


The Change in Microbial Diversity and Mycotoxins Concentration in Corn Silage after Addition of Silage Additives


 , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ensilage of Corn Matter
2.2. Microbiological Analyses
2.3. Mycotoxin Analysis
2.4. Statistical Evaluation of Results
3. Results and Discussion
3.1. Microbiota of Corn Silage
3.2. Mycotoxin Composition of Corn Silage
3.3. Relationship between the Mycotoxin Concentrations and Microscopic Fungi and Yeasts, Harmful Effects, and Mycotoxin Limits in Feeds
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, D.; Lee, K.D.; Choi, K.C. Role of LAB in silage fermentation: Effect on nutritional quality and organic acid production-an overview. AIMS Agric. Food 2021, 6, 216–234. [Google Scholar] [CrossRef]
- Fabiszewska, A.U.; Zielińska, K.J.; Wróbel, B. Trends in designing microbial silage quality by biotechnological methods using lactic acid bacteria inoculants: A minireview. World J. Microbiol. Biotechnol. 2019, 35, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jaipolsaen, N.; Sangsritavong, S.; Uengwetwanit, T.; Angthong, P.; Plengvidhya, V.; Rungrassamee, W.; Yammuenart, S. Comparison of the effects of microbial inoculants on fermentation quality and microbiota in napier grass (Pennisetum purpureum) and Corn (Zea mays L.) Silage. Front. Microbiol. 2022, 12, 784535. [Google Scholar] [CrossRef] [PubMed]
- Bryła, M.; Waśkiewicz, A.; Podolska, G.; Szymczyk, K.; Jędrzejczak, R.; Damaziak, K.; Sułek, A. Occurrence of 26 mycotoxins in the grain of cereals cultivated in Poland. Toxins 2016, 8, 160. [Google Scholar] [CrossRef]
- Juan, C.; Covarelli, L.; Beccari, G.; Colasante, V.; Mañes, J. Simultaneous analysis of twenty-six mycotoxins in durum wheat grain from Italy. Food Control. 2016, 62, 322–329. [Google Scholar] [CrossRef]
- Magan, N.; Medina, A.; Aldred, D. Possible climate-change effects on mycotoxin contamination of food crops pre-and postharvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Dzuman, Z.; Zachariasova, M.; Lacina, O.; Veprikova, Z.; Slavikova, P.; Hajslova, J. A rugged high-throughput analytical approach for the determination and quantification of multiple mycotoxins in complex feed matrices. Talanta 2014, 121, 263–272. [Google Scholar] [CrossRef]
- Santini, A.; Meca, G.; Uhlig, S.; Ritieni, A. Fusaproliferin, beauvericin and enniatins: Occurrence in food—A review. World Mycotoxin J. 2012, 5, 71–81. [Google Scholar] [CrossRef]
- Ivanova, L.; Egge-Jacobsen, W.M.; Solhaug, A.; Thoen, E.; Fæste, C.K. Lysosomes as a possible target of enniatin B-induced toxicity in Caco-2 cells. Chem. Res. Toxicol. 2012, 25, 1662–1674. [Google Scholar] [CrossRef]
- Renaud, J.B.; Kelman, M.J.; McMullin, D.R.; Yeung, K.K.C.; Sumarah, M.W. Application of C8 liquid chromatography-tandem mass spectrometry for the analysis of enniatins and bassianolides. J. Chromatogr. A 2017, 1508, 65–72. [Google Scholar] [CrossRef]
- Jajić, I.; Dudaš, T.; Krstović, S.; Krska, R.; Sulyok, M.; Bagi, F.; Savić, Z.; Guljaš, D.; Stankov, A. Emerging Fusarium mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin in Serbian maize. Toxins 2019, 11, 357. [Google Scholar] [CrossRef] [PubMed]
- Kolawole, O.; Graham, A.; Donaldson, C.; Owens, B.; Abia, W.A.; Meneely, J.; Alcorn, M.J.; Connolly, L.; Elliott, C.T. Low doses of mycotoxin mixtures below EU regulatory limits can negatively affect the performance of broiler chickens: A longitudinal study. Toxins 2020, 12, 433. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, A.; Berrada, H.; Ruiz, M.J.; Caloni, F.; Coccini, T.; Spicer, L.J.; Perego, M.C.; Lafranconi, A. A review of the mycotoxin enniatin B. Front. Public Health 2017, 5, 304. [Google Scholar] [CrossRef] [PubMed]
- Kalúzová, M.; Juráček, M.; Bíro, D.; Šimko, M.; Gálik, B.; Rolinec, M.; Hanušovský, O.; Mixtajová, E.; Kolláthová, R.; Brek, P. Mycotoxic contamination of silages: A review. In Proceedings of the NutriNET 2021, Košice, Slovakia, 9 September 2021; University of Veterinary Medicine and Pharmacy in Košice: Košice, Slovakia. [Google Scholar]
- Juráček, M.; Felšöciová, S.; Bíro, D.; Šimko, M.; Gálik, B.; Rolinec, M.; Hanušovský, O.; Kolláthová, R.; Kalúzová, M.; Kačániová, M. The effect of lactic acid bacteria addition on occurrence of microbiota and mycotoxins in rye silages. J. Cent. Eur. Agric. 2022, 23, 342–350. [Google Scholar] [CrossRef]
- Kalúzová, M.; Juráček, M.; Bíro, D.; Gálik, B.; Šimko, M.; Rolinec, M.; Hanušovský, O.; Mixtajová, E.; Kolláthová, R.; Drotárová, S. The effect of additives on mycotoxic contamination of maize silages. J. Cent. Eur. Agric. 2022, 23, 299–304. [Google Scholar] [CrossRef]
- Bíro, D.; Juráček, M.; Kačániová, M.; Šimko, M.; Gálik, B.; Michálková, J.; Gyongyova, E. Occurrence of microscopic fungi and mycotoxins in conserved high moisture corn from Slovakia. Ann. Agric. Environ. Med. 2009, 16, 227–232. [Google Scholar]
- Kung, L., Jr.; Stokes, M.R.; Lin, C. Silage Additives. Silage Sci. Technol. 2003, 42, 305–360. [Google Scholar]
- Schmidt, P.; Mari, L.J.; Nussio, L.G.; de Faria Pedroso, A.; de Fátima Paziani, S.; Wechsler, F.S. Aditivos químicos e biológicos na ensilagem de cana-de-açúcar: Composição química das silagens, ingestão, digestibilidade e comportamento ingestivo. Rev. Bras. de Zootec. 2007, 36, 1666–1675. [Google Scholar] [CrossRef]
- Araki, H.M.C.; De Oliveira, E.R.; Gandra, J.R.; De Goes, R.H.T.B.; Takiya, C.S.; Jacaúna, A.G.; De Oliveira, K.M.P.; Vasques, D.N.; Brandão Cônsolo, N.R.; Del Valle, T.A.; et al. Association of biological and chemical additives on nutrient composition, total losses, microbiological and fermentative profile of sugarcane silage. Iran. J. Appl. Anim. Sci. 2017, 7, 577–584. [Google Scholar]
- Santos, A.P.M.D.; Santos, E.M.; Oliveira, J.S.D.; Ribeiro, O.L.; Perazzo, A.F.; Pinho, R.M.A.; Macêdo, A.J.D.S.; Pereira, G.A. Effects of urea addition on the fermentation of sorghum (Sorghum bicolor) silage. Afr. J. Range Forage Sci. 2018, 35, 55–62. [Google Scholar] [CrossRef]
- Kaewpila, C.; Khota, W.; Gunun, P.; Kesorn, P.; Cherdthong, A. Strategic addition of different additives to improve silage fermentation, aerobic stability and in vitro digestibility of napier grasses at late maturity stage. Agriculture 2020, 10, 262. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.; Chen, S.; Shao, T.; Tao, X.; Yuan, X. Effect of lactic acid bacteria on the fermentation quality and mycotoxins concentrations of corn silage infested with mycotoxigenic fungi. Toxins 2021, 13, 699. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wang, W.; Wang, Z.; Tan, Z.; Qin, G.; Wang, Y.; Pang, H. Microbial population succession and community diversity and its correlation with fermentation quality in soybean meal treated with Enterococcus faecalis during fermentation and aerobic exposure. Microorganisms 2022, 10, 530. [Google Scholar] [CrossRef]
- Kačániová, M.; Kunová, S.; Horská, E.; Nagyová, Ľ.; Puchalski, C.; Haščík, P.; Terentjeva, M. Diversity of microorganisms in the traditional Slovak cheese. Potr. S.J.F.Sci. 2019, 13, 532–537. [Google Scholar] [CrossRef][Green Version]
- Singh, A.; Singh, P.K.; Kumar, A.; Chander, J.; Khanna, G.; Roy, P.; Meis, J.F.; Chowdhary, A. Molecular and matrix-assisted laser desorption ionization–time of flight mass spectrometry-based characterization of clinically significant melanized fungi in India. J. Clin. Microbiol. 2017, 55, 1090–1103. [Google Scholar] [CrossRef] [PubMed]
- SANTE. European Commission. European Commission Health & Consumer Protection Directorate-General. Document No. SANTE 12089/2016. Guidance Document on Identification of Mycotoxins in Food and Feed. Available online: https://ec.europa.eu/food/document/download/f16cac78-9318-4f1f-b2fa-efb25d2f1880_en.pdf (accessed on 20 June 2022).
