
Phylogeographic and Bioclimatic Determinants of the Dorsal Pattern Polymorphism in the Italian Wall Lizard, Podarcis siculus


Abstract
:1. Introduction
2. Material and Methods
2.1. Selection of Observations
2.2. Character and Phenotype
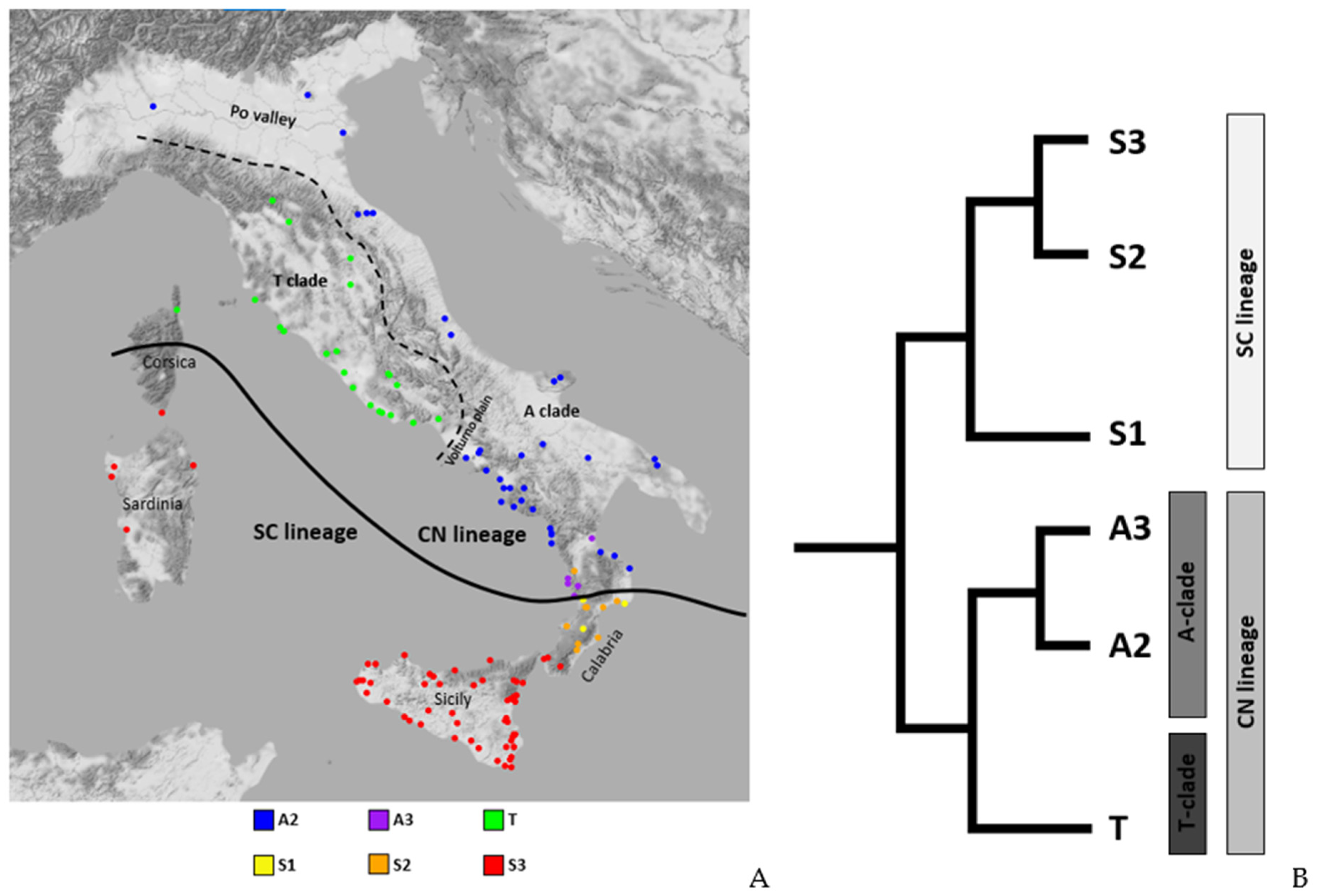
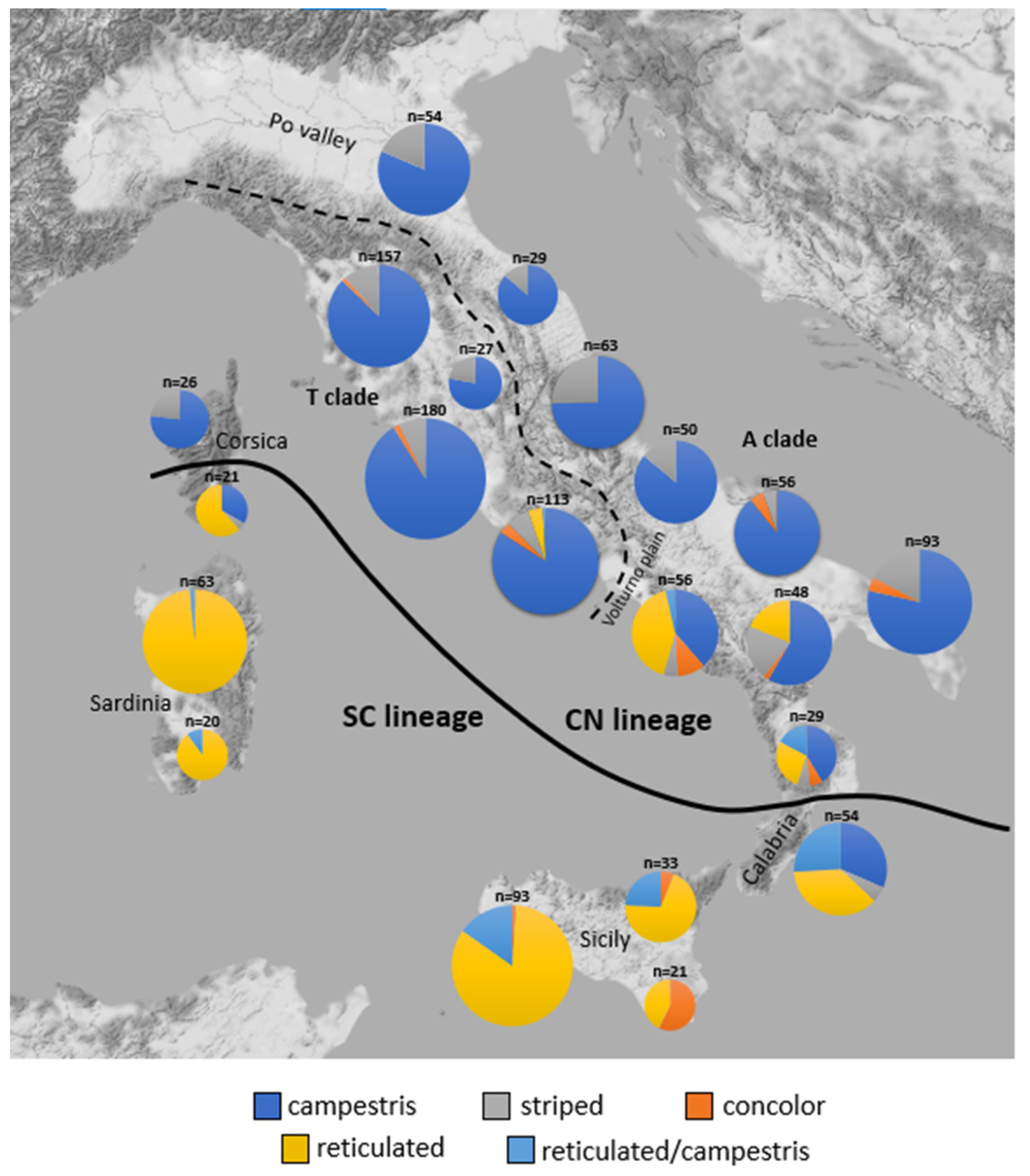
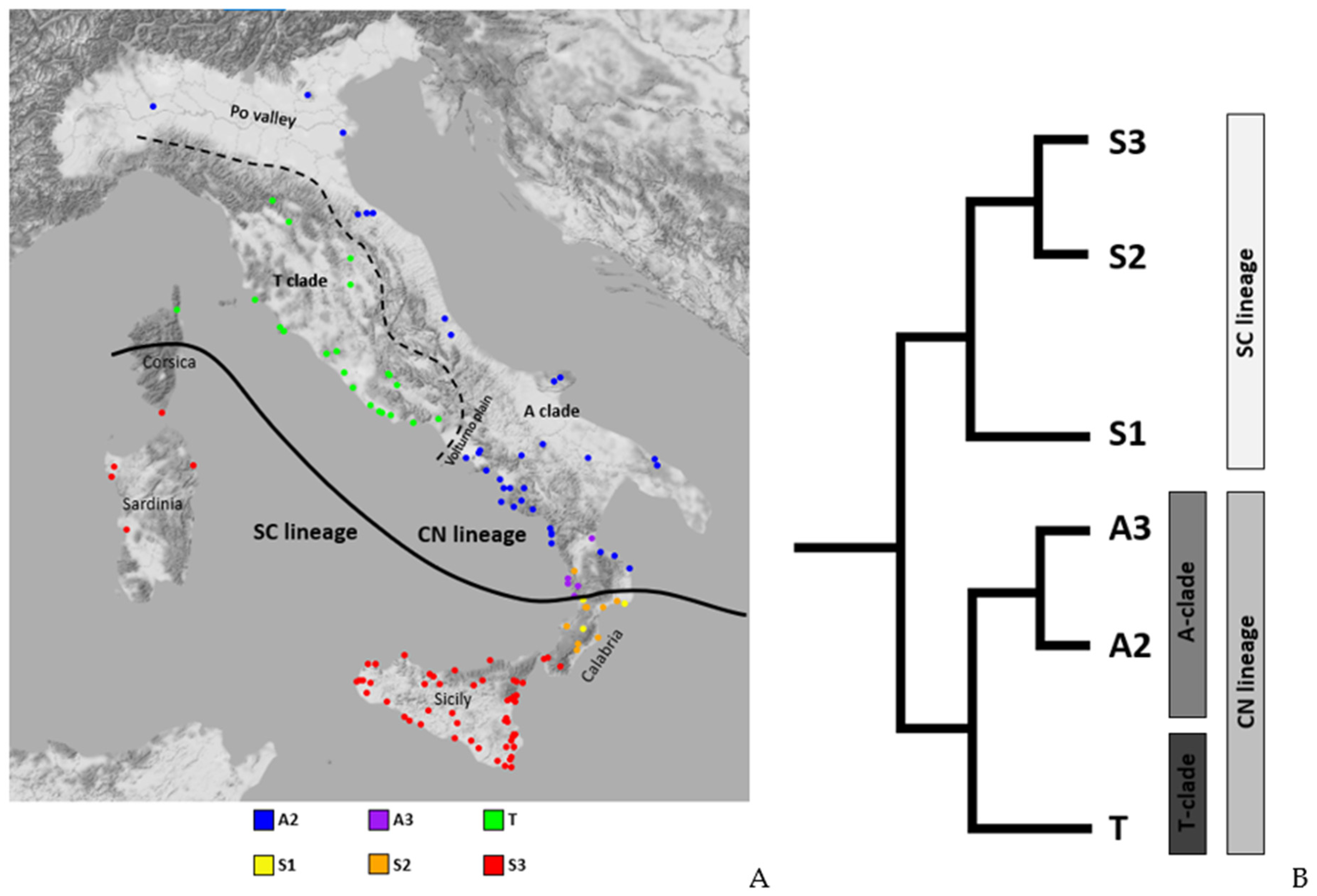
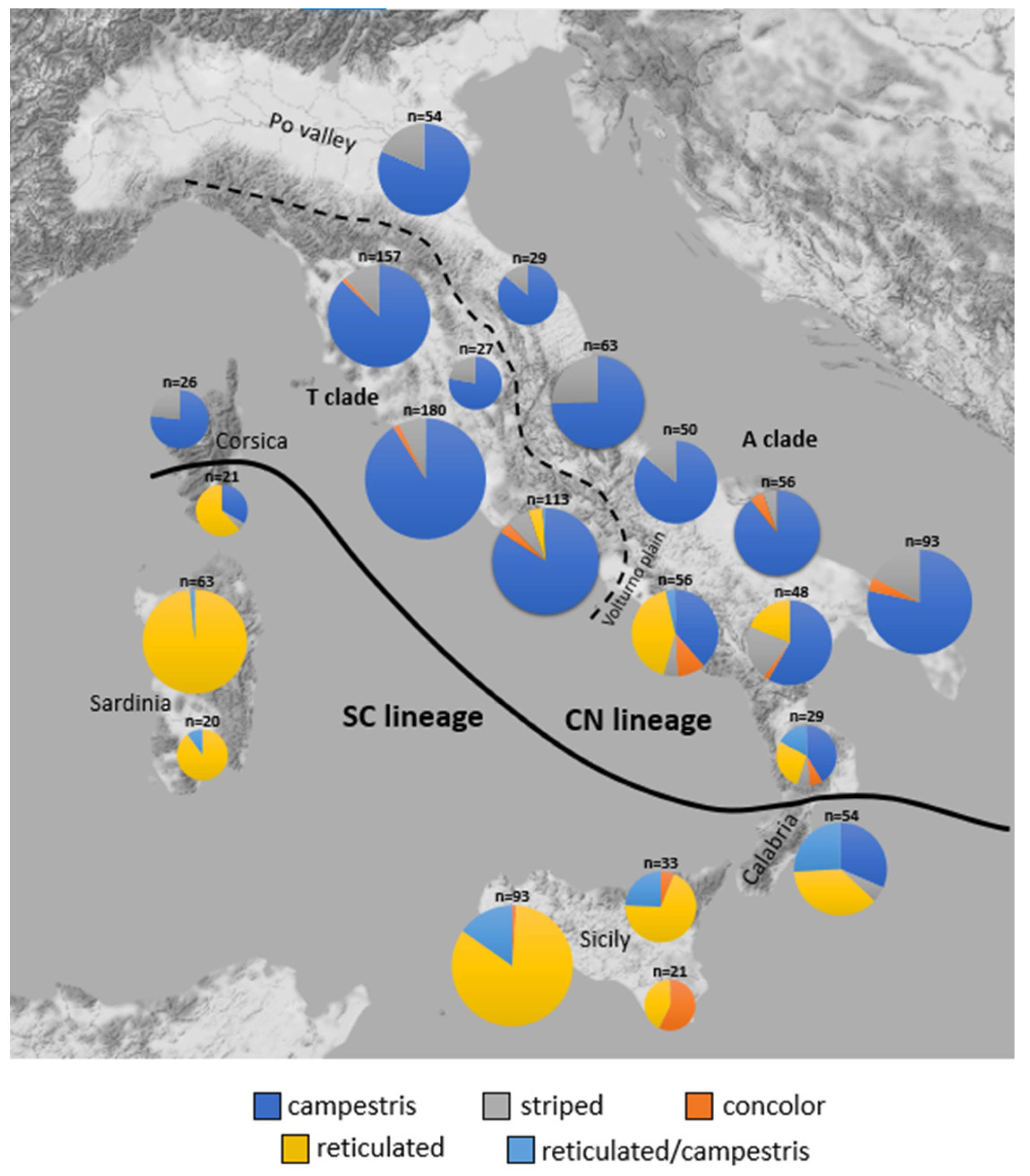
2.3. Distribution of Dorsal Pattern Phenotypes and mtDNA Lineages
2.4. GLM Modelling
3. Results
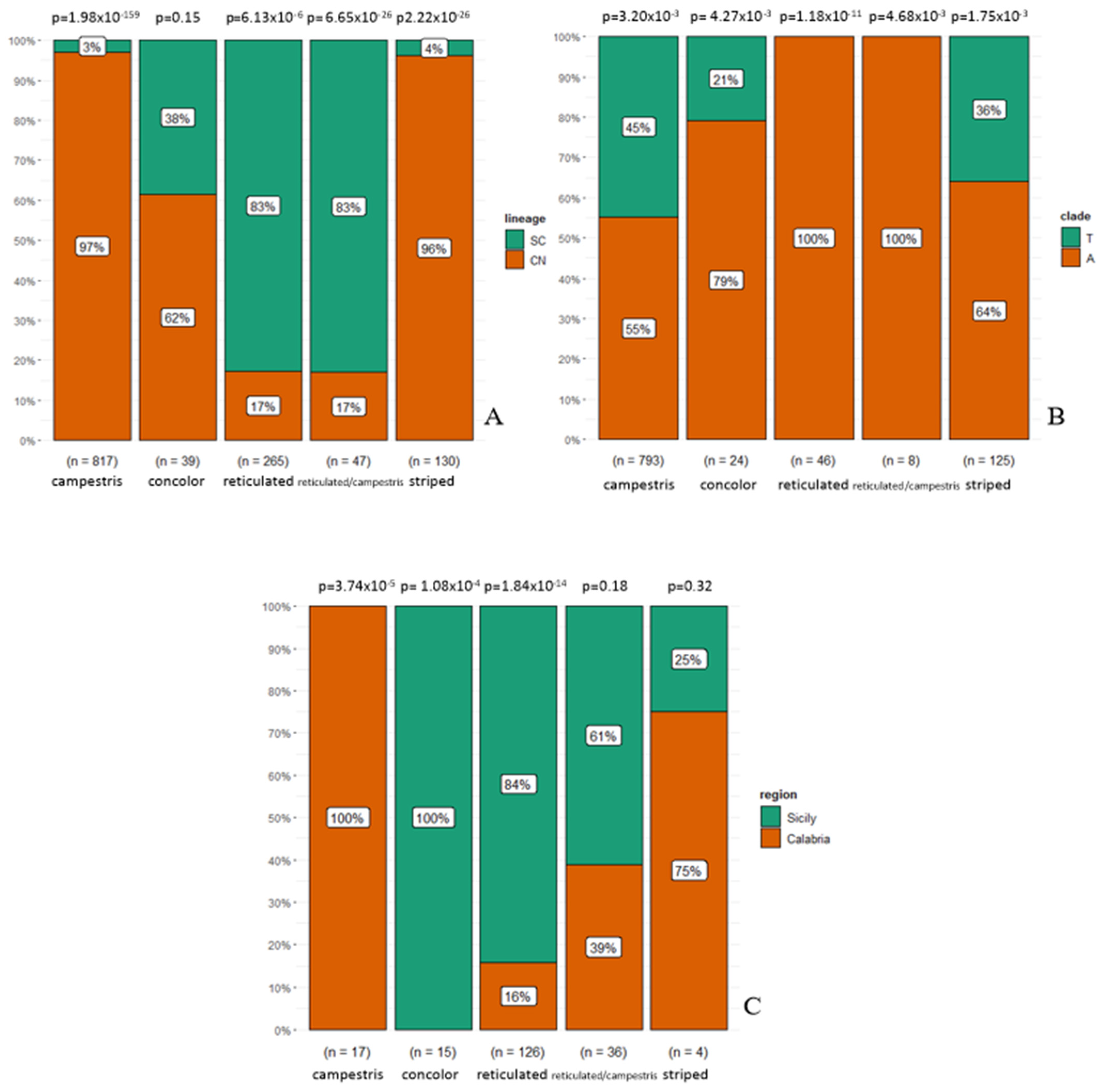
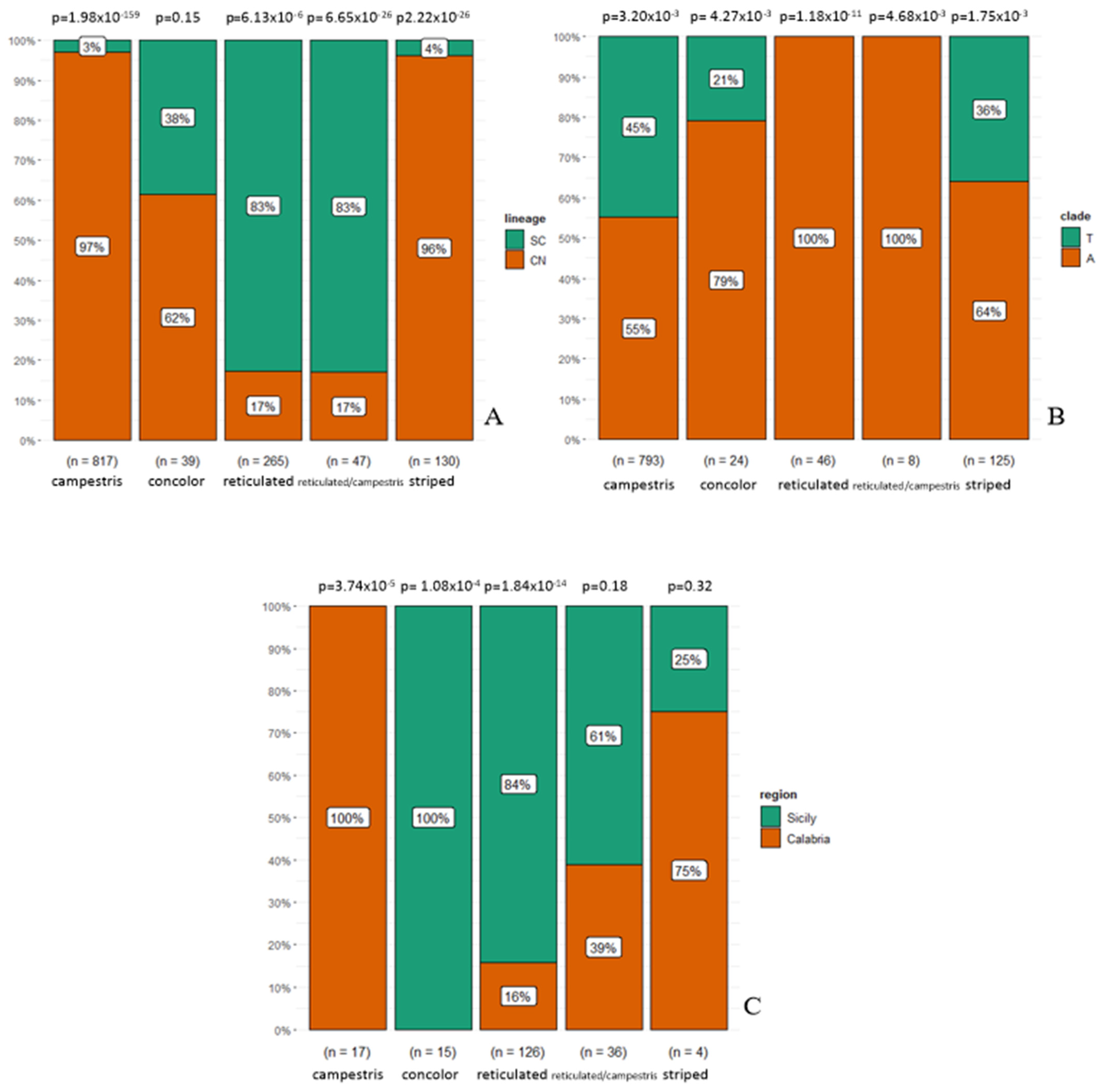
3.1. Phenotype Geographical Distribution and χ2 Test
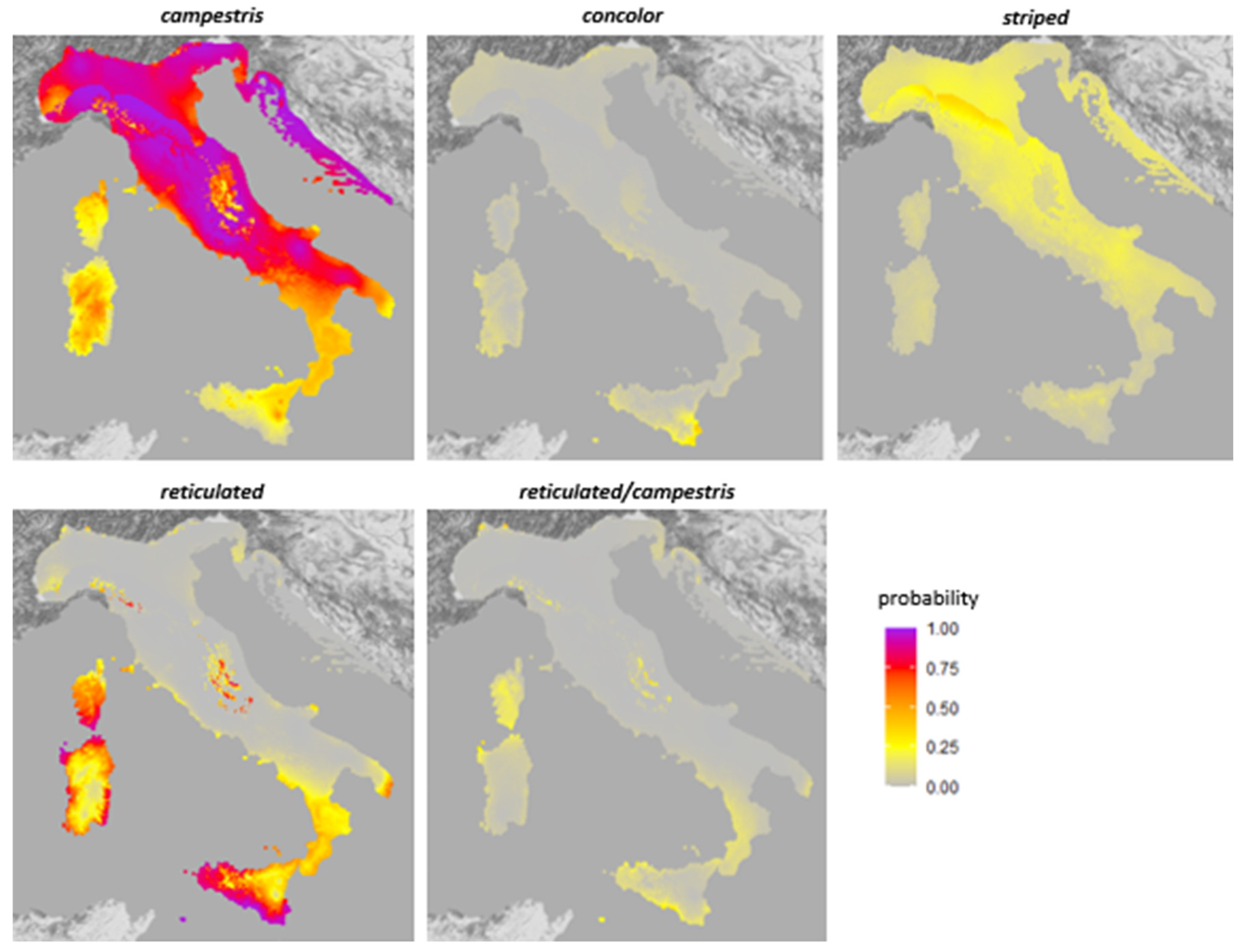
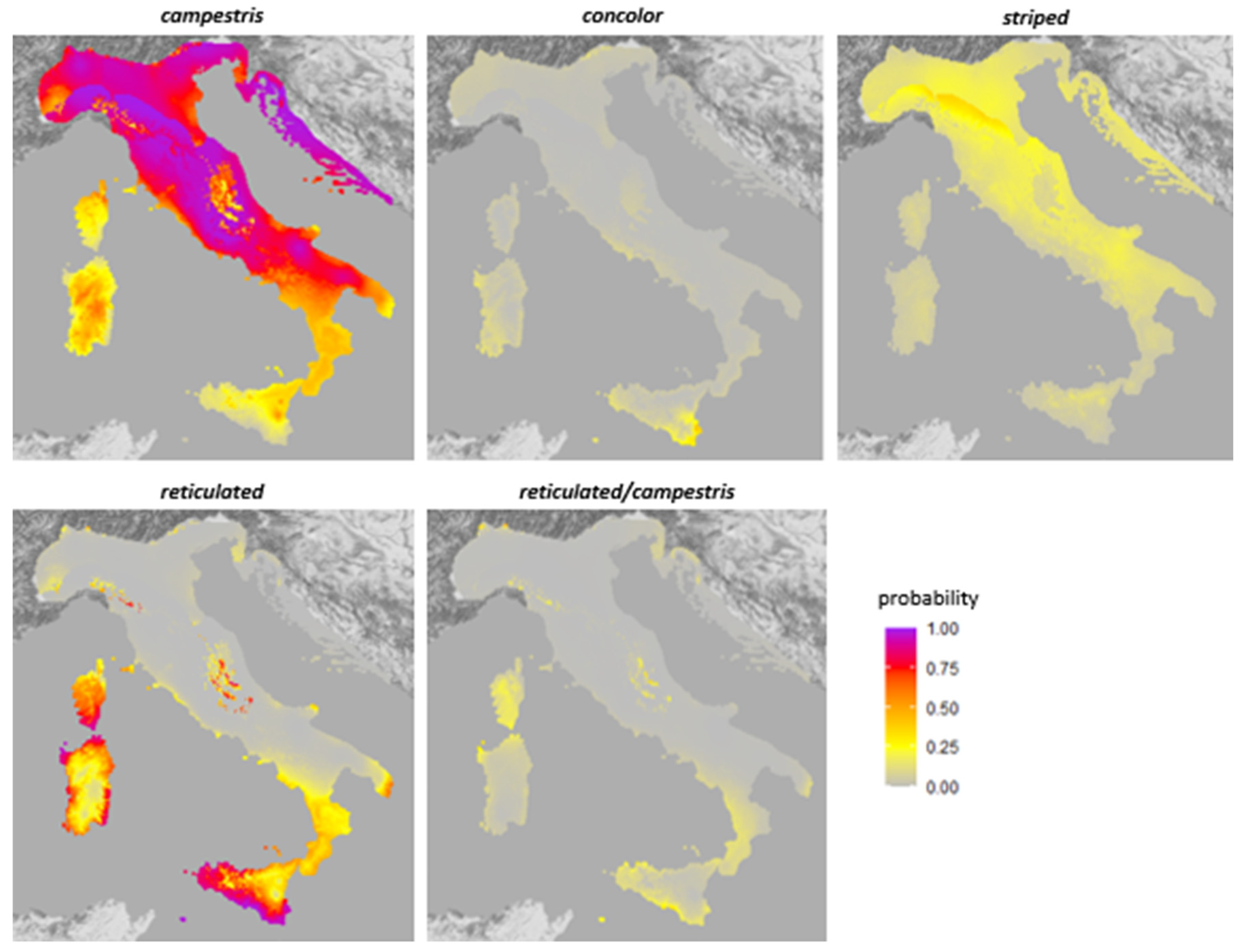
3.2. Generalized Linear Models
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Camargo, A.; Sinervo, B.; Sites, J.W., Jr. Lizards as model organisms for linking phylogeographic and speciation studies. Mol. Ecol. 2010, 19, 3250–3270. [Google Scholar] [CrossRef] [PubMed]
- Zamudio, K.R.; Bell, R.C.; Mason, N.A. Phenotypes in phylogeography: Species’ traits, environmental variation, and vertebrate diversification. Proc. Natl. Acad. Sci. USA 2016, 113, 8041–8048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lande, R. Natural selection and random genetic drift in phenotypic evolution. Evolution 1976, 30, 314–334. [Google Scholar] [CrossRef] [PubMed]
