
Comparative Analysis of the Polymorphism of the Casein Genes in Camels Bred in Kazakhstan

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
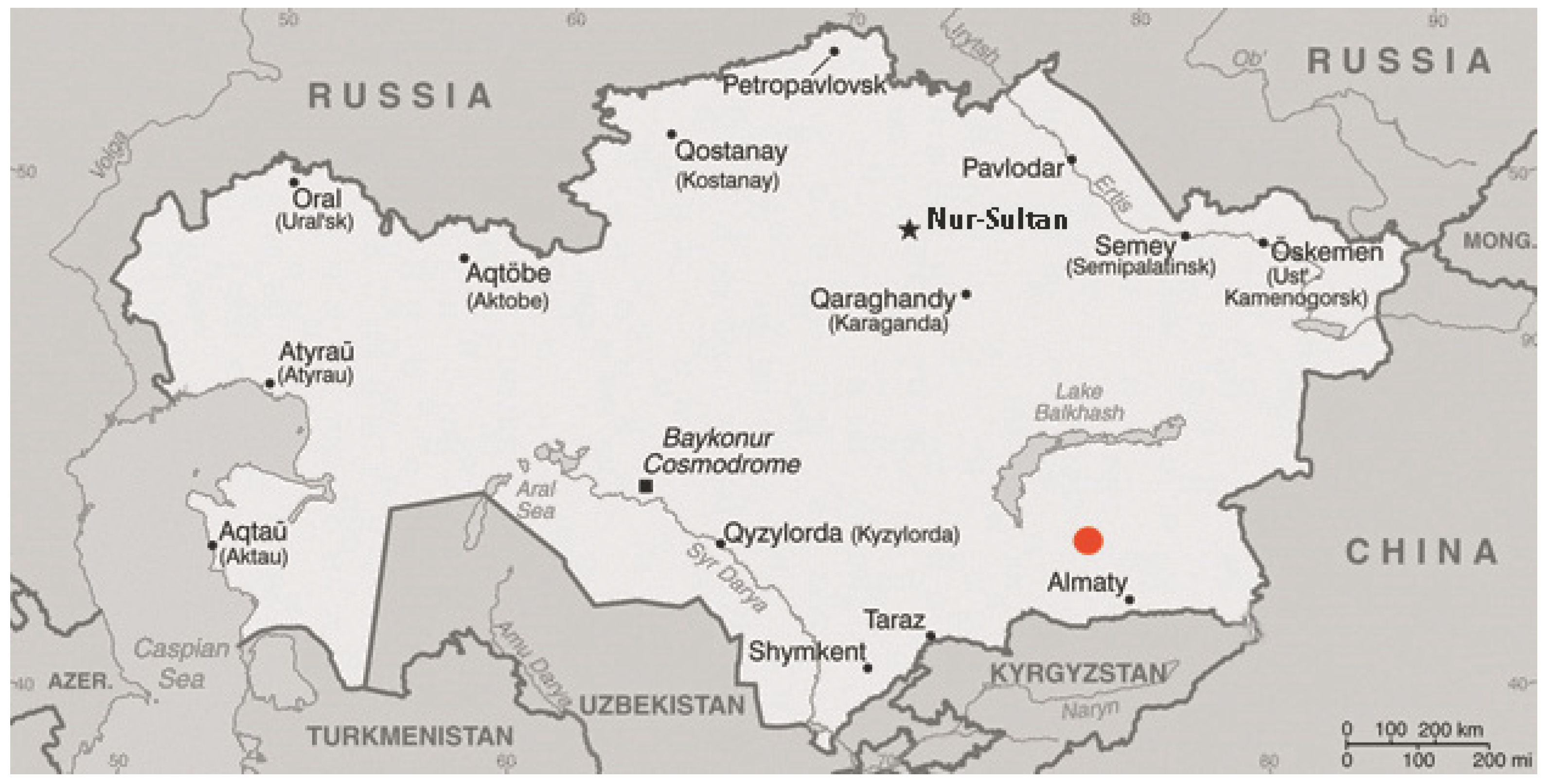
2.1. Collection of Materials
2.2. Genomic DNA Extraction and PCR-RFLP Analysis
2.3. Statistical Data Processing
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dioli, M. Dromedary (Camelus dromedarius) and Bactrian camel (Camelus bactrianus) crossbreeding husbandry practices in Turkey and Kazakhstan: An in-depth review. Pastor. Res. Policy Pract. 2020, 10, 6. [Google Scholar] [CrossRef]
- Ming, L.; Yuan, L.; Yi, L.; Ding, G.; Hasi, S.; Chen, G.; Jambl, T.; Hedayat-Evright, N.; Batmunkh, M.; Badmaevna, G.K.; et al. Whole-genome sequencing of 128 camels across Asia reveals origin and migration of domestic Bactrian camels. Commun. Biol. 2020, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Almathen, F.; Charruau, P.; Mohandesan, E.; Mwacharo, J.M.; Orozco-Terwengel, P.; Pitt, D.; Abdussamad, A.M.; Uerpmann, M.; Uerpmann, H.-P.; De Cupere, B.; et al. Ancient and modern DNA reveal dynamics of domestication and cross-continental dispersal of the dromedary. Proc. Natl. Acad. Sci. USA 2016, 113, 6707–6712. [Google Scholar] [CrossRef] [PubMed]
- Farah, Z. Composition and characteristics of camel milk. J. Dairy Res. 1993, 60, 603–626. [Google Scholar] [CrossRef] [PubMed]
- Mal, G.; Sena, D.S.; Jain, V.K.; Sahani, M.S. Therapeutic Value of Camel Milk as a Nutritional Supplement for Multiple Drug Resistant (MDR) Tuberculosis Patients. Isr. J. Vet. Med. 2006, 61, 88–91. [Google Scholar]
- Pauciullo, A.; Shuiep, E.; Cosenza, G.; Ramunno, L.; Erhardt, G. Molecular characterization and genetic variability at κ-casein gene (CSN3) in camels. Gene 2012, 15, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Shori, A.B. Camel milk as a potential therapy for controlling diabetes and its complications: A review of in vivo studies. J. Food Drug Anal. 2015, 23, 609–618. [Google Scholar] [CrossRef]
- Rachagani, S.; Gupta, I.D. Bovine kappa-casein gene polymorphism and its association with milk production traits. Genet. Mol. Biol. 2008, 31, 893–897. [Google Scholar] [CrossRef]
- Agrawal, R.P.; Beniwal, R.; Kochar, D.K.; Tuteja, F.C.; Ghorui, S.K.; Sahani, M.S.; Sharma, S. Camel milk as an adjunct to insulin therapy improves long-term glycemic control and reduction in doses of insulin in patients with type-1 diabetes: A 1 year randomized controlled trial. Diabetes Res. Clin. Pract. 2005, 68, 176–177. [Google Scholar] [CrossRef]
