
The Impact of Spatial Delineation on the Assessment of Species Recovery Outcomes
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dataset
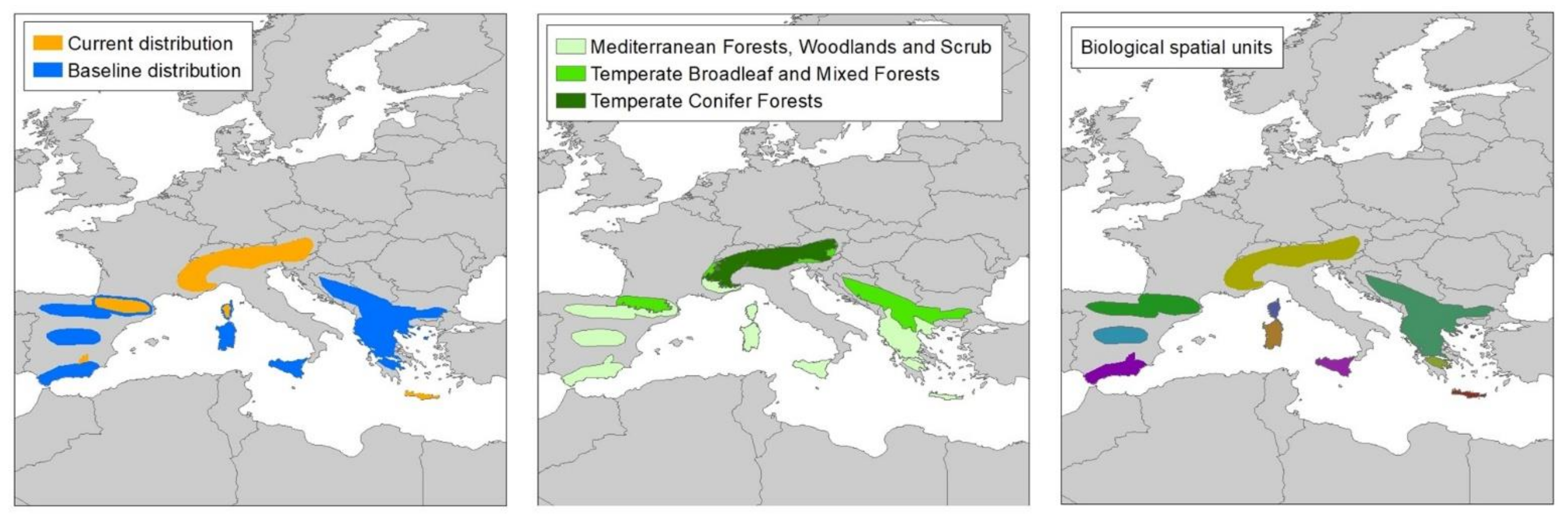
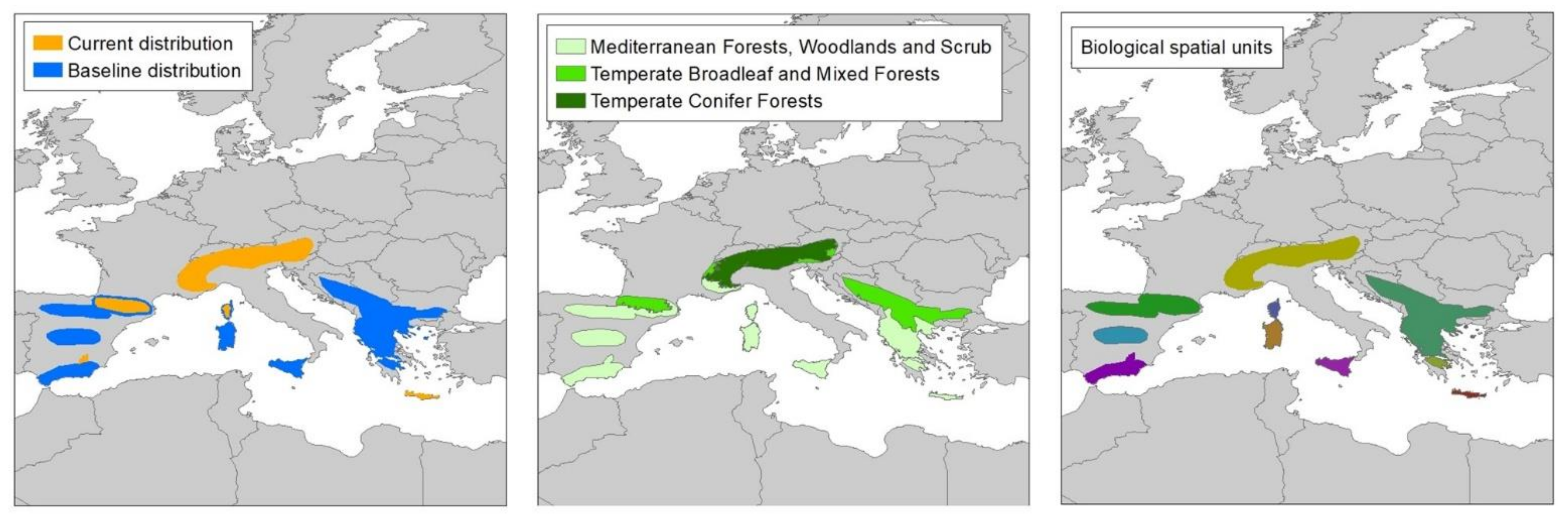
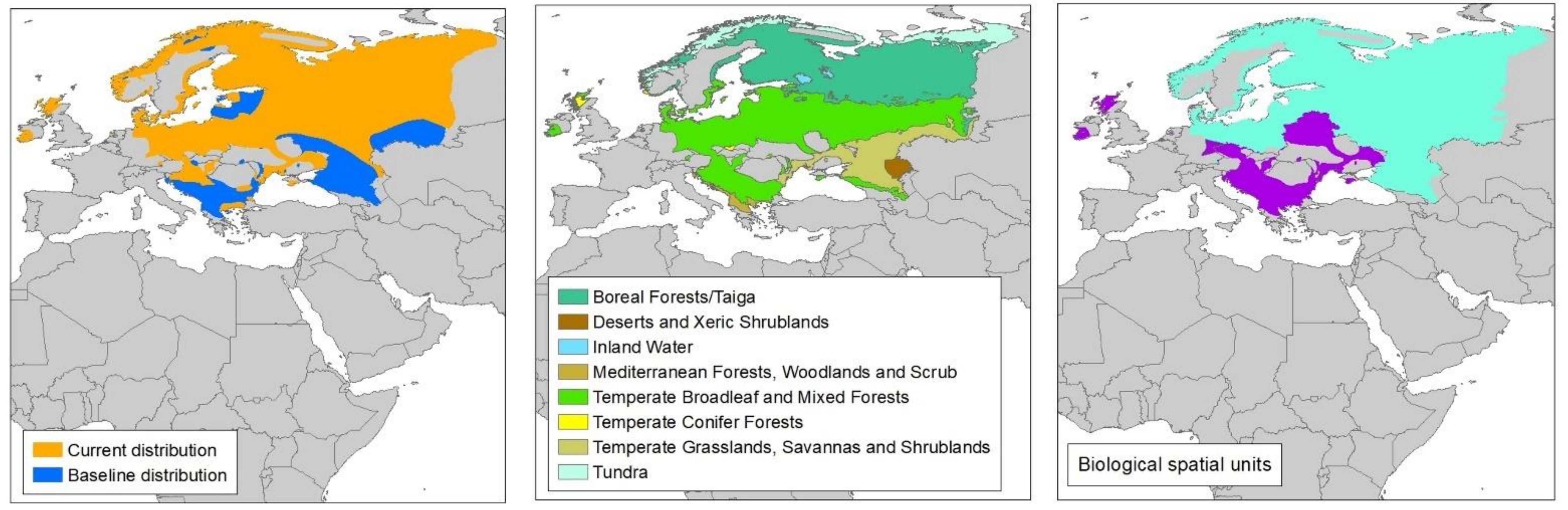
2.2. Species Distributions
2.3. Spatial Units
2.4. Population Estimates
2.5. Calculation of Green Score
2.6. Analysis
3. Results
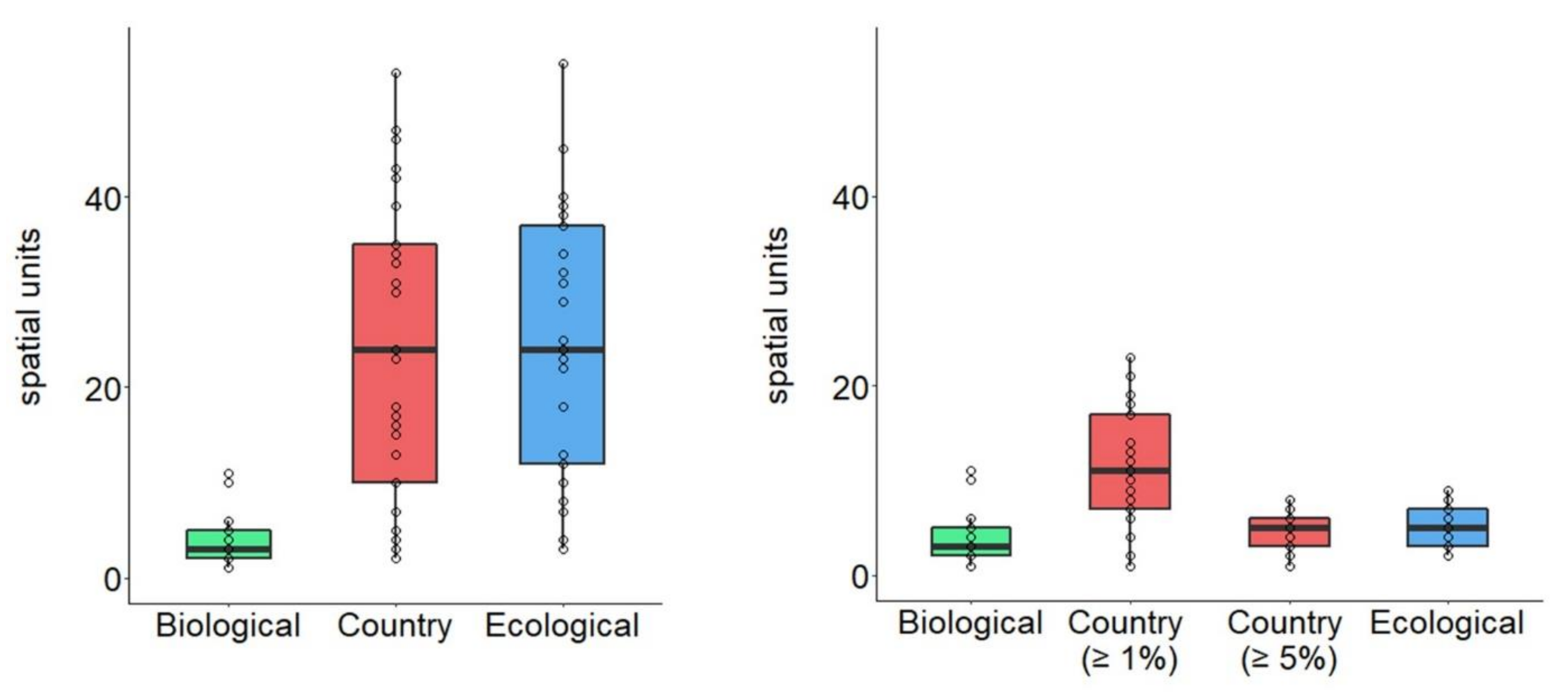
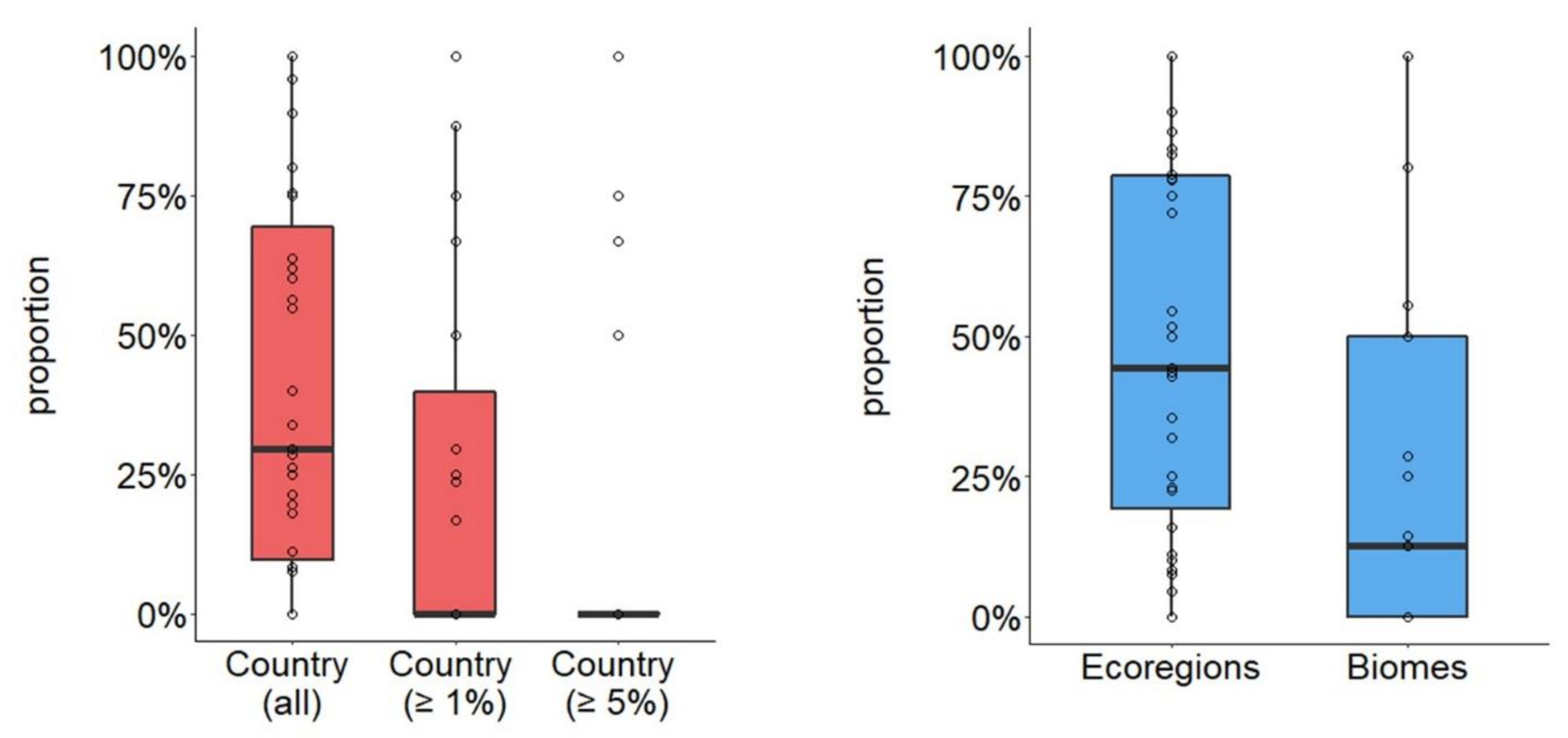
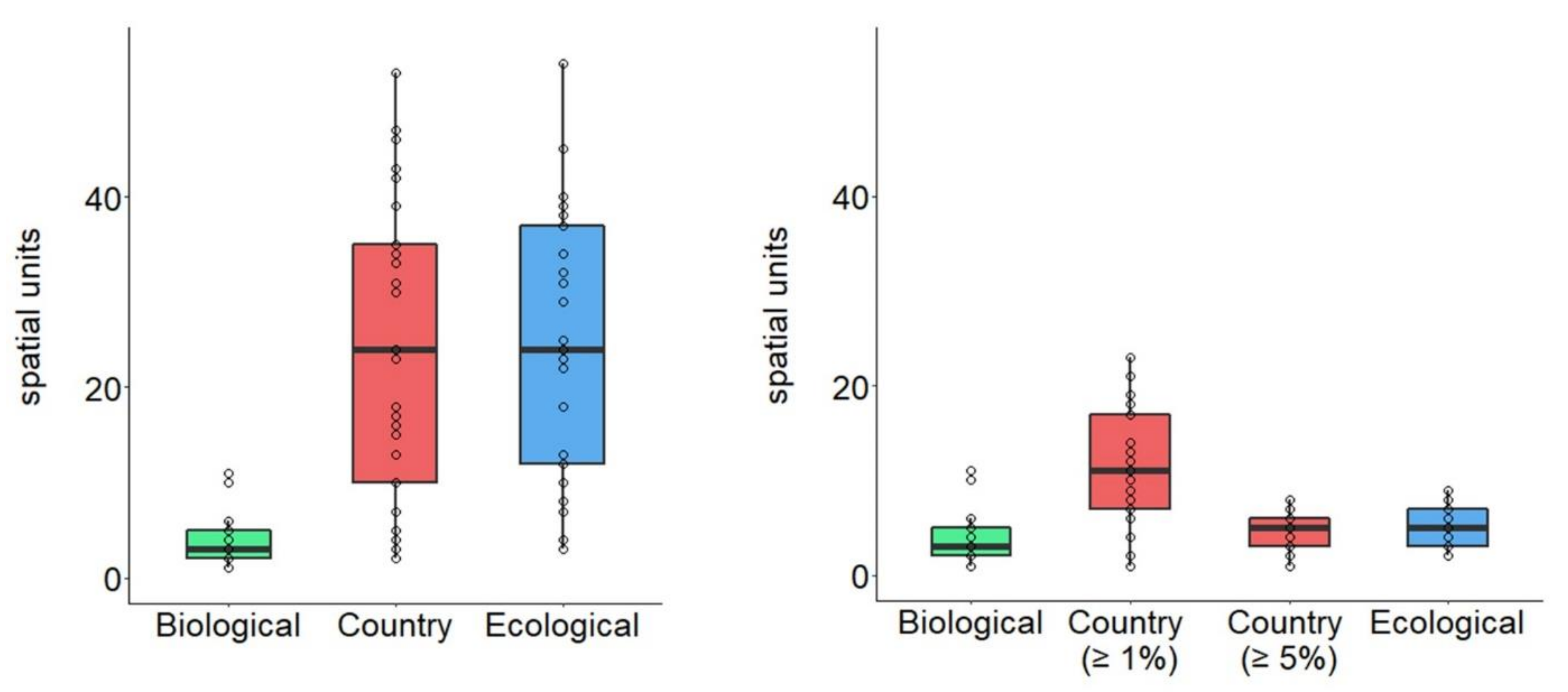
3.1. Average Changes in Number of Spatial Units Based on Delineation Method
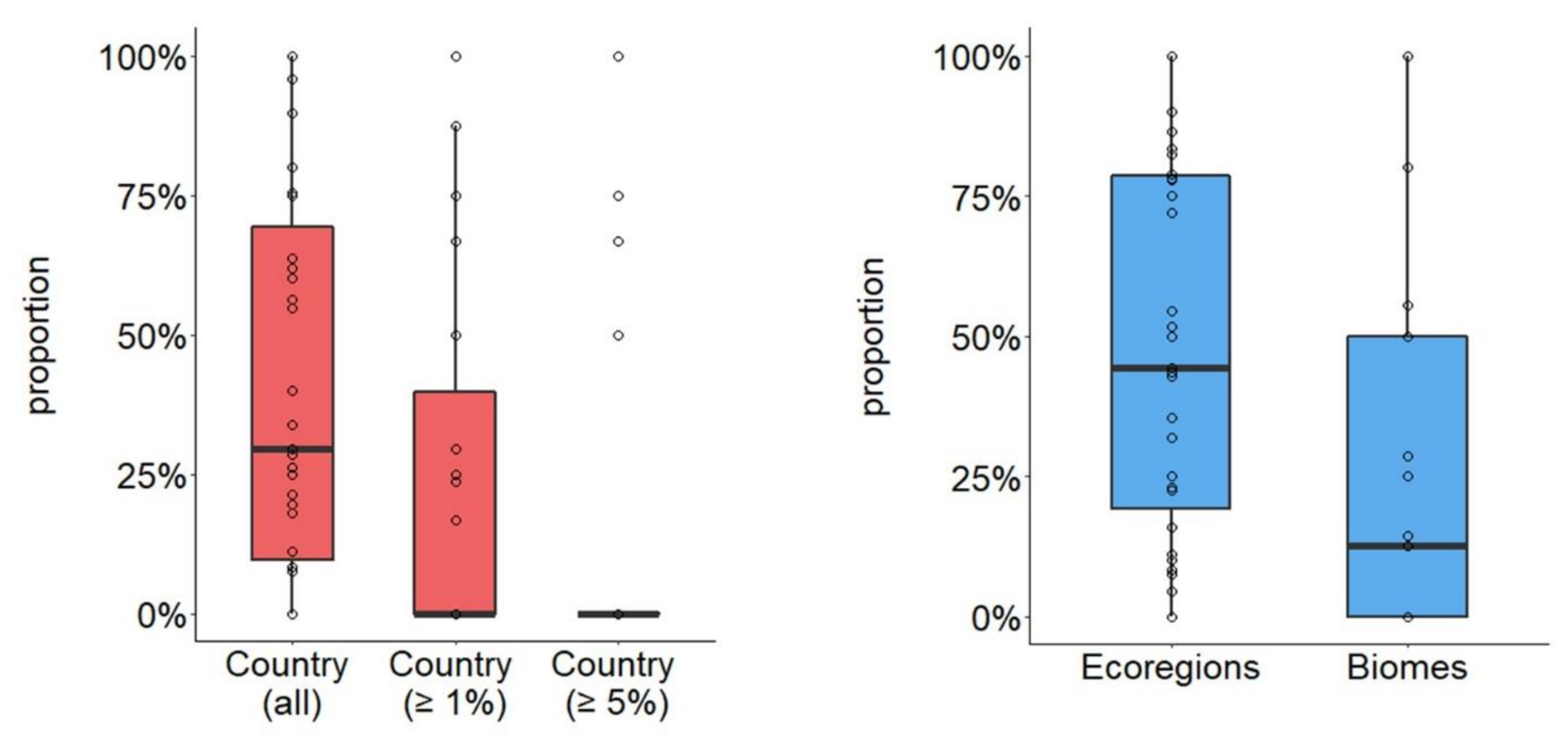
