
Biodiversity and Variations of Arbuscular Mycorrhizal Fungi Associated with Roots along Elevations in Mt. Taibai of China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Study Region
2.2. Collection of Samples
2.3. Bioinformatics Analysis of Sequence Data
2.4. Measurement of AM Colonization and Parameters of Soil and Plant Roots
2.5. Calculation of AM Colonization, Relative Abundance, and Occurrence Frequency
2.6. Statistics and Analysis of Data
3. Results
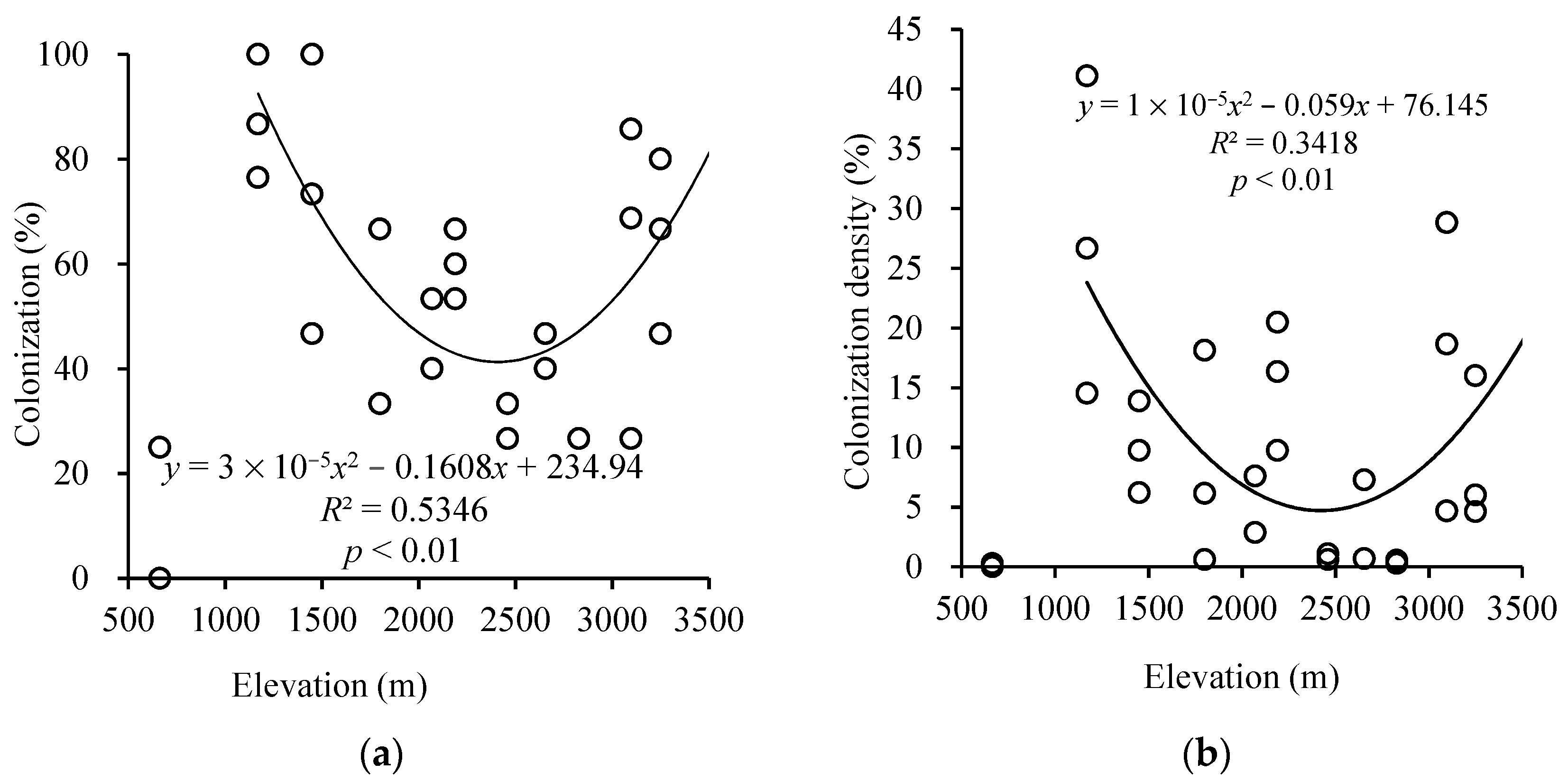
3.1. Arbuscular Mycorrhizal Colonization in Plant Roots at Different AltitudesSubsection
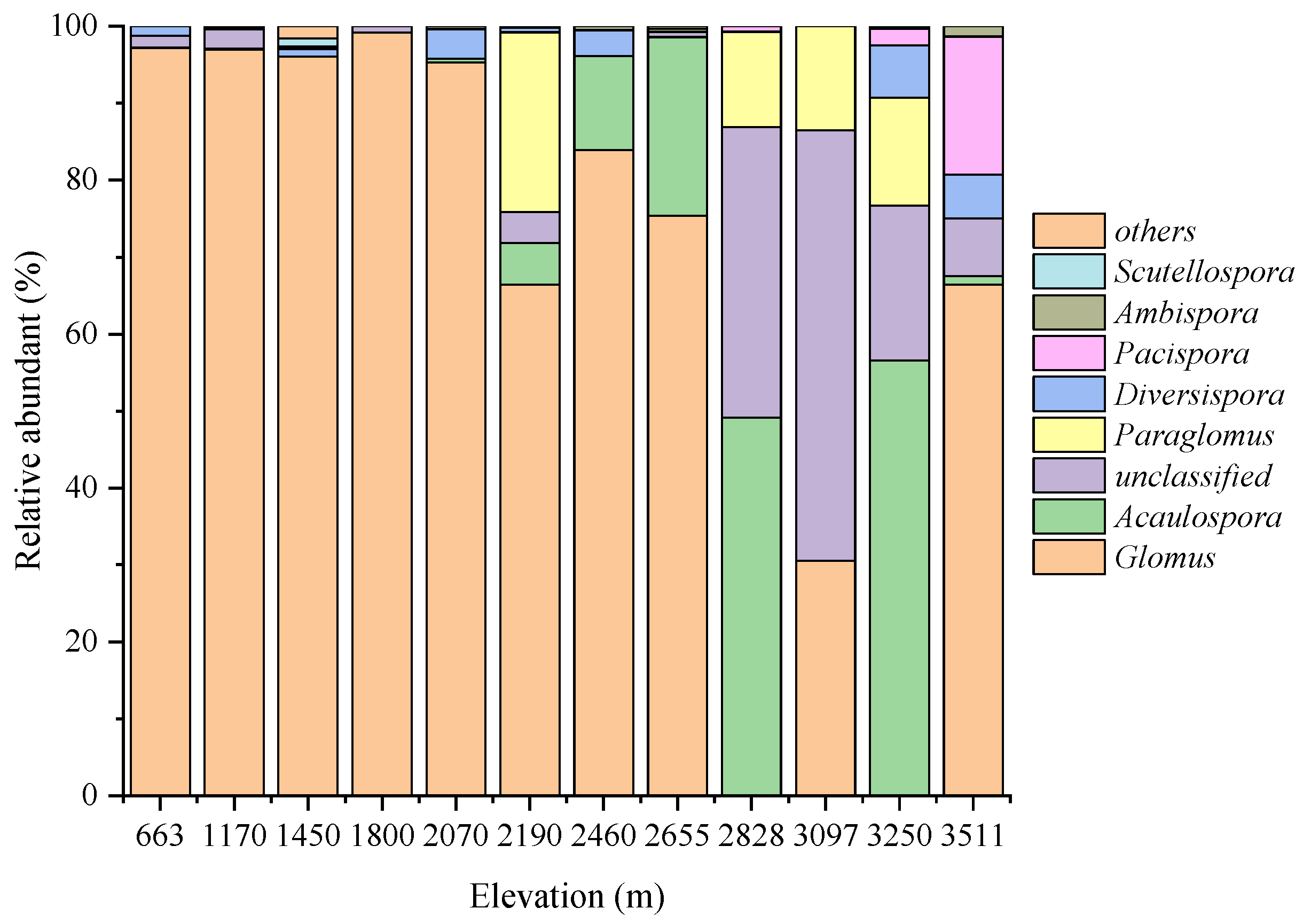
3.2. AMF Community Composition and Distribution at Different Altitudes
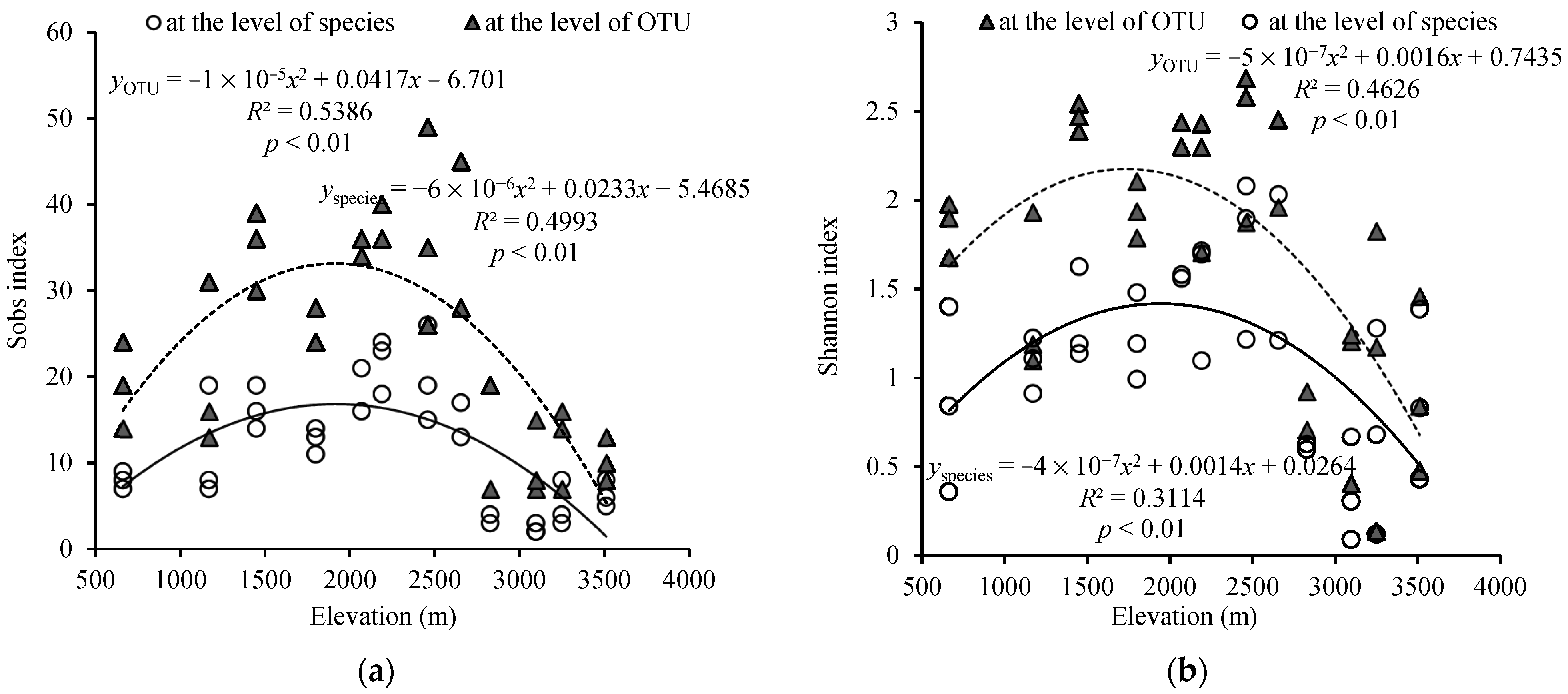
3.3. Diversity of AMF in Mt. Taibai
3.4. The Relative Abundance and Occurrence Frequency of AMF Genus in Mt. Taibai
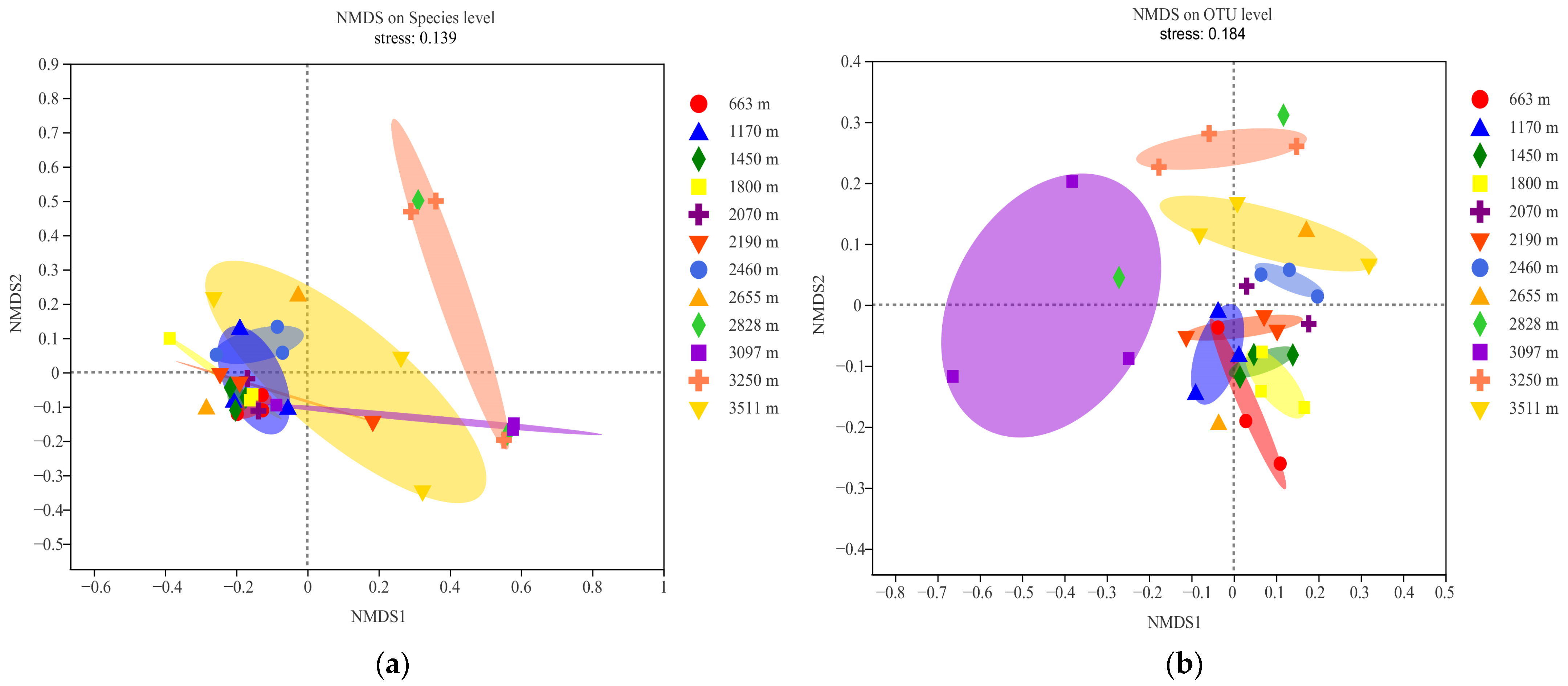
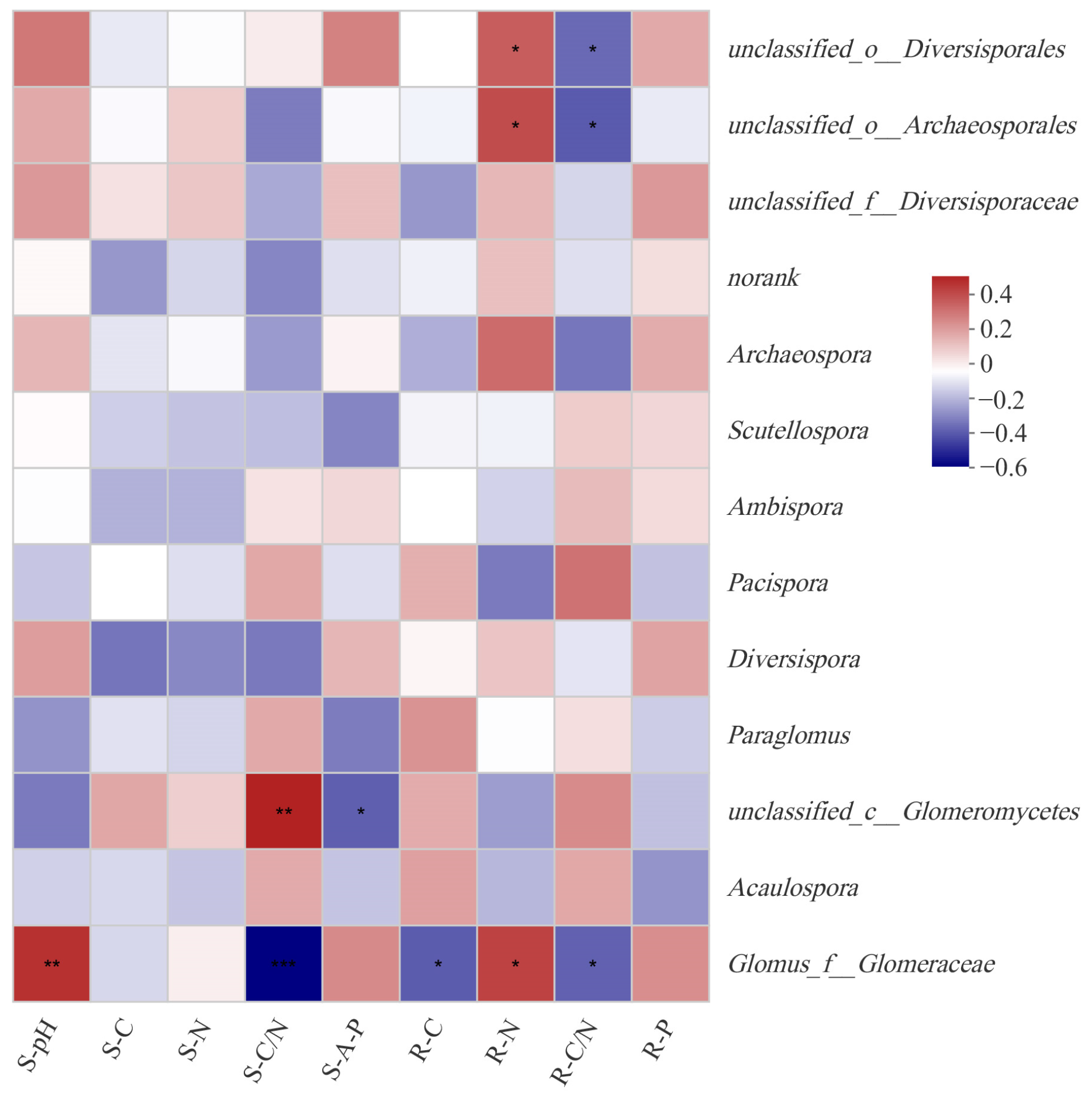
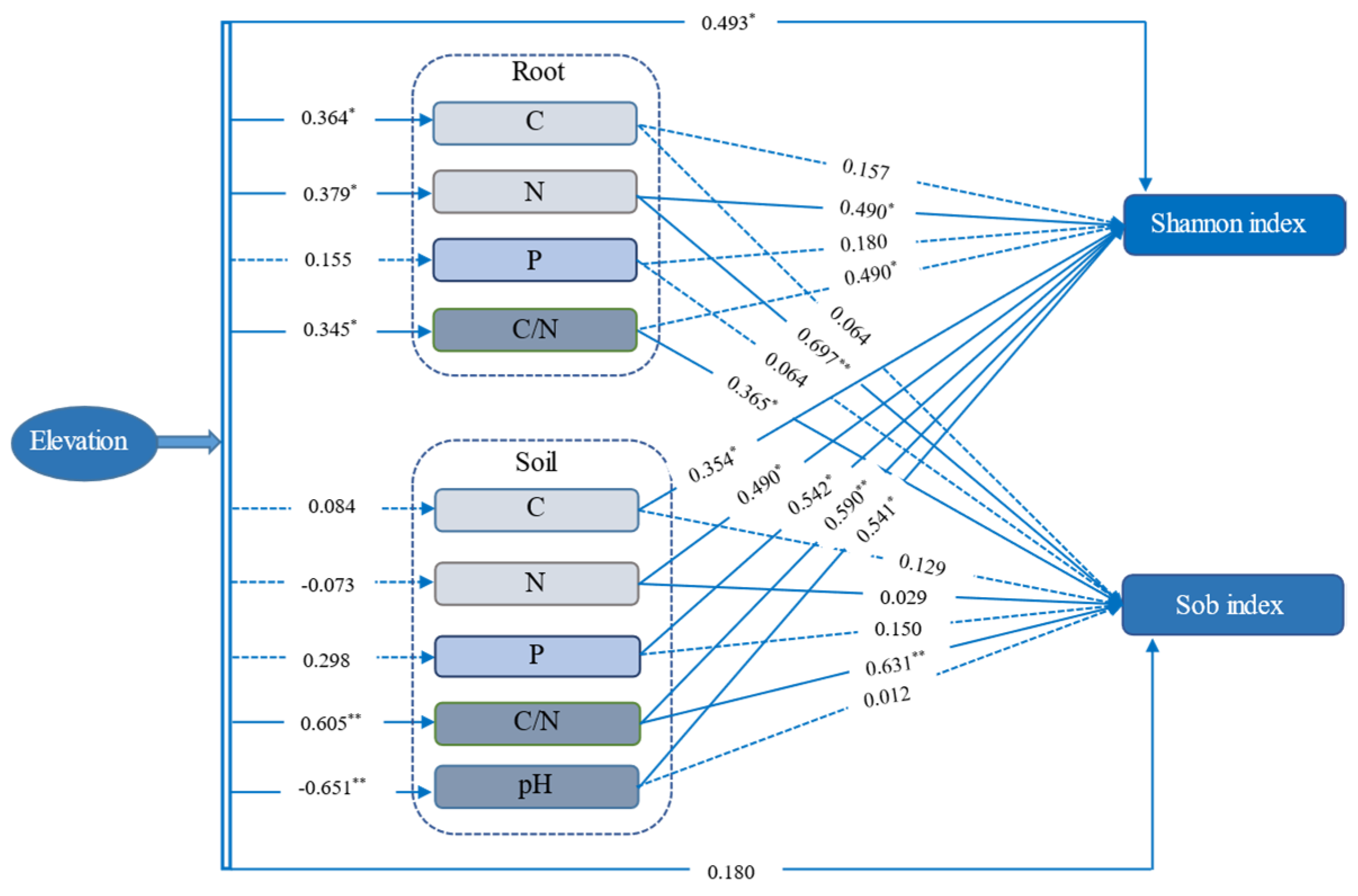
3.5. The Drive Factors of AMF Community and Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deng, W.; Wang, J.L.; Scott, M.B.; Fang, Y.H.; Liu, S.R.; Yang, X.Y.; Xiao, W. Sampling methods affect nematode-trapping fungi biodiversity patterns across an elevational gradient. BMC Microbiol. 2020, 20, 15. [Google Scholar] [CrossRef] [Green Version]
- Wagg, C.; Husband, B.C.; Green, D.S.; Massicotte, H.B.; Peterson, R.L. Soil microbial communities from an elevational cline differ in their effect on conifer seedling growth. Plant Soil 2011, 340, 491–504. [Google Scholar] [CrossRef]
- Peters, M.K.; Hemp, A.; Appelhans, T.; Behler, C.; Classen, A.; Detsch, F.; Ensslin, A.; Ferger, S.W.; Frederiksen, S.B.; Gebert, F.; et al. Predictors of elevational biodiversity gradients change from single taxa to the multi-taxa community level. Nat. Commun. 2016, 7, 13736. [Google Scholar] [CrossRef] [Green Version]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Microbes on mountain sides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitter, A.H. Darkness visible: Reflections on underground ecology. J. Ecol. 2005, 93, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Jing, X.; Sanders, N.J.; Shi, Y.; Chu, H.Y.; Classen, A.T.; Zhao, K.; Chen, L.T.; Shi, Y.; Jiang, Y.X.; He, J.S. The links between ecosystem multifunctionality and above-and belowground biodiversity are mediated by climate. Nat. Commun. 2015, 6, 8159. [Google Scholar] [CrossRef] [PubMed]
