Abstract
The conservation of pure wolf populations in Europe is currently threatened by anthropogenic hybridization with dogs, through potential extinction of local gene pools due to replacement with domestic gene variants. Distinction of wolf–dog hybrids from wild ancestors is essential for development and implementation of management and conservation plans. Morphological traits are rarely helpful, and a genetic approach is the most effective to distinguish admixed individuals back to old backcrosses. To provide a molecular tool to address the issue, we optimized and validated a cost-effective protocol in multiplexed PCR format based on 22 STR canine loci, which allows us to distinguish genetically pure wolves from dogs, and, when used in association with a Bayesian assignment approach, is capable of statistically assigning admixed individuals to classes of hybrids with different levels of dog ancestry. Our method demonstrated high reliability, showing full repeatability and reproducibility of data with as little as 0.125 ng of genomic DNA, and was therefore suitable for the analysis of non-invasively collected samples and degraded DNA. The application of our STR panel to the appropriate assignment procedure unambiguously defined two genetically separated clusters for wolves and dogs, and successfully identified known hybrids as admixed individuals, which eventually were classified as recent hybrids and older backcrosses. The protocol, which is described here in detail, can be adopted by various laboratories that need this kind of diagnosis; furthermore, it would be capable of producing concordant results through inter-laboratory comparisons with wolf and dog control DNAs.
Keywords:
multiplex STR panel; management; validation; wolf; dog; hybrids; conservation; Bayesian assignment 1. Introduction
The grey wolf (Canis lupus Linnaeus, 1758) is granted protection throughout Europe according to EU and national laws [1]. Nevertheless, wolf conservation faces serious threats in many European countries, from illegal killing and road accidents to anthropogenic hybridization with the dog (C. l. familiaris). The latter, in particular, is a sneaky threat, because introgression can lead to local extinction of the wolf gene pool in the wild through replacement with domestic gene variants. Dog–wolf gene flow is potentially negatively affecting the long-term survival of pure wolf populations in several areas of their distribution [2].
Dogs were domesticated from wolves some 30–15,000 years ago [3], and wolf-dog interbreeding has occurred as a recurrent event over time [4]. As a consequence, differentiating between genetically pure wolves, dogs and their hybrids is far from trivial, and the ambition to trace the exact generation with domestic ancestry for older backcrossed individuals is virtually unrealistic (cf. [2] and references therein). On the other hand, their distinction, at least in terms of inclusive classes/categories of hybrids with different levels of introgression, is an essential requirement for conservation and management decisions [5], as well as for forensic purposes [6].
Animal DNA profiling through panels of autosomal Short Tandem Repeat (STR) loci is widely used in a variety of applications requiring highly polymorphic genetic systems, such as research (phylogeny, conservation genetics, differentiation of closely related species, genetic linkage analysis, association mapping), diagnosis (parentage, pedigree assessment) and forensic testing (individual identification, DNA match).
Different methods are currently available for DNA typing in wild canids [7,8], but validation studies are lacking for most of them. On the other hand, STR-based commercial kits were developed and validated for some domestic animals, like dogs and cattle, while they are not available for non-model and wild species [9]. Specifically, commercial canine panels have been developed and validated to maximize intraspecific genetic variability for individual identification in forensics, kinship and breed testing in dogs. Therefore, genetic differentiation between dogs and wolves has not been the target of any kit supplied to the market so far.
Here we present a standardized molecular protocol based on a custom-made panel of canine STR markers with high variability between taxa and low variability within each taxon. Three multiplexed PCR reactions were developed that enabled co-amplification of 22 loci under the same PCR conditions, and basic validation steps were conducted to determine reliability, reproducibility and limitations of the assay.
The STR data obtained with the validated protocol can be combined with a model-based Bayesian statistical technique to assign multi-locus profiles to clusters of genetically similar individuals [10]. In order to test the effectiveness of our standardized 22-locus panel, in the present study a Bayesian approach was applied to the genetic profiles from 100 reference samples (50 wolves and 50 dogs) to define two assignment genetic classes, pure wolves and dogs, and to identify wolf × dog hybrids as recently admixed individuals and later backcrosses (introgressed wolves with minimal domestic ancestry).
2. Materials and Methods
2.1. Selection of STR Markers
Twenty-two canine STR markers were selected from literature data on wolf and dog genetics (cf. [11,12,13,14]), based on the following features: 1. High level of inter-taxon polymorphism, 2. Ease of locus amplification, 3. Easy-to-interpret electropherograms, 4. Possibility of multiplexing with clear resolution of bands, and 5. Small size of fragments for amplification in non-invasive and forensic samples. The selected markers (Table 1) are in linkage equilibrium (showing no co-segregation), with the exception of CPH16, C20.253, C20.622 and PRK-CD that are syntenic loci, forming a linkage group mapping in chromosome 20 (CanFam2.0, https://www.ncbi.nlm.nih.gov, accessed on 13 December 2021). The latter have been included in the STR panel to improve the genetic discrimination of hybrids from recently admixed taxa, as suggested when data are statistically processed using the “linkage model” of the Bayesian algorithm (cf. [13,15]), as implemented in STRUCTURE v 2.3.4 (see below).

Table 1.
STR loci, primer sequences, fluorescent dye labels at 5′ of the forward primer, final concentration of forward and reverse primers in their respective multiplex (M1, M2, M3), allelic ranges in 100 reference samples (50 Apennine wolves and 50 dogs), and repeat length (2 = dinucleotide, 4 = tetranucleotide).
