nine submarine canyons of Eastern Sardinia (central Mediterranean Sea). Deep. Sea Res. Part I: Oceanogr. Res. Pap. 2021, 167, 103422. [Google Scholar] [CrossRef]
- SPA/RAC–UN Environment/MAP (2019) Updated Classification of Benthic Marine Habitat Types for the Mediterranean Region. Available online: https://www.rac-spa.org/sites/default/files/doc_fsd/habitats_list_en.pdf (accessed on 14 July 2022).
- Fourt, M.; Goujard, A.; Perez, T.; Chevaldonné, P. Guide de la Faune Profonde de la mer Méditerranée. Exploration des Roches et Canyons Sous-marins des côtes Françaises; Muséum National d’Histoire Naturelle: Paris, France, 2017; p. 184. [Google Scholar]
- Anadón, N.; Fernández-Rodríguez, I.; Arias, A. Recent brachiopod diversity and distribution from the central Cantabrian Sea and the Avilés Canyons System (Bay of Biscay). Estuarine, Coast. Shelf Sci. 2022, 269, 107829. [Google Scholar] [CrossRef]
- Marine Species Identification Portal. Available online: http://www.specie-identification.org (accessed on 14 July 2022).
- Toma, M.; Betti, F.; Bavestrello, G.; Cattaneo-Vietti, R.; Canese, S.; Cau, A.; Andaloro, F.; Greco, S.; Bo, M. Diversity and abundance of heterobranchs (Mollusca, Gastropoda) from the mesophotic and bathyal zone of the Mediterranean Sea. Eur. Zool. J. 2022, 89, 167–189. [Google Scholar] [CrossRef]
- Toma, M.; Bo, M.; Cattaneo-Vietti, R.; Canese, S.; Canessa, M.; Cannas, R.; Cardone, F.; Carugati, L.; Cau, A.; Corriero, G.; et al. Basin-scale occurrence and distribution of mesophotic and upper bathyal red coral forests along the Italian coasts. Medi-Terranean Mar. Sci. 2022, 89, 167–189. [Google Scholar]
- Curry, G.B.; Endo, K. Migration of brachiopod species in the North Atlantic in response to Holocene climatic change. Geology 1991, 19, 1101. [Google Scholar] [CrossRef]
- Emig, C.C.; Geistdoerfer, P. The Mediterranean deep-sea fauna: Historical evolution, bathymetric variations and geographical changes. arXiv, 2005; arXiv:q-bio/0507003. [Google Scholar]
- Bianchi, C.N.; Morri, C.; Chiantore, M.; Montefalcone, M.; Parravicini, V.; Rovere, A. Mediterranean Sea biodiversity between the legacy from the past and a future of change. Life Mediterr. Sea A Look Habitat Chang. 2012, 1, 55. [Google Scholar]
- Enrichetti, F.; Bavestrello, G.; Coppari, M.; Betti, F.; Bo, M. Placogorgia coronate first documented record in Italian waters: Use of trawl bycatch to unveil vulnerable deep-sea ecosystems. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 1123–1138. [Google Scholar] [CrossRef]
- Sartoretto, S.; Zibrowius, H. Note on new records of living Scleractinia and Gorgonaria between 1700 and 2200 m depth in the western Mediterranean Sea. Mar. Biodivers. 2017, 48, 689–694. [Google Scholar] [CrossRef]