- Bauer, J.T.; Koziol, L.; Bever, J.D. Local adaptation of mycorrhizae communities changes plant community composition and increases aboveground productivity. Oecologia 2020, 192, 735–744. [Google Scholar] [CrossRef]
- Yang, W.; Zheng, Y.; Gao, C.; Duan, J.C.; Wang, S.P.; Guo, L.D. Arbuscular mycorrhizal fungal community composition affected by original elevation rather than translocation along an altitudinal gradient on the Qinghai-Tibet Plateau. Sci. Rep. 2016, 6, 36606. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.E.; Read, D. Mycorrhizal Symbiosis. Q. Rev. Biol. 2008, 3, 273–281. [Google Scholar] [CrossRef]
- Davison, J.; Moora, M.; Opik, M.; Adholeya, A.; Ainsaar, L.; Ba, A.; Burla, S.; Diedhiou, A.G.; Hiiesalu, I.; Jairus, T.; et al. Global assessment of arbuscular mycorrhizal fungus diversity reveals very low endemism. Science 2015, 349, 6251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.Y.; Zhang, J.C.; Lu, S.C.; Li, Y.; Wang, F.Y. Arbuscular mycorrhizal fungi improve the performance of sweet sorghum grown in a mo-contaminated soil. J. Fungi 2020, 6, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiang, W.; He, X.; Wang, J.; Zhao, L. Temporal and spatial variation of arbuscular mycorrhizal fungi under the canopy of Hedysarum scoparium in the northern desert, China. Appl. Soil Ecol. 2019, 136, 139–147. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Klironomos, J.N.; Ursic, M.; Moutoglis, P.; Streitwolf-engel, R.; Boller, T.; Wiemken, A.; Sanders, L.R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Shi, Z.Y.; Wang, F.Y.; Zhang, K.; Chen, Y.L. Diversity and distribution of arbuscular mycorrhizal fungi along altitudinal gradients in Mount Taibai of the Qinling Mountains. Can. J. Microbiol. 2014, 60, 811–818. [Google Scholar] [CrossRef]
- Li, X.L.; Xu, M.; Christie, P.; Li, X.L.; Zhang, J.L. Large elevation and small host plant differences in the arbuscular mycorrhizal communities of montane and alpine grasslands on the Tibetan Plateau. Mycorrhiza 2018, 28, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Shi, Z.Y.; Mickan, B.S.; Zhang, M.G.; Cao, L.B. Alterations to arbuscular mycorrhizal fungal community composition is driven by warming at specific elevations. Peer J. 2021, 9, e11792. [Google Scholar] [CrossRef]