- Wilkins, R.J. The biochemistry of silage. Anim. Feed Sci. Technol. 1982, 7, 317–318. [Google Scholar] [CrossRef]
- Namihira, T.; Shinzato, N.; Akamine, H.; Maekawa, H.; Matsui, T. Influence of nitrogen fertilization on tropical-grass silage assessed by ensiling process monitoring using chemical and microbial community analyses. J. Appl. Microbiol. 2010, 108, 1954–1965. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Tan, Z.; Gu, L.; Ma, H.; Wang, Z.; Wang, L.; Wu, G.; Qin, G.; Wang, Y.; Pang, H. Variation of microbial community and fermentation quality in corn silage treated with lactic acid bacteria and Artemisia argyi during aerobic exposure. Toxins 2022, 14, 349. [Google Scholar] [CrossRef]
- Krustev, E.; Khristov, B. Mikroflora v tsarevichen silazh Microflora in corn silage. Vet. Med. Nauki. 1981, 18, 88–91. [Google Scholar]
- dos Santos Leandro, E.; Ginani, V.C.; de Alencar, E.R.; Pereira, O.G.; Rose, E.C.P.; do Vale, H.M.M.; Pratesi, R.; Hecht, M.M.; Cavalcanti, M.H.; Tavares, C.S.O. Isolation, identification, and screening of lactic acid bacteria with probiotic potential in silage of different species of forage plants, Cocoa beans, and artisanal salami. Probiot. Antimicrob. Proteins 2020, 13, 173–186. [Google Scholar] [CrossRef]
- Paradhipta, D.H.V.; Lee, S.S.; Kang, B.; Joo, Y.H.; Lee, H.J.; Lee, Y.; Kim, J.; Kim, S.C. Dual-purpose inoculants and their effects on corn silage. Microorganisms 2020, 8, 765. [Google Scholar] [CrossRef] [PubMed]
- Tanizawa, Y.; Kobayashi, H.; Nomura, M.; Sakamoto, M.; Arita, M.; Nakamura, Y.; Ohkuma, M.; Tohno, M. Lactobacillus buchneri subsp. silagei subsp. nov., isolated from rice grain silage. Int. J. Systemat. Evolution. Microbiol 2020, 70, 3111–3116. [Google Scholar] [CrossRef] [PubMed]
- Wuyts, S.; Van Beeck, W.; Allonsius, C.N.; van den Broek, M.F.; Lebeer, S. Applications of plant-based fermented foods and their microbes. Curr. Opin. 2020, 61, 45–52. [Google Scholar] [CrossRef]
- Santos, A.O.; Ávila, C.L.S.; Schwan, R.F. Selection of tropical lactic acid bacteria for enhancing the quality of maize silage. J. Dairy Sci. 2013, 96, 7777–7789. [Google Scholar] [CrossRef]
- Da Silva, N.C.; Dos Santos, J.P.; Ávila, C.L.S.; Evangelista, A.R.; Casagrande, D.R.; Bernardes, T.F. Evaluation of the effects of two Lactobacillus buchneri strains and sodium benzoate on the characteristics of corn silage in a hot-climate environment. Grassl. Sci. 2014, 60, 169–177. [Google Scholar] [CrossRef]
- Danner, H.; Holzer, M.; Mayrhuber, E.; Braun, R. Acetic acid increases stability of silage under aerobic conditions. Appl. Environ. Microbiol. 2003, 69, 562–567. [Google Scholar] [CrossRef]
- Burns, P.; Borgo M., F.; Binetti, A.; Puntillo, M.; Bergamini, C.; Páez, R.; Mazzoni, R.; Reinheimer, J.; Vinderola, G. Isolation, characterization and performance of autochthonous spray dried lactic acid bacteria in maize micro and bucket-silos. Front. Microbiol. 2018, 9, 2861. [Google Scholar] [CrossRef]
- Driehuis, F.; Oude Elferink, S.J.W.H.; Spoelstra, S.F. Anaerobic lactic acid degradation during ensilage of whole crop maize inocu-lated with Lactobacillus buchneri inhibits yeast growth and improves aerobic stability. J. Appl. Microbiol. 1999, 87, 583–594. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Oude Elferink, S.J.W.H.; Krooneman, J.; Gottschal, J.C.; Spoelstra, S.F.; Faber, F.; Driehuis, F. Anaerobic conversion of lactic acid to acetic acid and 1, 2-propanediol by Lactobacillus buchneri. Appl. Environ. Microbiol. 2001, 67, 125–132. [Google Scholar] [CrossRef]
- Krooneman, J.; Faber, F.; Alderkamp, A.C.; Oude Elferink, S.J.W.H.; Driehuis, F.; Cleenwerck, I.; Swings, J.; Gottschal, J.C.; Vancanneyt, M. Lactobacillus diolivorans sp. nov., a 1, 2-propanediol-degrading bacterium isolated from aerobically stable maize silage. Int. J. Syst. Evol. Microbiol. 2002, 52, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Guaragnella, N.; Bobba, A.; Passarella, S.; Marra, E.; Giannattasio, S. Yeast acetic acid-induced programmed cell death can occur without cytochrome c release which requires metacaspase YCA1. FEBS Lett. 2010, 584, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, A.B.; Ascenso, J.R.; Sá-Correia, I. Metabolic insights into the yeast response to propionic acid based on high resolution 1H NMR spectroscopy. Metabolomics 2011, 7, 457–468. [Google Scholar] [CrossRef]