- Beg, O.U.; von Bahr-Lindström, H.; Zaidi, Z.H.; Jörnvall, H. Characterization of a camel milk protein rich in proline identifies a new β-casein fragment. Regul. Pept. 1986, 15, 55–61. [Google Scholar] [CrossRef]
- Shamsia, S.M. Nutritional and therapeutic properties of camel and human milks. Int. J. Genet. Mol. Biol. 2009, 1, 52–58. [Google Scholar]
- Faye, B. The enthusiasm for camel production. Editor. Emir. J. Food Agric. 2018, 30, 249–250. [Google Scholar]
- Nagy, P.; Fabri, Z.N.; Varga, L.; Reiczigel, J.; Juhasz, J. Effect of genetic and nongenetic factors on chemical composition of individual milk samples from dromedary camels (Camelus dromedarius) under intensive management. J. Dairy Sci. 2017, 100, 8680–8693. [Google Scholar] [CrossRef] [PubMed]
- Burger, P.A.; Clani, E.; Faye, B. Old World camels in a modern world—A balancing act between conservation and genetic improvement. Anim. Genet. 2019, 50, 598–612. [Google Scholar] [CrossRef]
- Hipp, N.J.; Groves, M.L.; Custer, J.N.; McMeekin, T.L. Separation of α-, β- and γ-casein. J. Dairy Sci. 1952, 35, 272–281. [Google Scholar] [CrossRef]
- Alhaj, O.A.; Al-kanhal, H.A. Compositional, technological and nutritional aspects of dromedary camel milk. Int. Dairy J. 2010, 20, 811–821. [Google Scholar] [CrossRef]
- Erhardt, G.; Shuiep, E.S.; Lisson, M.; Weimann, C.; Wang, Z.; Ibtisam, E.M.; Zubeir, E.I.; Pauciullo, A. Alpha S1-casein polymorphisms in camel (Camelus dromedarius) and descriptions of biological active peptides and allergenic epitopes. Trop. Anim. Health Prod. 2016, 48, 879–887. [Google Scholar] [CrossRef]
- Ryskaliyeva, A.; Henry, C.; Miranda, G.; Faye, B.; Konuspayeva, G.; Martin, P. Combining different proteomic approaches to resolve complexity of the milk protein fraction of dromedary, Bactrian camels and hybrids, from different regions of Kazakhstan. PLoS ONE 2018, 13, e0197026. [Google Scholar] [CrossRef]
- Singh, K.; Jayakumar, S.; Dixit, S.P.; Malik, S.Z. Molecular characterization and genetic variability of Alpha Casein gene, CSN1S1 in Bikaneri camel (Camelus dromedarius) milk. Indian J. Anim. Res. 2018, 53, 67–70. [Google Scholar] [CrossRef]
- Mati, A.; Senoussi-Ghezali, C.; Si Ahmed Zennia, S.; Almi-Sebbane, D.; El-Hatmi, H.; Girardet, J.M. Dromedary camel milk proteins, a source of peptides having biological activities—A review. Int. Dairy J. 2017, 73, 25–37. [Google Scholar] [CrossRef]