- Gough, E.C.; Owen, K.J.; Zwart, R.S.; Thompson, J.P. A systematic review of the effects of arbuscular mycorrhizal fungi on root-lesion nematodes, Pratylenchus spp. Front. Plant Sci. 2020, 11, 923. [Google Scholar] [CrossRef]
- Zhang, M.G.; Shi, Z.Y.; Yang, M.; Lu, S.C.; Wang, X.G. Molecular diversity and distribution of arbuscular mycorrhizal fungi at different elevations in Mt. Taibai of Qinling Mountain. Front. Microbiol. 2021, 12, 609386. [Google Scholar] [CrossRef] [PubMed]
- Shuai, L.Y.; Ren, C.L.; Yan, W.B.; Song, Y.L.; Zeng, Z.G. Different elevational patterns of rodent species richness between the southern and northern slopes of a mountain. Sci. Rep. 2017, 7, 8743. [Google Scholar] [CrossRef]
- Guo, Y.X.; Ren, C.J.; Yi, J.J.; Doughty, R.; Zhao, F.Z. Contrasting responses of rhizosphere bacteria, fungi and arbuscular mycorrhizal fungi along an elevational gradient in a temperate montane forest of China. Front. Microbiol. 2020, 11, 2042. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.Z.; Feng, X.X.; Guo, Y.X.; Ren, C.J.; Wang, J.; Doughty, R. Elevation gradients affect the differences of arbuscular mycorrhizal fungi diversity between root and rhizosphere soil. Agric. For. Meteorol. 2020, 284, 107894. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agrochemical Analysis; China Agricultural Press: Beijing, China, 2000; pp. 176–185. [Google Scholar]
- Koske, R.E.; Tessier, B. A convenient, permanent slide mounting medium. Mycol. Soc. Am. Newsl. 1983, 34, 59. [Google Scholar]
- Bhople, P.; Samad, A.; Sisic, A.; Antonielli, L.; Sessitsch, A.; Keiblinger, K.; Djukic, I.; Zehetner, F.; Zechmeister-Boltenstern, S.; Joergensen, R.G.; et al. Variations in fungal community structure along elevation gradients in contrasting Austrian Alpine ecosystems. Appl. Soil Ecol. 2022, 177, 104508. [Google Scholar] [CrossRef]
- Xu, D.P.; Kong, H.J.; Yang, E.J.; Wang, Y.; Li, X.R.; Sun, P.; Jiao, N.Z.; Lee, Y.J.; Jung, J.Y.; Cho, K.H. Spatial dynamics of active microeukaryotes along a latitudinal gradient: Diversity, assembly process, and co-occurrence relationships. Environ. Res. 2022, 212, 113234. [Google Scholar] [CrossRef] [PubMed]
