
Effects of Landscape Configuration on the Body Condition of Migratory and Resident Tropical Birds

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
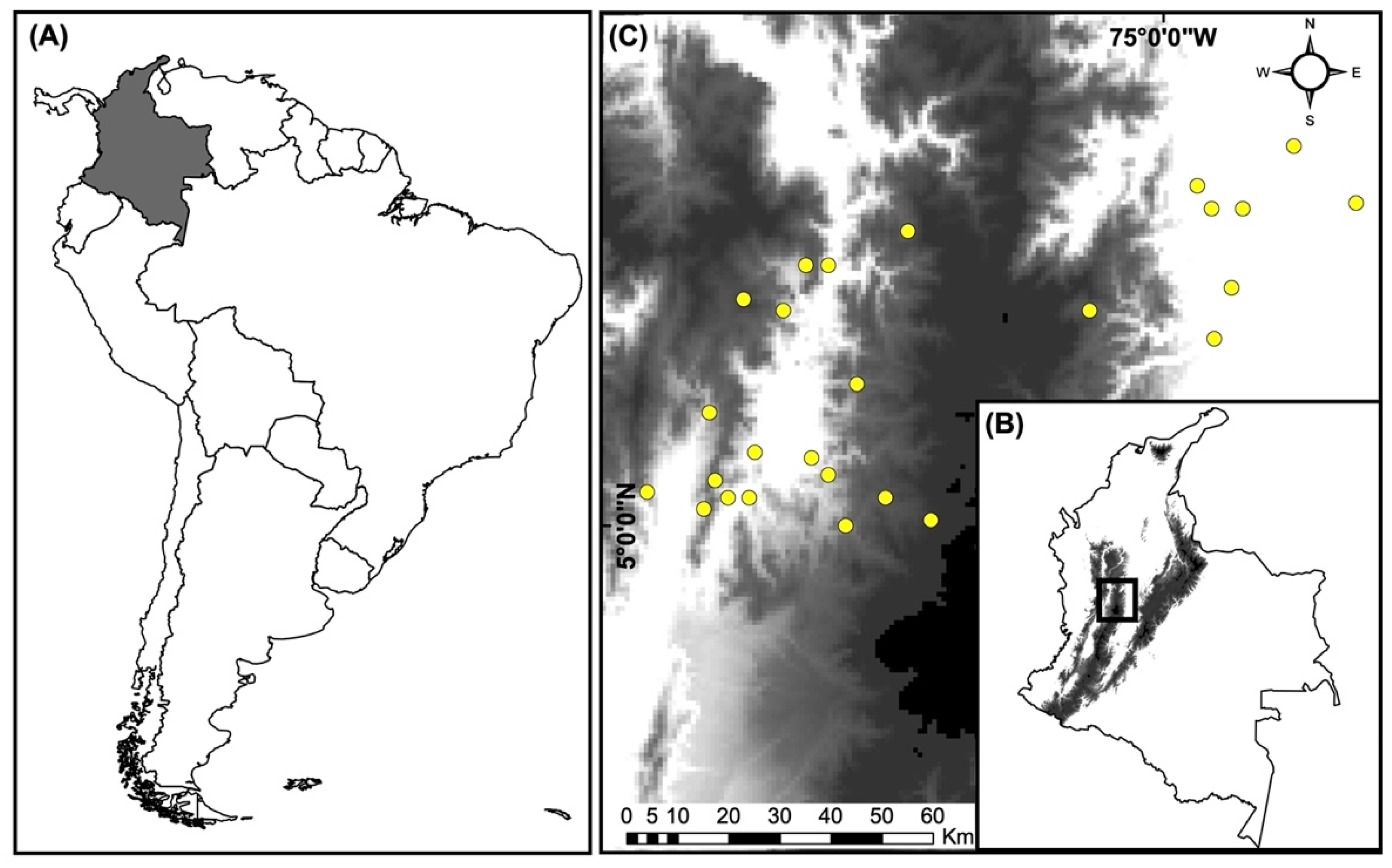
2.1. Study Area
2.2. Information Compilation on Landscapes and Birds
2.3. Data Analysis
3. Results
3.1. Bird Occurrence across Landscapes
3.2. Landscape Characterization
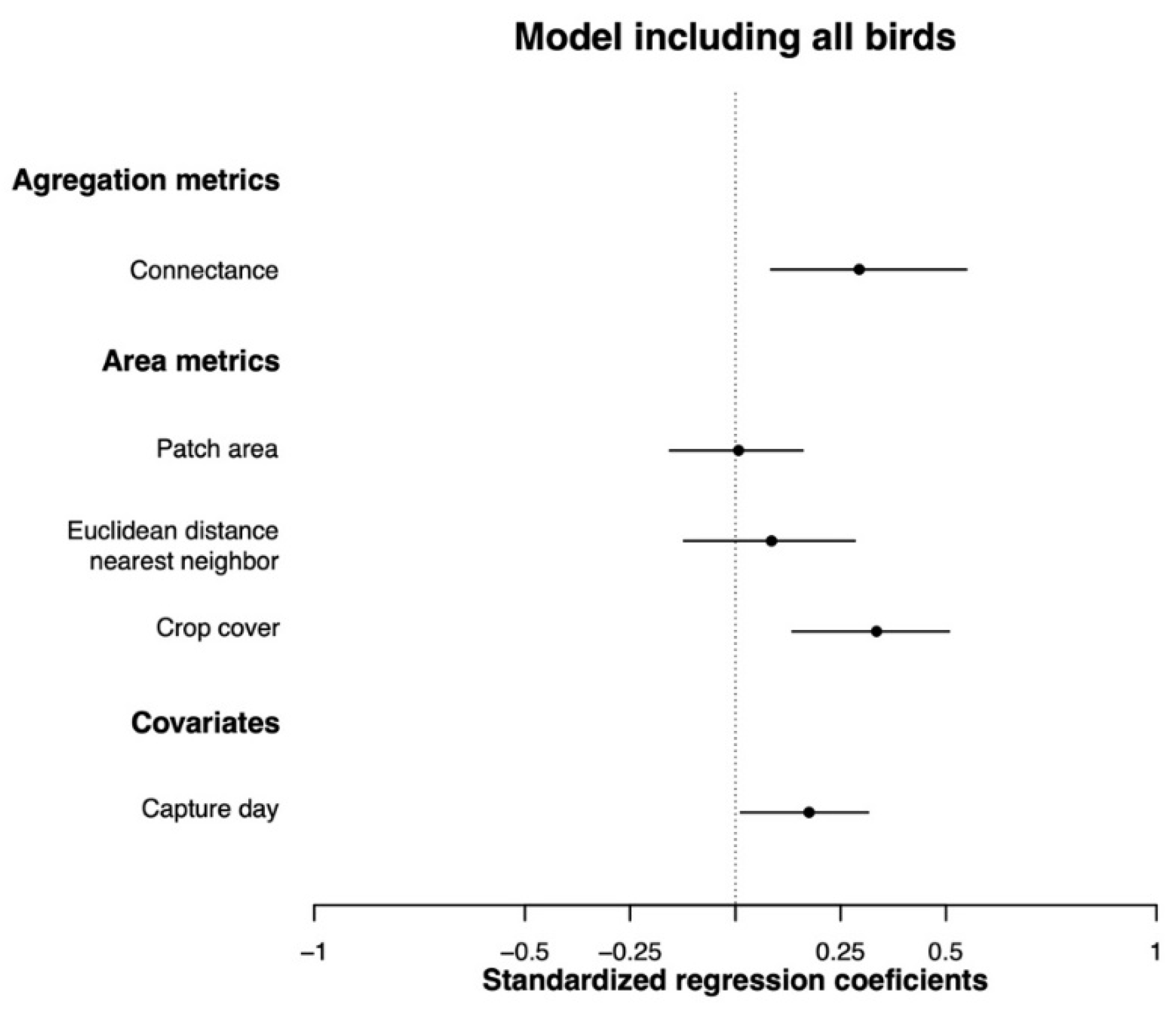
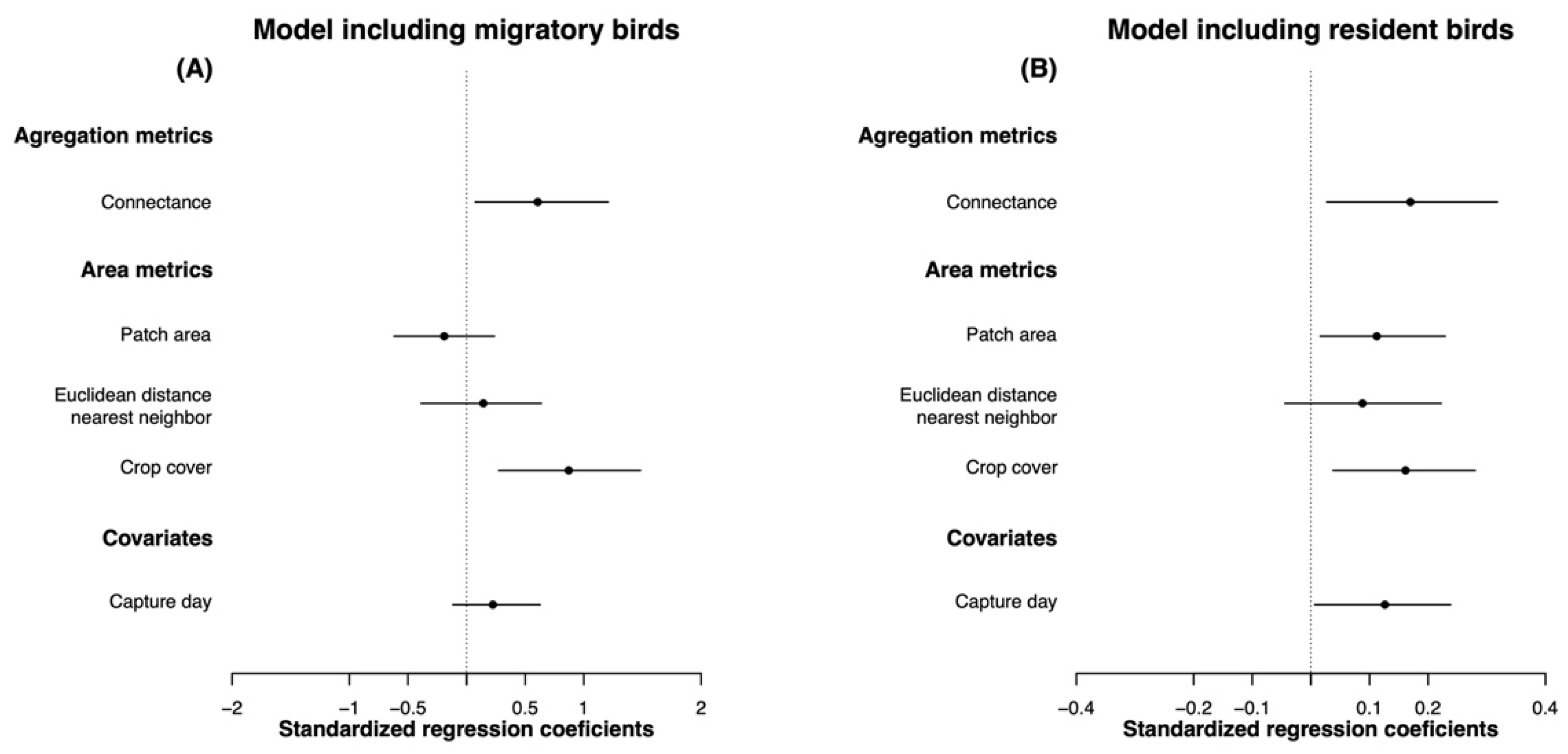
3.3. Effects of Landscape Metrics on Body Condition
4. Discussion
4.1. The Role of Landscape Connectivity
4.2. The Role of Landscape Heterogeneity and Forest Patch Area
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Groombridge, B.; Jenkins, M. World Atlas of Biodiversity: Earth’s Living Resources in the 21st Century; University of California Press: Berkeley, CA, USA, 2002; p. 340. [Google Scholar]
- Murphy, G.E.P.; Romanuk, T.N. Data gaps in anthropogenically driven local-scale species richness change studies across the Earth’s terrestrial biomes. Ecol. Evol. 2016, 6, 2938–2947. [Google Scholar] [CrossRef] [PubMed]
- FAO; UNEP. The State of the World’s Forests 2020: Forests, Biodiversity and People; FAO and UNEP: Rome, Italy, 2020; p. 214. [Google Scholar]