- Barton, N.H.; Mallet, J. Natural selection and random genetic drift as causes of evolution on islands. Philos. Trans. R. Soc. B Biol. Sci. 1996, 351, 785–795. [Google Scholar]
- Marshall, K.L.A.; Philpot, K.E.; Damas-Moreira, I.; Stevens, M. Intraspecific colour variation among lizards in distinct island environments enhances local camouflage. PLoS ONE 2015, 10, e0135241. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.R.; Cadena, V.; Endler, J.A.; Kearney, M.R.; Porter, W.P.; Stuart-Fox, D. Color Change for Thermoregulation versus Camouflage in Free-Ranging Lizards. Am. Nat. 2016, 188, 668–678. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Rueda, G.; González-Granda, L.G.; Reguera, S.; Zamora-Camacho, F.J.; Melero, E. Crypsis Decreases with Elevation in a Lizard. Diversity 2019, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Ortega, J.; López, P.; Martín, J. Conspicuous blue tails, dorsal pattern morphs and escape behaviour in hatchling Iberian wall lizards (Podarcis hispanicus). Biol. J. Linn. Soc. 2014, 113, 1094–1106. [Google Scholar] [CrossRef] [Green Version]
- Ortega, J.; Pellitteri-Rosa, D.; López, P.; Martín, J. Dorsal pattern polymorphism in female Iberian wall lizards: Differences in morphology, dorsal coloration, immune response, and reproductive investment. Biol. J. Linn. Soc. 2015, 116, 352–363. [Google Scholar] [CrossRef] [Green Version]