- Bo, M.; Coppari, M.; Betti, F.; Massa, F.; Gay, G.; Cattaneo-Vietti, R.; Bavestrello, G. Unveiling the deep biodiversity of the Janua Seamount (Ligurian Sea): First Mediterranean sighting of the rare Atlantic bamboo coral Chelidonisis aurantiaca Studer, 1890. Deep Sea Res. Part Oceanogr. Res. Pap. 2019, 156, 103186. [Google Scholar] [CrossRef]
- Hiller, N. The environment, biogeography, and origin of the southern African Recent brachiopod fauna. J. Paléontol. 1994, 68, 776–786. [Google Scholar] [CrossRef]
- Kowalewski, M.; Simões, M.G.; Carroll, M.; Rodland, D.L. Abundant Brachiopods on a Tropical, Upwelling-Influenced Shelf (Southeast Brazilian Bight, South Atlantic). Palaios 2002, 17, 277–286. [Google Scholar] [CrossRef]
- Baumgarten, S.; Laudien, J.; Jantzen, C.; Häussermann, V.; Försterra, G. Population structure, growth and production of a recent brachiopod from the Chilean fjord region. Mar. Ecol. 2013, 35, 401–413. [Google Scholar] [CrossRef]
- Álvarez, F.; Tréguier, J.; Emig, C.C. Brachiopodes actuels: Historique et révision de la collection D.-P. Œhlert (Laval); bra-chiopodes des côtes françaises métropolitaines. Carnets De Géologie 2017, 386. [Google Scholar]
- Vacelet, J.; Boury-Esnault, N.; Harmelin, J.G. Hexactinellid cave, a unique deep-sea habitat in the scuba zone. Deep. Sea Re-Search Part I Oceanogr. Res. Pap. 1994, 41, 965–973. [Google Scholar] [CrossRef]
- Tsiamis, K.; Aydogan, O.; Bailly, N.; Balistreri, P.; Bariche, M.; Carden-Noad, S.; Corsini-Foka, M.; Crocetta, F.; Davidov, B.; Dimitriadis, C.; et al. New Mediterranean Biodiversity Records (July 2015). Mediterr. Mar. Sci. 2015, 16, 472–488. [Google Scholar] [CrossRef]
- Vacelet, J.; Boury-Esnault, N. A new species of carnivorous sponge (Demospongiae: Cladorhizidae) from a Mediterranean cave. Bull. Inst. R. Sci. Nat. Belg. Biol. 1996, 66, 109–115. [Google Scholar]
- Taviani, M.; Angeletti, L.; Canese, S.; Cannas, R.; Cardone, F.; Cau, A.; Follesa, M.; Marchese, F.; Montagna, P.; Tessarolo, C. The “Sardinian cold-water coral province” in the context of the Mediterranean coral ecosystems. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 145, 61–78. [Google Scholar] [CrossRef]
- James, M.A.; Ansell, A.D.; Collins, M.J.; Curry, G.B.; Peck, L.S.; Rhodes, M.C. Biology of living brachiopods. In Advances in Marine Biology; Academic Press: Cambridge, MA, USA, 1992; Volume 28, p. 175. [Google Scholar]
- Álvarez, F.; Emig, C.C. Brachiopoda from the Luso-Iberian zone. I. Biology and ecology. In Proceedings of the Fourth, Millennium, International Brachiopod Congress, London, UK, 10–14 July 2000. [Google Scholar]
- Enrichetti, F.; Bavestrello, G.; Betti, F.; Rindi, F.; Tregrosso, A.; Bo, M. Fate of lost fishing gears: Experimental evidence of biofouling colonization patterns from the northwestern Mediterranean Sea. Environ. Pollut. 2020, 268, 115746. [Google Scholar] [CrossRef]