- Lewis, S.L.; Edwards, D.P.; Galbraith, D. Increasing human dominance of tropical forests. Science 2015, 349, 827–832. [Google Scholar] [CrossRef] [PubMed]
- FAO. FAOSTAT Statistics Database; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019. [Google Scholar]
- Dixon, J.; Gibbon, D.P.; Gulliver, A.; Hall, M. Farming Systems and Poverty: Improving Farmers’ Livelihoods in a Changing World; FAO—World Bank: Rome, Italy; Washington, DC, USA, 2001; p. 412. [Google Scholar]
- Taubert, F.; Fischer, R.; Groeneveld, J.; Lehmann, S.; Müller, M.S.; Rödig, E.; Wiegand, T.; Huth, A. Global patterns of tropical forest fragmentation. Nature 2018, 554, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Kattan, G.H.; Alvarez-Lopez, H.; Giraldo, M. Forest Fragmentation and Bird Extinctions: San Antonio Eighty Years Later. Conserv. Biol. 1994, 8, 138–146. [Google Scholar] [CrossRef]
- Laurance, W.F.; Lovejoy, T.E.; Vasconcelos, H.L.; Bruna, E.M.; Didham, R.K.; Stouffer, P.C.; Gascon, C.; Bierregaard, R.O.; Laurance, S.G.; Sampaio, E. Ecosystem Decay of Amazonian Forest Fragments: A 22-Year Investigation. Conserv. Biol. 2002, 16, 605–618. [Google Scholar] [CrossRef] [Green Version]