In order to encompass loci that maximized the variability between wolf and dog and met our specific criteria, the STR panel included as many as 17 loci with dinucleotide repeats. These are more prone than tri- or tetranucleotides to amplification of stutter bands, broken repeats, and unbalanced alleles in heterozygotes, which makes their interpretation and standardization potentially challenging (cf. [9]). However, the selected dinucleotide loci showed high levels of dog-wolf polymorphism (and, ultimately, relative ease of interpretation), thus making their use particularly suitable for our purposes.
2.2. Sample Collection and DNA Extraction
The samples used for method development and validation, as well as for databasing, consisted of muscle tissues from Apennine wolves and dogs that died of disease, accidentally, or due to direct and indirect poaching. Carcasses were recovered from central and southern Italy by local authorities (public veterinary service, rangers of State Forestry Service, provincial police, guards of protected areas, animal welfare associations), and delivered to Centro di Referenza Nazionale per la Medicina Forense Veterinaria at the Istituto Zooprofilattico Sperimentale delle Regioni Lazio e Toscana “M. Aleandri” for diagnostic or forensic purposes. Individuals were classified as pure wolves by expert veterinarians and zoologists based on morphology and body mass [16,17].
Five samples (3 wolves and 2 dogs) were used in the validation steps, while samples from 50 wolves and 50 medium- and large-sized dogs, both mongrels and pure bred (Siberian husky, golden retriever, German shepherd, English setter, Labrador, rottweiler, border collie, Maremma sheepdog, Belgian shepherd, Alaskan Malamute, bracco italiano), were used for databasing and genotype assignments (see below). Ten known admixed wolf x dog individuals (first-generation offspring of a female wolf and a male dog, as well as individuals showing clear dog-like morphological traits, cf. [5,16,17]), were identified by qualified wildlife scientists and genotyped with the validated protocol to check for signals of genetic admixture. Muscle tissues were stored at −20 °C or in five volumes of 95% ethanol at room temperature (or at +4 °C) until processed. DNA was isolated from approximately 15 mg of muscle using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) and the Maxwell16 Instrument (Promega, Madison, WI, USA) for automated genomic DNA isolation following the user handbooks, and diluted in 200 µL RNAse-free molecular grade water. DNA quantification was performed using the Quantus Fluorometer (Promega) according to the manufacturer’s instructions. Extracted DNA was stored at +4 °C until amplification steps. One mock tube with reagents and no sample DNA was included in each extraction session.
2.3. Multiplex PCR Amplification
PCR conditions for co-amplification of 22 STR markers were optimized for three multiplexes (M1, M2, M3, Table 1). The combinations of primers and fluorescent dye labeling blended in each multiplex were set up according to the allele range at single locus (to avoid overlap of ranges in the same color channel) and lack of interference among primer pairs. Optimization involved basically primer concentrations for inter-locus balance, while the effects of multiplexing (e.g., artifacts) were evaluated through direct observation of the electropherograms to check for amplification of aspecific products inside and outside the defined allelic ranges, that might hinder and yield misinterpretations in allele scoring. After optimization, each multiplex contained 2 µL of genomic DNA, primers at the concentrations listed in Table 1, 3.6 µL of master mix (Qiagen Multiplex PCR Kit, code 206143, Hilden, Germany), and PCR grade H2O in 12 µL total reaction volume. Multiplexed PCR tubes were loaded onto an ABI Veriti® 96-Well Thermal Cycler (Applied Biosystems, Waltham, MA, USA) under the following thermal cycling conditions: an initial 15 min activation step at 95 °C, followed by 32 cycles of denaturation at 94 °C for 30 s, annealing at 58 °C for 90 s, and extension at 72 °C for 60 s, and a final extension step at 60 °C for 30 min. Extraction negative controls and PCR mock tubes were included in each amplification round both in validation steps and databasing.
2.4. Visualization of STR Genotypes
A volume of 2 μL of five-fold diluted amplified reaction products were denaturated with 14 μL HiDiTM formamide (Applied Biosystems, code 4311320) at room temperature and combined with 0.2 μL GeneScanTM 500 LIZTM size standard (Applied Biosystems, code 4322682) for fragment sizing on an ABI PrismTM 3130 Genetic Analyzer (Applied Biosystems). The end-labeled primers were compatible with filter dye set G5, requiring the use of DS-33 Dye Primer Matrix Standard (Applied Biosystems, code 4345833) for optimized allele calling. GeneMapper Software 5.0 (Applied Biosystems, Waltham, MA, USA) was used for fragment sizing and automated allele identification. Unless otherwise stated, default settings were used in the “Analysis Method”, selecting the “Advanced” peak detection algorithm in combination with the “Local Southern” method for implemented fragment sizing. Since our panel consisted of STR with tandem repeat units of not less than two nucleotides, the maximum offset of each bin for allele calling in the bin sets was rounded off to integer values at ±1 bp in the “Panel Manager”.
2.5. Validation
We conducted a basic validation study, following the revised guidelines and recommendations of the Scientific Working Group on DNA Analysis Methods [18] for forensic testing.