3.2. Generation of Spatial Units too Small to Hold a Minimum Viable Population
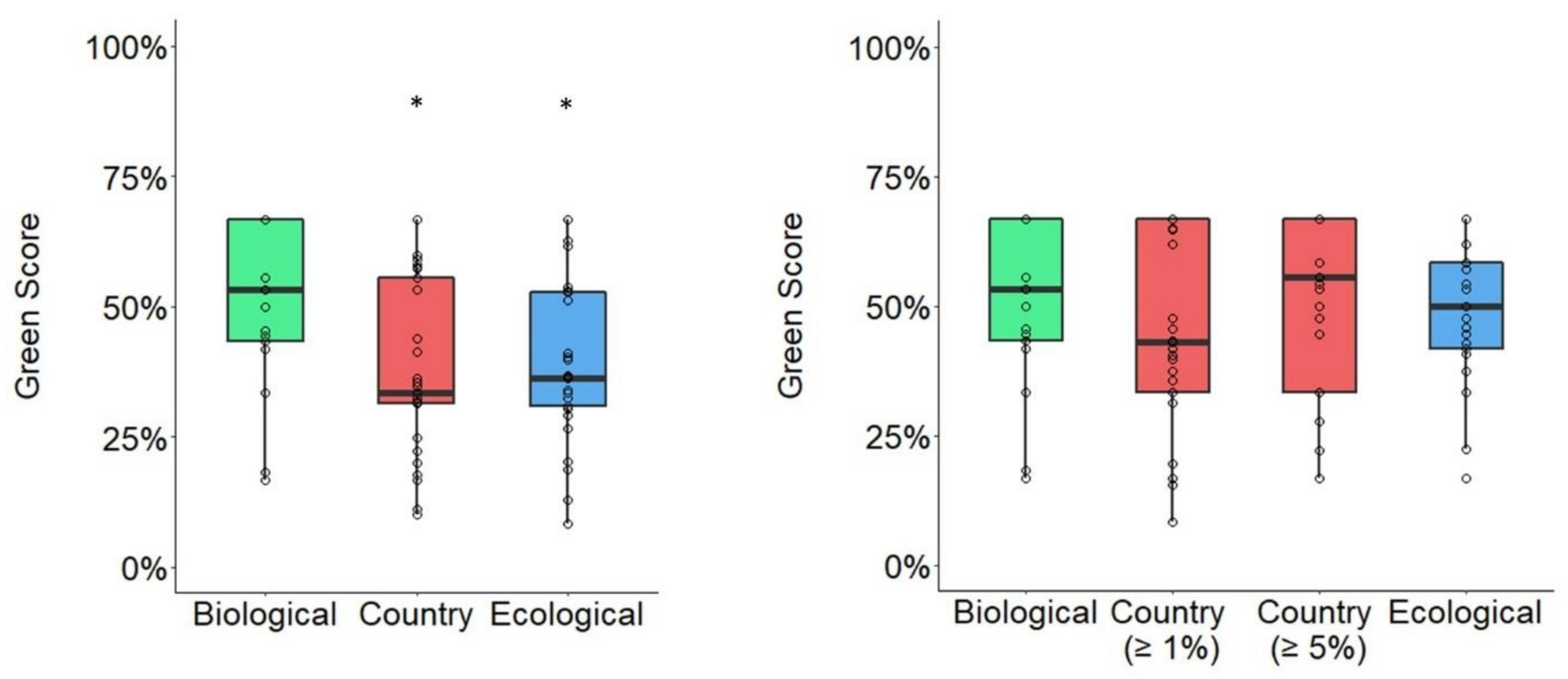
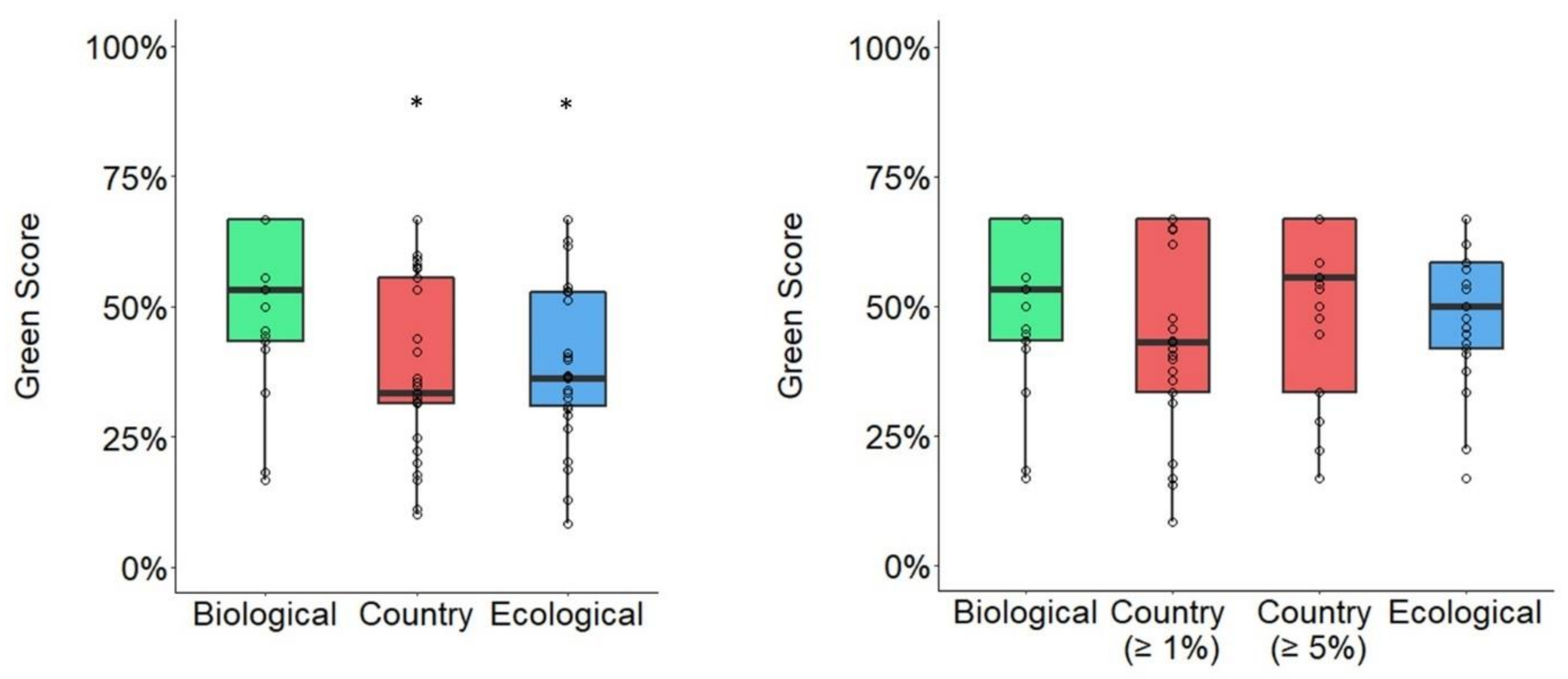
3.3. Average Changes in Green Score Based on Spatial Unit Delineation Method
3.4. Species-Level Changes in Green Score Based on Spatial Unit Delineation Method
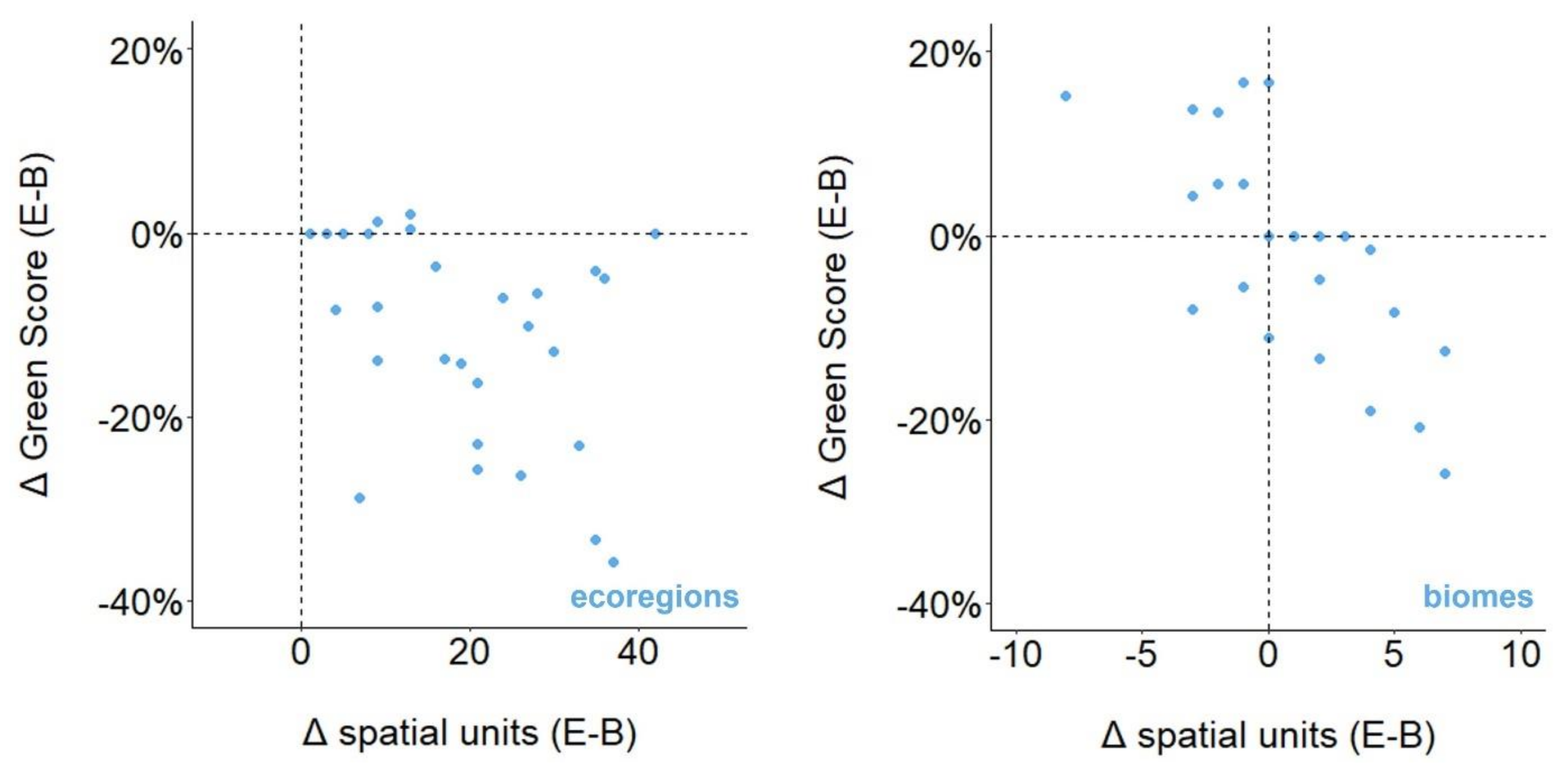
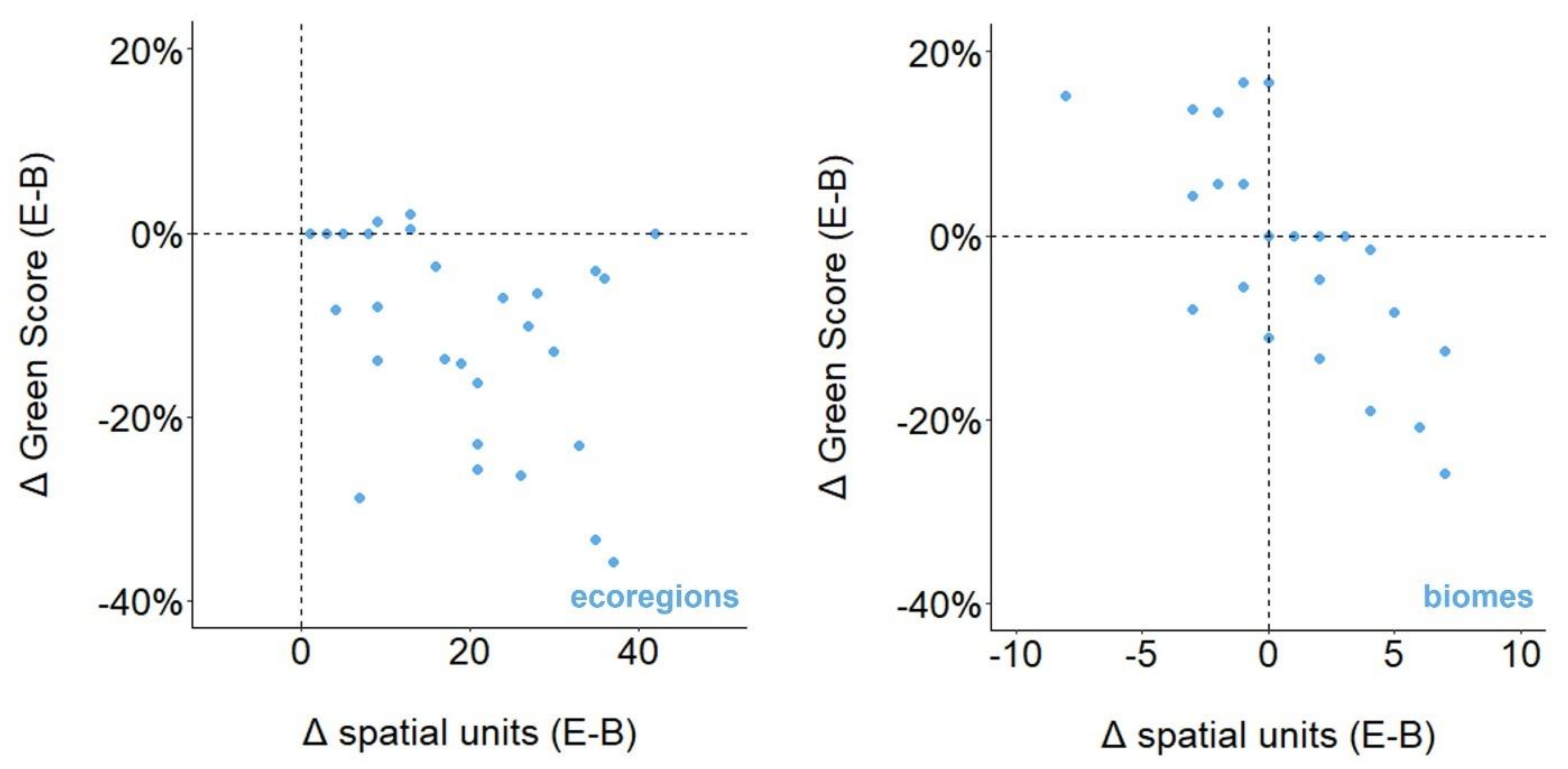
3.5. Ecological vs. Biological Case Studies
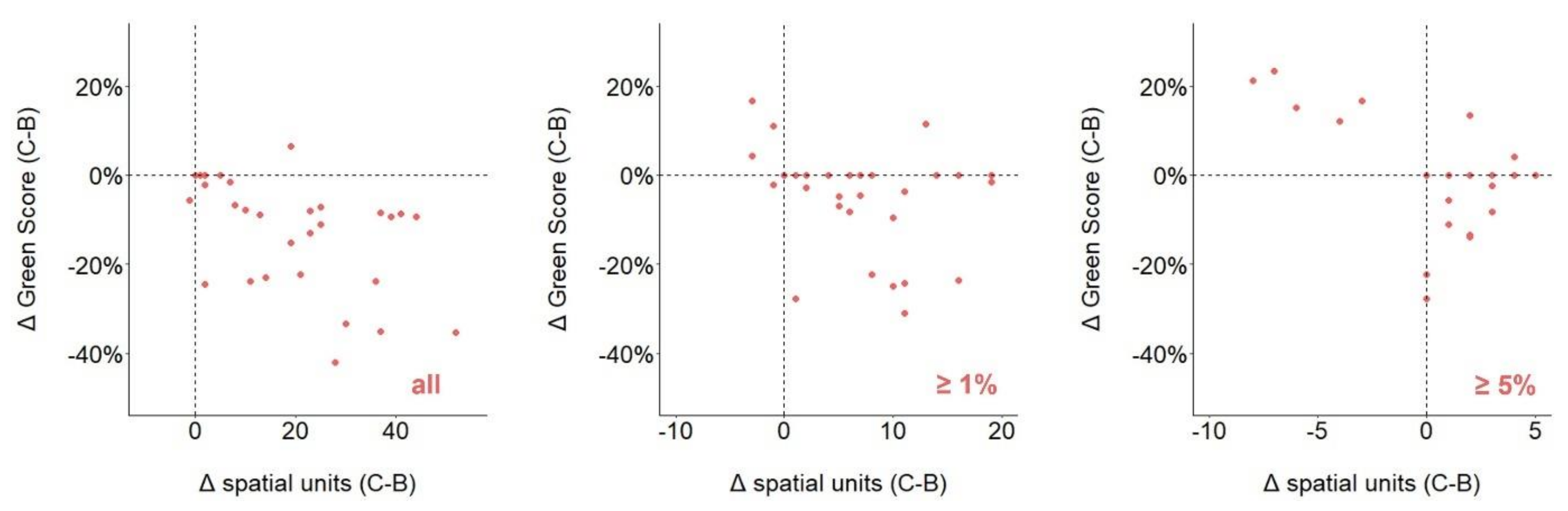
3.6. Country vs. Biological Case Studies
3.7. Cases Consistent across Spatial Unit Delineation Methods
4. Discussion
4.1. Findings and Recommendations