- Ramirez-Llodra, E.; De Mol, B.; Company, J.B.; Coll, M.; Sardà, F. Effects of natural and anthropogenic processes in the dis-tribution of marine litter in the deep Mediterranean Sea. Prog. Oceanogr. 2013, 118, 273–287. [Google Scholar] [CrossRef]
- Song, X.; Lyu, M.; Zhang, X.; Ruthensteiner, B.; Ahn, I.-Y.; Pastorino, G.; Wang, Y.; Gu, Y.; Ta, K.; Sun, J.; et al. Large Plastic Debris Dumps: New Biodiversity Hot Spots Emerging on the Deep-Sea Floor. Environ. Sci. Technol. Lett. 2021, 8, 148–154. [Google Scholar] [CrossRef]
- Brennan, M.L.; Ballard, R.D. Deep-water ancient shipwrecks of the Mediterranean, Aegean, and Black Seas: 1988. Oceanography 2013, 26 (Suppl. S1), 22–25. [Google Scholar]
- Secci, M.; Demesticha, S.; Jimenez, C.; Papadopoulou, C.; Katsouri, I. A LIVING SHIPWRECK: An integrated three-dimensional analysis for the understanding of site formation processes in archaeological shipwreck sites. J. Archaeol. Sci. Rep. 2020, 35, 102731. [Google Scholar] [CrossRef]
- Wisshak, M.; López Correa, M.; Gofas, S.; Salas, C.; Taviani, M.; Jakobsen, J.; Freiwald, A. Shell architecture, element composition, and stable isotope signature of the giant deep-sea oyster Neopycnodonte zibrowii sp. n. from the NE Atlantic. Deep Sea Res. Part I 2009, 56, 374–407. [Google Scholar] [CrossRef]
- Aguilar, R.; Correa, M.L.; Calcinai, B.; Pastor, X.; De la Torriente, A.; Garcia, S. First records of Asbestopluma hypogea Vacelet and Boury-Esnault, 1996 (Porifera, Demospongiae Cladorhizidae) on seamounts and in bathyal settings of the Mediterra-nean Sea. Zootaxa 2011, 2925, 33–40. [Google Scholar] [CrossRef]
- Buhl-Mortensen, L.; Vanreusel, A.; Gooday, A.J.; Levin, L.; Priede, I.; Mortensen, P.; Gheerardyn, H.; King, N.J.; Raes, M. Biological structures as a source of habitat heterogeneity and biodiversity on the deep ocean margins. Mar. Ecol. 2010, 31, 21–50. [Google Scholar] [CrossRef]
- Maldonado, M.; Aguilar, R.; Bannister, R.; Bell, J.; Conway, J.; Dayton, P.; Diaz, P.; Gutt, C.; Kelly, J.; Kenchington, M.; et al. Sponge grounds as key marine habitats: A synthetic review of types, structure, functional roles, and conservation concerns. In Marine animal forests: The Ecology of Benthic Biodiversity Hotspots; Springer: Berlin, Germany, 2017. [Google Scholar]
- Gori, A.; Bavestrello, G.; Grinyó, J.; Dominguez-Carrió, C.; Ambroso, S.; Bo, M. Animal Forests in Deep Coastal Bottoms and Continental Shelf of the Mediterranean Sea. In Marine animal forests: The Ecology of Benthic Biodiversity Hotspots; Springer: Berlin, Germany, 2017; pp. 1–27. [Google Scholar] [CrossRef]
- Delance, J.H.; Emig, C.C. Drilling predation on Gryphus vitreus (Brachiopoda) off the French Mediterranean coasts. Palaeogeogr. Palaeoclim. Palaeoecol. 2004, 208, 23–30. [Google Scholar] [CrossRef]
- Ruggiero, E.T.; Buono, G.; Raia, P. Bioerosion on Brachiopod Shells of a Thanatocoenosis of Alboràn Sea (Spain). Ichnos 2006, 13, 175–184. [Google Scholar] [CrossRef]