Genomic DNA samples from 5 individuals (3 wolves and 2 dogs) were used as independent replicates to assess reliability of the markers and reproducibility of our multiplex PCR assays. We evaluated sensitivity and repeatability of the method, determining the template Limit Of Detection (LOD) for reliable profiles, and variation in the performance of different replicates, respectively.
As part of the validation process, we also evaluated whether the amplification of possible locus-specific backstutters, plusA peaks, and peak imbalance in the heterozygotes affected the interpretation of electropherograms and correct allele calling. Artifacts, like dye blobs, were also monitored. Stutter bands originate from Taq-polymerase-dependent PCR errors (slipped strand mispairing, cf. [19]), and appear as sequences that are 1 bp shorter than the true allele. PlusA peaks result from the addition of non-templated nucleotides (primarily an extra adenosine) at the 3′-end of the amplicon, which is favoured by the non-proofreading activity (3′ to 5′ exonuclease activity) of the Qiagen HotStart Taq polymerase. Tetranucleotides typically show only a single stutter band (around 15% of the main allele height), and a straightforward interpretation of electropherograms. Dinucleotides, on the other hands, can yield more stutters, which may show as high as the true allele. In these cases, any measure to quantify stutters relative to the main allele (for example, stutter per cent) is not applicable. Therefore, we monitored the stutters through direct visual inspection of electrophergrams to verify that locus-specific patterns and correct allele calling were consistent across all samples during validation and databasing. The same monitoring strategy was adopted for plusA peaks and dye blobs.
To define the lowest template amount for detection, DNA serial dilutions for each replicate (10, 5, 2.5, 1, 0.5, 0.25, 0.125, 0.025 and 0.0125 ng) were amplified with the three multiplex reactions in duplicate under standard conditions. In order to assess the reproducibility of results, the tests were repeated by two operators on different days using different ABI Veriti thermal cyclers.
Peak imbalance in heterozygous individuals may be due to competition of alleles with different sizes during amplification (larger alleles are usually underdog) or to primer binding site mutation. To assess heterozygote (im)balance, we calculated the peak height ratio (PHR) as the RFU (relative fluorescent unit) value of the lowest allele divided by the RFU of the highest allele, and the ratio was multiplied by 100 to express PHR as a percentage. According to the SWGDAM recommendations for forensic samples, an acceptable single-locus peak balance of alleles in heterozygotes should be >60%. In any case, the peak height of the lowest allele in a true heterozygote should exceed the minimum detection threshold selected for a validated STR panel (in our case, 150 RFU, see below). Calculations were based on 20 heterozygous genotypes (10 wolves and 10 dogs from databasing, see below) at each locus. Results were then validated using the full set of wolf and dog samples (n = 100) typed for the reference population database.
2.6. Genotype Assignment by Bayesian Analysis
Statistical analyses to assign unknown individuals to their source population (pure wolves or dogs), and to identify admixed genotypes (wolf × dog hybrids and older backcrosses) were conducted following the Bayesian assignment procedure as implemented in STRUCTURE v 2.3.4 [10,15]. Details of the statistical approach are beyond the scope of this paper, but they are fully described in [5]. Here, we applied the same procedure, with only minor changes in the input setting of the software STRUCTURE, due to use of a slightly different STR panel. The modifications consisted of including map distances for linked loci in the input file (1, 1.3, 4, 2.1 Morgans, respectively for CPH16, C20.253, C20.622 and PRK-CD; CanFam2.0, https://www.ncbi.nlm.nih.gov, accessed on 13 December 2021) and selecting the linkage model and correlated allele frequencies in the parameter settings. As illustrated in [5], results from the highest posterior probability approach and the Evanno’s Δk method [20] suggested that three clusters better described the whole dataset, so K = 3 was defined in the parameter set.
Simulations using empirical data from 100 reference wolves and dogs typed with the current marker set were conducted with HYBRIDLAB [21] to generate two classes of admixed genotypes corresponding to recent generation hybrids (F1s, F2s, first and second generation backcrosses, BW1 and BW2 respectively) and introgressed wolves (older backcrosses with minimal dog ancestry). Subsequently, the simulated genotypes were inputted in STRUCTURE to establish the appropriate thresholds of proportional membership (q) to the inferred clusters for single genotypes. Q-threshold values were derived to minimize type I errors (false positives) and type II errors (false negatives) in the assignment of individuals to their parental classes (pure wolves and dogs) and hybrid classes (recently admixed wolf x dog hybrids and older backcrosses) (cf. [5,22]).
3. Results
3.1. Characterization of Electropherograms and Automated Allele Identification
Five replicates (three wolf samples and two dog samples) were used to validate our protocol. The complete spectrum of a genetic profile at 22 STR loci is displayed for a wolf sample (Figure 1).
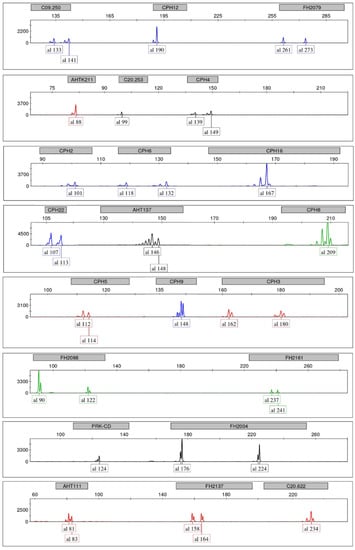
Figure 1.