4.2. Scope of Results and Future Directions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IUCN. IUCN Green Status of Species: A Global Standard for Measuring Species Recovery and Assessing Conservation Impact. Version 2.0; IUCN: Gland, Switzerland, 2021. [Google Scholar] [CrossRef]
- Akcakaya, R.; Bennett, E.; Brooks, T.; Grace, M.K.; Heath, A.; Hedges, S.; Hilton-Taylor, C.; Hoffmann, M.; Keith, D.A.; Long, B.; et al. Quantifying Species Recovery and Conservation Success to develop an IUCN Green List of Species. Conserv. Biol. 2018, 32, 1128–1138. [Google Scholar] [CrossRef] [PubMed]
- Grace, M.; Akçakaya, H.R.; Bennett, E.L.K.; Brooks, T.M.; Heath, A.; Hedges, S.; Hilton-Taylor, C.; Hoffmann, M.; Hochkirch, A.; Jenkins, R.; et al. Testing a global standard for quantifying species recovery and assessing conservation impact. Conserv. Biol. 2021, 35, 1833–1849. [Google Scholar] [CrossRef] [PubMed]
- Grace, M.K.; Timmins, H.L.; Bennett, E.L.; Long, B.; Milner-Gulland, E.J.; Dudley, N. Engaging End-Users to Maximise Uptake and Effectiveness of a New Species Recovery Assessment. Conserv. Soc. 2021, 19, 150–160. [Google Scholar] [CrossRef]
- Sanderson, E.W. A full and authentic reckoning of species ranges for conservation. Conserv. Biol. 2019, 33, 1208–1210. [Google Scholar] [CrossRef]
