
First Record of Ophidonais serpentina (Müller, 1773) (Oligochaeta: Naididae) in China: The Occurrence or Absence of Needles Are Intraspecific Differences


Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling and Collection of Specimens
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Alignments and Phylogenetic Analysis
3. Results
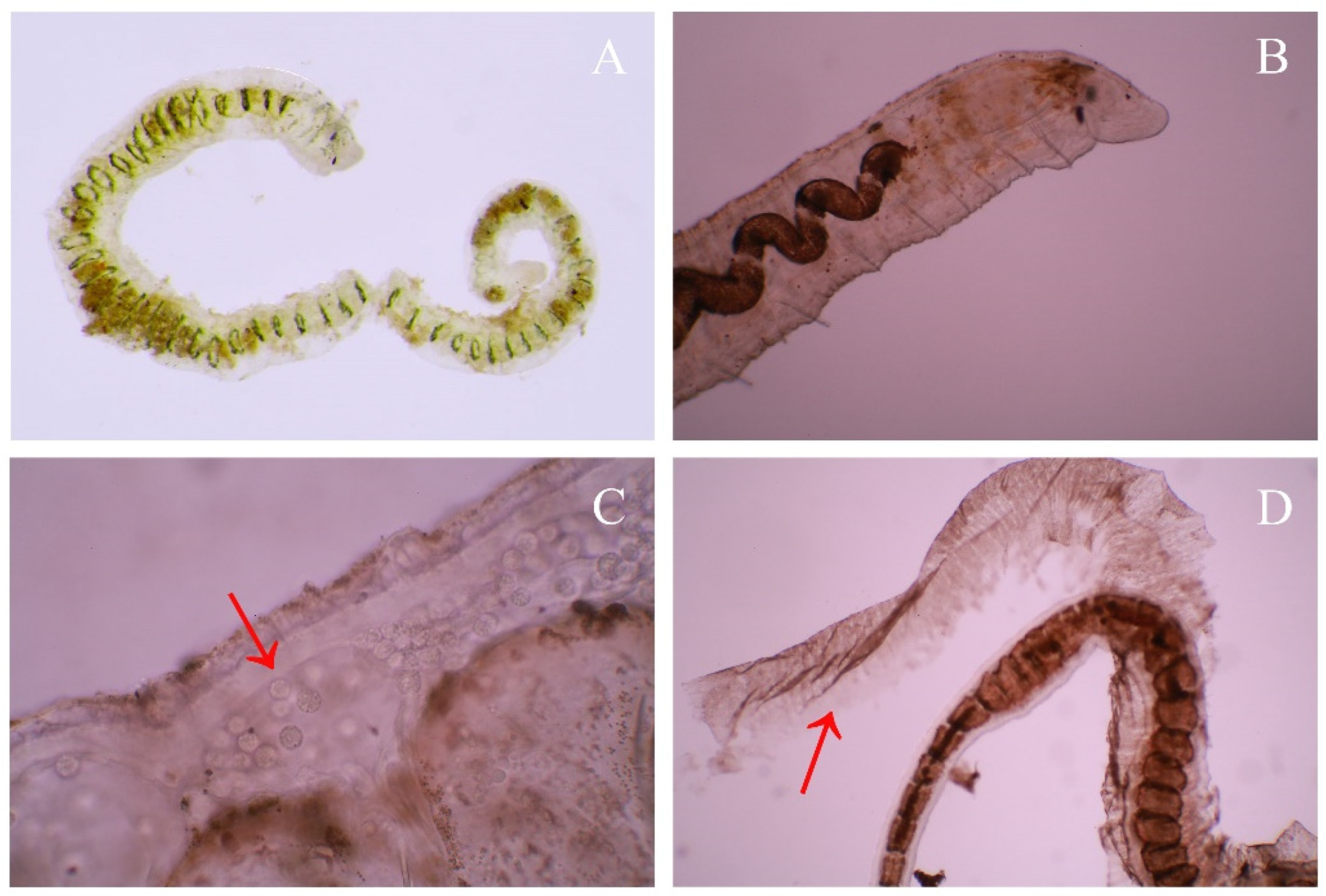
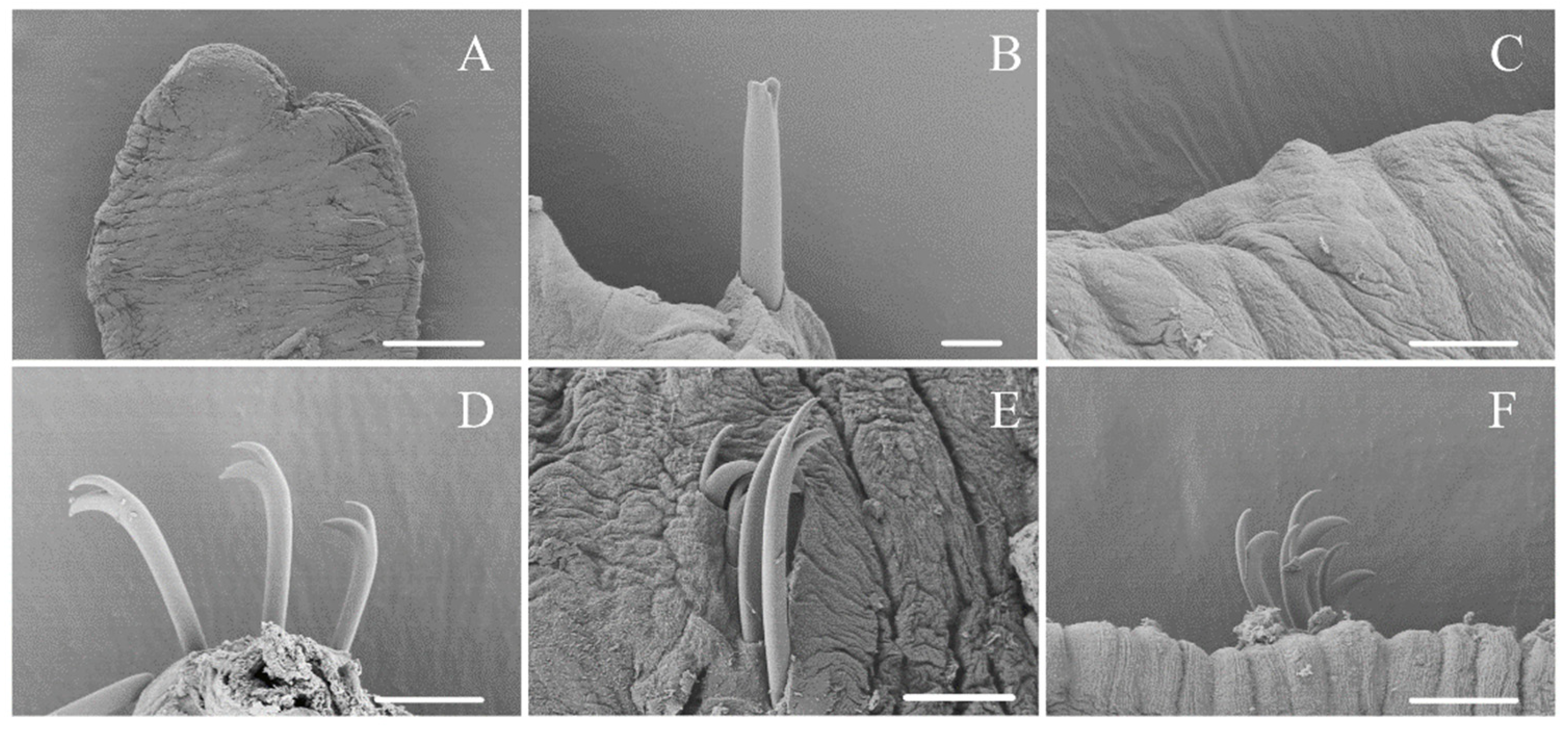
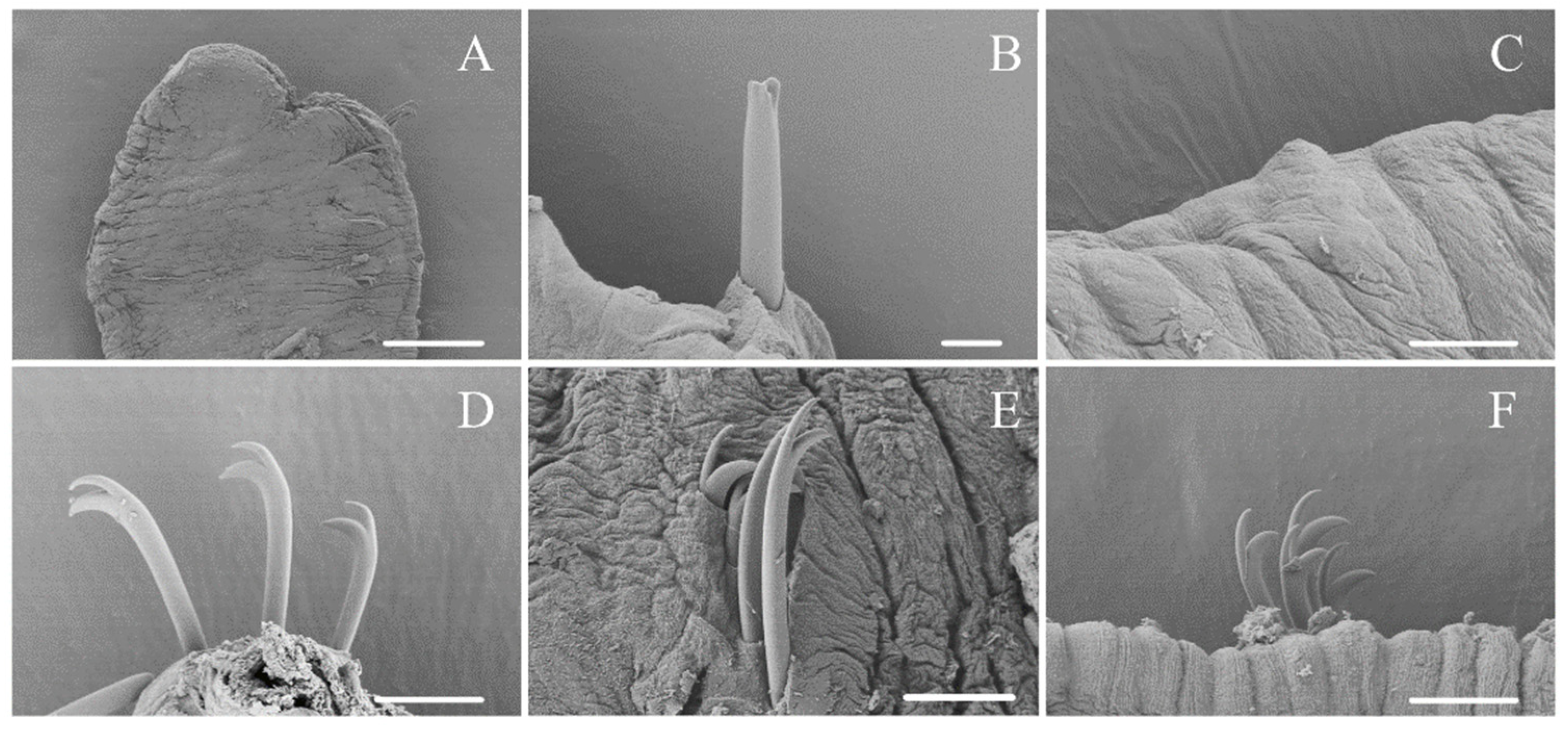
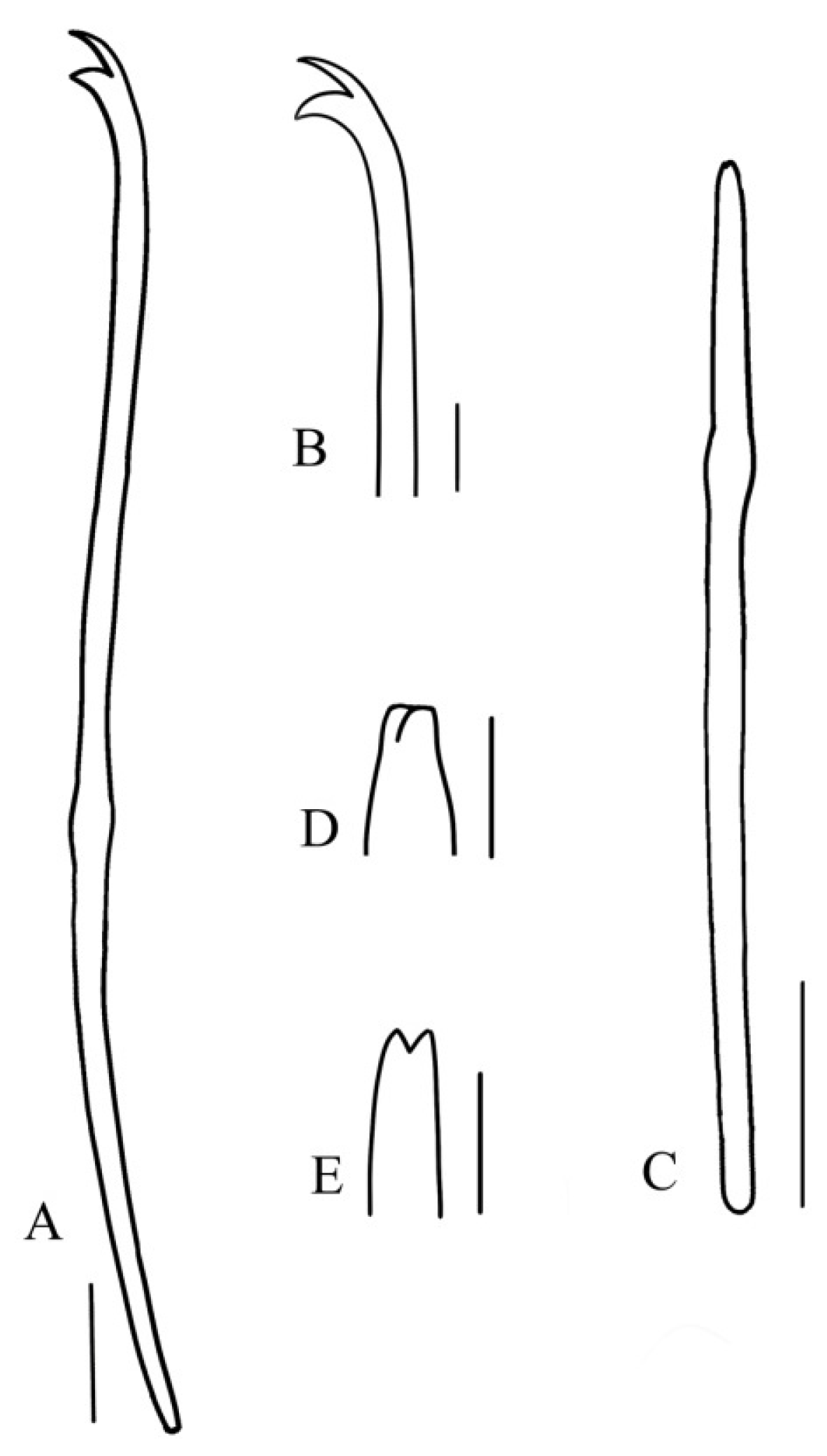