GeneMapper output showing complete genetic profile at 22 STR loci obtained using the ideal template amount of 5 ng from a wolf muscle sample in the amplification reactions of three multiplexes, and loading 2 µL of five-fold dilutions of the amplicons onto an ABI PRISM 3130 Genetic Analyzer. Allele calls in base pairs are indicated in boxes on the x-axis, while the y-axis shows relative fluorescent units (RFU) at different scales in the different panels. For graphic convenience, markers are not clustered according to their respective amplification multiplex.
Initially, based on the observed single-locus electropherograms in homozygotes, we identified the peaks with the highest RFU values as the true alleles, while the remaining lowest peaks were considered as stutters or plusA bands. Furthermore, we ascertained that the heights of nonspecific peaks both inside and outside the allele ranges (if not already filtered out by range filters, see below) were negligible and that in no case did they interfere with the correct calling of alleles.
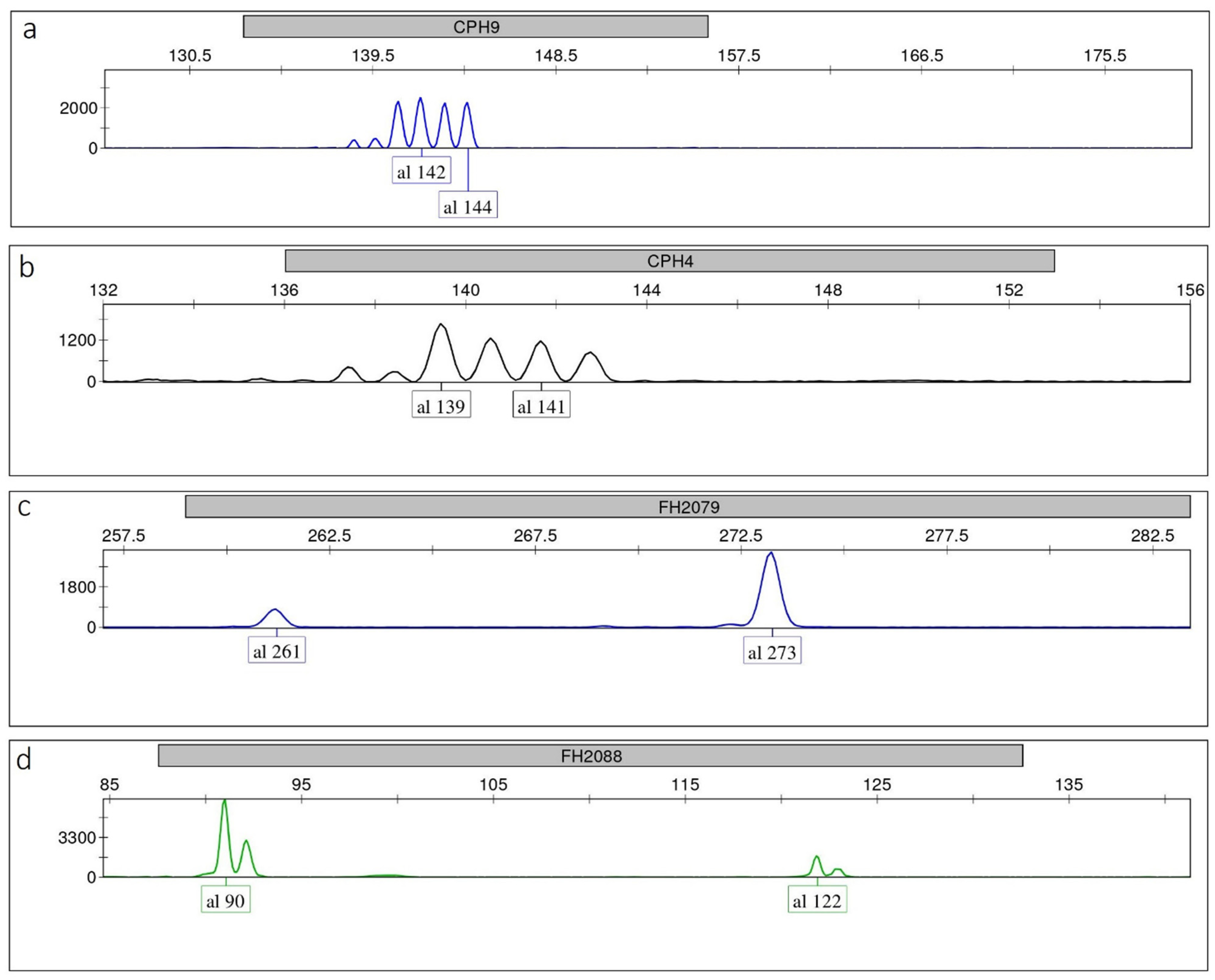
We also observed that backstutters at specific loci (e.g., CPH9) could be as high as the main allele. However, they were always clearly identifiable, even in heterozygotes where alleles differed in size by 2 bp (Figure 2a), which are potentially more prone to misinterpretation. Similarly, plusA bands at some loci (e.g., CPH4, FH2137, Figure 1 and Figure 2b) could reach up to 70–80% of the true allele (even using extension times longer than that indicated in the Qiagen user handbook to promote the adenylate products; data not shown), yet without affecting the correct allele calling in heterozygotes. Dye blobs were never observed in any color within any allele range. Background signals were low across multiplexes and never interfered with signals from amplified STR loci. Stutter and plusA patterns of each allele within each locus and multiplex were consistent and reproducible in five replicates. We observed that this held also true for heterozygotes with alleles differing by 2 bp in size.