- IUCN. IUCN Red List Categories and Criteria: Version 3.1, 2nd ed.; IUCN: Gland, Switzerland; Cambridge, UK, 2012; 32p. [Google Scholar]
- IUCN. Guidelines for Application of IUCN Red List Criteria at Regional and National Levels: Version 4.0. 2012. Available online: https://www.iucn.org/content/guidelines-application-iucn-red-list-criteria-regional-andnational-levels-version-40 (accessed on 27 July 2022).
- IUCN Species Conservation Success Task Force. Background and Guidelines for the IUCN Green Status of Species. Version 1.0. Prepared by the Species Conservation Success Task Force. 2020. Available online: https://www.iucnredlist.org/resources/green-status-assessment-materials (accessed on 27 July 2022).
- Moritz, C.D. Defining ‘Evolutionarily Significant Units’ for conservation. Trends Ecol. Evol. 1994, 9, 373–375. [Google Scholar] [CrossRef]
- Deinet, S.; Ieronymidou, C.; McRae, L.; Burfield, I.J.; Foppen, R.P.; Collen, B.; Böhm, M. Wildlife Comeback in Europe: The Recovery of Selected Mammal and Bird Species. Final Report to Rewilding Europe by ZSL, BirdLife International and the European Bird Census Council; ZSL: London, UK, 2013. [Google Scholar]
- Annoni, A.; Luzet, C.; Gubler, E.; Ihde, J. Map Projections for Europe; Report EUR 20120 EN; JRC: Ispra, Italy, 2001; 131p.