- Tyler, C.L.; Leighton, L.R.; Carlson, S.J.; Huntley, J.W.; Kowalewski, M. Predation on Modern and Fossil brachiopods: As-sessing chemical defenses and palatability. Palaios 2013, 28, 724–735. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Coastal Macro-Areas | Offshore Reliefs | TOT | |||
|---|---|---|---|---|---|---|
| LIG | NCT | ST | SC | SEAM | ||
| CRANIIDA | ||||||
| Craniidae | ||||||
| Novocrania anomala (Müller, 1776) | 11 | 21 | 1 | 0 | 5 | 38 |
| TEREBRATULIDA | ||||||
| Terebratulidae | ||||||
| Gryphus vitreus (Born, 1778) | 7 | 37 | 3 | 3 | 15 | 65 |
| Kraussinidae | ||||||
| Megerlia truncata (Linnaeus, 1767) | 13 | 43 | 32 | 17 | 17 | 122 |
| Cancellothyrididae | ||||||
| Terebratulina retusa (Linnaeus, 1758) | 4 | 13 | 2 | 0 | 4 | 23 |
| Macro-Area | Depth Range (m) | N. anomala | G. vitreus | M. truncata | T. retusa | ||||||||
| 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 40–80 | 1 | ||||||||||||
| 80–150 | 2 | 1 | 4 | 3 | 1 | ||||||||
| LIG | 150–250 | 2 | 2 | 1 | 1 | 3 | 1 | 1 | |||||
| 250–500 | 4 | 2 | 5 | 3 | 1 | 1 | |||||||
| >500 | 2 | 1 | 1 | ||||||||||
| 40–80 | |||||||||||||
| 80–150 | 7 | 18 | 1 | 33 | 1 | 4 | |||||||
| NCT | 150–250 | 6 | 1 | 1 | 13 | 1 | 1 | 18 | 1 | 4 | 1 | 1 | |
| 250–500 | 12 | 8 | 1 | 1 | 3 | 1 | |||||||
| >500 | |||||||||||||
| 40–80 | 3 | ||||||||||||
| 80–150 | 1 | 23 | 1 | 1 | |||||||||
| ST | 150–250 | 1 | 3 | 8 | |||||||||
| 250–500 | 2 | 1 | |||||||||||
| >500 | |||||||||||||
| 40–80 | 1 | ||||||||||||
| 80–150 | 10 | 1 | |||||||||||
| SC | 150–250 | 1 | 3 | ||||||||||
| 250–500 | 2 | 1 | 2 | ||||||||||
| >500 | |||||||||||||
| 40–80 | |||||||||||||
| 80–150 | 3 | 1 | 6 | 1 | |||||||||
| SEAM | 150–250 | 1 | 5 | 6 | 4 | ||||||||
| 250–500 | 10 | 3 | 4 | ||||||||||
| >500 | 2 | ||||||||||||
| Species | Density Value | Density Category | ||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 1 | 2 | 3 | |
| Novocrania anomala | 10.0 ± 1.3 | 52.5 ± 7.0 | 271.7 ± 61.7 | 0–20 | 21–200 | >200 |
| Gryphus vitreus | 4.3 ± 0.3 | 13.1 ± 1.0 | 23.8 ± 0.8 | 0–10 | 11–20 | >20 |
| Megerlia truncata | 23.2 ± 1.9 | 103.5 ± 5.2 | 426.7 ± 133.0 | 0–50 | 51–200 | >200 |
| Terebratulina retusa | 1.3 ± 0.3 | 11.1 ± 1.8 | nd | 0–3 | 4–50 | >50 |
| Associated Megabenthic Invertebrates | Brachiopod Species | |||||
|---|---|---|---|---|---|---|
| Phylum | Class | Taxon | N. anomala | G. vitreus | M. truncata | T. retusa |
| Porifera | Demospongiae | Hamacantha (Vomerula) falcula (Bowerbank, 1874) | + | + | + | + |
| Lycopodina hypogea (Vacelet & Boury-Esnault, 1996) | + | |||||
| Pachastrella monilifera Schmidt, 1868 | + | + | + | |||
| Poecillastra compressa (Bowerbank, 1866) | + | + | + | |||
| Polymastia polytylota Vacelet, 1969 | + | |||||
| Rhabderemia sp. | + | + | + | |||
| Cnidaria | Anthozoa | Corallium rubrum (Linnaeus, 1758) | + | + | ||
| Bebryce mollis Philippi, 1842 | + | |||||