- Ferrero, F.; Piano, S.; Tabacco, E.; Borreani, G. Effects of conservation period and Lactobacillus hilgardii inoculum on the fermentation profile and aerobic stability of whole corn and sorghum silages. J. Sci. Food Agric. 2019, 99, 2530–2540. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.B.; Smith, M.L.; Savage, R.M.; Polukis, S.A.; Drouin, P.; Kung, L. Effects of Lactobacillus hilgardii 4785 and Lactobacillus buchneri 40788 on the bacterial community, fermentation and aerobic stability of high-moisture corn silage. J. Appl. Microbiol. 2020, 130, 1481–1493. [Google Scholar] [CrossRef]
- Drouin, P.; Tremblay, J.; Chaucheyras-Durand, F. Dynamic succession of microbiota during ensiling of whole plant corn following inoculation with Lactobacillus buchneri and Lactobacillus hilgardii alone or in combination. Microorganisms 2019, 7, 595. [Google Scholar] [CrossRef]
- Hu, Z.; Chang, J.; Yu, J.; Li, S.; Niu, H. Diversity of bacterial community during ensiling and subsequent exposure to air in wholeplant maize silage. Asian-australas. J. Anim. 2018, 31, 1464–1473. [Google Scholar] [CrossRef]
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef]
- Bull, L.; Chalupa, W.; Owens, F.; Satter, L.D.; Sniffen, C.J.; Trenkle, A.H.; Waldo, D.R. National Research Council-NRC. Ruminant Nitrogen Usage; The National Academies Press: Washington, DC, USA, 1985; p. 148. [Google Scholar]
- Sumadong, P.; Cherdthong, A.; So, S.; Wanapat, M. Sulfur, fresh cassava root, and urea independently enhanced gas production, ruminal characteristics, and in vitro degradability. BMC Vet. Res. 2021, 17, 304. [Google Scholar] [CrossRef]
- Cherdthong, A.; Wanapat, M. Development of urea products as rumen slow-release feed for ruminant production: A review. Aust. J. Basic Appl. Sci. 2010, 4, 2232–2241. [Google Scholar]
- Pang, H.; Qin, G.; Tan, Z.; Li, Z.; Wang, Y.; Cai, Y. Natural populations of lactic acid bacteria associated with silage fermentation as determined by phenotype, 16S ribosomal RNA and recA gene analysis. Syst. Appl. Microbiol. 2011, 34, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Zhao, J.; Zhu, B.; Su, R.; Pan, Y.; Ma, J.; Zhou, G.; Tao, Y.; Liu, X.; Zhong, J. Assessing the fermentation quality and microbial community of the mixed silage of forage soybean with crop corn or sorghum. Bioresour. Technol. 2018, 265, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Liu, Y.; Wang, S.; Zhang, Y.; Wang, W.; Yang, H.; Lu, N.; Li, S. Nutrient digestibility, microbial fermentation, and response in bacterial composition to methionine dipeptide: An in vitro study. Biology 2022, 11, 93. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Gao, Y.; Tang, M.; Fu, T.; Diao, Q.; Bi, Y.; Tu, Y. Effects of dietary rumen-protected Lys levels on rumen fermentation and bacterial community composition in Holstein heifers. Appl. Microbiol. Biotechnol. 2020, 104, 6623–6634. [Google Scholar] [CrossRef] [PubMed]
- Penagos-Tabares, F.; Khiaosa-Ard, R.; Schmidt, M.; Pacífico, C.; Faas, J.; Jenkins, T.; Nagl, V.; Sulyok, M.; Labuda, R.; Zebeli, Q. Fungal species and mycotoxins in mouldy spots of grass and maize silages in Austria. Mycotoxin Res. 2022, 38, 117–136. [Google Scholar] [CrossRef] [PubMed]
- Whitlow, L.W.; Hagler, W.M. Mycotoxins in dairy cattle: Occurrence, toxicity, prevention and treatment. In Proceedings of the Southwest Nutrition Conference, Tempe, AZ, USA, 24–25 February 2005. [Google Scholar]
- Dong, J.; Li, S.; Chen, X.; Sun, Z.; Sun, Y.; Zhen, Y.; Qin, G.; Wang, T.; Demelash, N.; Zhang, X. Effects of Lactobacillus plantarum inoculation on the quality and bacterial community of whole-crop corn silage at different harvest stages. Res. Sq. preprint. 2022. [Google Scholar] [CrossRef]
- Drouin, P.; Tremblay, J.; Renaud, J.; Apper, E. Microbiota succession during aerobic stability of maize silage inoculated with Lentilactobacillus buchneri NCIMB 40788 and Lentilactobacillus hilgardii CNCM-I-4785. Microbiologyopen 2021, 10, e1153. [Google Scholar] [CrossRef]
- Bakri, M.M. Evaluating the effects of cellulolytic enzymes and Lactobacillus bulgaricus on mycotoxins production and the quality of maize silage. BioResources 2021, 16, 8366–8378. [Google Scholar] [CrossRef]
- Gallo, A.; Bernardes, T.F.; Copani, G.; Fortunati, P.; Giuberti, G.; Bruschi, S.; Bryan, K.A.; Nielsen, N.G.; Witt, K.L.; Masoero, F. Effect of inoculation with Lactobacillus buchneri LB1819 and Lactococcus lactis O224 on fermentation and mycotoxin production in maize silage compacted at different densities. Anim. Feed Sci. Technol. 2018, 246, 36–45. [Google Scholar] [CrossRef]