- Cotton, T.E.A. Arbuscular mycorrhizal fungal communities and global change: An uncertain future. FEMS Microbiol. Ecol. 2018, 94, 14. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.W.; Liu, X.; Zhou, S.R. Nitrogen addition altered the plant-arbuscular mycorrhizal fungi network through reducing redundant interactions in an alpine meadow. Soil Biol. Biochem. 2022, 171, 108727. [Google Scholar] [CrossRef]
- Thangavel, P.; Anjum, N.A.; Muthukumar, T.; Sridevi, G.; Vasudhevan, P.; Maruthupandian, A. Arbuscular mycorrhizae: Natural modulators of plant-nutrient relation and growth in stressful environments. Arch. Microbiol. 2022, 204, 264. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Guo, M.; Wang, E.; Yin, C.Y.; Wang, Y.J.; He, H.L.; Zhao, C.Z. Effects of mycorrhiza and hyphae on the response of soil microbial community to warming in eastern Tibetan Plateau. Sci. Total Environ. 2022, 837, 155498. [Google Scholar] [CrossRef] [PubMed]
- Gai, J.P.; Tian, H.; Yang, F.Y.; Christie, P.; Li, X.L.; Klironomos, J.N. Arbuscular mycorrhizal fungal diversity along a Tibetan elevation gradient. Pedobiologia 2012, 55, 145–151. [Google Scholar] [CrossRef]
- Kotilinek, M.; Hiiesalu, I.; Kosnar, J.; Smilauerov, M.; Smilauer, P.; Altman, J.; Dvorsky, M.; Kopecky, M.; Dolezal, J. Fungal root symbionts of high-altitude vascular plants in the Himalayas. Sci. Rep. 2017, 7, 6562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.L.; Xu, M.; Li, X.L.; Christie, P.; Wagg, C.; Zhang, J.L. Linkages between changes in plant and mycorrhizal fungal community composition at high versus low elevation in alpine ecosystems. Environ. Microbiol. Rep. 2020, 12, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Liu, M. Research on the Arbuscular Mycorrhizal Fungal Diversity in the Rhizosphere of Clematis fruticosa and the Mechanism of Mycorrhizal Seedlings Response to Drought Stress. Ph.D. Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2017. [Google Scholar]