- Pauciullo, A.; Ogah, D.M.; Iannaccone, M.; Erhardt, G.; Stasio, L.D.; Cosenza, G. Genetic characterization of the oxytocinneurophysin I gene (OXT) and its regulatory regions analysis in domestic Old and New World camelids. PLoS ONE 2018, 13, e0195407. [Google Scholar] [CrossRef] [PubMed]
- Alim, N.; Fondrini, F.; Ionizzi, I.; Feligini, M.; Enne, G. Characterization of casein fractions from Algerian dromedary (Camelus dromedarius) milk. Pak. J. Nutr. 2005, 4, 112–116. [Google Scholar] [CrossRef]
- Yelubayeva, M.E.; Buralkhiyev, B.A.; Tyshchenko, V.I.; Terletskiy, V.P.; Usenbekov, Y.S. Results of Camelus dromedarius and Camelus bactrianus Genotyping by Alpha-S1-Casein, Kappa-Casein Loci, and DNA Fingerprinting. Cytol. Genet. 2018, 52, 179–185. [Google Scholar] [CrossRef]
- Shild, T.A.; Geldermann, H. Variants within the 5’-flanking regions of bovine milk-protein-encoding genes. III. Genes encoding the Ca-sensitive Caseins alpha-s1, alpha-s2 and beta. Theor. Appl. Genet. 1996, 93, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, S.A.; Umrikar, U.D.; Sawane, M.P.; Pawar, V.D.; Deshmukh, R.S.; Dahiya, S.S.; Mehta, S.C. Genetic polymorphism at k-casein gene in Indian camel breeds (Camelus dromedarius). J. Camel Pract. Res. 2020, 27, 201–206. [Google Scholar] [CrossRef]
- Pauciullo, A.; Giambra, I.; Iannuzzi, L.; Erhardt, G. The β-casein in camels: Molecular characterization of the CSN2 gene, promoter analysis and genetic variability. Gene 2014, 547, 159–168. [Google Scholar] [CrossRef]
- Amigo, L.; Recio, I.; Ramos, M. Genetic polymorphism of ovine milk proteins: Its influence on technological properties of milk—A review. Int. Dairy J. 2000, 10, 135–149. [Google Scholar] [CrossRef]
- Soyudal, B.; Ardicli, S.; Samli, H.; Dincel, D.; Balci, F. Association of polymorphisms in the CSN2, CSN3, LGB and LALBA genes with milk production traits in Holstein cows raised in Turkey. J. Hellenic Vet. Med. Soc. 2018, 69, 1271–1282. [Google Scholar] [CrossRef]
- Nowier, A.M.; Ramadan, S.I. Association of β-casein gene polymorphism with milk composition traits of Egyptian Maghrebi camels (Camelus dromedarius). Arch. Anim. Breed. 2020, 63, 493–500. [Google Scholar] [CrossRef]
- Giambra, I.J.; Chianese, L.; Ferranti, P.; Erhardt, G. Short communication: Molecular genetic characterization of ovine αS1-casein allele H caused by alternative splicing. J. Dairy Sci. 2010, 93, 792–795. [Google Scholar] [CrossRef]