- Teller, R.S.; Schmidt, R.J.; Whitlow, L.W.; Kung, L., Jr. Effect of physical damage to ears of corn before harvest and treatment with various additives on the concentration of mycotoxins, silage fermentation, and aerobic stability of corn silage. J. Dairy Sci. 2012, 95, 1428–1436. [Google Scholar] [CrossRef]
- Wang, M.; Xu, S.; Wang, T.; Jia, T.; Xu, Z.; Wang, X.; Yu, Z. Effect of inoculants and storage temperature on the microbial, chemical and mycotoxin composition of corn silage. Asian-australas. J. Anim. Sci. 2018, 31, 1903–1912. [Google Scholar] [CrossRef] [PubMed]
- Eckard, S.; Wettstein, F.E.; Forrer, H.R.; Vogelgsang, S. Incidence of Fusarium species and mycotoxins in silage maize. Toxins 2011, 3, 949–967. [Google Scholar] [CrossRef] [PubMed]
- Latorre, A.; Dagnac, T.; Lorenzo, B.F.; Llompart, M. Occurrence and stability of masked fumonisins in corn silage samples. Food Chem. 2015, 189, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Driehuis, F.; Wilkinson, J.M.; Jiang, Y.; Ogunade, I.; Adesogan, A.T. Silage review: Animal and human health risks from silage. J. Dairy Sci. 2018, 101, 4093–4110. [Google Scholar] [CrossRef]
- Zhang, D.; Zhao, L.; Chen, Y.; Gao, H.; Hua, Y.; Yuan, X.; Yang, H. Mycotoxins in maize silage from China in 2019. Toxins 2022, 14, 241. [Google Scholar] [CrossRef]
- Zachariasova, M.; Dzuman, Z.; Veprikova, Z.; Hajkova, K.; Jiru, M.; Vaclavikova, M.; Zachariasova, A.; Pospichalova, M.; Hajslova, J. Occurrence of multiple mycotoxins in European feedingstuffs, assessment of dietary intake by farm animals. Anim. Feed Sci. Technol. 2014, 193, 124–140. [Google Scholar] [CrossRef]
- Reisinger, N.; Schürer-Waldheim, S.; Mayer, E.; Debevere, S.; Antonissen, G.; Sulyok, M.; Nagl, V. Mycotoxin occurrence in maize silage—A neglected risk for bovine gut health? Toxins 2019, 11, 577. [Google Scholar] [CrossRef]
- Sørensen, J.L.; Nielsen, K.F.; Rasmussen, P.H.; Thrane, U. Development of a LC-MS/MS method for the analysis of enniatins and beauvericin in whole fresh and ensiled maize. J. Agric. Food Chem. 2008, 56, 10439–10443. [Google Scholar] [CrossRef]
- Rasmussen, R.R.; Storm, I.M.L.D.; Rasmussen, P.H.; Smedsgaard, J.; Nielsen, K.F. Multi-mycotoxin analysis of maize silage by LC-MS/MS. Anal. Bioanal. Chem. 2010, 397, 765–776. [Google Scholar] [CrossRef]
- Storm, I.M.; Rasmussen, R.R.; Rasmussen, P.H. Occurrence of pre-and post-harvest mycotoxins and other secondary metabolites in Danish maize silage. Toxins 2014, 6, 2256–2269. [Google Scholar] [CrossRef]
- Van Pamel, E.; Verbeken, A.; Vlaemynck, G.; De Boever, J.; Daeseleire, E. Ultrahigh-performance liquid chromatographic–tandem mass spectrometric multimycotoxin method for quantitating 26 mycotoxins in maize silage. J. Agric. Food Chem. 2011, 59, 9747–9755. [Google Scholar] [CrossRef] [PubMed]
- Shimshoni, J.A.; Cuneah, O.; Sulyok, M.; Krska, R.; Galon, N.; Sharir, B.; Shlosberg, A. Mycotoxins in corn and wheat silage in Israel. Food Addit. Contam. Part A Chem. Anal. 2013, 30, 1614–1625. [Google Scholar] [CrossRef] [PubMed]
- McElhinney, C.; Danaher, M.; Elliott, C.T.; O′Kiely, P. Mycotoxins in farm silages–a 2-year I rish national survey. Grass Forage Sci. 2016, 72, 339–352. [Google Scholar] [CrossRef]
- Wambacq, E.; Vanhoutte, I.; Audenaert, K.; De Gelder, L.; Haesaert, G. Occurrence, prevention and remediation of toxigenic fungi and mycotoxins in silage: A review. J. Sci. Food Agric. 2016, 96, 2284–2302. [Google Scholar] [CrossRef]
- Gräfenhan, T.; Patrick, S.K.; Roscoe, M.; Trelka, R.; Gaba, D.; Chan, J.M.; McKendry, T.; Clear, R.M.; Tittlemier, S.A. Fusarium damage in cereal grains from Western Canada. 1. Phylogenetic analysis of moniliformin-producing Fusarium species and their natural occurrence in mycotoxin-contaminated wheat, oats, and rye. J. Agric. Food Chem. 2013, 61, 5425–5437. [Google Scholar] [CrossRef]
- Bazin, I.; Faucet-Marquis, V.; Monje, M.-C.; El Khoury, M.; Marty, J.-L.; Pfohl-Leszkowicz, A. Impact of pH on the stability and the cross-reactivity of ochratoxin A and citrinin. Toxins 2013, 5, 2324–2340. [Google Scholar] [CrossRef]
- Mannaa, M.; Kim, K.D. Influence of temperature and water activity on deleterious fungi and mycotoxin production during grain storage. Mycobiology 2017, 45, 240–254. [Google Scholar] [CrossRef]
- Garcia-Cela, E.; Kiaitsi, E.; Sulyok, M.; Krska, R.; Medina, A.; Petit Damico, I.; Magan, N. Influence of storage environment on maize grain: CO2 production, dry matter losses and aflatoxins contamination. Food Addit. Contam. Part A 2019, 36, 175–185. [Google Scholar] [CrossRef]