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- South, A. rworldmap: A New R package for Mapping Global Data. R J. 2011, 3, 35–43. [Google Scholar] [CrossRef]
- Mace, G.M.; Collar, N.J.; Gaston, K.J.; Hilton-Taylor, C.; Akcakaya, H.R.; Leader-Williams, N.; Milner-Gulland, E.J.; Stuart, S.N. Quantification of extinction risk: IUCN’s system for classifying threatened species. Conserv. Biol. 2008, 22, 1424–1442. [Google Scholar] [CrossRef]
- Grace, M.K.; Akçakaya, H.R. Green Status of Species Recovery State Calculator. 2020. Available online: https://oxford.onlinesurveys.ac.uk/species-recovery-status-calculator (accessed on 27 July 2022).
- Cribari-Neto, F.; Zeileis, A. Beta Regression in R. J. Stat. Softw. 2010, 34, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Douma, J.C.; Weedon, J.T. Analysing continuous proportions in ecology and evolution: A practical introduction to beta and Dirichlet regression. Methods Ecol. Evol. 2019, 10, 1412–1430. [Google Scholar] [CrossRef]
- Huber, D. (Large Carnivore Initiative for Europe/Bear Specialist Group). Ursus arctos. The IUCN Red List of Threatened Species 2007, e.T41688A10514791. Available online: https://www.iucnredlist.org/species/41688/10514791 (accessed on 28 July 2022).
- Halley, D.; Rosell, F.; Saveljev, A. Population and distribution of Eurasian beaver (Castor fiber). Balt. For. 2012, 18, 168–175. [Google Scholar]
- Tokarska, M.; Pertoldi, C.; Kowalczyk, R.; Perzanowski, K. Genetic status of the European bison Bison bonasus after extinction in the wild and subsequent recovery. Mammal Rev. 2011, 41, 151–162. [Google Scholar] [CrossRef]
- Olech, W.; Bison Specialist Group. Bison bonasus. The IUCN Red List of Threatened Species 2007, e.T2814A9484514. Available online: https://www.iucnredlist.org/species/2814/9484514 (accessed on 16 August 2022).
- Ranc, N.; Krofel, M.; Ćirović, D. Canis aureus (errata version published in 2019). The IUCN Red List of Threatened Species 2018, e.T118264161A144166860. Available online: https://www.iucnredlist.org/species/118264161/163507876 (accessed on 16 August 2022).
- Rodríguez, A.; Calzada, J. Lynx pardinus (Errata Version Published in 2020). The IUCN Red List of Threatened Species 2015, e.T12520A174111773. Available online: https://www.iucnredlist.org/species/12520/174111773 (accessed on 26 January 2021).
- Simón, M.A.; Gil-Sánchez, J.M.; Ruiz, G.; Garrote, G.; Mccain, E.B.; Fernandez, L. Reverse of the decline of the endangered Iberian lynx. Conserv. Biol. 2012, 26, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Lovari, S.; Lorenzini, R.; Masseti, M.; Pereladova, O.; Carden, R.F.; Brook, S.M.; Mattioli, S. Cervus elaphus (Errata Version Published in 2019). The IUCN Red List of Threatened Species 2018, e.T55997072A142404453. Available online: https://www.iucnredlist.org/species/55997072/142404453 (accessed on 10 February 2021).
- Meiri, M.; Kosintsev, P.; Conroy, K.; Meiri, S.; Barnes, I.; Lister, A. Subspecies dynamics in space and time: A study of the red deer complex using ancient and modern DNA and morphology. J. Biogeogr. 2018, 45, 367–380. [Google Scholar] [CrossRef]
- Randi, E.; Alves, P.C.; Carranza, J.; Milošević-Zlatanović, S.; Sfougaris, A.; Mucci, N. Phylogeography of roe deer (Capreolus capreolus) populations: The effects of historical genetic subdivisions and recent nonequilibrium dynamics. Molec. Ecol. 2004, 13, 3071–3083. [Google Scholar] [CrossRef]
- Herrero, J.; Conroy, J.; Maran, T.; Giannatos, G.; Stubbe, M.; Aulagnier, S. Capreolus capreolus. The IUCN Red List of Threatened Species 2007, e.T42395A10693900. Available online: https://www.iucnredlist.org/species/42395/10693900 (accessed on 10 February 2021).
- Meijaard, E.; d’Huart, J.P.; Oliver, W.L.R. Family Suidae (Pigs). In Handbook of the Mammals of the World, Volume 2: Hoofed Mammals; Wilson, D.E., Wilson, D.E., Mittermeier, R.A., Eds.; Lynx Ediciones: Barcelona, Spain, 2011; pp. 248–291. [Google Scholar]
- Boitani, L. Canis lupus (Errata Version Published in 2019). The IUCN Red List of Threatened Species 2018, e.T3746A144226239. Available online: https://www.iucnredlist.org/species/3746/144226239 (accessed on 12 February 2021).
- White, C.M.; Sonsthagen, S.A.; Sage, G.K.; Anderson, C.; Talbot, S.L. Genetic relationships among some subspecies of the Peregrine Falcon (Falco peregrinus L.), inferred from mitochondrial DNA control-region sequences. Auk 2013, 130, 78–87. [Google Scholar] [CrossRef]
- Knott, J.P.; Newbery, P.; Barov, B. Action Plan for the Red Kite Milvus Milvus in the European Union. International Red Kite Symposium 2009, MontbéLiard, Franche-Comte. Available online: https://ec.europa.eu/environment/nature/conservation/wildbirds/action_plans/docs/milvus_milvus.pdf (accessed on 12 February 2021).
- Kojola, I.; Danilov, P.I.; Laitala, H.M.; Belkin, V.; Yakimov, A. Brown bear population structure in core and periphery: Analysis of hunting statistics from Russian Karelia and Finland. Ursus 2003, 14, 17–20. [Google Scholar]
- Campbell, R.D. Demography and Life History of the Eurasian Beaver Castor Fiber. Ph.D. Thesis, 2010. Available online: https://ora.ox.ac.uk/objects/uuid:9cb53e73-edd0-47bd-a132-380e359dd5d1/download_file?file_format=pdf&safe_filename=RDC_Thesis.pdf&type_of_work=Thesis (accessed on 15 February 2021).
- Mysterud, A.; Bartoń, K.A.; Jędrzejewska, B.; Krasiński, Z.A.; Niedziałkowska, M.; Kamler, J.F. Population ecology and conservation of endangered megafauna: The case of European bison in Białowieża Primeval Forest, Poland. Anim. Conserv. 2007, 10, 77–87. [Google Scholar] [CrossRef]
- Pañella, P.; Herrero, J.; Canut, J.; Garcìa-Serrano, A. Long-term monitoring of Pyrenean chamois in a protected area reveals a fluctuating population. Hystrix 2011, 21, 183–188. [Google Scholar]
- Gashe, T.; Yihune, M. Population status, foraging ecology and activity pattern of golden jackal (Canis aureus) in Guangua Ellala Forest, Awi Zone, north west Ethiopia. PLoS ONE 2020, 15, e0233556. [Google Scholar] [CrossRef]
- Jedrzejewski, W.; Jedrzejewska, B.; Okarma, H.; Schmidt, K.; Bunevich, A.N.; Milkowski, L. Population dynamics (1869–1994), demography, and home ranges of the lynx in Bialowieza Primeval Forest (Poland and Belarus). Ecography 1996, 19, 122–138. [Google Scholar] [CrossRef]
- Escos, J.; Alados, C.L. Influence of weather and population characteristics of free-ranging Spanish ibex in the Sierra de Cazorla y Segura and in the eastern Sierra Nevada. Mammalia 1991, 55, 67–78. [Google Scholar] [CrossRef]
- Larkin, J.L.; Maehr, D.S.; Cox, J.J.; Wichrowski, M.W.; Crank, R.D. Factors affecting reproduction and population growth in a restored elk Cervus elaphus nelsoni population. Wildl. Biol. 2002, 8, 49–54. [Google Scholar] [CrossRef]
- Kjellander, P. Density Dependence in Roe Deer Population Dynamics. Ph.D. Thesis; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2000. Available online: https://core.ac.uk/download/pdf/335350641.pdf (accessed on 31 March 2021).