- Pérez i de Lanuza, G.; Font, E.; Carazo, P. Color-assortative mating in a color-polymorphic lacertid lizard. Behav. Ecol. 2013, 24, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Eitzel, M.V.; Cappadonna, J.L.; Santos-Lang, C.; Duerr, R.E.; Virapongse, A.; West, S.E.; Kyba, C.C.M.; Bowser, A.; Cooper, C.B.; Sforzi, A.; et al. Citizen Science Terminology Matters: Exploring Key Terms. Citiz. Sci. Theory Pract. 2017, 2, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Crall, A.W.; Jordan, R.; Holfelder, K.; Newman, G.J.; Graham, J.; Waller, D.M. The impacts of an invasive species citizen science training program on participant attitudes, behavior, and science literacy. Public Underst. Sci. 2012, 22, 745–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tulloch, A.I.; Possingham, H.P.; Joseph, L.N.; Szabo, J.; Martin, T.G. Realising the full potential of citizen science monitoring programs. Biol. Conserv. 2013, 165, 128–138. [Google Scholar] [CrossRef] [Green Version]
- Simoniello, C.; Jencks, J.; Lauro, F.M.; Loftis, J.D.; Weslawski, J.M.; Deja, K.; Forrest, D.R.; Gossett, S.; Jeffries, T.; Jensen, R.M.; et al. Citizen-Science for the Future: Advisory Case Studies from Around the Globe. Front. Mar. Sci. 2019, 6, 225. [Google Scholar] [CrossRef] [Green Version]