- Barug, D.; Bhatnagar, D.; van Egmond, H.; Van Der Kamp, J.; Van Osenbruggen, W.; Visconti, A. The Mycotoxin Factbook: Food & Feed Topics; Wageningen Academic Publishers: Wageningen, The Netherlands, 2006. [Google Scholar]
- Vandicke, J.; Visschere, K.D.; Ameye, M.; Croubels, S.; Saeger, S.D.; Audenaert, K.; Haesaert, G. Multi-mycotoxin contamination of maize silages in Flanders, Belgium: Monitoring mycotoxin levels from seed to feed. Toxins 2021, 13, 202. [Google Scholar] [CrossRef]
- Schenck, J.; Müller, C.; Djurle, A.; Jensen, D.F.; O’Brien, M.; Johansen, A.; Rasmussen, P.H.; Spörndly, R. Occurrence of filamentous fungi and mycotoxins in wrapped forages in Sweden and Norway and their relation to chemical composition and management. Grass Forage Sci. 2019, 74, 613–625. [Google Scholar] [CrossRef]
- Ogunade, I.; Martinez-Tuppia, C.; Queiroz, O.; Jiang, Y.; Drouin, P.; Wu, F.; Vyas, D.; Adesogan, A. Silage review: Mycotoxins in silage: Occurrence, effects, prevention, and mitigation. J. Dairy Sci. 2018, 101, 4034–4059. [Google Scholar] [CrossRef] [PubMed]
- Commission Directive 2003/100/EC of 31 October 2003 Amending Annex I to Directive 2002/32/EC of the European Parliament and of the Council on Undesirable Substances in Animal Feed. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32003L0100&rid=2 (accessed on 20 June 2022).
- Mostrom, M.S.; Jacobsen, B.J. Ruminant mycotoxicosis: An update. Vet. Clin. Food Anim. Pract. 2020, 36, 745–774. [Google Scholar] [CrossRef] [PubMed]
- Commission recommendation 2006/576/EC of 17 august 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2006:229:0007:0009:EN:PDF (accessed on 20 June 2022).
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on mycotoxin issues in ruminants: Occurrence in forages, effects of mycotoxin ingestion on health status and animal performance and practical strategies to counteract their negative effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef] [PubMed]
- Valgaeren, B.; Théron, L.; Croubels, S.; Devreese, M.; Baere, S.D.; Pamel, E.V.; Daeseleire, E.; Boevre, M.D.; Saeger, S.D.; Vidal, A.; et al. The role of roughage provision on the absorption and disposition of the mycotoxin deoxynivalenol and its acetylated derivatives in calves: From field observations to toxicokinetics. Arch. Toxicol. 2018, 93, 293–310. [Google Scholar] [CrossRef] [PubMed]
- Diaz, D.; Hopkins, B.A.; Leonard, L.M.; Hagler, W.M., Jr.; Whitlow, L.W. Effect of fumonisin on lactating dairy cattle. J. Dairy Sci. 2000, 83, 1171. [Google Scholar]
- Roberts, H.L.; Bionaz, M.; Jiang, D.; Doupovec, B.; Faas, J.; Estill, C.T.; Schatzmayr, D.; Duringer, J.M. Effects of deoxynivalenol and fumonisins fed in combination to beef cattle: Immunotoxicity and gene expression. Toxins 2021, 13, 714. [Google Scholar] [CrossRef]
- Vardon, P.; McLaughlin, C.; Nardinelli, C. CAST (Council for Agricultural Science and Technology) Technology Mycotoxins: Risks in Plant, Animal, and Human Systems; Task Force Report, No. 139; Council for Agricultural Science and Technology: Ames, IA, USA, 2003; ISBN 978-1-887383-22-6. [Google Scholar]
- Benford, D.; Ceccatelli, S.; Cottrill, B.; DiNovi, M.; Dogliotti, E.; Edler, L.; Farmer, P.; Fürst, P.; Hoogenboom, L.; Knutsen, H.K.; et al. Scientific opinion on risks for animal and public health related to the presence of nivalenol in food and feed. EFSA J. 2013, 11, 3262. [Google Scholar] [CrossRef]
- Eriksen, G.S.; Pettersson, H.; Lundh, T. Comparative cytotoxicity of deoxynivalenol, nivalenol, their acetylated derivatives and de-epoxy metabolites. Food Chem. Toxicol. 2004, 42, 619–624. [Google Scholar] [CrossRef]
- Benford, D.; Ceccatelli, S.; Cottrill, B.; DiNovi, M.; Dogliotti, E.; Edler, L.; Farmer, P.; Fürst, P.; Hoogenboom, L.; Knutsen, H.K.; et al. EFSA Panel on contaminants in the food chain (CONTAM). Scientific opinion on the risks for human and animal health related to the presence of modified forms of certain mycotoxins in food and feed. EFSA J. 2014, 12, 3916. [Google Scholar] [CrossRef]
- Kouri, K.; Lemmens, M.; Lemmens-Gruber, R. Beauvericin-induced channels in ventricular myocytes and liposomes. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2003, 1609, 203–210. [Google Scholar] [CrossRef]