3.1. Taxonomy and Morphology
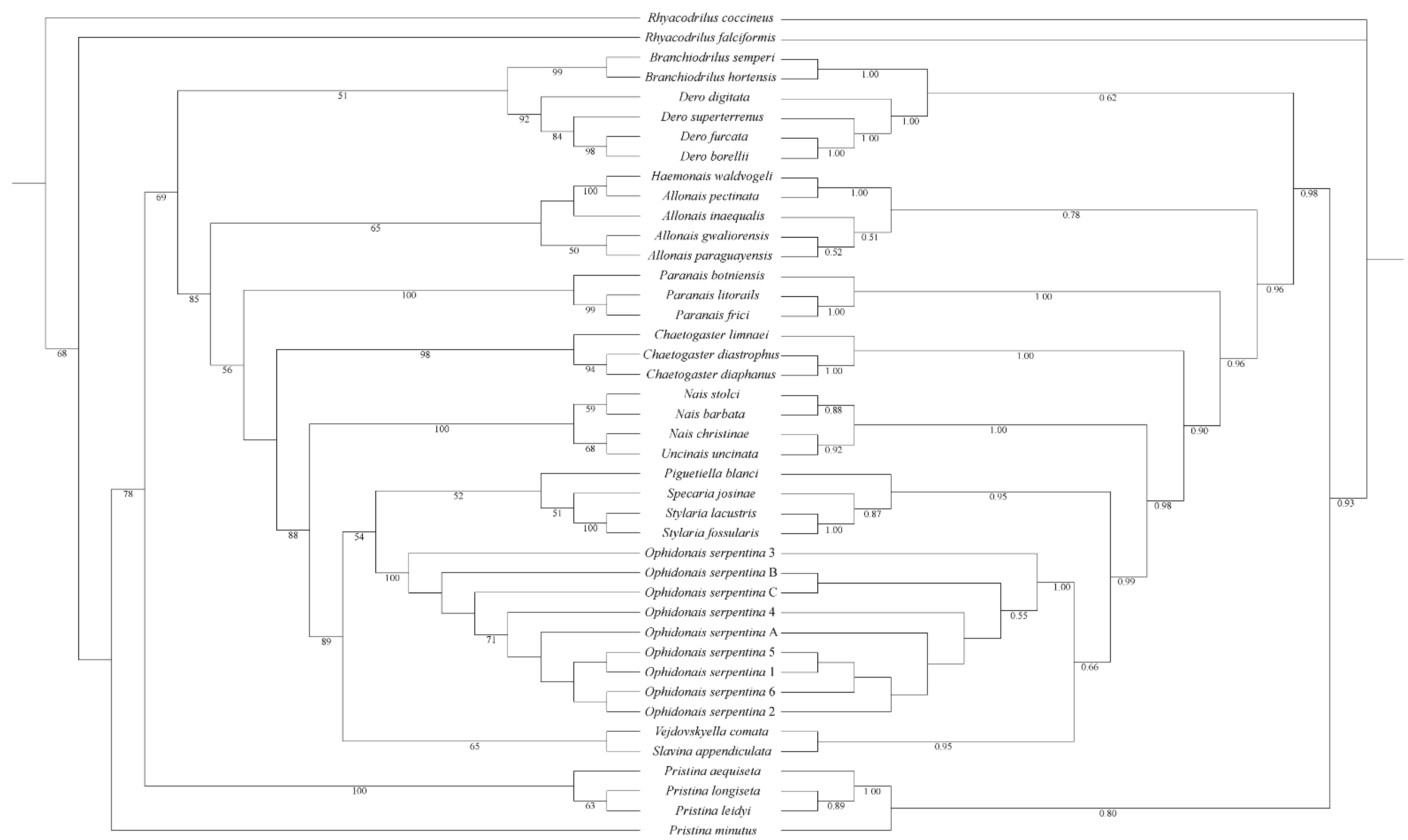
3.2. Phylogenetic Analyses
3.3. Pairwise Genetic Distances
4. Discussion
4.1. Morphological Characters of Ophidonais Serpentina
4.2. Intraspecies Analysis
4.3. Distribution and Habitat
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Müller, O.F. Vermium Terrestrium et Fluviatilium II. Hafniae Lipsiae 1773. Available online: https://www.biodiversitylibrary.org/item/50344 (accessed on 26 October 2021).