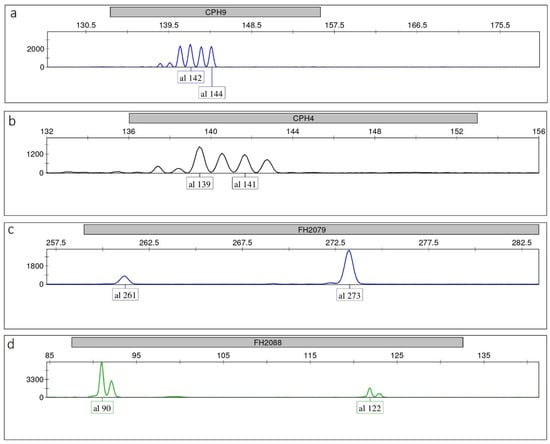
Figure 2.
Single-locus electropherograms in heterozygotes showing: (a) CPH9—alleles differing in size by 2 bp with backstutters that are as high as the true alleles; (b) CPH4—alleles differing in size by 2 bp with plusA bands that reach up to 70–80% of the true alleles; (c) FH2079—severe peak imbalance due to primer binding site mutation in one allele; (d) FH2088—competition of the smallest allele for best amplification.
To optimize parameters for reproducible automated data processing and accurate allele calling with GeneMapper software v 5.0, we made only a few minor changes to the default settings in the “Analysis Methods”. These consisted of increasing the minimum peak half width to 4 data points (to minimize the recognition of nonspecific peaks as real alleles) for M3, while leaving the default value (2) for M1 and M2. Range filters were specified for each color to filter out nonspecific peaks outside the defined allele ranges and avoid any interference with allele identification. Furthermore, a maximum offset of each bin rounded to integer values at ±1 bp (selected because our panel consisted of STRs with tandem repeat units of no less than two nucleotides) proved to be adequate for allele calling across all bin sets. The minimum detection threshold for allele calling was set at 150 RFU (see below).
3.2. Validation
The sensitivity of our 22-locus panel was evaluated by using different amounts of DNA template. Serial dilutions (from 10 to 0.0125 ng) of five replicates were amplified in duplicate under same standard PCR conditions to establish the optimal DNA amount and the template limit of detection (LOD) for each multiplex.
Dilution tests showed that our panel delivered optimal results in the signal range of 6000–500 RFU, when 5–0.125 ng of genomic DNA is applied in PCR reaction volume of 12 µL. Using these amounts, no off-scale peaks occur, artifacts (pull-up, off-scale peaks, noise) are negligible and do not affect the interpretation of electropherograms. The lowest amount of DNA that yielded complete and reliable profiles was LOD = 0.125 ng for all three multiplexes. Comparable LOD values and correct allele calls (all RFUs > 150) were obtained from the five replicates amplified in duplicate, with 100% repeatability. Same results were obtained when analyses were performed by two operators in different days on different thermal cyclers, thus yielding 100% reproducibility of the test. With DNA amounts lower than the LOD value, we observed stochastic fluctuations for several markers, with incorrect allele calling and genotype misidentification experienced by at least one multiplex. Furthermore, allelic dropout (loss of at least one allele in heterozygotes), amplification failure in homozygotes (no signals), alleles fallen below the minimum detection threshold of 150 RFU (see below) were recorded across all multiplexes. Loading 2 µL of non-diluted PCR products (instead of five-fold diluted amplicons) onto the 3130 Genetic Analyzer did not produce better results.
Once the LOD value has been established, we defined the minimum detection threshold as follows: we selected CPH3 as the locus that systematically showed the lowest height across all loci, then we calculated the mean RFU value and standard deviation (SD) of the lowest alleles at this locus across five replicates (mean = 395, SD = 76). We determined that the RFU value of a true allele should reach at least the mean value decreased by three times the SD (395 − 228 = 167) to attain approximately 99 per cent confidence level. Finally, RFU = 150 was selected as the minimum cautionary detection threshold for our panel, meaning that peaks with RFU values lower than 150, obtained under standard conditions, are indicative of possible unreliable genetic profiles.
Twenty heterozygotes were used in the peak height ratio study to calculate PHR values at single loci (Table 2). PHRs (×100) (excluding loci FH2088 and FH2079, see below) averaged between 61.56 ± 6.75 to 92.88 ± 4.55. When considering the mean values of PHR decreased by three times the SD as the lowest PHR range, 13 out of 22 loci (59%) reached PHR values ≥ 60% with 99 per cent confidence. It should be emphasized, however, that this level of peak balance in heterozygotes is highly stringent, and indeed it is suggested in forensic DNA typing. When 95 per cent confidence level is considered (i.e., mean PHRs decreased by two times the SD), three additional loci reached PHR values ≥ 60%.
Table 2.
Peak height ratios (PHR) multiplied by 100, based on 20 heterozygotes analyzed for each locus. Lower PHR ranges are assessed as the mean decrease by three (and two) times the standard deviation (SD). NE = negative value. Values in parentheses were obtained by excluding from calculations the alleles that caused strong peak imbalance (see text for details).
