
A New Species of the Genus Niphargus Schiödte, 1849 (Crustacea: Amphipoda: Niphargidae) from Groundwater Habitats of the Tarkhankut Upland, Crimean Peninsula

Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Sampling
2.2. Morphological Studies
2.3. Molecular and Phylogenetic Study
2.4. Stable Isotope Analysis
3. Results
3.1. Biological Assemblage of the Studied Wells
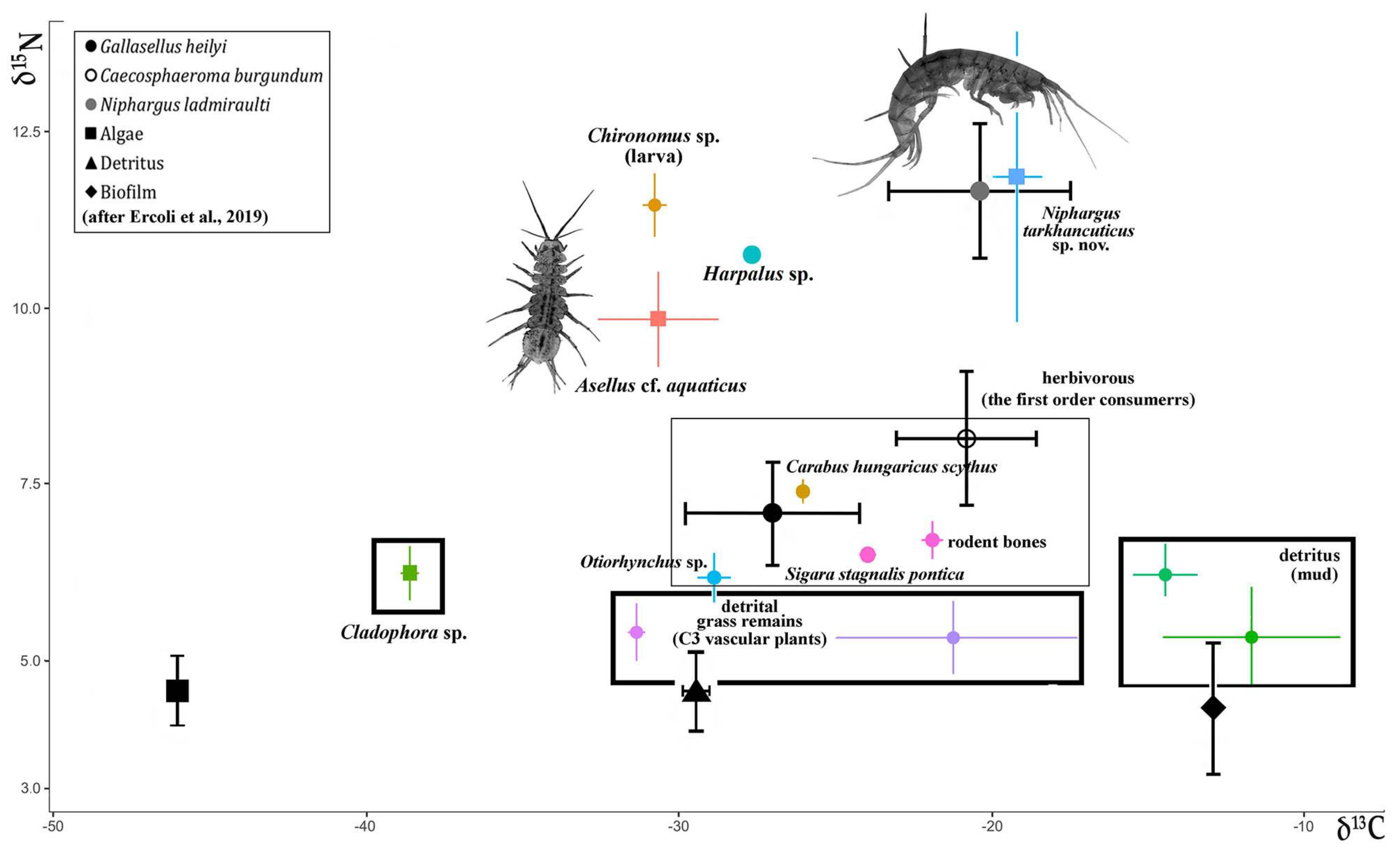
3.2. Trophic Relationships with the Studied Wells