- Devictor, V.; Whittaker, R.J.; Beltrame, C. Beyond scarcity: Citizen science programmes as useful tools for conservation biogeography. Divers. Distrib. 2010, 16, 354–362. [Google Scholar] [CrossRef]
- Hurlbert, A.H.; Liang, Z. Spatiotemporal Variation in Avian Migration Phenology: Citizen Science Reveals Effects of Climate Change. PLoS ONE 2012, 7, e31662. [Google Scholar] [CrossRef] [Green Version]
- Putman, B.J.; Williams, R.; Li, E.; Pauly, G.B. The power of community science to quantify ecological interactions in cities. Sci. Rep. 2021, 11, 1–8. [Google Scholar] [CrossRef]
- Silvertown, J.; Cook, L.; Cameron, R.; Dodd, M.; McConway, K.; Worthington, J.; Skelton, P.; Anton, C.; Bossdorf, O.; Baur, B.; et al. Citizen Science Reveals Unexpected Continental-Scale Evolutionary Change in a Model Organism. PLoS ONE 2011, 6, e18927. [Google Scholar] [CrossRef] [Green Version]
- Kerstes, N.A.; Breeschoten, T.; Kalkman, V.J.; Schilthuizen, M. Snail shell colour evolution in urban heat islands detected via citizen science. Commun. Biol. 2019, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Drury, J.P.; Barnes, M.; Finneran, A.E.; Harris, M.; Grether, G.F. Continent-scale phenotype mapping using citizen scientists’ photographs. Ecography 2019, 42, 1436–1445. [Google Scholar] [CrossRef] [Green Version]