- Li, X.L.; Gai, J.P.; Cai, X.B.; Li, X.L.; Christie, P.; Zhang, F.S.; Zhang, J.L. Molecular diversity of arbuscular mycorrhizal fungi associated with two co-occurring perennial plant species on a Tibetan altitudinal gradient. Mycorrhiza 2014, 24, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, X.Z.; Zhang, Z.M.; Zhao, Y.; Yang, J.T.; Zhu, Y.W. Species diversity and drivers of arbuscular mycorrhizal fungal communities in a semi-arid mountain in China. Peer J. 2017, 5, e4155. [Google Scholar] [CrossRef] [Green Version]
- Camenzind, T.; Hempel, S.; Homeier, J.; Horn, S.; Velescu, A.; Wilcke, W.; Rillig, M.C. Nitrogen and phosphorus additions impact arbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest. Glob. Change Biol. 2014, 20, 3646–3659. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.Y.; Yin, K.J.; Wang, F.Y.; Mickan, B.S.; Wang, X.G.; Zhou, W.L.; Li, Y.J. Alterations of arbuscular mycorrhizal fungal diversity in soil with elevation in tropical forests of China. Diversity 2019, 11, 181. [Google Scholar] [CrossRef] [Green Version]
- Oehl, F.; Sykorova, Z.; Redecker, D.; Wiemken, A.; Sieverding, E. Acaulospora alpina, a new arbuscular mycorrhizal fungal species characteristic for high mountainous and alpine regions of the Swiss Alps. Mycologia 2006, 98, 286–294. [Google Scholar] [CrossRef]
- Haug, I.; Setaro, S.; Suarez, J.P. Species composition of arbuscular mycorrhizal communities changes with elevation in the Andes of south Ecuador. PLoS ONE 2019, 14, e0221091. [Google Scholar] [CrossRef] [Green Version]
- Egan, C.P.; Callaway, R.M.; Hart, M.M.; Pither, J.; Klironomos, J. Phylogenetic structure of arbuscular mycorrhizal fungal communities along an elevation gradient. Mycorrhiza 2017, 27, 273–282. [Google Scholar] [CrossRef]
- Hiiesalu, I.; PärtelM, D.J.; Gerhold, P.; Metsis, M.; Moora, M.; Öpik, M.; Vasar, M.; Zobel, M.; Wilson, S.D. Species richness of arbuscular mycorrhizal fungi: Associations with grassland plant richness and biomass. New Phytol. 2014, 203, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Bonfim, J.A.; Vascon, R.L.F.; Gumiere, T.; Mescolotti, D.D.L.C.; Oehl, F.; Cardoso, E.J.B.N. Diversity of arbuscular mycorrhizal fungi in a Brazilian atlantic forest toposequence. Microb. Ecol. 2016, 71, 164–177. [Google Scholar] [CrossRef]