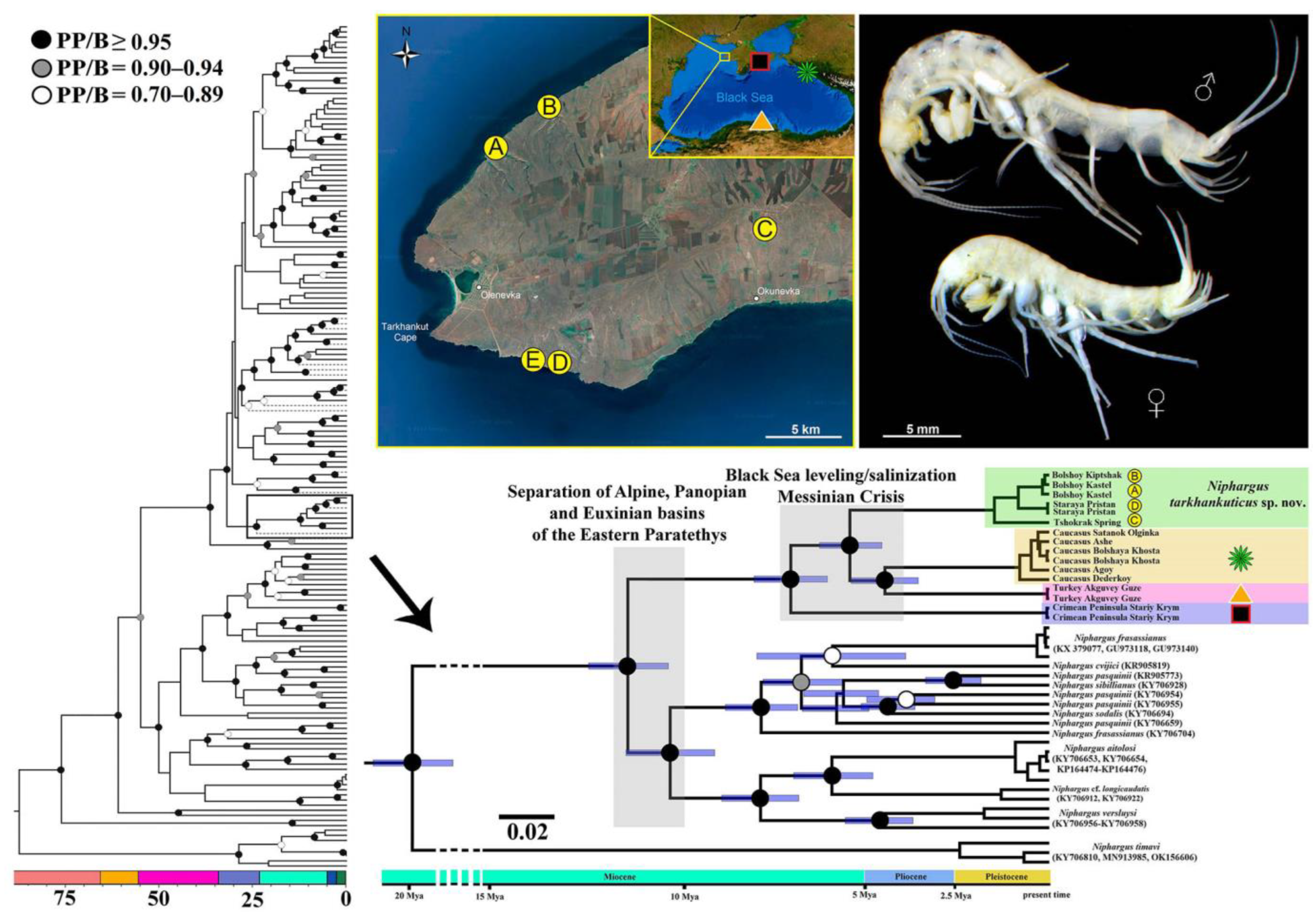
3.3. Phylogenetic Study of Newly Discovered Niphargus


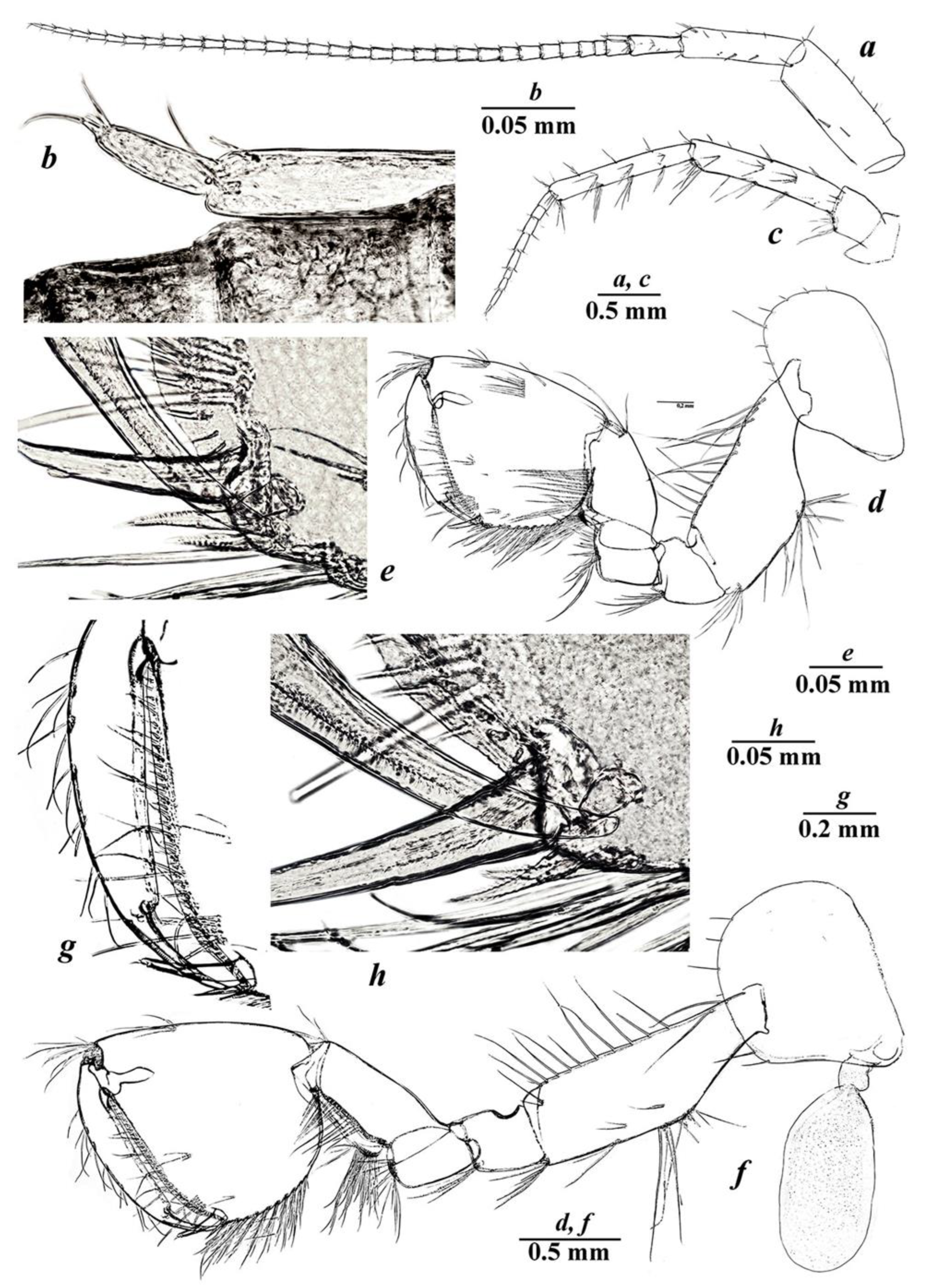