- Prugh, L.R.; Hodges, K.E.; Sinclair, A.R.E.; Brashares, J.S. Effect of habitat area and isolation on fragmented animal populations. Proc. Natl. Acad. Sci. USA 2008, 105, 20770–20775. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef] [Green Version]
- Henle, K.; Davies, K.F.; Kleyer, M.; Margules, C.; Settele, J. Predictors of Species Sensitivity to Fragmentation. Biodivers. Conserv. 2004, 13, 207–251. [Google Scholar] [CrossRef]
- Rodríguez-Cabal, M.A.; Aizen, M.A.; Novaro, A.J. Habitat fragmentation disrupts a plant-disperser mutualism in the temperate forest of South America. Biol. Conserv. 2007, 139, 195–202. [Google Scholar] [CrossRef]
- Sherry, T.W.; Holmes, R.T. Winter Habitat Quality, Population Limitation, and Conservation of Neotropical-Nearctic Migrant Birds. Ecology 1996, 77, 36–48. [Google Scholar] [CrossRef]
- Hinsley, S.A. The costs of multiple patch use by birds. Landsc. Ecol. 2000, 15, 765–775. [Google Scholar] [CrossRef]
- Keinath, D.A.; Doak, D.F.; Hodges, K.E.; Prugh, L.R.; Fagan, W.; Şekercioğlu, H.; Buchart, S.H.M.; Kauffman, M. A global analysis of traits predicting species sensitivity to habitat fragmentation. Glob. Ecol. Biogeogr. 2017, 26, 115–127. [Google Scholar] [CrossRef]
- Deikumah, J.P.; McAlpine, C.; Maron, M. Matrix Intensification Affects Body and Physiological Condition of Tropical Forest-Dependent Passerines. PLoS ONE 2015, 10, e0128521. [Google Scholar] [CrossRef]