- Moora, M.; Davison, J.; Opik, M.; Metsis, M.; Saks, U.; Jairus, T.; Vasar, M.; Zobel, M. Anthropogenic land use shapes the composition and phylogenetic structure of soil arbuscular mycorrhizal fungal communities. FEMS Microbiol. Ecol. 2014, 90, 609–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geml, J. Altitudinal gradients in mycorrhizal symbioses. In Biogeography of Mycorrhizal Symbiosis. Ecological Studies (Analysis and Synthesis); Tedersoo, L., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; Volume 230, pp. 107–123. [Google Scholar] [CrossRef]
- Tunnisa, R.; Ezawa, T. Nestedness in arbuscular mycorrhizal fungal communities in a volcanic ecosystem: Selection of disturbance-tolerant fungi along an elevation gradient. Microbes. Environ. 2019, 34, 327–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.M.; Chen, B.D.; Yin, R.B.; Xing, S.P.; Fu, W.; Wu, H.; Hao, Z.P.; Ma, Y.B.; Zhang, X. Long-term nickel contamination increased soil fungal diversity and altered fungal community structure and co-occurrence patterns in agricultural soils. J. Hazard. Mater. 2022, 436, 129113. [Google Scholar] [CrossRef]
- Ma, X.C.; Geng, Q.H.; Zhang, H.G.; Bian, C.Y.; Chen, H.Y.H.; Jiang, D.L.; Xu, X. Global negative effects of nutrient enrichment on arbuscular mycorrhizal fungi, plant diversity and ecosystem multifunctionality. New Phytol. 2021, 229, 2957–2969. [Google Scholar] [CrossRef]
- Jia, Y.Y.; Zhang, T.; Walder, F.; Sun, Y.; Shi, Z.Y.; Wagg, C.; Tian, C.Y.; Feng, G. Can mycorrhizal fungi alleviate plant community instability caused by increased precipitation in arid ecosystems? Plant Soil 2022. [Google Scholar] [CrossRef]
- Adnan, M.; Islam, W.; Gang, L.; Chen, H.Y.H. Advanced research tools for fungal diversity and its impact on forest ecosystem. Environ. Sci. Pollut. Res. 2022, 29, 45044–45062. [Google Scholar] [CrossRef]
- Montiel-Rozas, M.D.M.; Lopez-Garcia, A.; Madejon, P.; Madejon, E. Native soil organic matter as a decisive factor to determine the arbuscular mycorrhizal fungal community structure in contaminated soils. Biol. Fertil. Soils 2017, 53, 327–338. [Google Scholar] [CrossRef]
- Maitra, P.; Zheng, Y.; Wang, Y.L.; Mandal, D.; Lu, P.P.; Gao, C.; Babalola, B.J.; Ji, N.N.; Li, X.C.; Guo, L.D. Phosphorus fertilization rather than nitrogen fertilization, growing season and plant successional stage structures arbuscular mycorrhizal fungal community in a subtropical forest. Biol. Fertil. Soils 2021, 57, 685–697. [Google Scholar] [CrossRef]