- Fernández-Llario, P.; Carranza, J.; De Trucios, S.H. Social organization of the wild boar (Sus scrofa) in Doñana National Park. Miscellània Zool. 1996, 19, 9–18. [Google Scholar]
- Lovari, S.; Sforzi, A.; Scala, C.; Fico, R. A wolf in the hand is worth two in the bush: A response to Ciucci et al.(2007). J. Zool. 2007, 273, 128–130. [Google Scholar] [CrossRef]
- Landa, A.; Skogland, T. The relationship between population density and body size of wolverines Gulo gulo in Scandinavia. Wildl. Biol. 1995, 1, 165–175. [Google Scholar] [CrossRef]
- Owen, M. Dynamics and age structure of an increasing goose population: The Svalbard barnacle goose Branta leucopsis. Nor. Polarinst. Skr. 1984, 181, 37–47. [Google Scholar]
- Cranswick, P.A.; Colhoun, K.; Einarsson, O.; McElwaine, J.G.; Gardarsson, A.; Pollitt, M.S.; Rees, E.C. The status and distribution of the Icelandic Whooper Swan population: Results of the international Whooper Swan census 2000. Waterbirds 2002, 25, 37–48. [Google Scholar]
- Shao, M.; Chen, B.; Cui, P.; Dai, N.; Chen, H. Sex ratios and age structure of several waterfowl species wintering at Poyang Lake, China. Pak. J. Zool. 2016, 48, 839–844. [Google Scholar]
- Christensen, T.K.; Fox, A.D. Changes in age and sex ratios amongst samples of hunter-shot wings from common duck species in Denmark 1982–2010. Eur. J. Wildl. Res. 2014, 60, 303–312. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Species | Biological SUs | Green Score | ΔSUEco * | ΔGSEco * | ΔSUCountry * | ΔGSCountry * | Baseline Distribution |
|---|---|---|---|---|---|---|---|---|
| Bird | Anser brachyrhynchus | 2 | 67% | 1 | 0% | 0 | 0% | D |
| Bird | Aquila adalberti | 1 | 33% | 1 | 0% | 0 | 0% | C |
| Bird | Ciconia ciconia | 2 | 67% | 3 | 0% | 5 | 0% | C |
| Bird | Oxyura leucocephala | 2 | 17% | 2 | 0% | 2 | 0% | C |
| Mammal | Cervus elaphus | 4 | 67% | 3 | 0% | 4 | 0% | C |
| Mammal | Gulo gulo | 2 | 33% | 3 | 0% | 2 | 0% | C |
| Mammal | Sus scrofa | 5 | 67% | 2 | 0% | 1 | 0% | C |
| Bird | Aegypius monachus | 4 | 42% | 0 | 0% | 2 | −14% | S |
| Bird | Aquila heliaca | 3 | 44% | 4 | −2% | 0 | 0% | S |
| Bird | Branta leucopsis | 3 | 67% | 2 | −13% | 2 | 0% | D |
| Bird | Cygnus cygnus | 2 | 67% | 6 | −21% | 1 | 0% | S |
| Bird | Grus grus | 3 | 67% | 5 | −8% | 2 | 0% | C |
| Bird | Gyps fulvus | 4 | 50% | 0 | 0% | 3 | −2% | S |
| Mammal | Bison bonasus | 2 | 33% | 0 | 17% | 3 | 0% | D |
| Mammal | Capreolus capreolus | 3 | 67% | 5 | −8% | 3 | 0% | C |
| Mammal | Castor fiber | 5 | 67% | 2 | −5% | 2 | 0% | S |
| Bird | Falco naumanni | 3 | 67% | 4 | −19% | 2 | −13% | D |
| Bird | Falco peregrinus | 1 | 67% | 7 | −13% | 3 | −8% | S |
| Bird | Gypaetus barbatus | 10 | 18% | −7 | 15% | −6 | 15% | D |
| Bird | Haliaeetus albicilla | 2 | 67% | 7 | −26% | 1 | −11% | C |
| Bird | Milvus milvus | 6 | 56% | 0 | −11% | 0 | −28% | S |
| Mammal | Canis aureus | 4 | 50% | −1 | 17% | 4 | 4% | C |
| Mammal | Canis lupus | 10 | 43% | −3 | 4% | −4 | 12% | C |
| Mammal | Capra pyrenaica | 3 | 44% | −1 | 6% | −2 | 23% | D |
| Mammal | Lynx lynx | 11 | 45% | −3 | −8% | −8 | 21% | S |
| Mammal | Lynx pardinus | 4 | 17% | −2 | 6% | −3 | 17% | D |
| Mammal | Rupicapra pyrenaica | 3 | 56% | −1 | −6% | 1 | −6% | D |
| Mammal | Rupicapra rupicapra | 5 | 53% | −2 | 13% | 2 | 13% | D |
| Mammal | Ursus arctos | 10 | 43% | −3 | 14% | −7 | 23% | S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grace, M.K.; Akçakaya, H.R.; Bennett, E.L.; Boyle, M.J.W.; Hilton-Taylor, C.; Hoffmann, M.; Money, D.; Prohaska, A.; Young, R.; Young, R.; et al. The Impact of Spatial Delineation on the Assessment of Species Recovery Outcomes. Diversity 2022, 14, 742. https://doi.org/10.3390/d14090742
Grace MK, Akçakaya HR, Bennett EL, Boyle MJW, Hilton-Taylor C, Hoffmann M, Money D, Prohaska A, Young R, Young R, et al. The Impact of Spatial Delineation on the Assessment of Species Recovery Outcomes. Diversity. 2022; 14(9):742. https://doi.org/10.3390/d14090742
Chicago/Turabian StyleGrace, Molly K., H. Resit Akçakaya, Elizabeth L. Bennett, Michael J. W. Boyle, Craig Hilton-Taylor, Michael Hoffmann, Daniel Money, Ana Prohaska, Rebecca Young, Richard Young, and et al. 2022. "The Impact of Spatial Delineation on the Assessment of Species Recovery Outcomes" Diversity 14, no. 9: 742. https://doi.org/10.3390/d14090742
APA StyleGrace, M. K., Akçakaya, H. R., Bennett, E. L., Boyle, M. J. W., Hilton-Taylor, C., Hoffmann, M., Money, D., Prohaska, A., Young, R., Young, R., & Long, B. (2022). The Impact of Spatial Delineation on the Assessment of Species Recovery Outcomes. Diversity, 14(9), 742. https://doi.org/10.3390/d14090742