- Pavlacky, D.C.; Possingham, H.P.; Goldizen, A.W. Integrating life history traits and forest structure to evaluate the vulnerability of rainforest birds along gradients of deforestation and fragmentation in eastern Australia. Biol. Conserv. 2015, 188, 89–99. [Google Scholar] [CrossRef]
- Norris, D.R.; Marra, P.; Kyser, T.K.; Sherry, T.W.; Ratcliffe, L.M. Tropical winter habitat limits reproductive success on the temperate breeding grounds in a migratory bird. Proc. R. Soc. B Biol. Sci. 2004, 271, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bearhop, S.; Hilton, G.M.; Votier, S.C.; Waldron, S. Stable isotope ratios indicate that body condition in migrating passerines is influenced by winter habitat. Proc. R. Soc. B Biol. Sci. 2004, 271, S215–S218. [Google Scholar] [CrossRef] [Green Version]
- Kattan, G.H.; Franco, P. Bird diversity along elevational gradients in the Andes of Colombia: Area and mass effects. Glob. Ecol. Biogeogr. 2004, 13, 451–458. [Google Scholar] [CrossRef]
- Fierro, K. Aves migratorias en Colombia. In Plan Nacional de Las Especies Migratorias Diagnóstico e Identificación de Acciones Para la Conservación y el Manejo Sostenible de Las Especies Migratorias de La Biodiversidad en Colombia; Naranjo, L.G., Amaya, J.D., Eds.; Ministro de Ambiente, Vivienda y Desarrollo Territorial: Bogotá, Colombia, 2009; pp. 63–75. [Google Scholar]
- Avendaño, J.E.; Bohórquez, C.I.; Rosselli, L.; Arzuza-Buelvas, D.; Cuervo, A.M.; Stiles, F.G.; Renjifo, L.M. Lista de chequeo de las aves de Colombia: Una síntesis del estado del conocimiento desde Hilty & Brown (1986). Ornitol. Colomb. 2017, 16, eA01. [Google Scholar]
- Armenteras, D.; Rodríguez, N.; Retana, J.; Morales, M. Understanding deforestation in montane and lowland forests of the Colombian Andes. Reg. Environ. Chang. 2011, 11, 693–705. [Google Scholar] [CrossRef]
- Renjifo, L.M. Composition Changes in a Subandean Avifauna after Long-Term Forest Fragmentation. Conserv. Biol. 1999, 13, 1124–1139. [Google Scholar] [CrossRef]
- Castaño-Villa, G.J.; Patiño-Zabala, Y.J.C. Extinciones locales de aves en fragmentos de bosque en la región de Santa Elena, Andes Centrales. Colombia. Hornero 2008, 23, 23–34. [Google Scholar]
- Velásquez-Chaverra, E.d.J.; Arias, C.H. Anuario Estadístico de Frutas y Hortalizas 2004–2008 y Sus Calendarios de Siembras y Cosechas; Ministerio de Agricultura y Desarrollo Rural: Bogotá, Colombia, 2009.