- Lehtinen, R.M.; Carlson, B.M.; Hamm, A.R.; Riley, A.G.; Mullin, M.M.; Gray, W.J. Dispatches from the neighborhood watch: Using citizen science and field survey data to document color morph frequency in space and time. Ecol. Evol. 2020, 10, 1526–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, E.N.; Burton, J.A.; Ovenden, D. Field Guide to the Reptiles and Amphibians of Britain and Europe; Collins: New York, NY, USA, 1978. [Google Scholar]
- Corti, C.; Lo Cascio, P. Lacertidi Italiani; L’epos: Milan, Italy, 1999. [Google Scholar]
- Lepetz, V.; Massot, M.; Chaine, A.S.; Clobert, J. Climate warming and the evolution of morphotypes in a reptile. Glob. Chang. Biol. 2009, 15, 454–466. [Google Scholar] [CrossRef]
- Sherbrooke, W.C.; Castrucci, A.M.D.L.; Hadley, M.E. Temperature Effects on in vitro Skin Darkening in the Mountain Spiny Lizard, Sceloporus jarrovi: A Thermoregulatory Adaptation? Physiol. Zool. 1994, 67, 659–672. [Google Scholar] [CrossRef]
- Gabirot, M.; Balleri, A.; López, P.; José, M. Differences in Thermal Biology Between Two Morphologically Distinct Populations of Iberian Wall Lizards Inhabiting Different Environments. In Annales Zoologici Fennici; Finnish Zoological and Botanical Publishing Board: Helsinki, Finland, 2013; Volume 50, pp. 225–236. [Google Scholar]