- Ramunno, L.; Cosenza, G.; Rando, A.; Pauciullo, A.; Illario, R.; Gallo, D.; Di Berardino, D.; Masina, P. Comparative analysis of gene sequence of goat CSN1S1 F and N alleles and characterization of CSN1S1 transcript variants in mammary gland. Gene 2005, 345, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Kappeler, S.; Farah, Z.; Puhan, Z. Sequence analysis of Camelus dromedarius milk caseins. J. Dairy Res. 1998, 65, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Shuiep, E.; Giambra, I.J.; El Zubeir, I.E.M.; Erhardt, G. Biochemical and molecular characterization of polymorphisms of αs1-casein in Sudanese camel (Camelus dromedarius) milk. Int. Dairy J. 2013, 28, 88–93. [Google Scholar] [CrossRef]
- Kim, J.; Yu, J.; Bag, J.; Bakovic, M.; Cant, J.P. Translation attenuation via 3′ terminal codon usage in bovine csn1s2 is responsible for the difference in αs2- and β-casein profile in milk. RNA Biol. 2015, 12, 354–367. [Google Scholar] [CrossRef][Green Version]
- Kharrati, H.K.; Esmailizadeh, A.K. SNPs genotyping technologies and their applications in farm animals breeding programs: Review. Braz. Arch. Biol. Technol. 2014, 57, 87–95. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlex 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Yeh, F.C.; Yang, R.C.; Boyle, T. POPGENE Version 1.32: Microsoft Window-Based Freeware for Population Genetics Analysis; University of Alberta: Edmonton, AB, Canada, 1999. [Google Scholar]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987; pp. 176–187. [Google Scholar] [CrossRef]
- Kimura, M.; Crow, F.J. The number of alleles that can be maintained in a finite population. Genetics 1964, 49, 725–738. [Google Scholar] [CrossRef]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed]
- Lewontin, R.C. The Apportionment of Human Diversity. Evol. Biol. 1972, 6, 381–398. [Google Scholar]
- Smekenov, I.T.; Akishev, Z.D.; Altybayeva, N.A.; Mukhitdinov, N.M.; Bisenbayev, A.K. Evaluation of genetic polymorphism of the Berberis iliensis population in the Ili-Balkhash region based on ISSR markers. Rep. Natl. Acad. Sci. Repub. Kazakhstan 2012, 4, 49–57. [Google Scholar]
- Reiner, A.P.; Gross, M.D.; Carlson, C.S.; Bielinski, S.J.; Lange, L.A.; Fornage, M.; Jenny, N.S.; Walston, J.; Tracy, R.P.; Williams, O.D.; et al. Common coding variants of the HNF1A gene are associated with multiple cardiovascular risk phenotypes in community-based samples of younger and older European American adults: The coronary artery risk development in young adults study and the cardiovascular health study. Circ. Cardiovasc. Genet. 2009, 2, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Othman, E.O.; Nowier, A.M.; Medhat, E.E. Genetic Variations in Two Casein Genes Among Maghrabi Camels Reared in Egypt. Biosci. Biotechnol. Res. Asia 2016, 13, 473–480. [Google Scholar] [CrossRef][Green Version]