- Gervais, P. Note sur la disposition systématique des Annélides chétopodes de la famille des Nais. Bull. L’académie R. Des Sci. Belles-Lett. Brux. 1838, 5, 13–20. [Google Scholar]
- Erséus, C.; Envall, I.; De Wit, P.; Gustavsson, L.M. Molecular data reveal a tropical freshwater origin of Naidinae (Annelida, Clitellata, Naididae). Mol. Phylogenet. Evol. 2017, 115, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Timm, T. A guide to the freshwater Oligochaeta and Polychaeta of northern and central Europe. Lauterbornia 2009, 66, 1–235. [Google Scholar]
- Sperber, C. A taxonomical study of the Naididae. Zool. Bidr. Fran Upps. 1948, 28, 1–296. [Google Scholar]
- Kathman, D.D.; Brinkhurst, R.O. Guide to the Freshwater Oligochaetes of North America; Revised Version; Aquatic Resources Center: Nashville, TN, USA, 1999; p. 264. [Google Scholar]
- Ohtaka, A.; Iwakuma, T. Redescription of Ophidonais serpentina (Muller, 1773) (Naididae, Oligochaeta) from Lake Yunoko, Central Japan, with record of the oligochaete composition in the lake. Jpn. J. Limnol. 1993, 54, 251–259. [Google Scholar] [CrossRef]
- Envall, I.; Källersjö, M.; Erséus, C. Molecular evidence for the non-monophyletic status of Naidinae (Annelida, Clitellata, Tubificidae). Mol. Phylogenet. Evol. 2006, 40, 570–584. [Google Scholar] [CrossRef]
- Bely, A.E.; Sikes, J.M. Latent regeneration abilities persist following recent evolutionary loss in asexual annelids. Proc. Natl. Acad. Sci. USA 2010, 107, 1464–1469. [Google Scholar] [CrossRef] [Green Version]
- Bely, A.E.; Wray, G.A. Molecular phylogeny of naidid worms (Annelida: Clitellata) based on cytochrome oxidase I. Mol. Phylogenet. Evol. 2004, 30, 50–63. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, T.T.; Wang, H.Z.; Yu, P.; Erseus, C.; Cui, Y.D. Genetic and morphological analyses uncover a new record and a cryptic species in Allonais (Clitellata: Naididae). Biologia 2021, 76, 1705–1714. [Google Scholar] [CrossRef]
- Envall, I.; Gustavsson, L.M.; Erseus, C. Genetic and chaetal variation in Nais worms (Annelida, Clitellata, Naididae). Zool. J. Linn. Soc. 2012, 165, 495–520. [Google Scholar] [CrossRef] [Green Version]
- Martinsson, S.; Achurra, A.; Svensson, M.; Erseus, C. Integrative taxonomy of the freshwater worm Rhyacodrilus falciformis s.l. (C litellata: N aididae), with the description of a new species. Zool. Scr. 2013, 42, 612–622. [Google Scholar]
- Vivien, R.; Holzmann, M.; Werner, I.; Pawlowski, J.; Lafont, M.; Ferrari, B.J.D. Cytochrome c oxidase barcodes for aquatic oligochaete identification: Development of a Swiss reference database. PeerJ 2017, 5, e4122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, P.; Martinsson, S.; Wuillot, J.; Erséus, C. Integrative species delimitation and phylogeny of the branchiate worm Branchiodrilus (Clitellata, Naididae). Zool. Scr. 2018, 47, 727–742. [Google Scholar] [CrossRef]
- Vivien, R.; Wyler, S.; Lafont, M.; Pawlowski, J.J.P.O. Molecular barcoding of aquatic oligochaetes: Implications for biomonitoring. PLoS ONE 2015, 10, e0125485. [Google Scholar] [CrossRef] [PubMed]
- Sjölin, E.; Erséus, C.; Källersjö, M. Phylogeny of Tubificidae (Annelida, Clitellata) based on mitochondrial and nuclear sequence data. Mol. Phylogenet. Evol. 2005, 35, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Erseus, C.; Rota, E.; Matamoros, L.; De Wit, P. Molecular phylogeny of Enchytraeidae (Annelida, Clitellata). Mol. Phylogenet. Evol. 2010, 57, 849–858. [Google Scholar] [CrossRef]
- Mejlon, E.; De Wit, P.; Matamoros, L.; Erséus, C. DNA-based phylogeny of the marine genus Heterodrilus (Annelida, Clitellata, Naididae). J. Zool. Syst. Evol. Res. 2015, 53, 194–199. [Google Scholar] [CrossRef]
- Palumbi, S.R.; Martin, A.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; Version 2.0; University of Hawaii: Honolulu, HI, USA, 1991; Volume 45. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Liu, Y.; Erséus, C. New specific primers for amplification of the Internal Transcribed Spacer region in Clitellata (Annelida). Ecol. Evol. 2017, 7, 10421–10439. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.; Rouse, G.; Hutchings, P.; Colgan, D. Assessing the usefulness of histone H3, U2 snRNA and 28S rDNA in analyses of polychaete relationships. Aust. J. Zool. 1999, 47, 499–516. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Ohtaka, A.; Nishino, M. Studies on the aquatic oligochaete fauna in Lake Biwa, central Japan. II. Records and taxonomic remarks of nine species. Hydrobiologia 1999, 406, 33–47. [Google Scholar] [CrossRef]