- Podnar, M.; Mayer, W.; Tvrtković, N. Phylogeography of the Italian wall lizard, Podarcis sicula, as revealed by mitochondrial DNA sequences. Mol. Ecol. 2005, 14, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Senczuk, G.; Colangelo, P.; De Simone, E.; Aloise, G.; Castiglia, R. A combination of long term fragmentation and glacial persistence drove the evolutionary history of the Italian wall lizard Podarcis siculus. BMC Evol. Biol. 2017, 17, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Senczuk, G.; Havenstein, K.; Milana, V.; Ripa, C.; De Simone, E.; Tiedemann, R.; Castiglia, R. Spotlight on islands: On the origin and diversification of an ancient lineage of the Italian wall lizard Podarcis siculus in the western Pontine Islands. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kahle, D.; Wickham, H. ggmap: Spatial Visualization with ggplot2. R J. 2013, 5, 144–161. [Google Scholar] [CrossRef] [Green Version]
- Senczuk, G.; Harris, D.J.; Castiglia, R.; Mizan, V.L.; Colangelo, P.; Canestrelli, D.; Salvi, D. Evolutionary and demographic correlates of Pleistocene coastline changes in the Sicilian wall lizard Podarcis wagleriana. J. Biogeogr. 2019, 46, 224–237. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; van Etten, J. Raster: Geographic Analysis and Modeling with Raster Data. R Package Version 2.0-12. 2012. Available online: http://cran.r-project.org/web/packages/raster/raster.pdf (accessed on 5 April 2021).