- Castaño-Villa, G.J.; Estevez, J.V.; Fontúrbel, F.E. The role of native forest plantations in the conservation of Neotropical birds: The case of the Andean alder. J. Nat. Conserv. 2014, 22, 547–551. [Google Scholar] [CrossRef]
- Castaño-Villa, G.J.; Ramos-Valencia, S.A.; Fontúrbel, F.E. Fine-scale habitat structure complexity determines insectivorous bird diversity in a tropical forest. Acta Oecologica 2014, 61, 19–23. [Google Scholar] [CrossRef]
- Martínez-Sánchez, E.T.; Romero, M.C.; Páez, F.A.R.; Cárdenas, J.E.P.; Castaño-Villa, G.J. Contribution of agroecosystems to the conservation of bird diversity in the department of Caldas. Rev. Fac. Nac. Agron. 2018, 71, 8445–8457. [Google Scholar] [CrossRef] [Green Version]
- Cardona-Salazar, L.J.; Benavides-Ossa, Y.A.; Vargas-Daza, M.; Betancurt-Grisales, J.F.; Bohada-Murillo, M.; Martínez-Sánchez, E.T.; Cardona-Romero, M.; Busi, A.; Tobón-Escobar, W.D.; Ortíz-Giraldo, M.; et al. A morphological, reproductive, and molt phenology database for 379 bird species from the Colombian Tropical Andes. Ecology 2020, 101, e03016. [Google Scholar] [CrossRef] [Green Version]
- Hilty, S.L.; Brown, W.L. A Guide to the Birds of Colombia; Princeton University Press: Princeton, NJ, USA, 1986; p. 836. [Google Scholar]
- Marra, P.P.; Francis, C.M.; Mulvihill, R.S.; Moore, F.R. The influence of climate on the timing and rate of spring bird migration. Oecologia 2005, 142, 307–315. [Google Scholar] [CrossRef]
- Ayerbe-Quiñones, F. Guía Ilustrada de La Avifauna Colombiana, 1st ed.; Wildlife Conservation Society: Bogotá, Colombia, 2018. [Google Scholar]
- Remsen, J.V.; Areta, J.I.; Bonaccorso, S.; Claramunt, S.; Jaramillo, A.; Pacheco, J.F.; Ribas, C.; Robbins, M.B.; Stiles, F.G.; Stotz, D.F.; et al. A classification of the bird species of South America. Am. Ornithol. Soc. 2019. Available online: http://www.museum.lsu.edu/~Remsen/SACCBaseline.html (accessed on 5 September 2019).
- Aborn, D.A.; Moore, F.R. Pattern of Movement by Summer Tanagers (Piranga Rubra) During Migratory Stopover: A Telemetry Study. Behaviour 1997, 134, 1077–1100. [Google Scholar] [CrossRef]
- Boscolo, D.; Metzger, J.P. Is bird incidence in Atlantic forest fragments influenced by landscape patterns at multiple scales? Landsc. Ecol. 2009, 24, 907–918. [Google Scholar] [CrossRef]
- Peig, J.; Green, A.J. New perspectives for estimating body condition from mass/length data: The scaled mass index as an alternative method. Oikos 2009, 118, 1883–1891. [Google Scholar] [CrossRef]
- Peig, J.; Green, A.J. The paradigm of body condition: A critical reappraisal of current methods based on mass and length. Funct. Ecol. 2010, 24, 1323–1332. [Google Scholar] [CrossRef]
- Bell, S.C.; Owens, I.P.; Lord, A.M. Quality of breeding territory mediates the influence of paternal quality on sex ratio bias in a free-living bird population. Behav. Ecol. 2014, 25, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Forman, R.T.T.; Godron, M. Patches and Structural Components for a Landscape Ecology. BioScience 1981, 31, 733–740. [Google Scholar] [CrossRef]
- Turner, M.G. Landscape Ecology: The Effect of Pattern on Process. Annu. Rev. Ecol. Syst. 1989, 20, 171–197. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.; Ene, E. FRAGSTATS 4.2: Spatial Pattern Analysis Program for Categorical Maps; University of Massachusetts: Amherst, MA, USA, 2012. [Google Scholar]
- Gergel, S.E.; Turner, M.G.; Miller, J.R.; Melack, J.M.; Stanley, E.H. Landscape indicators of human impacts to riverine systems. Aquat. Sci. 2002, 64, 118–128. [Google Scholar] [CrossRef]