- Ornelis, V.; Rajkovic, A.; Decleer, M.; Sas, B.; Saeger, S.D.; Madder, A. Counteracting in vitro toxicity of the ionophoric mycotoxin beauvericin—synthetic receptors to the rescue. J. Org. Chem. 2019, 84, 10422–10435. [Google Scholar] [CrossRef] [PubMed]
- Maranghi, F.; Tassinari, R.; Narciso, L.; Tait, S.; Rocca, C.L.; Felice, G.D.; Butteroni, C.; Corinti, S.; Barletta, B.; Cordelli, E.; et al. In vivo toxicity and genotoxicity of beauvericin and enniatins. Combined approach to study in vivo toxicity and genotoxicity of mycotoxins geauvericin (BEA) and enniatin B (ENNB). EFSA Support. Publ. 2018, 15, 1406E. [Google Scholar] [CrossRef]
- Huang, C.-H.; Wang, F.-T.; Chan, W.-H. Enniatin B1 exerts embryotoxic effects on mouse blastocysts and induces oxidative stress and immunotoxicity during embryo development. Environ. Toxicol. 2019, 34, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; DiNovi, M.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Risks to human and animal health related to the presence of moniliformin in food and feed. EFSA J. 2018, 16, 5082. [Google Scholar] [CrossRef]
- Lamprecht, S.C. Incidence and toxigenicity of seedborne Fusarium species from annual Medicago species in South Africa. Phytopathology 1986, 76, 1040–1042. [Google Scholar] [CrossRef]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Risks to human and animal health related to the presence of deoxyniva-lenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, e04718. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycotoxin Group | Analyte |
|---|---|
| Aflatoxins | Aflatoxin B1, Aflatoxin B2, Aflatoxin G1, Aflatoxin G2 |
| Alternaria Metabolites | Alternariol |
| Aspergillus Metabolites | Gliotoxin, Sterigmatocystin |
| A-Trichothecene | Diacetoxyscirpenol, HT-2 Toxin, Neosolaniol, T-2 Toxin, T-2 Triol, T-2-Tetraol |
| B-Trichothecene | Deoxynivalenol, Deoxynivalenol-3-Glucoside, 15Acetoxyscirpenol, 15AcetylDeoxynivalenol, 3AcetylDeoxynivalenol, Nivalenol |
| Enniatins and Beauvericin | Beauvericin, Enniatin A, Enniatin A1, Enniatin B, Enniatin B1 |
| Ergot Alkaloids | Agroclavine, alpha-Ergocryptine, alpha-Ergocryptinine, Dihydrolysergol, Elymoclavine, Ergine, Ergocornine, Ergocorninine, Ergocristine, Ergocristinine, Ergometrine, Ergometrinine, Ergosine, Ergotamine |
| Fumonisins | Fumonisin B1, Fumonisin B2, Fumonisin B3 |
| Fusarium Metabolites | Fusarenon X, Moniliformin |
| Ochratoxins | Ochratoxin A, Ochratoxin B |
| Penicillium Metabolites | Mycophenolic Acid, Patulin, Penicillic Acid, Roquefortine C |
| Zearalenone + Metabolites | alpha-Zearalenol, beta-Zearalenol, Zearalanone |
| MSE | MRS | APT | MC | EA | TSA | MEA | |
|---|---|---|---|---|---|---|---|
| CONT1 | ND | 5.11 | ND | ND | ND | 3.89 | 2.84 |
| CONT2 | ND | 5.08 | ND | ND | ND | 3.21 | 2.58 |
| CONT3 | ND | 4.22 | 2.69 | ND | ND | 3.52 | 2.78 |
| Σ CONT | ND | 4.81 ± 0.48 a | 2.69 ± 0.36 a | ND | ND | 3.54 ± 0.43 | 2.73 ± 0.33 a |
| ALAB1 | ND | 5.05 | 3.21 | ND | ND | 3.46 | 2.21 |
| ALAB2 | ND | 4.11 | 3.32 | ND | ND | 3.23 | 2.13 |
| ALAB3 | 2.83 | 4.12 | 3.12 | ND | ND | 3.51 | 2.15 |
| Σ ALAB | / | 4.43 ± 0.49 a,b | 3.22 ± 0.21 b | ND | ND | 3.40 ± 0.20 | 2.16 ± 0.16 b |
| NAUR1 | ND | 4.21 | ND | ND | ND | 3.34 | 3.83 |
| NAUR2 | ND | 4.15 | ND | ND | ND | 3.25 | 3.41 |
| NAUR3 | ND | 4.14 | ND | ND | ND | 3.18 | 2.89 |
| Σ NAUR | ND | 4.17 ± 0.13 b | ND | ND | ND | 3.26 ± 0.22 | 3.38 ± 0.45 c |
| Total | / | 4.47 ± 0.47 | 3.09 ± 0.34 | / | / | 3.40 ± 0.31 | 2.76 ± 0.60 |
| p value | / | 0.010 | 0.010 | / | / | 0.157 | 0.000 |
| Species of Bacteria/Sample | CONT | ALAB | NAUR | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | ||
| Acinetobacter radioresistens | 3 | 3 | ||||||||
| Arthrobacter crystallopoietes | 4 | 4 | ||||||||
| Alkalihalobacillus gibsonii | 2 | 3 | 5 | |||||||
| Priestia megaterium | 3 | 4 | 4 | 11 | ||||||
| Bacillus pumilus | 4 | 5 | 9 | |||||||
| Bacillus safensis | 3 | 3 | 3 | 9 | ||||||
| Bacillus spp. | 2 | 3 | 4 | 9 | ||||||
| Blastomonas natatoria | 2 | 2 | ||||||||
| Cellulosimicrobium cellulans | 3 | 3 | ||||||||
| Citrobacter freundii | 4 | 5 | 9 | |||||||
| Clostridium cadaveris | 2 | 3 | 5 | |||||||
| Lacrimispora sphenoides | 3 | 3 | ||||||||
| Cryptococcus neoformans | 3 | 3 | ||||||||
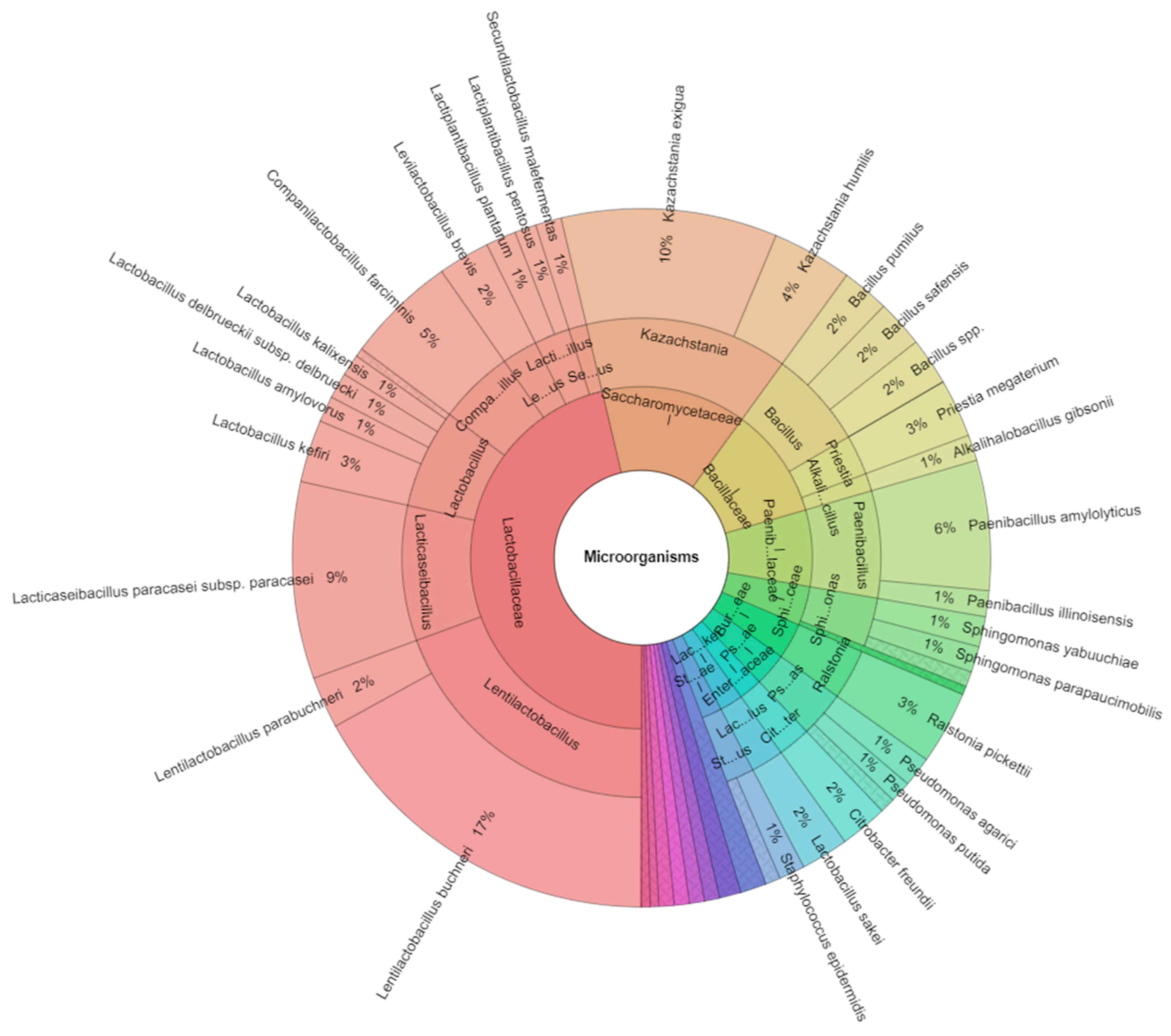
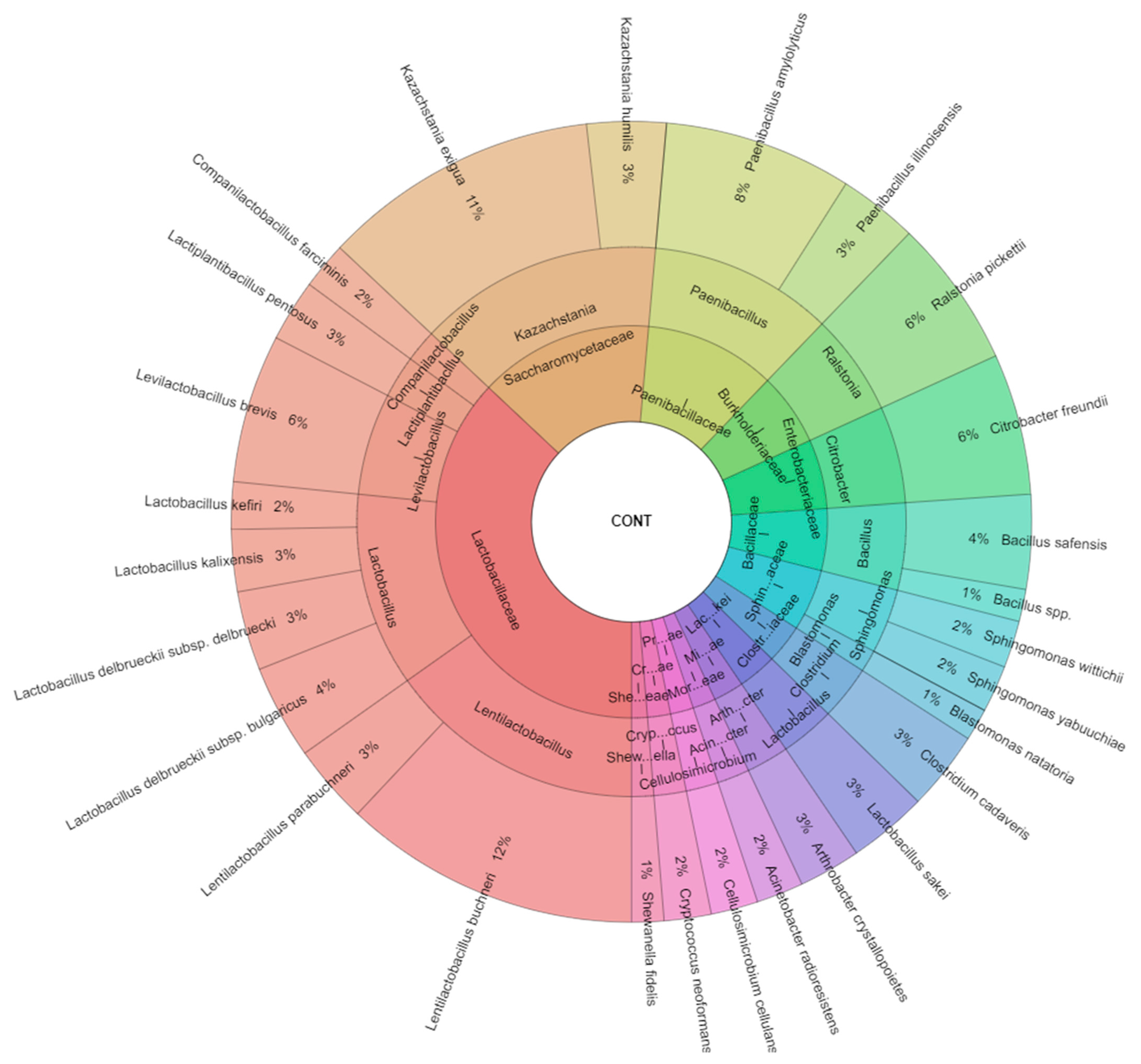
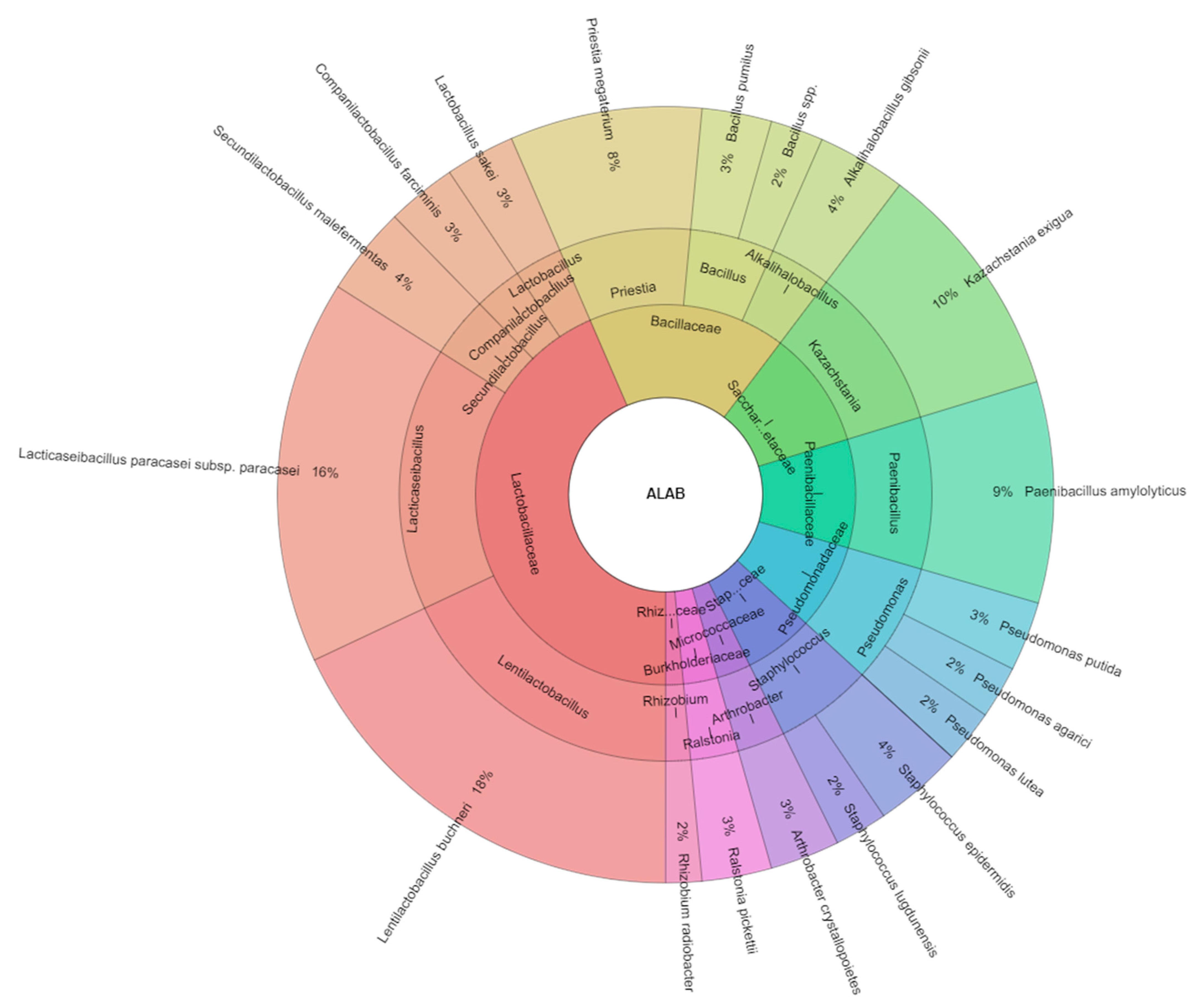
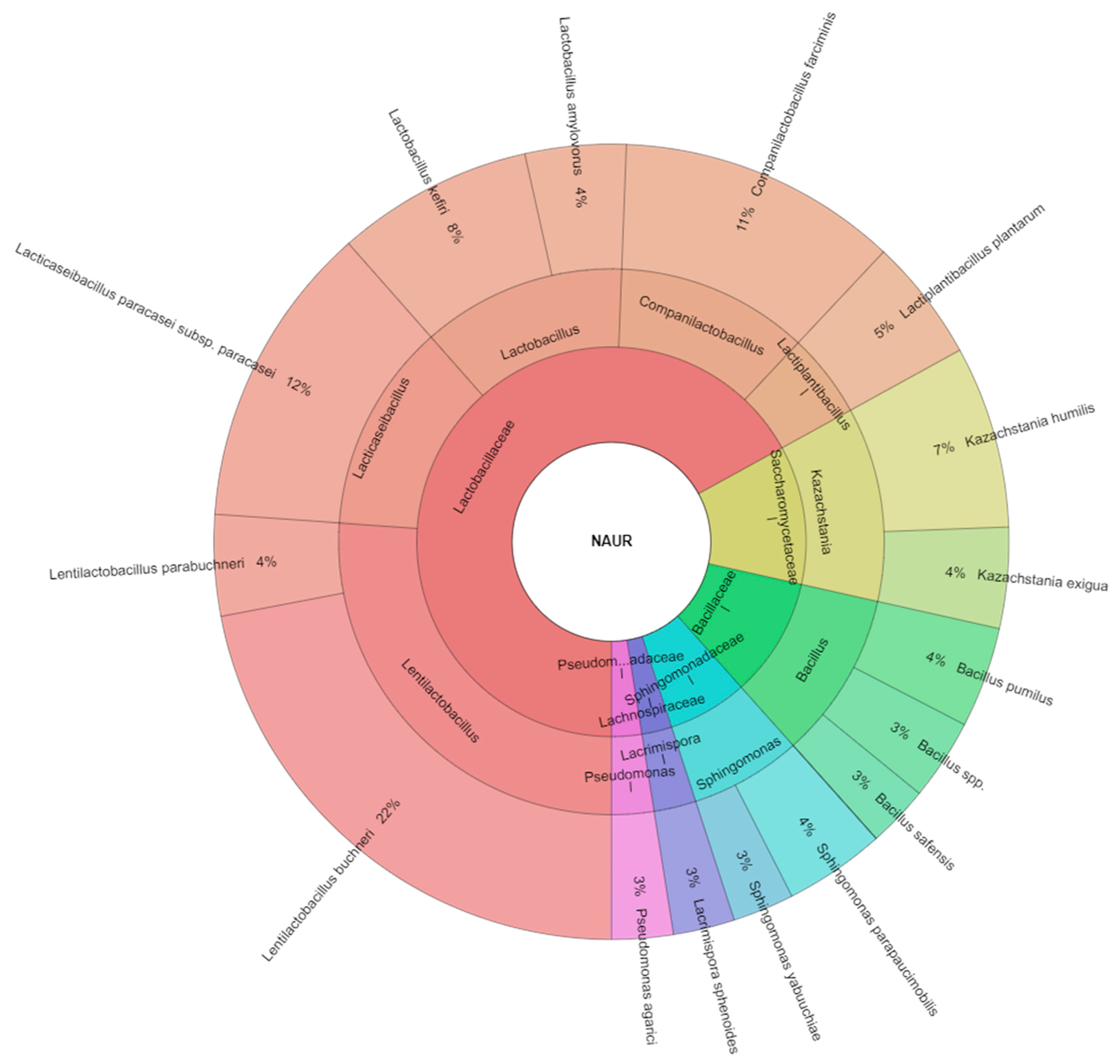
| Kazachstania exigua | 7 | 5 | 6 | 4 | 5 | 6 | 5 | 38 | ||
| Kazachstania humilis | 5 | 4 | 6 | 15 | ||||||
| Lactobacillus amylovorus | 5 | 5 | ||||||||