- Avramo, V.; Senczuk, G.; Corti, C.; Böhme, W.; Capula, M.; Castiglia, R.; Colangelo, P. Evaluating the island effect on phenotypic evolution in the Italian wall lizard, Podarcis siculus (Reptilia: Lacertidae). Biol. J. Linn. Soc. 2021, 132, 655–665. [Google Scholar] [CrossRef]
- Yoon, S.; Lee, W.-H. Methodological analysis of bioclimatic variable selection in species distribution modeling with application to agricultural pests (Metcalfa pruinosa and Spodoptera litura). Comput. Electron. Agric. 2021, 190, 106430. [Google Scholar] [CrossRef]
- Xie, C.; Zhang, G.; Jim, C.; Liu, X.; Zhang, P.; Qiu, J.; Liu, D. Bioclimatic Suitability of Actual and Potential Cultivation Areas for Jacaranda mimosifolia in Chinese Cities. Forests 2021, 12, 951. [Google Scholar] [CrossRef]
- Gusmão, A.C.; Evangelista-Vale, J.C.; Pires-Oliveira, J.C.; Barnett, A.A.; da Silva, O.D. New records and modelling the impacts of climate change on the black-tailed marmosets. PLoS ONE 2021, 16, e0256270. [Google Scholar] [CrossRef] [PubMed]
- Janžekovič, F.; Novak, T. PCA—A powerful method for analyze ecological niches. In Principal Component Analysis–Multidisciplinary Applications; Intech Open: Vienna, Austria, 2012; pp. 127–142. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.-C.; Müller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- Speybroeck, J.; Beukema, W.; Dufresnes, C.; Fritz, U.; Jablonski, D.; Lymberakis, P.; Martínez-Solano, I.; Razzetti, E.; Vamberger, M.; Vences, M.; et al. Species list of the European herpetofauna–2020 update by the Taxonomic Committee of the Societas Europaea Herpetologica. Amphibia-Reptilia 2020, 41, 139–189. [Google Scholar] [CrossRef]
- Pellitteri-Rosa, D.; Gazzola, A.; Todisco, S.; Mastropasqua, F.; Liuzzi, C. Lizard colour plasticity tracks background seasonal changes. Biol. Open 2020, 9, bio052415. [Google Scholar] [CrossRef]
- Storniolo, F.; Zuffi, M.A.L.; Coladonato, A.J.; Di Vozzo, L.; Giglio, G.; Gini, A.E.; Leonetti, F.L.; Luccini, S.; Mangiacotti, M.; Scali, S.; et al. Patterns of variations in dorsal colouration of the Italian wall lizard Podarcis siculus. Biol. Open 2021, 10, bio058793. [Google Scholar] [CrossRef]
- Romano, P.; Santo, A.; Voltaggio, M. Evoluzione geomorfologica della piana del fiume Volturno (Campania) durante il tardo Quaternario (Pleistocene medio-superiore—Olocene). Il Quat. 1994, 7, 41–56. [Google Scholar]
- Barra, D.; Romano, P.; Santo, A.; Campajola, L.; Roca, V.; Tuniz, C. The versilian transgression in the Volturno River plain (Campania, Italy): Palaeoenvironmental evolution and chronological data. Il Quat. 1996, 9, 445–458. [Google Scholar]
- Canestrelli, D.; Cimmaruta, R.; Nascetti, G. Population genetic structure and diversity of the Apennine endemic stream frog, Rana italica–insights on the Pleistocene evolutionary history of the Italian peninsular biota. Mol. Ecol. 2008, 17, 3856–3872. [Google Scholar] [CrossRef]
- Bisconti, R.; Porretta, D.; Arduino, P.; Nascetti, G.; Canestrelli, D. Hybridization and extensive mitochondrial introgression among fire salamanders in peninsular Italy. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chiocchio, A.; Arntzen, J.; Martinez-Solano, I.; de Vries, W.; Bisconti, R.; Pezzarossa, A.; Maiorano, L.; Canestrelli, D. Reconstructing hotspots of genetic diversity from glacial refugia and subsequent dispersal in Italian common toads (Bufo bufo). Sci. Rep. 2021, 11, 1–14. [Google Scholar]
- Michaelides, S.N.; While, G.M.; Zajac, N.; Uller, T. Widespread primary, but geographically restricted secondary, human introductions of wall lizards, Podarcis muralis. Mol. Ecol. 2015, 24, 2702–2714. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; While, G.M.; Laakkonen, H.; Sacchi, R.; Zuffi, M.A.L.; Scali, S.; Salvi, D.; Uller, T. Genomic evidence for asymmetric introgression by sexual selection in the common wall lizard. Mol. Ecol. 2018, 27, 4213–4224. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Feiner, N.; Laakkonen, H.; Sacchi, R.; Zuffi, M.A.L.; Scali, S.; While, G.; Uller, T. Spatial variation in gene flow across a hybrid zone reveals causes of reproductive isolation and asymmetric introgression in wall lizards*. Evolution 2020, 74, 1289–1300. [Google Scholar] [CrossRef]
- Corti, C.; Capula, M.; Luiselli, L.; Razzetti, E.; Sindaco, R. (Eds.) Fauna d’Italia, Reptilia; Calderini De Il Sole: Bologna, Italy, 2010; Volume XLV. [Google Scholar]