- Schielzeth, H. Simple means to improve the interpretability of regression coefficients. Methods Ecol. Evol. 2010, 1, 103–113. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing, Reference Index Version 3.6.1; Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Naimi, B. Usdm: Uncertainty Analysis for Species Distribution Models, R package version 1.1–15; R Core Team: Vienna, Austria, 2010. [Google Scholar]
- Hadfield, J.D. MCMC Methods for Multi-Response Generalized Linear Mixed Models: TheMCMCglmmRPackage. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Tscharntke, T.; Tylianakis, J.; Rand, T.A.; Didham, R.K.; Fahrig, L.; Batary, P.; Bengtsson, J.; Clough, Y.; Crist, T.O.; Dormann, C.; et al. Landscape moderation of biodiversity patterns and processes—Eight hypotheses. Biol. Rev. 2012, 87, 661–685. [Google Scholar] [CrossRef]
- Camacho, C.; Palacios, S.; Sáez, P.; Sánchez, S.; Potti, J. Human-Induced Changes in Landscape Configuration Influence Individual Movement Routines: Lessons from a Versatile, Highly Mobile Species. PLoS ONE 2014, 9, e104974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, S.N.; Rodewald, P.G. Movement behaviour of a forest songbird in an urbanized landscape: The relative importance of patch-level effects and body condition during migratory stopover. Landsc. Ecol. 2010, 25, 955–965. [Google Scholar] [CrossRef]
- Richard, Y.; Armstrong, D.P. Cost distance modelling of landscape connectivity and gap-crossing ability using radio-tracking data. J. Appl. Ecol. 2010, 47, 603–610. [Google Scholar] [CrossRef]
- Amos, J.N.; Balasubramaniam, S.; Grootendorst, L.; Harrisson, K.A.; Lill, A.; Mac Nally, R.; Pavlova, A.; Radford, J.Q.; Takeuchi, N.; Thomson, J.R.; et al. Little evidence that condition, stress indicators, sex ratio, or homozygosity are related to landscape or habitat attributes in declining woodland birds. J. Avian Biol. 2013, 44, 45–54. [Google Scholar] [CrossRef]
- Loiselle, B.; Blake, J.G. Temporal Variation in Birds and Fruits Along an Elevational Gradient in Costa Rica. Ecology 1991, 72, 180–193. [Google Scholar] [CrossRef]
- Morales-Betancourt, J.A.; Castaño-Villa, G.J.; Fontúrbel, F.E. Resource abundance and frugivory in two manakin species (Aves: Pipridae) inhabiting a reforested area in Colombia. J. Trop. Ecol. 2012, 28, 511–514. [Google Scholar] [CrossRef]
- Ewers, R.M.; Didham, R.K. Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. 2006, 81, 117–142. [Google Scholar] [CrossRef]
- Hewison, A.M.; Morellet, N.; Verheyden, H.; Daufresne, T.; Angibault, J.-M.; Cargnelutti, B.; Merlet, J.; Picot, D.; Rames, J.-L.; Joachim, J.; et al. Landscape fragmentation influences winter body mass of roe deer. Ecography 2009, 32, 1062–1070. [Google Scholar] [CrossRef]
- Öberg, S. Influence of landscape structure and farming practice on body condition and fecundity of wolf spiders. Basic Appl. Ecol. 2009, 10, 614–621. [Google Scholar] [CrossRef]
- Salazar, D.A.; Fontúrbel, F.E. Beyond habitat structure: Landscape heterogeneity explains the monito del monte (Dromiciops gliroides) occurrence and behavior at habitats dominated by exotic trees. Integr. Zool. 2016, 11, 413–421. [Google Scholar] [CrossRef]
- Fontúrbel, F.E.; Salazar, D.A.; Medel, R. Increased resource availability prevents the disruption of key ecological interactions in disturbed habitats. Ecosphere 2017, 8, e01768. [Google Scholar] [CrossRef]
- Hansen, N.A.; Scheele, B.C.; Driscoll, D.A.; Lindenmayer, D.B. Amphibians in agricultural landscapes: The habitat value of crop areas, linear plantings and remnant woodland patches. Anim. Conserv. 2019, 22, 72–82. [Google Scholar] [CrossRef]