- Giambra, I.J.; Brandt, H.; Erhardt, G. Milk protein variants are highly associated with milk performance traits in East Friesian Dairy and Lacaune sheep. Small Rumin. Res. 2014, 121, 382–394. [Google Scholar] [CrossRef]
- Caroli, A.M.; Chessa, S.; Erhardt, G.J. Invited review: Milk protein genetic variation in cattle: Impact on animal breeding and human nutrition. J. Dairy Sci. 2009, 92, 5335–5352. [Google Scholar] [CrossRef]
- Imamura, K. Camel Production in Kazakhstan. J. Nagoya Gakuin Univ. 2015, 52, 1–13. [Google Scholar]
- Tanana, L.A.; Karaba, V.I.; Peshko, V.V. Breeding of Farm Animals and the Basics of Selection: A Tutorial; Republican Institute of Professional Education: Minsk, Belarus, 2017; pp. 236–237. [Google Scholar]



{kind=link}
{kind=link}
| № | SNP | Fragment Length | Primers | Annealing Temperature | Restriction Enzymes | References |
|---|---|---|---|---|---|---|
| 1 | CSN3 g.1029T > C | 488 bp | 5′-CACAAAGATGACTCTGCTATCG-3′ 5′-GCCCTCCACATATGTCTG-3′. | 60 °C | AluI | [6] |
| 2 | CSN2 g.2126A > G | 659 bp | 5′-GTTTCTCCATTACAGCATC-3′ 5′-TCAAATCTATACAGGCACTT-3′. | 53 °C | HphI | [26] |
| 3 | CSN1S1 c.150G > T | 930 bp | 5′-TGAACCAGACAGCATAGAG-3′ 5′-CTAAACTGAATGGGTGAAAC-3′ | 55 °C | SmoI | [17] |
| Population | Overall | CSN3 | CSN2 | CSN1S1 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identified Genotypes | Allele Frequency | Identified Genotypes | Allele Frequency | Identified Genotypes | Allele Frequency | |||||||||||
| CC | CT | TT | C | T | AA | AG | GG | A | G | GG | GT | TT | G | T | ||
| 1 | 8 | 1 | 3 | 4 | 0.30 | 0.70 | 5 | 2 | 1 | 0.75 | 0.25 | 6 | 2 | 0 | 0.88 | 0.12 |
| 2 | 9 | 1 | 2 | 6 | 0.18 | 0.82 | 4 | 2 | 3 | 0.60 | 0.40 | 8 | 1 | 0 | 0.94 | 0.06 |
| 3 | 20 | 2 | 10 | 8 | 0.37 | 0.63 | 18 | 2 | 0 | 0.95 | 0.05 | 20 | 0 | 0 | 1.00 | 0 |
| 4 | 16 | 8 | 3 | 5 | 0.44 | 0.56 | 1 | 6 | 9 | 0.25 | 0.75 | 14 | 2 | 0 | 0.94 | 0.06 |
| Overall | 53 | 12 | 18 | 23 | 0.40 | 0.60 | 28 | 12 | 13 | 0.64 | 0.36 | 48 | 5 | 0 | 0.94 | 0.06 |
| χ2 = 12.1 | χ2 = 8.6 | χ2 = 14.5 | ||||||||||||||
| Populations | Overall | CSN3 | CSN2 | CSN1S1 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identified Genotypes | Allele Frequency | Identified Genotypes | Allele Frequency | Identified Genotypes | Allele Frequency | |||||||||||