- Ceulemans, T.; Van, G.M.; Jacquemyn, H.; Boeraeve, M.; Plue, J.; Saar, L.; Kasari, L.; Peeters, G.; van Acker, K.; Crauwels, S.; et al. Arbuscular mycorrhizal fungi in European grasslands under nutrient pollution. Glob. Ecol. Biogeogr. 2019, 28, 1796–1805. [Google Scholar] [CrossRef]
- Ji, B.; Bever, J.D. Plant preferential allocation and fungal reward decline with soil phosphorus enrichment: Implications for mycorrhizal mutualism. Ecosphere 2016, 7, e01256. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.Q.; Gao, W.; Bol, R.; Xiao, Q.; Wu, L.; Zhang, W.J. Microbial regulation of net N mineralisation is driven by C, N, P content and stoichiometry. Eur. J. Soil Sci. 2022, 73, e13257. [Google Scholar] [CrossRef]
- He, D.; Xiang, X.J.; He, J.S.; Wang, C.; Cao, G.M.; Adams, J.; Chu, H.Y. Composition of the soil fungal community is more sensitive to phosphorus than nitrogen addition in the alpine meadow on the Qinghai-Tibetan Plateau. Biol. Fertil. Soils 2016, 52, 1059–1072. [Google Scholar] [CrossRef]
- Garo, G.; Van Geel, M.; Eshetu, F.; Swennen, R.; Honnay, O.; Vancampenhout, K. Arbuscular mycorrhizal fungi community composition, richness and diversity on enset (Ensete ventricosum (Welw.) Cheesman) in Ethiopia is influenced by manure application intensity in low-input farming systems. Plant Soil 2022. [Google Scholar] [CrossRef]
- Wang, G.Z.; Koziol, L.; Foster, B.L.; Bever, J.D. Microbial mediators of plant community response to long-term N and P fertilization: Evidence of a role of plant responsiveness to mycorrhizal fungi. Glob. Change Biol. 2022, 28, 2721–2735. [Google Scholar] [CrossRef] [PubMed]
- Turrini, A.; Bedini, A.; Loor, M.B.; Santini, G.; Sbrana, C.; Giovannetti, M.; Avio, L. Local diversity of native arbuscular mycorrhizal symbionts differentially affects growth and nutrition of three crop plant species. Biol. Fertil. Soils 2018, 54, 203–217. [Google Scholar] [CrossRef]
- Chen, K.; Huang, G.; Li, Y.; Zhang, X.R.; Lei, Y.H.; Li, Y.; Xiong, J.; Sun, Y.F. Illumina MiSeq sequencing reveals correlations among fruit ingredients, environmental factors, and amf communities in three lycium barbarum producing regions of China. Microbiol. Spectr. 2022, 10, e0229321. [Google Scholar] [CrossRef]