| Callogorgia verticillata (Pallas, 1766) | + | |||||
| Eunicella cavolini (Koch, 1887) | + | |||||
| Swiftia dubia (Thomson, 1929) | + | |||||
| Caryophyllia (Caryophyllia) cyathus (Ellis & Solander, 1786) | + | + | ||||
| Caryophyllia (Caryophyllia) smithii Stokes & Broderip, 1828 | + | + | ||||
| Dendrophyllia cornigera (Lamarck, 1816) | + | + | + | |||
| Desmophyllum dianthus (Esper, 1794) | + | |||||
| Madrepora oculata Linnaeus, 1758 | + | + | ||||
| Thalamophyllia gasti (Döderlein, 1913) | + | + | ||||
| Antipathes dichotoma Pallas, 1766 | + | |||||
| Leiopathes glaberrima (Esper, 1792) | + | + | + | |||
| Parantipathes larix (Esper, 1788) | + | + | ||||
| Annelida | Polychaeta | Apomatus/Protula complex | + | + | ||
| Bonellia viridis Rolando, 1822 | + | |||||
| Filogranula annulata(Costa, 1861) | + | + | ||||
| Mollusca | Bivalvia | Neopycnodonte cochlear (Poli, 1795) | + | |||
| Neopycnodonte zibrowii Gofas, Salas & Taviani, 2009 (fossil) | + | |||||
| Crustacea | Malacostraca | Munida tenuimana Sars, 1872 | + | |||
| Munida spp. | + | + | ||||
| Paromola cuvieri (Risso, 1816) | + | |||||
| Plesionika giglioli (Senna, 1902) | + | + | ||||
| Plesionika narval (Fabricius, 1787) | + | + | ||||
| Echinodermata | Crinoidea | Leptometra phalangium (Müller, 1841) | + | |||
| Asteroidea | Hacelia attenuata Gray, 1840 | + | + | |||
| Echinoidea | Cidaris cidaris (Linnaeus, 1758) | + | + | + | + | |
| Echinus melo Lamarck, 1816 | + | + | ||||
| Stylocidaris affinis (Philippi, 1845) | + | |||||
| Ophiuroidea | Ophiacantha setosa (Bruzelius, 1805) | + | + | |||
| Holothuroidea | Holothuria (Roweothuria) poli Delle Chiaje, 1824 | + | + | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toma, M.; Enrichetti, F.; Bavestrello, G.; Canese, S.; Cau, A.; Andaloro, F.; Angiolillo, M.; Greco, S.; Bo, M. Brachiopod Fauna from the Deep Mediterranean Sea: Distribution Patterns and Ecological Preferences. Diversity 2022, 14, 753. https://doi.org/10.3390/d14090753
Toma M, Enrichetti F, Bavestrello G, Canese S, Cau A, Andaloro F, Angiolillo M, Greco S, Bo M. Brachiopod Fauna from the Deep Mediterranean Sea: Distribution Patterns and Ecological Preferences. Diversity. 2022; 14(9):753. https://doi.org/10.3390/d14090753
Chicago/Turabian StyleToma, Margherita, Francesco Enrichetti, Giorgio Bavestrello, Simonepietro Canese, Alessandro Cau, Franco Andaloro, Michela Angiolillo, Silvestro Greco, and Marzia Bo. 2022. "Brachiopod Fauna from the Deep Mediterranean Sea: Distribution Patterns and Ecological Preferences" Diversity 14, no. 9: 753. https://doi.org/10.3390/d14090753
APA StyleToma, M., Enrichetti, F., Bavestrello, G., Canese, S., Cau, A., Andaloro, F., Angiolillo, M., Greco, S., & Bo, M. (2022). Brachiopod Fauna from the Deep Mediterranean Sea: Distribution Patterns and Ecological Preferences. Diversity, 14(9), 753. https://doi.org/10.3390/d14090753