| CC | CT | TT | C | T | AA | AG | GG | A | G | GG | GT | TT | G | T | ||
| One-humped camels (Camelus dromedarius) | 9 | 1 | 4 | 4 | 0.33 | 0.67 | 5 | 2 | 2 | 0.66 | 0.33 | 7 | 2 | 0 | 0.89 | 0.11 |
| Two-humped camels (Camelus bactrianus) | 22 | 8 | 7 | 7 | 0.52 | 0.48 | 7 | 7 | 8 | 0.48 | 0.52 | 20 | 2 | 0 | 0.95 | 0.05 |
| Hybrids | 22 | 3 | 7 | 12 | 0.30 | 0.70 | 16 | 3 | 3 | 0.80 | 0.20 | 21 | 1 | 0 | 0.98 | 0.02 |
| Overall | 53 | 12 | 18 | 23 | 0.40 | 0.60 | 28 | 12 | 13 | 0.64 | 0.36 | 48 | 5 | 0 | 0.94 | 0.06 |
| χ2 = 0.40 | χ2 = 0.98 | χ2 = 36.32 | ||||||||||||||
| Locus | Na | Ne | h | I | |
|---|---|---|---|---|---|
| CSN3 | CC | 2.0000 | 1.5392 | 0.3503 | 0.5349 |
| CT | 2.0000 | 1.8134 | 0.4486 | 0.6408 | |
| TT | 2.0000 | 1.9657 | 0.4913 | 0.6844 | |
| CSN2 | AA | 2.0000 | 1.9824 | 0.4956 | 0.6887 |
| AG | 2.0000 | 1.5879 | 0.3702 | 0.5571 | |
| GG | 2.0000 | 1.4902 | 0.3289 | 0.5107 | |
| CSN1S1 | GG | 2.0000 | 1.2061 | 0.1709 | 0.3125 |
| GT | 2.0000 | 1.2061 | 0.1709 | 0.3125 | |
| TT | 1.0000 | 1.0000 | 0.0000 | 0.0000 | |
| Mean | 1.8889 | 1.5323 | 0.3141 | 0.4713 | |
| Statistical error | 0.3333 | 0.3483 | 0.1684 | 0.2255 | |
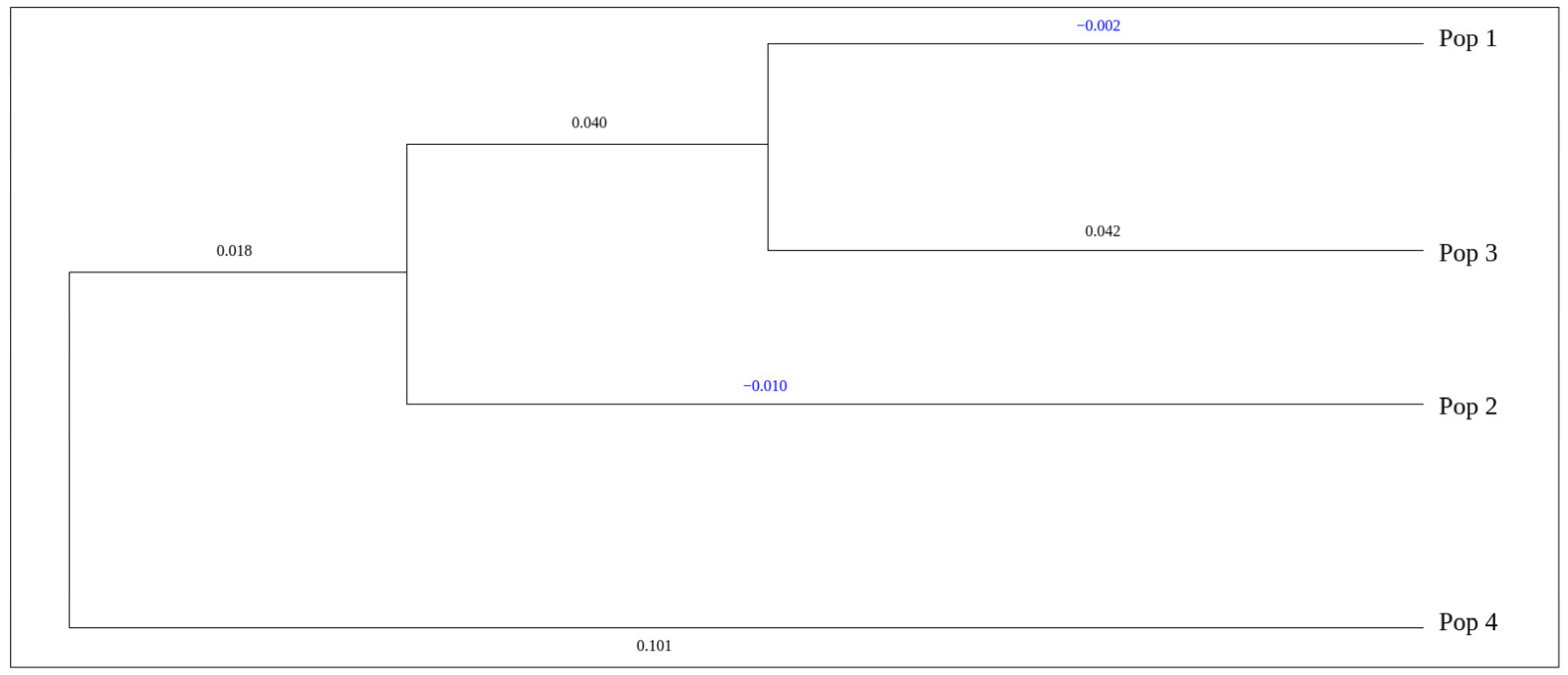
| Population | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| 1 | – | 0.9567 | 0.9603 | 0.8684 |
| 2 | 0.0443 | – | 0.9447 | 0.8969 |
| 3 | 0.0405 | 0.0569 | – | 0.8046 |
| 4 | 0.1411 | 0.1088 | 0.2174 | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amandykova, M.; Dossybayev, K.; Mussayeva, A.; Bekmanov, B.; Saitou, N. Comparative Analysis of the Polymorphism of the Casein Genes in Camels Bred in Kazakhstan. Diversity 2022, 14, 285. https://doi.org/10.3390/d14040285
Amandykova M, Dossybayev K, Mussayeva A, Bekmanov B, Saitou N. Comparative Analysis of the Polymorphism of the Casein Genes in Camels Bred in Kazakhstan. Diversity. 2022; 14(4):285. https://doi.org/10.3390/d14040285
Chicago/Turabian StyleAmandykova, Makpal, Kairat Dossybayev, Aizhan Mussayeva, Bakytzhan Bekmanov, and Naruya Saitou. 2022. "Comparative Analysis of the Polymorphism of the Casein Genes in Camels Bred in Kazakhstan" Diversity 14, no. 4: 285. https://doi.org/10.3390/d14040285
APA StyleAmandykova, M., Dossybayev, K., Mussayeva, A., Bekmanov, B., & Saitou, N. (2022). Comparative Analysis of the Polymorphism of the Casein Genes in Camels Bred in Kazakhstan. Diversity, 14(4), 285. https://doi.org/10.3390/d14040285