- Yanez, E.; Cuadrado, S.; Martinez-Ansemil, E. External sense organs in freshwater oligochaetes (Annelida, Clitellata) revealed by scanning electron microscopy. J. Morphol. 2006, 267, 198–207. [Google Scholar] [CrossRef]
- Spencer, D.R.; Wisseman, R.E. Some new records of Naididae and Tubificidae (Annelida, Oligochaeta) from Washingtons. Great Basin Nat. 1993, 53, 395–401. [Google Scholar]
- Conn, D.B. Invading the invaders: Infestation of zebra mussels by native parasites in the St. Lawrence River. In Proceedings of the Fourth International Zebra Mussel Conference, Madison, WI, USA, 7–10 March 1994; p. 8. [Google Scholar]
- Jablonska, A.; Pesic, V. Five species of aquatic oligochaetes new to Iran with an updated checklist. Oceanol. Hydrobiol. Stud. 2014, 43, 100–105. [Google Scholar] [CrossRef]
- Ardalan, A.A.; Mooraki, N.; Sadeghi, M.S. Occurrence of Ophidonais serpentina in Potamon persicum from Jajrood River, Iran. Iran. J. Fish. Sci. 2011, 10, 177–180. [Google Scholar]
- George, A.D.I.; Abowei, J.F.N.; Daka, E.R. Benthic Macro Invertebrate Fauna and Physico-chemical Parameters in Okpoka Creek Sediments, Niger Delta, Nigeria. Int. J. Anim. Vet. Adv. 2009, 1, 59–65. [Google Scholar]
- Sinsch, U.; Dehling, M.; Scheid, P.; Balczun, C. A new African species of parasitic Dero (Annelida, Clitellata, Naididae) in the urinary tract of reed frogs. Parasitol. Res. 2019, 118, 3359–3370. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.M.; Childress, J.N.; Iakovidis, T.J.; Langford, G.J. Elucidating the Life History and Ecological Aspects of Allodero hylae (Annelida: Clitellata: Naididae), A Parasitic Oligochaete of Invasive Cuban Tree Frogs in Florida. J. Parasitol. 2015, 101, 275–281. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No Needles Group | With Needles Group | |
|---|---|---|
| Specimens | 53 | 11 |
| Body length | 4–30 mm | 4–30 mm |
| N segments | 27–84 | 38–87 |
| First segment of needles | none | variable, beginning at segment between XIV to XXXXV, lacking in most of the segments |
| Ventral chaetae of II segment | length 150–178 μm, width 5 μm | length 150–178 μm, width 5 μm |
| Ventral chaetae of the remaining segment | length 95–125 μm, width 5 μm | length 95–125 μm, width 5 μm |
| Papillae | dorsal side, scattered, the beginning is variable most appear at the posterior segment; some individuals have abundant papillae, begin at X, then each segment has one papilla | few, only appear at those segments have no needles |
| Species | Collection Site or Source; Collector | 16S | COI | ITS2 | H3 |
|---|---|---|---|---|---|
| Ingroup | |||||
| Allonais gwaliorensis | Moat at Angor Wat, Cambodia; A. Ohtaka | KY633311 [3] | KY633391 [3] | KY633363 [3] | - |
| Allonais inaequalis | Pacaya-Samiria Reserve, Peruvian Amazon, Peru; D. Shain | DQ459952 [8] | KY633390 [3] | - | - |
| Allonais paraguayensis | Ward’s Natural Science (sold as Stylaria); A. E. Bely | GQ355399 [9] | AF534828 [10] | - | - |
| Allonais pectinata | East Lake Scenic Area of Wuhan, China; W. jiang | MN914711 [11] | MN935212 [11] | - | - |
| Branchiodrilus hortensis | Lake Tehang, Central Kalimantan, Indonesia; A. Ohtaka | KY633312 [3] | KY633393 [3] | KY633378 [3] | - |
| Branchiodrilus semperi | Pond in Bogor Botanical Garden, Bogor, West Java, Indonesia; A. Ohtaka | KY633315 [3] | KY633396 [3] | KY633379 [3] | - |
| Chaetogaster diaphanus | Lake Lången, Vårgårda, Sweden; C. Erséus | DQ459956 [8] | JQ519897 [12] | KY633380 [3] | - |
| Chaetogaster limnaei | Sacramento, CA, USA; A. Bely/J. Sikes | GQ355405 [9] | KF952355 [13] | - | - |
| Chaetogaster diastrophus | Hällekis, Götene, Sweden; C. Erséus | JQ424952 [12] | LT904771 [14] | - | - |
| Dero borellii | Experimental biofilter, Manchester Metropolitan Univ.,UK; M. Dempsey | KY633324 [3] | KY633385 [3] | KY633364 [3] | - |
| Dero digitata | Lake Lången, Vårgårda, Sweden; C. Erséus | DQ459954 [8] | KY633397 [3] | KY633381 [3] | MH744978 [15] |
| Dero furcata | Ditch, Fengshan, Kaohsiung, Taiwan; C.-R. Li | KY633325 [3] | KY633388 [3] | - | MH744979 [15] |
| Dero superterrenus | Eastern Melrose, Alachua Co., Fl., USA; D. Strom & M. Wetzel | KY633326 [3] | KY633389 [3] | - | - |
| Nais barbata | Lake Låttern, Vingåker, Sweden; C. Erséus | JQ424993 [12] | JQ519861 [12] | - | - |
| Nais christinae | Upper Kenai River, Moose beach, AK, USA; L. Arsan & S. Atkinson | JQ424969 [12] | JQ519824 [12] | - | - |