| Levilactobacillus brevis | 5 | 5 | 10 | |||||||
| Lentilactobacillus buchneri | 5 | 6 | 8 | 10 | 9 | 6 | 9 | 10 | 7 | 70 |
| Lactobacillus delbrueckii subsp. delbruecki | 5 | 5 | ||||||||
| Lactobacillus delbrueckii subsp. bulgaricus | 6 | 6 | ||||||||
| Companilactobacillus farciminis | 3 | 4 | 5 | 3 | 6 | 21 | ||||
| Lactobacillus kalixensis | 4 | 4 | ||||||||
| Lactobacillus kefiri | 3 | 4 | 5 | 12 | ||||||
| Secundilactobacillus malefermentas | 5 | 5 | ||||||||
| Lentilactobacillus parabuchneri | 5 | 5 | 10 | |||||||
| Lacticaseibacillus paracasei subsp. paracasei | 5 | 8 | 10 | 7 | 8 | 38 | ||||
| Lactiplantibacillus pentosus | 4 | 4 | ||||||||
| Lactiplantibacillus plantarum | 6 | 6 | ||||||||
| Lactobacillus sakei | 5 | 4 | 9 | |||||||
| Paenibacillus amylolyticus | 5 | 4 | 3 | 4 | 6 | 3 | 25 | |||
| Paenibacillus illinoisensis | 5 | 5 | ||||||||
| Pseudomonas agarici | 3 | 3 | 6 | |||||||
| Pseudomonas lutea | 3 | 3 | ||||||||
| Pseudomonas putida | 4 | 4 | ||||||||
| Ralstonia pickettii | 5 | 3 | 2 | 4 | 14 | |||||
| Rhizobium radiobacter | 2 | 2 | ||||||||
| Shewanella fidelis | 2 | 2 | ||||||||
| Sphingomonas parapaucimobilis | 2 | 3 | 5 | |||||||
| Sphingomonas wittichii | 3 | 3 | ||||||||
| Sphingomonas yabuuchiae | 3 | 3 | ||||||||
| Sphingomonas yabuuchiae | 3 | 3 | ||||||||
| Staphylococcus epidermidis | 5 | 5 | ||||||||
| Staphylococcus lugdunensis | 3 | 3 | ||||||||
| Total isolates | 63 | 33 | 63 | 47 | 35 | 54 | 42 | 34 | 45 | 416 |
| Species | Genera | Family | % |
|---|---|---|---|
| Acinetobacter radioresistens | Acinetobacter | Moraxellaceae | 0.7 |
| Arthrobacter crystallopoietes | Arthrobacter | Micrococcaceae | 1.0 |
| Alkalihalobacillus gibsonii | Alkalihalobacillus | Bacillaceae | 1.2 |
| Priestia megaterium | Priestia | Bacillaceae | 2.7 |
| Bacillus pumilus | Bacillus | Bacillaceae | 2.2 |
| Bacillus safensis | Bacillus | Bacillaceae | 2.2 |
| Bacillus spp. | Bacillus | Bacillaceae | 2.2 |
| Blastomonas natatoria | Blastomonas | Sphingomonadaceae | 0.4 |
| Cellulosimicrobium cellulans | Cellulosimicrobium | Promicromonosporaceae | 0.7 |
| Citrobacter freundii | Citrobacter | Enterobacteriaceae | 2.2 |
| Clostridium cadaveris | Clostridium | Clostridiaceae | 1.2 |
| Lacrimispora sphenoides | Lacrimispora | Lachnospiraceae | 0.7 |
| Cryptococcus neoformans | Cryptococcus | Cryptococcaceae | 0.7 |
| Kazachstania exigua | Kazachstania | Saccharomycetaceae | 10.0 |
| Kazachstania humilis | Kazachstania | Saccharomycetaceae | 3.7 |
| Lactobacillus amylovorus | Lactobacillus | Lactobacillaceae | 1.2 |
| Levilactobacillus brevis | Levilactobacillus | Lactobacillaceae | 2.4 |
| Lentilactobacillus buchneri | Lentilactobacillus | Lactobacillaceae | 17.0 |
| Lactobacillus delbrueckii subsp. delbruecki | Lactobacillus | Lactobacillaceae | 1.2 |
| Lactobacillus delbrueckii subsp. bulgaricus | Lactobacillus | Lactobacillaceae | 0.4 |
| Companilactobacillus farciminis | Companilactobacillus | Lactobacillaceae | 5.1 |
| Lactobacillus kalixensis | Lactobacillus | Lactobacillaceae | 1.0 |
| Lactobacillus kefiri | Lactobacillus | Lactobacillaceae | 2.9 |
| Secundilactobacillus malefermentas | Secundilactobacillus | Lactobacillaceae | 1.2 |
| Lentilactobacillus parabuchneri | Lentilactobacillus | Lactobacillaceae | 2.4 |
| Lacticaseibacillus paracasei subsp. paracasei | Lacticaseibacillus | Lactobacillaceae | 9.1 |
| Lactiplantibacillus pentosus | Lactiplantibacillus | Lactobacillaceae | 1.0 |
| Lactiplantibacillus plantarum | Lactiplantibacillus | Lactobacillaceae | 1.4 |
| Lactobacillus sakei | Lactobacillus | Lactobacillaceae | 2.2 |
| Paenibacillus amylolyticus | Paenibacillus | Paenibacillaceae | 6.0 |
| Paenibacillus illinoisensis | Paenibacillus | Paenibacillaceae | 1.2 |
| Pseudomonas agarici | Pseudomonas | Pseudomonadaceae | 1.4 |
| Pseudomonas lutea | Pseudomonas | Pseudomonadaceae | 0.7 |
| Pseudomonas putida | Pseudomonas | Pseudomonadaceae | 1.0 |
| Ralstonia pickettii | Ralstonia | Burkholderiaceae | 3.4 |
| Rhizobium radiobacter | Rhizobium | Rhizobiaceae | 0.4 |
| Shewanella fidelis | Shewanella | Shewanellaceae | 0.4 |
| Sphingomonas parapaucimobilis | Sphingomonas | Sphingomonadaceae | 1.2 |
| Sphingomonas wittichii | Sphingomonas | Sphingomonadaceae | 0.7 |
| Sphingomonas yabuuchiae | Sphingomonas | Sphingomonadaceae | 0.7 |
| Sphingomonas yabuuchiae | Sphingomonas | Sphingomonadaceae | 0.7 |
| Staphylococcus epidermidis | Staphylococcus | Staphylococcaceae | 1.2 |