Four markers (CPH6, ATH111, ATH137, FH2004) showed PHRs with 95% confidence level = 53.26, 52.65, 48.06, 47.78%, respectively, therefore never exceeding the value of 60%. However, as the minimum detection threshold of 150 RFU was attained by all alleles and very low backgrounds were produced for each dye in all three multiplexes, correct peak sizing and allele callings in heterozygous genotypes were never compromised. Markers FH2088 and FH2079 exhibited negative values of mean PHR–3SD (mean PHR–2SD as well), due to severe peak imbalance in some samples, where specific alleles clearly amplified more efficiently than others in the heterozygotes. This is possibly explained by primer binding site mutation (e.g., alleles 261 at FH2079, Figure 2c), and by competition of the smallest allele (e.g., allele 90 at FH2088, Figure 2d). When these alleles were excluded from the calculations, the mean values of PHR–3SD highly exceeded 60% at both loci (68% and 80%, respectively).
3.3. Application to Reference Samples
To verify that our validated protocol and automated data processing were appropriate to process field samples, we analyzed 50 Apennine wolves and 50 dogs for databasing, using 5 ng template DNA under standardized conditions. Reference samples were also used to define complete bins for each marker. Results showed that consistency of stutter and plusA patterns were always obtained at single loci across all multiplexes in the field samples. However, some deviations from validation samples were observed.
Signal intensity of 35 alleles at eight loci dropped below the pre-set minimum detection threshold of 150 RFU in five samples, while amplification failure was observed in four samples at three loci (due presumably to primer binding site mutation). However, in all instances where alleles fell below the threshold, peak signals were still significantly higher than the background, so that in no case were the genotypes lost.
Thirty-six out of 2200 single-locus genotypes (1.6%) were left unassigned by automated allele calling due to peaks exceeding the ±1-bp range variation of bins. After visual inspection, however, peaks were easily identified from the background as real alleles and the genotypes were all recovered. Thirty-eight single alleles (0.86%) were left undetermined, and nine complete genotypes at single loci (0.41%) proved definitively inconclusive by both automated and manual allele calling, due to difficult peak interpretation of the electropherograms.
During databasing we observed that marker FH2161, mapping on chromosome 21, showed a tri-allelic pattern (three alleles with same RFU values) in a French Bulldog, suggesting that the dog might suffer from some inherited chromosome disorder [23]. Furthermore, dogs showed various microvariants (i.e., clearly resolved alleles which differed by 2 bp instead of 4) at three tetranucleotide loci (FH2161, FH2137, FH2004), indicating broken repeats.
Based on our 22-locus validated panel, a total of 202 allele variants were found in the entire population dataset. Only 33% of the alleles were shared between wolves and dogs, while 14% and 53% were private to the wolf and the dog, respectively. Allele frequencies in the Apennine wolf and dog populations from our database are available in Table S1 (Supplementary Materials).
3.4. Bayesian Analysis
Simulations were conducted with HYBRIDLAB on empirical data from 50 wolf samples and 50 dog samples typed with our validated protocol. Results allowed the establishment of a q-value threshold of 0.01 to uniquely assign multilocus genotypes to the wolf and dog genetic clusters, and to identify admixed wild genotypes with different levels of domestic ancestry, according to the following categories: (a) q-value threshold > 0.990 for assignment of individuals to the pure wolf cluster (or, alternatively, to the dog cluster); (b) q-value threshold between 0.960 and 0.990 for assignment of older backcrosses (introgressed wolves with old domestic ancestry); (c) q-value threshold < 0.960 for assignment of individuals as admixed wolf × dog hybrids. According to the derived q-values, all wolves were probabilistically assigned to one cluster with a mean Q = 0.990 (CI 0.989–1.00). Similarly, all dogs fell into an alternative cluster with a mean Q = 0.995 (CI 0.991–1.00).
When typing 10 known hybrids, the results (Figure S1, Supplementary Materials) correctly showed that none of them were assigned to the pure wolf or dog cluster (q-value threshold < 0.990 in both clusters). Three of them were classified as recently admixed hybrids (q-value threshold < 0.960), while seven were classified as later backcrosses (0.960 ≤ q-value ≤ 0.990).
4. Discussion
The evaluation of the level of anthropogenic hybridization with the dog is increasingly relevant in the conservation and management of the wolf in Europe. This is especially true in the light of the conflicting ideas and motivations of various stakeholders (scientists, conservationists, hunters, livestock farmers, wildlife managers, practitioners, politicians/decision makers) [24,25,26].
To provide a molecular tool to help addressing the issue, we optimized and validated a protocol in multiplexed PCR format based on 22 STR canine loci, which is capable of distinguishing genetically pure wolves from dogs, and statistically assigning admixed individuals to either recent hybrids or hybrids with older dog ancestry, when used in association with a Bayesian assignment procedure.
In order to generate reliable genetic profiles with high diagnostic power, dinucleotide and tetranucleotide markers were selected from the literature based on their inter-taxon polymorphism on one hand, and clear resolution of specific bands, peak detection and fragment sizing on the other. The validation process of the molecular protocol presented here involved sensitivity, repeatability and reproducibility of data in compliance with the SWGDAM [18] and SWFS [27] recommendations and guidelines for animal and wildlife forensic analyses.
Our optimized assay showed good sensitivity and minimizes sample requirement for complete genotyping, using as low as 0.125 ng of DNA in each of three multiplexes of 12 µL PCR reactions. One-hundred per cent repeatability and reproducibility was obtained when tests were run in replicates by different operators in separated sessions, using different thermal cyclers. Furthermore, the performance of PHR values reached highly conservative levels of acceptability, which are proper to forensic testing.