| Nais stolci | Charlottenlund, Ystad, Sweden; C. Erséus | JQ425026 [12] | JQ519894 [12] | - | - |
| Ophidonais serpentina 1 | Xinkai River of Shaoxing, China; T. T. Zhou & J. F. Yu | OM033727 | OM033378 | OM033228 | SRR17607623 |
| Ophidonais serpentina 2 | Xinkai River of Shaoxing, China; T. T. Zhou & J. F. Yu | OM033728 | OM033379 | OM033229 | SRR17607622 |
| Ophidonais serpentina 3 | Xinkai River of Shaoxing, China; T. T. Zhou & J. F. Yu | OM033729 | OM033380 | OM033230 | SRR17607621 |
| Ophidonais serpentina 4 | Xinkai River of Shaoxing, China; T. T. Zhou & J. F. Yu | OM033730 | OM033381 | OM033231 | SRR17607620 |
| Ophidonais serpentina 5 | Xinkai River of Shaoxing, China; T. T. Zhou & J. F. Yu | OM033731 | OM033382 | OM033232 | SRR17607619 |
| Ophidonais serpentina 6 | Xinkai River of Shaoxing, China; T. T. Zhou & J. F. Yu | OM033732 | OM033383 | OM033233 | SRR17607618 |
| Ophidonais serpentina A | Kungsbackaån River, Kungsbacka, Sweden; S. Kvist &M. Lindström | KY633327 [3] | KY633398 [3] | LN810239 [16] | - |
| Ophidonais serpentina B | San Francisco Creek, San Mateo Co., California, USA; S. Fend | DQ459939 [8] | LT903820 [14] | KY633367 [3] | - |
| Ophidonais serpentina C | Wildcat Creek, Richmond, CA, USA; A. Bely/J. Sikes | GQ355411 [9] | KY633398 [3] | KY633366 [3] | - |
| Piguetiella blanci | Lake Jäsen, Orsa, Sweden; M. Lindström; C. Erséus | KY633320 [3] | KY633402 [3] | KY633370 [3] | - |
| Paranais botniensis | Viken, Höganäs, Sweden; C. Erséus | KY633316 [3] | KY633399 [3] | KY633368 [3] | - |
| Paranais frici | Rappahannock River (brackish), Middlesex Co., VA, USA; S. Kvist | KY633318 [3] | KY633415 [3] | KY633369 [3] | - |
| Paranais litorails | Rhode River, Edgewater, MD, USA; A. Bely/J. Sikes | KY633319 [3] | KY633401 [3] | - | - |
| Slavina appendiculata | Lången Lake, near Alingsås, Västergötland, Sweden; C. Erséus | AY885582 [17] | KY633405 [3] | KY633371 [3] | - |
| Stylaria fossularia | Kampong Chhnang, Lake Tonle Sap, Cambodia; A. Ohtaka | KY633322 [3] | KY633408 [3] | KY633374 [3] | - |
| Specaria josinae | Lake Lången, Vårgårda, Sweden; C. Erséus | KY633321 [3] | KY633407 [3] | KY633372 [3] | - |
| Stylaria lacustris | Lake Lången, Vårgårda, Sweden; C. Erséus | DQ459947 [8] | KY633409 [3] | KY633375 [3] | - |
| Uncinais uncinata | Lången Lake, near Alingsås, Västergötland, Sweden; C.Erséus | DQ459942 [8] | KY633410 [3] | KY633376 [3] | - |
| Vejodovskyella comata | Lången Lake, near Alingsås, Västergötland, Sweden; C. Erséus | AY885584 [17] | KY633411 [3] | KY633377 [3] | - |
| Pristina aequiseta | Paint Branch, College Park, MD, USA; A. Bely/J. Sikes | GQ355415 [9] | GQ355374 [9] | - | - |
| Pristina leidyi | Carolina Biological Supply (sold as Stylaria). | GQ355416 [9] | AF534853 [10] | - | - |
| Pristina longiseta | Lizard Island (freshwater), Great Barrier Reef, Queensland, Australia; C. Erséus | GU901850 [18] | GU902108 [18] | - | - |
| Pristina minutus | Tjärnö, Strömstad, Sweden; C. Erséus | DQ459958 [8] | KJ753865 [19] | - | - |
| Haemonais waldvogeli | Naolihe wuxinghu River of Helongjiang, China; T. T. Zhou | OM264280 | MW888774 | MW885234 | - |
| Outgroup | |||||
| Rhyacodrilus coccineus | Lake Lången, Vårgårda, Sweden; C. Erséus | DQ459931 [8] | GU902110 [18] | - | KF267971 [13] |
| Rhyacodrilus falciformis | Vitärtskällan Spring, Kappelshamn, Gotland, Sweden; C. Erséus | DQ459938 [8] | KF267935 [13] | - | KF267970 [13] |
| Gene | Primer | Sequence 5′-3′ | The Program of PCR | Reference |
|---|---|---|---|---|
| 16S | 16SAR-L | CGCCTGTTTATCAAAAACAT | 30 s at 98 °C; 10 s at 98 °C; 45 s at 60 °C; 35 cycles of 1 min at 72 °C; 2 min at 72 °C. | Palumbi et al., 1991 [20] |
| 16SBRH | CCGGTCTGAACTCAGATCACGT | Palumbi et al., 1991 | ||
| COI | LCO1490 | GGTCAACAAATCATAAAGATATTGG | 30 s at 98 °C; 10 s at 98 °C; 45 s at 45 °C; 35 cycles of 45 s at 72 °C; 3 min at 72 °C. | Folmer et al., 1994 [21] |
| HCO2198 | TAAACTTCAGGGTGACCAAAAAATCA | Folmer et al., 1994 | ||
| COI-E | TATACTTCTGGGTGTCCGAAGAATCA | Bely and Wray 2004 [10] | ||
| ITS2 | 606F | GTCGATGAAGAGCGCAGCCA | 30 s at 98 °C; 10 s at 98 °C; 45 s at 55 °C; 35 cycles of 45 s at 72 °C; 3 min at 72 °C. | Liu, Erséus 2017 [22] |
| 1082R | TTAGTTTCTTTTCCTCCGCTT | Liu, Erséus 2017 | ||
| H3 | H3F | ATGGCTCGTACCAAGCAGACVGC | 5 min at 95 °C; 30 s at 95 °C; 30 s at 50 °C; 35 cycles of 90 s at 72 °C; 8 min at 72 °C | Brown et al., 1999 [23] |