- Wang, J.P.; Wang, G.G.; Zhang, B.; Yuan, Z.M.; Fu, Z.Y.; Yuan, Y.D.; Zhu, L.J.; Ma, S.L.; Zhang, J.C. Arbuscular mycorrhizal fungi associated with tree species in a planted forest of Eastern China. Forests 2019, 105, 424. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Neuberger, P.; Daly, E.J.; Gorzelak, M.; Hernandez-Ramirez, G. Arbuscular mycorrhizal fungi community linkages to soil nutrient availability across contrasting agroecosystems. Appl. Soil Ecol. 2022, 176, 104464. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Relative Abundance (Species)/% | Relative Abundance (OTU)/% | Occurrence Frequency/% |
|---|---|---|---|
| Acaulospora | 9.46b | 5.32 | 66.67 |
| Ambispora | 2.18 | 1.09 | 50.00 |
| Archaeospora | 5.72 | 1.82 | 41.67 |
| Diversispora | 4.36 | 3.58 | 66.67 |
| Glomus | 61.29 | 53.58 | 91.67 |
| Pacispora | 2.18 | 0.72 | 33.33 |
| Paraglomus | 4.96 | 5.45 | 66.67 |
| Scutellospora | 2.57 | 0.65 | 33.33 |
| no rank | 0.81 | 0.34 | 8.33 |
| unclassified in Diversisporaceae | 0.76 | 0.24 | 8.33 |
| unclassified in Archaeosporales | 2.18 | 0.85 | 16.67 |
| unclassified in Diversisporales | 1.36 | 0.37 | 16.67 |
| unclassified in Glomeromycetes | 2.18 | 25.97 | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Yang, M.; Shi, Z.; Gao, J.; Wang, X. Biodiversity and Variations of Arbuscular Mycorrhizal Fungi Associated with Roots along Elevations in Mt. Taibai of China. Diversity 2022, 14, 626. https://doi.org/10.3390/d14080626
Zhang M, Yang M, Shi Z, Gao J, Wang X. Biodiversity and Variations of Arbuscular Mycorrhizal Fungi Associated with Roots along Elevations in Mt. Taibai of China. Diversity. 2022; 14(8):626. https://doi.org/10.3390/d14080626
Chicago/Turabian StyleZhang, Mengge, Mei Yang, Zhaoyong Shi, Jiakai Gao, and Xugang Wang. 2022. "Biodiversity and Variations of Arbuscular Mycorrhizal Fungi Associated with Roots along Elevations in Mt. Taibai of China" Diversity 14, no. 8: 626. https://doi.org/10.3390/d14080626
APA StyleZhang, M., Yang, M., Shi, Z., Gao, J., & Wang, X. (2022). Biodiversity and Variations of Arbuscular Mycorrhizal Fungi Associated with Roots along Elevations in Mt. Taibai of China. Diversity, 14(8), 626. https://doi.org/10.3390/d14080626