From a reference sample set of 100 wolves and dogs typed with the validated protocol, we verified that consistency of peak patterns in the electropherograms occurred across loci, and defined complete bins for each marker to enable automated identification of single alleles. Changes from the default parameter setting in GeneMapper for automatic allele calling procedure showed appropriate to our STR panel, and only a very low number of missing data at single loci in the whole population dataset (1.6%) needed to be manually identified and recovered after unsuccessful automated data processing. In these cases, as also suggested for human forensic DNA typing [28], we suggest that final interpretation of ambiguous profiles should always be sifted by the scientist’s professional experience and judgement. Eventually, the validated 22-locus panel and data processing yielded few inconclusive genotypes (0.41%) in the whole dataset, where 91 out of 100 reference profiles showed no missing data at single loci.
The application of our STR panel to a Bayesian assignment approach proved to be adequate in order to unambiguously define two genetically separated clusters for wolves and dogs, and to successfully identify 10 known hybrids as admixed individuals. The latter were assigned to their class of admixture according to individual q-values of membership, and were classified as recent hybrids and older backcrosses.
Our approach has proven to be an excellent starting point for a cost-effective, reliable and reproducible process to diagnose pure and admixed wolf genotypes. We are also confident, however, that our protocol would be amenable to resolution improvement through the incorporation of additional selected markers with only minor optimization into the existing 22-locus assay format and analysis method. More importantly, the protocol can be adopted by various laboratories that need this kind of diagnosis, and inter-laboratory comparisons to calibrate allele sizes would produce concordant results, even in the absence of allelic ladders, through the organization of ring tests with wolf and dog control DNAs, scheduled over time.
Although future applications are likely to involve panels of highly informative markers at the genomic level, such as SNPs (Single Nucleotide Polymorphisms) through NGS (Next Generation Sequencing) technology [29], STRs are currently the most widely used markers in animal DNA typing, especially for use with opportunistically-collected samples, where the DNA is often scarce and degraded.
If no less than 0.125 ng template DNA is applied, as LOD value suggested from this study, our protocol is compatible with genomic DNA from various types of biological samples, such as blood and saliva from buccal swabs. With allele size ranges not exceeding 300 base pairs in length, the presented STR-based method is suitable for amplification of degraded and low quality DNA samples. In addition, after internal laboratory validation to estimate genotyping errors (i.e., allelic drop-out and false alleles), the protocol can be applied to non-invasively collected samples, such as feces, shed hairs, wolf saliva from wounds on preys, and forensic evidence.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d14070511/s1, Table S1: Allele frequencies in Apennine wolves and dogs. N = allele number, Figure S1: Bar plotting of the results from a Bayesian analysis conducted with STRUCTURE. Each vertical bar represents one individual and the length of the different colored sections is proportional to its membership (q-values) in the three inferred genetic clusters. Wolf component is colored green. Hybrids: recently admixed individuals; old backcrosses: introgressed wolves with minimal domestic ancestry.
Author Contributions
Conceptualization, R.L. and L.G.; laboratory procedures and validation, C.T., R.F. and L.A.; writing—original draft preparation, R.L.; writing—review and editing, L.G. and L.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Istituto Zooprofilattico Sperimentale delle Regioni Lazio e Toscana “M. Aleandri”.
Institutional Review Board Statement
No ethics permit in accordance with the EU Directives was required for this study, as no live animals were involved, nor was there any transboundary passage of samples from protected species. Only samples from individuals that died of natural causes, accidentally or killed illegally, were analyzed for our purposes.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We acknowledge all the people who provided us with wolf and dog samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mech, L.D.; Boitani, L. Wolves: Behavior, Ecology, and Conservation; University of Chicago Press: Chicago, IL, USA, 2007; p. 472. [Google Scholar]
- Donfrancesco, V.; Ciucci, P.; Salvatori, V.; Benson, D.; Andersen, L.W.; Bassi, E.; Blanco, J.C.; Boitani, L.; Caniglia, R.; Canu, A.; et al. Unravelling the scientific debate on how to address wolf-dog hybridization in Europe. Front. Ecol. Evol. 2019, 7, 175. [Google Scholar] [CrossRef] [Green Version]
- Skoglund, P.; Götherström, A.; Jakobsson, M. Estimation of population divergence times from non-overlapping genomic sequences: Examples from dogs and wolves. Mol. Biol. Evol. 2011, 28, 1505–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilà, C.; Wayne, R.K. Hybridization between wolves and dogs. Conserv. Biol. 1999, 13, 195–198. [Google Scholar] [CrossRef]