| H3R | ATATCCTTRGGCATKATRGTGAC | Brown et al., 1999 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Slavina appendiculata | 0.024 | 0.029 | 0.033 | 0.035 | 0.040 | 0.033 | 0.035 | 0.032 | 0.028 | 0.032 | 0.032 | 0.032 | 0.034 | 0.031 | |
| 2. Vejdovskyella comata | 0.131 | 0.021 | 0.031 | 0.031 | 0.033 | 0.026 | 0.028 | 0.026 | 0.025 | 0.026 | 0.026 | 0.030 | 0.027 | 0.029 | |
| 3. Piguetiella blanci | 0.150 | 0.131 | 0.019 | 0.023 | 0.023 | 0.029 | 0.031 | 0.029 | 0.023 | 0.029 | 0.029 | 0.027 | 0.029 | 0.029 | |
| 4. Specaria josinae | 0.142 | 0.142 | 0.114 | 0.031 | 0.029 | 0.042 | 0.044 | 0.042 | 0.038 | 0.042 | 0.042 | 0.041 | 0.042 | 0.042 | |
| 5. Stylaria lacustris | 0.157 | 0.134 | 0.126 | 0.143 | 0.017 | 0.031 | 0.033 | 0.031 | 0.030 | 0.031 | 0.031 | 0.032 | 0.031 | 0.033 | |
| 6. Stylaria fossularis | 0.158 | 0.132 | 0.138 | 0.134 | 0.114 | 0.044 | 0.044 | 0.044 | 0.040 | 0.044 | 0.044 | 0.046 | 0.044 | 0.046 | |
| 7. Ophidonais serpentina 1 | 0.132 | 0.137 | 0.120 | 0.128 | 0.131 | 0.147 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| 8. Ophidonais serpentina 2 | 0.132 | 0.137 | 0.120 | 0.128 | 0.131 | 0.147 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| 9. Ophidonais serpentina 3 | 0.132 | 0.137 | 0.120 | 0.128 | 0.131 | 0.147 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| 10. Ophidonais serpentina 4 | 0.132 | 0.137 | 0.120 | 0.128 | 0.131 | 0.147 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| 11. Ophidonais serpentina 5 | 0.137 | 0.143 | 0.124 | 0.131 | 0.134 | 0.151 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| 12. Ophidonais serpentina 6 | 0.139 | 0.144 | 0.126 | 0.131 | 0.136 | 0.147 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| 13. Ophidonais serpentina A | 0.131 | 0.136 | 0.120 | 0.128 | 0.130 | 0.148 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| 14. Ophidonais serpentina B | 0.133 | 0.138 | 0.122 | 0.127 | 0.130 | 0.147 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| 15. Ophidonais serpentina C | 0.131 | 0.136 | 0.120 | 0.128 | 0.130 | 0.148 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Information | Asia | Europe | America |
|---|---|---|---|
| Body length (mm) | 4–30 | 6–36 | 6 |
| Segment | 27–126 | 23–51 | 62 |
| Eyes | present | present | present or absent |
| Ventral chaetae in | |||
| length (μm) | 150–208 | 152–179 | 130–174 |
| no./bundle | 2–5 | 2–6 | 2–4 |
| teeth | distal teeth shorter and thinner | distal teeth shorter and thinner | equal, or distal teeth slightly longer |
| Ventral chaetae after | |||
| length (μm) | 152–168 | 128–158 | 100–130 |
| no./bundle | 2–4 | 2–6 | 3–4 |
| teeth | distal teeth shorter and thinner in anterior, equal in posterior | distal teeth shorter than proximal teeth | equal, or distal teeth slightly shorter |
| Needle | |||
| length (μm) | 140–172 | 150–168 | 130 |
| staring segment | VI, or absent, not clear in the specimens in China | VI | VI |
| no./bundle | 1 | 1 | 1 |
| no. of teeth | 1–2, sometimes 3 | 1–2 | saw-toothed |
| Penial chaetae | two teeth closed, and the end swell and sag | the end swell and sag | immature |
| Habitat | shallow lake, the surface of hydrophytes | freshwater | common in rivers in North America, or live as parasites |
| Reference | Ohtaka and Iwakuma 1993; this study | Sperber 1948 | Spencer et al., 1993 [33]; Conn et al., 1994 [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Zhou, T.; Wang, H.; Cui, Y. First Record of Ophidonais serpentina (Müller, 1773) (Oligochaeta: Naididae) in China: The Occurrence or Absence of Needles Are Intraspecific Differences. Diversity 2022, 14, 265. https://doi.org/10.3390/d14040265
Yu J, Zhou T, Wang H, Cui Y. First Record of Ophidonais serpentina (Müller, 1773) (Oligochaeta: Naididae) in China: The Occurrence or Absence of Needles Are Intraspecific Differences. Diversity. 2022; 14(4):265. https://doi.org/10.3390/d14040265
Chicago/Turabian StyleYu, Jiefeng, Tingting Zhou, Hongzhu Wang, and Yongde Cui. 2022. "First Record of Ophidonais serpentina (Müller, 1773) (Oligochaeta: Naididae) in China: The Occurrence or Absence of Needles Are Intraspecific Differences" Diversity 14, no. 4: 265. https://doi.org/10.3390/d14040265
APA StyleYu, J., Zhou, T., Wang, H., & Cui, Y. (2022). First Record of Ophidonais serpentina (Müller, 1773) (Oligochaeta: Naididae) in China: The Occurrence or Absence of Needles Are Intraspecific Differences. Diversity, 14(4), 265. https://doi.org/10.3390/d14040265