- Okada, S.; Lindenmayer, D.B.; Wood, J.T.; Crane, M.J.; Pierson, J. How does a transforming landscape influence bird breeding success? Landsc. Ecol. 2017, 32, 1039–1048. [Google Scholar] [CrossRef]
- Greenberg, R.; Bichier, P.; Angon, A.C.; Reitsma, R. Bird Populations in Shade and Sun Coffee Plantations in Central Guatemala. Conserv. Biol. 1997, 11, 448–459. [Google Scholar] [CrossRef]
- Boesing, A.L.; Nichols, E.; Metzger, J.P. Effects of landscape structure on avian-mediated insect pest control services: A review. Landsc. Ecol. 2017, 32, 931–944. [Google Scholar] [CrossRef]
- Murillo, M.B.; Castaño-Villa, G.J.; Fontúrbel, F.E. The effects of forestry and agroforestry plantations on bird diversity: A global synthesis. Land Degrad. Dev. 2019, 31, 646–654. [Google Scholar] [CrossRef]
- Campo-Celada, M.; Jordano, P.; Benítez-López, A.; Gutiérrez-Expósito, C.; Rabadán-González, J.; Mendoza, I. Assessing short and long-term variations in diversity, timing and body condition of frugivorous birds. Oikos 2022, 2022, e08387. [Google Scholar] [CrossRef]
- Marra, P.P.; Holmes, R.T. Consequences of Dominance-Mediated Habitat Segregation in American Redstarts During the Nonbreeding Season. Ornithology 2001, 118, 92–104. [Google Scholar] [CrossRef]
- Johnson, M.D.; Sherry, T.W.; Holmes, R.T.; Marra, P.P. Assessing Habitat Quality for a Migratory Songbird Wintering in Natural and Agricultural Habitats. Conserv. Biol. 2006, 20, 1433–1444. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Category | Index | Description |
|---|---|---|
| Aggregation metrics | Connectance index (CONNECT) | The percentage of functional joinings between forest patches at a maximum distance of 200 m |
| Euclidean nearest-neighbor distance (ENN_MN) | Average straight-line distance in meters to the nearest forest | |
| Area metrics | Largest patch index, crop or grassland patch index (LPI) | Percentage of the landscape area that is occupied by the largest area of forest (LPI_forest), crops (LPI_crops), or grasslands (LPI_grasslands) |
| Patch area (PATCH_AREA_MN) | Mean area in hectares of forest patches |
| Category | Index | Mean | Min | Max |
|---|---|---|---|---|
| Aggregation | CONNECT (%) | 32.43 | 30.89 | 285.30 |
| ENN_MN (m) | 62.46 | 30.00 | 344.80 | |
| Area | LPI_CROP (%) | 14.40 | 0.97 | 69.36 |
| LPI_FOREST (%) | 47.12 | 3.87 | 79.52 | |
| LPI_GRASSLAND (%) | 7.41 | 0.02 | 80.53 | |
| PATCH_AREA_MN (ha) | 128.62 | 18.76 | 336.76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molina-Marin, D.A.; Rodas-Rua, J.C.; Lara, C.E.; Rivera-Páez, F.A.; Fontúrbel, F.E.; Castaño-Villa, G.J. Effects of Landscape Configuration on the Body Condition of Migratory and Resident Tropical Birds. Diversity 2022, 14, 432. https://doi.org/10.3390/d14060432
Molina-Marin DA, Rodas-Rua JC, Lara CE, Rivera-Páez FA, Fontúrbel FE, Castaño-Villa GJ. Effects of Landscape Configuration on the Body Condition of Migratory and Resident Tropical Birds. Diversity. 2022; 14(6):432. https://doi.org/10.3390/d14060432
Chicago/Turabian StyleMolina-Marin, Dimas A., Juan C. Rodas-Rua, Carlos E. Lara, Fredy A. Rivera-Páez, Francisco E. Fontúrbel, and Gabriel J. Castaño-Villa. 2022. "Effects of Landscape Configuration on the Body Condition of Migratory and Resident Tropical Birds" Diversity 14, no. 6: 432. https://doi.org/10.3390/d14060432
APA StyleMolina-Marin, D. A., Rodas-Rua, J. C., Lara, C. E., Rivera-Páez, F. A., Fontúrbel, F. E., & Castaño-Villa, G. J. (2022). Effects of Landscape Configuration on the Body Condition of Migratory and Resident Tropical Birds. Diversity, 14(6), 432. https://doi.org/10.3390/d14060432