- Lorenzini, R.; Fanelli, R.; Grifoni, G.; Sholl, F.; Fico, R. Wolf-dog crossbreeding: “Smelling” a hybrid may not be easy. Mamm. Biol. 2014, 79, 149–156. [Google Scholar] [CrossRef]
- Amorim, A.; Pereira, F.; Alves, C.; García, O. Species assignment in forensics and the challenge of hybrids. Forensic Sci. Int. Genet. 2020, 48, 102333. [Google Scholar] [CrossRef]
- Galov, A.; Fabbri, E.; Caniglia, R.; Arbanasić, H.; Lapalombella, S.; Florijančić, T.; Bošković, I.; Galaverni, M.; Randi, E. First evidence of hybridization between golden jackal (Canis aureus) and domestic dog (Canis familiaris) as revealed by genetic markers. R. Soc. Open Sci. 2015, 2, 150450. [Google Scholar] [CrossRef] [Green Version]
- Dziech, A. Identification of wolf-dog hybrids in Europe—An overview of genetic studies. Front. Ecol. Evol. 2021, 9, 760160. [Google Scholar] [CrossRef]
- Lorenzini, R.; Garofalo, L. Wildlife Forensics. In Forensic Genetics: New Technology and Applications; Pilli, E., Berti, A., Eds.; Apple Academic Press: Palm Bay, FL, USA, 2021; pp. 357–384. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Richman, M.; Mellersh, C.S.; André, C.; Galibert, F.; Ostrander, E.A. Characterization of a minimal screening set of 172 microsatellite markers for genome-wide screens of the canine genome. J. Biochem. Biophys. Methods 2001, 47, 137–149. [Google Scholar] [CrossRef]
- Randi, E.; Lucchini, V. Detecting rare introgression of domestic dog genes into wild wolf (Canis lupus) populations by Bayesian admixture analyses of microsatellite variation. Conserv. Genet. 2002, 3, 31–45. [Google Scholar] [CrossRef]
- Verardi, A.V.; Lucchini, E.; Randi, E. Detecting introgressive hybridization between free-ranging domestic dogs and wild wolves (Canis lupus) by admixture linkage disequilibrium analysis. Mol. Ecol. 2006, 15, 2845–2855. [Google Scholar] [CrossRef] [PubMed]
- Godinho, R.; Llaneza, L.; Blanco, J.C.; Lopes, S.; Álvares, F.; García, E.J.; Palacios, V.; Cortés, Y.; Talegòn, J.; Ferrand, N. Genetic evidence for multiple events of hybridization between wolves and domestic dogs in the Iberian Peninsula. Mol. Ecol. 2011, 20, 5154–5166. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef] [PubMed]
- Ciucci, P.; Lucchini, V.; Boitani, L.; Randi, E. Dewclaws in wolves as evidence of admixed ancestry with dogs. Can. J. Zool. 2003, 81, 2077–2081. [Google Scholar] [CrossRef]
- Caniglia, R.; Fabbri, E.; Greco, C.; Galaverni, M.; Manghi, L.; Boitani, L.; Sforzi, A.; Randi, E. Black coats in an admixed wolf × dog pack is melanism an indicator of hybridization in wolves? Eur. J. Wildl. Res. 2013, 59, 543–555. [Google Scholar] [CrossRef]
- Scientific Working Group on DNA Analysis Methods (SWGDAM). Validation Guidelines for Forensic DNA Analysis Methods. 2016. Available online: https://www.swgdam.org/publications (accessed on 20 April 2022).
- Goldstein, D.B.; Schlötterer, C. Microsatellites: Evolution and Applications; Oxford University Press: Oxford, UK, 1999; p. 368. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, E.E.; Bach, L.A.; Kotlicki, P. Hybridlab (version 1.0): A program for generating simulated hybrids from population samples. Mol. Ecol. Notes 2006, 6, 972–973. [Google Scholar] [CrossRef]
- Caniglia, R.; Galaverni, M.; Velli, E.; Mattucci, F.; Canu, A.; Apollonio, M.; Mucci, N.; Scandura, M.; Fabbri, E. A standardized approach to empirically define reliable assignment thresholds and appropriate management categories in deeply introgressed populations. Sci. Rep. 2020, 10, 2862. [Google Scholar] [CrossRef] [Green Version]
- Szczerbal, I.; Switonski, M. Clinical cytogenetics of the dog: A Review. Animals 2021, 11, 947. [Google Scholar] [CrossRef]
- Musiani, M.; Boitani, L.; Paquet, P.C. The World of Wolves: New Perspectives on Ecology, Behaviour, and Management; University of Calgary Press: Calgary, AB, Canada, 2010; p. 399. [Google Scholar] [CrossRef]
- Mech, L.D. Where can wolves live and how can we live with them? Biol. Conserv. 2017, 210, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.; Chamaillé-Jammes, S.; Waller, D.M. Deer, wolves, and people: Costs, benefits and challenges of living together. Biol. Rev. 2020, 95, 782–801. [Google Scholar] [CrossRef] [PubMed]
- SWFS Technical Working Group. Standards and Guidelines for Wildlife Forensic Analysis, Version 3; Webster, L.M.I., Ed.; Society for Wildlife Forensic Science: Ashland, OR, USA, 2018; p. 21. Available online: https://www.wildlifeforensicscience.org/documents/ (accessed on 10 April 2022).
- Holt, C.L.; Buoncristiani, M.; Wallin, J.M.; Nguyen, T.; Lazaruk, K.D.; Walsh, P.S. TWGDAM validation of AmpF STR™ PCR amplification kits for forensic DNA casework. J. Forensic Sci. 2002, 47, 66–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caniglia, R.; Fabbri, E.; Hulva, P.; Bolfíková, B.Č.; Jindřichová, M.; Stronen, A.V.; Dykyy, I.; Camassa, A.; Carnier, P.; Randi, E.; et al. Wolf outside, dog inside? The genomic make-up of the Czechoslovakian Wolfdog. BMC Genom. 2018, 19, 533. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).