- Di Matteo, N.; Cavigioli, L.; Luiselli, L.; Andreone, F. Anfibi e Rettili D’Italia; Edizioni Belvedere: Latina, Italy, 2019. [Google Scholar]
- Zuffi, M.A.L.; Casu, V.; Marino, S. The Italian wall lizard, Podarcis siculus, along the Tuscanian coast of central Italy: Biometrical features and phenotypic patterns. Herpetol. J. 2012, 22, 207–212. [Google Scholar]
- Dietvorst, P.J.M.; Strijbosch, H.; Bonnemayer, J.J.A.M. The Northernmost Population of Podarcis muralis (Lacertilia, Lacertidae). Amphibia-Reptilia 1980, 1, 161–172. [Google Scholar] [CrossRef]
- Chondropoulos, B.P.; Lykakis, J.J. Ecology of the Balkan Wall Lizard, Podarcis taurica ionica (Sauria: Lacertidae) from Greece. Copeia 1983, 4, 991–1001. [Google Scholar] [CrossRef]
- Perez i de Lanuza, G.; Carretero, M.A.; Font, E. Intensity of male-male competition predicts morph diversity in a color polymorphic lizard. Evolution 2017, 71, 1832–1840. [Google Scholar] [CrossRef]
- Díaz, J.A.; Verdú-Ricoy, J.; Iraeta, P.; Llanos-Garrido, A.; Pérez-Rodríguez, A.; Salvador, A. There is more to the picture than meets the eye: Adaptation for crypsis blurs phylogeographical structure in a lizard. J. Biogeogr. 2017, 44, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Miñano, M.R.; While, G.M.; Yang, W.; Burridge, C.P.; Sacchi, R.; Zuffi, M.; Scali, S.; Salvi, D.; Uller, T. Climate Shapes the Geographic Distribution and Introgressive Spread of Color Ornamentation in Common Wall Lizards. Am. Nat. 2021, 198, 379–393. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AIC | |||
|---|---|---|---|
| Phenotype | Null | Env. Het. | AUC |
| campestris | 1713.4 | 1251.5 | 0.843 |
| concolor | 352.21 | 317.83 | 0.713 |
| striped | 846.79 | 817.84 | 0.660 |
| reticulated | 1315.9 | 694.46 | 0.934 |
| reticulated/campestris | 406.21 | 340.37 | 0.841 |
| Estimate | Std. Error | Z Value | p-Value | ||
|---|---|---|---|---|---|
| campestris | Intercept | 1.20836 | 0.12066 | 10.015 | <0.001 |
| PC1 | 0.49878 | 0.06012 | 8.297 | <0.001 | |
| PC2 | 0.43560 | 0.05718 | 7.617 | <0.001 | |
| PC3 | −0.19821 | 0.09257 | −2.141 | <0.05 | |
| PC4 | −1.35633 | 0.20879 | −6.496 | <0.001 | |
| concolor | Intercept | −4.38803 | 0.42049 | −10.436 | <0.001 |
| PC1 | −0.18869 | 0.18397 | −1.026 | >0.05 | |
| PC2 | 0.20144 | 0.14434 | 1.396 | >0.05 | |
| PC3 | −0.05147 | 0.24937 | −0.206 | >0.05 | |
| PC4 | 1.66756 | 0.55162 | 3.023 | <0.01 | |
| striped | Intercept | −2.025614 | 0.142466 | −14.218 | <0.001 |
| PC1 | 0.174922 | 0.075795 | 2.308 | <0.05 | |
| PC2 | −0.001489 | 0.079173 | −0.019 | >0.05 | |
| PC3 | 0.243598 | 0.144348 | 1.688 | >0.05 | |
| PC4 | −0.500197 | 0.237406 | −2.107 | <0.05 | |
| reticulated | Intercept | −3.65142 | 0.28494 | −12.815 | <0.001 |
| PC1 | −0.82899 | 0.12458 | −6.654 | <0.001 | |
| PC2 | −0.76792 | 0.09469 | −8.110 | <0.001 | |
| PC3 | −0.11727 | 0.13413 | −0.874 | >0.05 | |
| PC4 | 2.49372 | 0.42512 | 5.866 | <0.001 | |
| reticulated/campestris | Intercept | −4.7651 | 0.5621 | −8.478 | <0.001 |
| PC1 | −0.4345 | 0.2523 | −1.722 | >0.05 | |
| PC2 | −0.5734 | 0.1727 | −3.321 | <0.001 | |
| PC3 | −0.7313 | 0.2540 | −2.879 | <0.001 | |
| PC4 | 1.1870 | 0.7948 | 1.493 | >0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallozzi, F.; Colangelo, P.; Senczuk, G.; Castiglia, R. Phylogeographic and Bioclimatic Determinants of the Dorsal Pattern Polymorphism in the Italian Wall Lizard, Podarcis siculus. Diversity 2022, 14, 519. https://doi.org/10.3390/d14070519
Gallozzi F, Colangelo P, Senczuk G, Castiglia R. Phylogeographic and Bioclimatic Determinants of the Dorsal Pattern Polymorphism in the Italian Wall Lizard, Podarcis siculus. Diversity. 2022; 14(7):519. https://doi.org/10.3390/d14070519
Chicago/Turabian StyleGallozzi, Francesco, Paolo Colangelo, Gabriele Senczuk, and Riccardo Castiglia. 2022. "Phylogeographic and Bioclimatic Determinants of the Dorsal Pattern Polymorphism in the Italian Wall Lizard, Podarcis siculus" Diversity 14, no. 7: 519. https://doi.org/10.3390/d14070519
APA StyleGallozzi, F., Colangelo, P., Senczuk, G., & Castiglia, R. (2022). Phylogeographic and Bioclimatic Determinants of the Dorsal Pattern Polymorphism in the Italian Wall Lizard, Podarcis siculus. Diversity, 14(7), 519. https://doi.org/10.3390/d14070519