| Staphylococcus lugdunensis | Staphylococcus | Staphylococcaceae | 0.7 |
| Mycotoxin Species | Variant | Mean | S.D. | Minimum | Maximum | p Value |
|---|---|---|---|---|---|---|
| Deoxynivalenol | CONT | 163.46 | 80.23 | 117.03 | 256.10 | 0.404 |
| ALAB | 260.85 | 250.09 | 116.33 | 549.62 | ||
| NAUR | 373.33 | 158.00 | 215.00 | 531.00 | ||
| Total | 265.88 | 178.21 | 116.33 | 549.62 | / | |
| Aflatoxins | CONT | <0.3/1 * | / | / | / | / |
| ALAB | <0.3/1 * | / | / | / | ||
| NAUR | <0.3/1 * | / | / | / | ||
| Total | <0.3/1 * | / | / | / | ||
| Ochratoxins | CONT | <0.5/2 * | / | / | / | / |
| ALAB | <0.5/2 * | / | / | / | ||
| NAUR | <0.5/2 * | / | / | / | ||
| Total | <0.5/2 * | / | / | / | ||
| Zearalenone | CONT | 56.61 | 43.09 | 31.52 | 106.36 | 0.626 |
| ALAB | 36.47 | 6.89 | 28.57 | 41.23 | ||
| NAUR | 41.03 | 8.55 | 36.00 | 50.90 | ||
| Total | 44.71 | 24.04 | 28.57 | 106.36 | / | |
| Fumonisin B1 | CONT | 94.35 | 42.47 | 50.75 | 135.59 | 0.076 |
| ALAB | 96.71 | 54.95 | 57.02 | 159.42 | ||
| NAUR | 208.33 | 67.16 | 144.00 | 278.00 | ||
| Total | 133.13 | 74.27 | 50.75 | 278.00 | / | |
| Nivalenol | CONT | 203.26 | 108.71 | 140.44 | 328.79 | 0.277 |
| ALAB | 592.26 | 454.57 | 139.52 | 1048.63 | ||
| NAUR | 331.33 | 48.21 | 277.00 | 369.00 | ||
| Total | 375.62 | 290.98 | 139.52 | 1048.63 | / | |
| Type A trichothecenes | CONT | <3/10 * | / | / | / | / |
| ALAB | <3/10 * | / | / | / | ||
| NAUR | <3/10 * | / | / | / | ||
| Total | <3/10 * | / | / | / | ||
| Ergot alkaloids | CONT | <2/6 * | / | / | / | / |
| ALAB | <2/6 * | / | / | / | ||
| NAUR | <2/6 * | / | / | / | ||
| Total | <2/6 * | / | / | / |
| Mycotoxin Species | Variant | Mean | S.D. | Minimum | Maximum | p-Value |
| Beauvericin | CONT | <2/6 * | / | / | / | / |
| ALAB | <2/6 * | / | / | / | ||
| NAUR | 496.67 | 175.51 | 355.00 | 693.00 | ||
| Total | 496.67 | 175.51 | 355.00 | 693.00 | ||
| Enniatin A1 | CONT | 16.36 | 14.78 | 5.72 | 33.24 | 0.428 |
| ALAB | 6.67 | 1.99 | 5.30 | 8.95 | ||
| NAUR | 13.17 | 0.99 | 12.50 | 14.30 | ||
| Total | 12.07 | 8.61 | 5.30 | 33.24 | / | |
| Enniatin B | CONT | 44.99 | 48.93 | 16.49 | 101.49 | 0.368 |
| ALAB | 14.55 | 5.05 | 9.21 | 19.25 | ||
| NAUR | 13.53 | 0.78 | 12.90 | 14.40 | ||
| Total | 24.36 | 29.06 | 9.21 | 101.49 | / | |
| Enniatin B1 | CONT | 30.50 | 37.58 | 4.50 | 73.59 | 0.463 |
| ALAB | 7.11 | 2.42 | 4.37 | 8.95 | ||
| NAUR | 16.57 | 1.46 | 15.40 | 18.20 | ||
| Total | 18.06 | 21.42 | 4.37 | 73.59 | / | |
| Moniliformin | CONT | 42.30 | 12.32 | 28.56 | 52.34 | 0.323 |
| ALAB | 24.73 | 18.64 | 13.95 | 46.25 | ||
| NAUR | 35.13 | 3.55 | 31.30 | 38.30 | ||
| Total | 34.06 | 13.65 | 13.95 | 52.34 | / |
| DON | ZEA | FUMB1 | NIV | ENNA1 | ENNB | ENNB1 | MON | |
|---|---|---|---|---|---|---|---|---|
| MaY | 0.567 | 0.179 | 0.876 | −0.629 | 0.629 | −0.066 | 0.367 | 0.558 |
| DON | −0.709 | 0.894 | 0.283 | −0.283 | −0.859 | −0.558 | −0.368 | |
| ZEA | −0.318 | −0.877 | 0.877 | 0.970 | 0.981 | 0.916 | ||
| FUM | −0.176 | 0.176 | −0.540 | −0.128 | 0.088 | |||
| NIV | −1.000 ** | −0.734 | −0.954 | −0.996 * | ||||
| ENNA1 | 0.734 | 0.954 | 0.996 * | |||||
| ENNB | 0.904 | 0.791 | ||||||
| ENNB1 | 0.977 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalúzová, M.; Kačániová, M.; Bíro, D.; Šimko, M.; Gálik, B.; Rolinec, M.; Hanušovský, O.; Felšöciová, S.; Juráček, M. The Change in Microbial Diversity and Mycotoxins Concentration in Corn Silage after Addition of Silage Additives. Diversity 2022, 14, 592. https://doi.org/10.3390/d14080592
Kalúzová M, Kačániová M, Bíro D, Šimko M, Gálik B, Rolinec M, Hanušovský O, Felšöciová S, Juráček M. The Change in Microbial Diversity and Mycotoxins Concentration in Corn Silage after Addition of Silage Additives. Diversity. 2022; 14(8):592. https://doi.org/10.3390/d14080592
Chicago/Turabian StyleKalúzová, Mária, Miroslava Kačániová, Daniel Bíro, Milan Šimko, Branislav Gálik, Michal Rolinec, Ondrej Hanušovský, Soňa Felšöciová, and Miroslav Juráček. 2022. "The Change in Microbial Diversity and Mycotoxins Concentration in Corn Silage after Addition of Silage Additives" Diversity 14, no. 8: 592. https://doi.org/10.3390/d14080592
APA StyleKalúzová, M., Kačániová, M., Bíro, D., Šimko, M., Gálik, B., Rolinec, M., Hanušovský, O., Felšöciová, S., & Juráček, M. (2022). The Change in Microbial Diversity and Mycotoxins Concentration in Corn Silage after Addition of Silage Additives. Diversity, 14(8), 592. https://doi.org/10.3390/d14080592