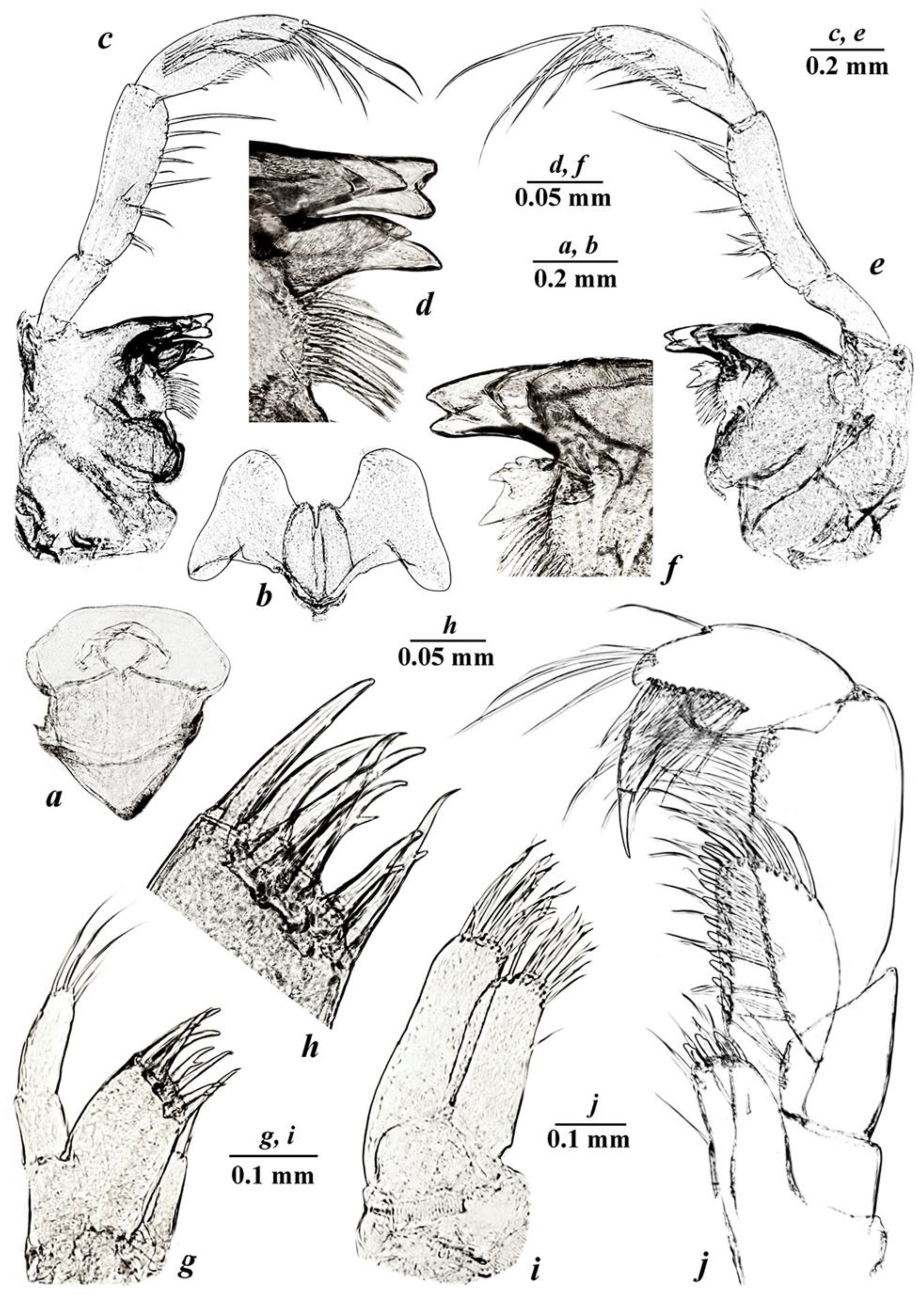
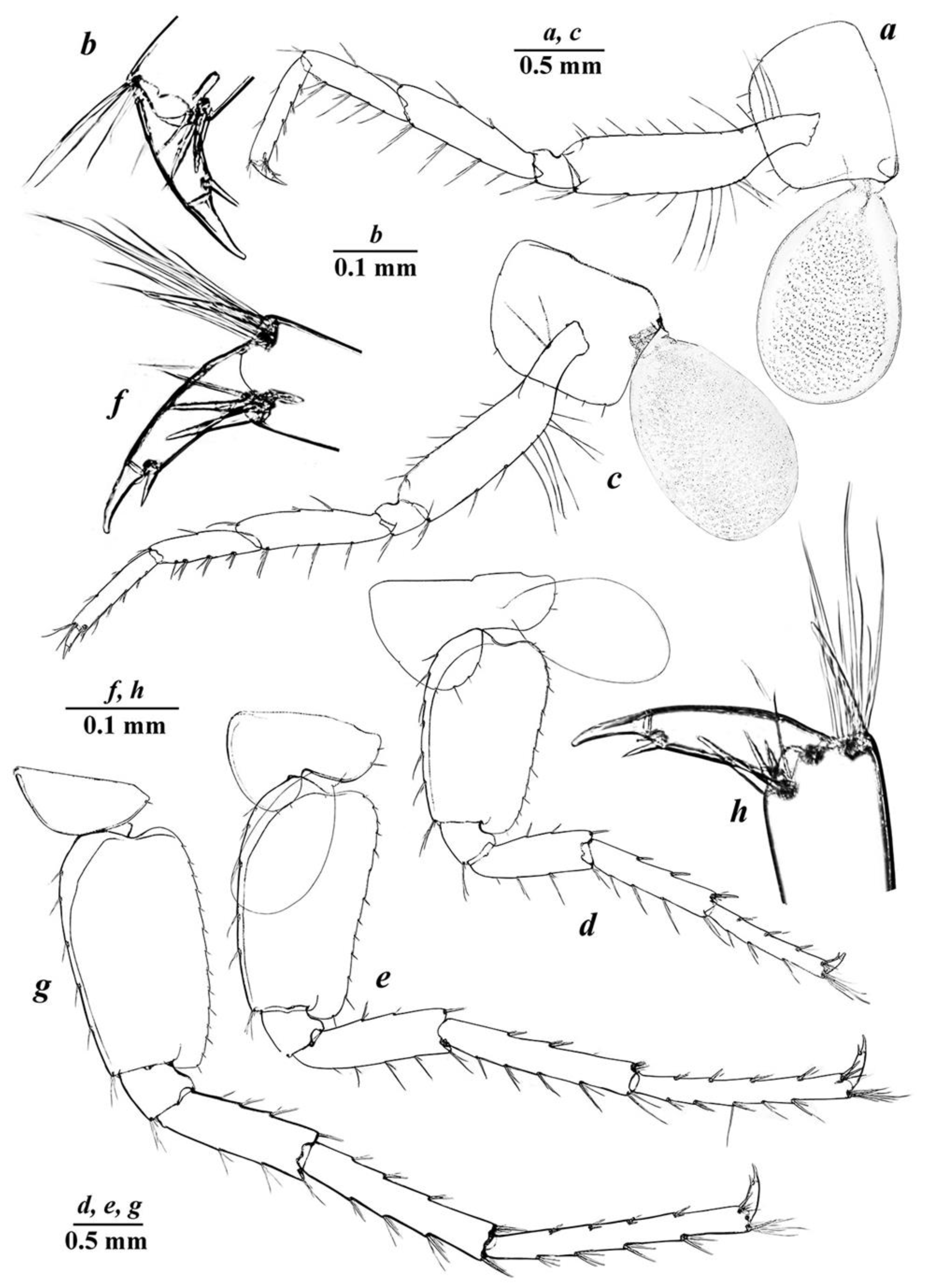
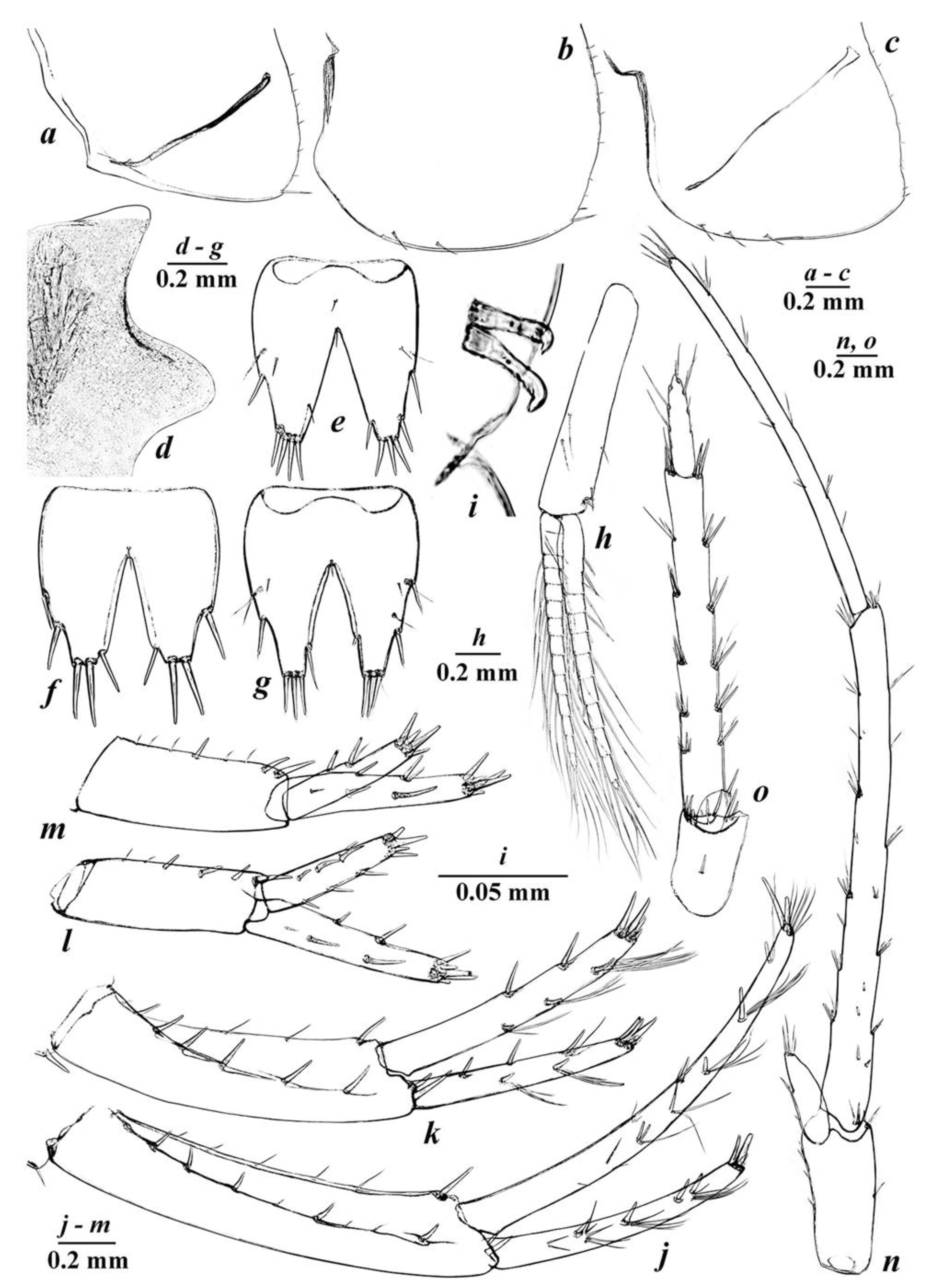
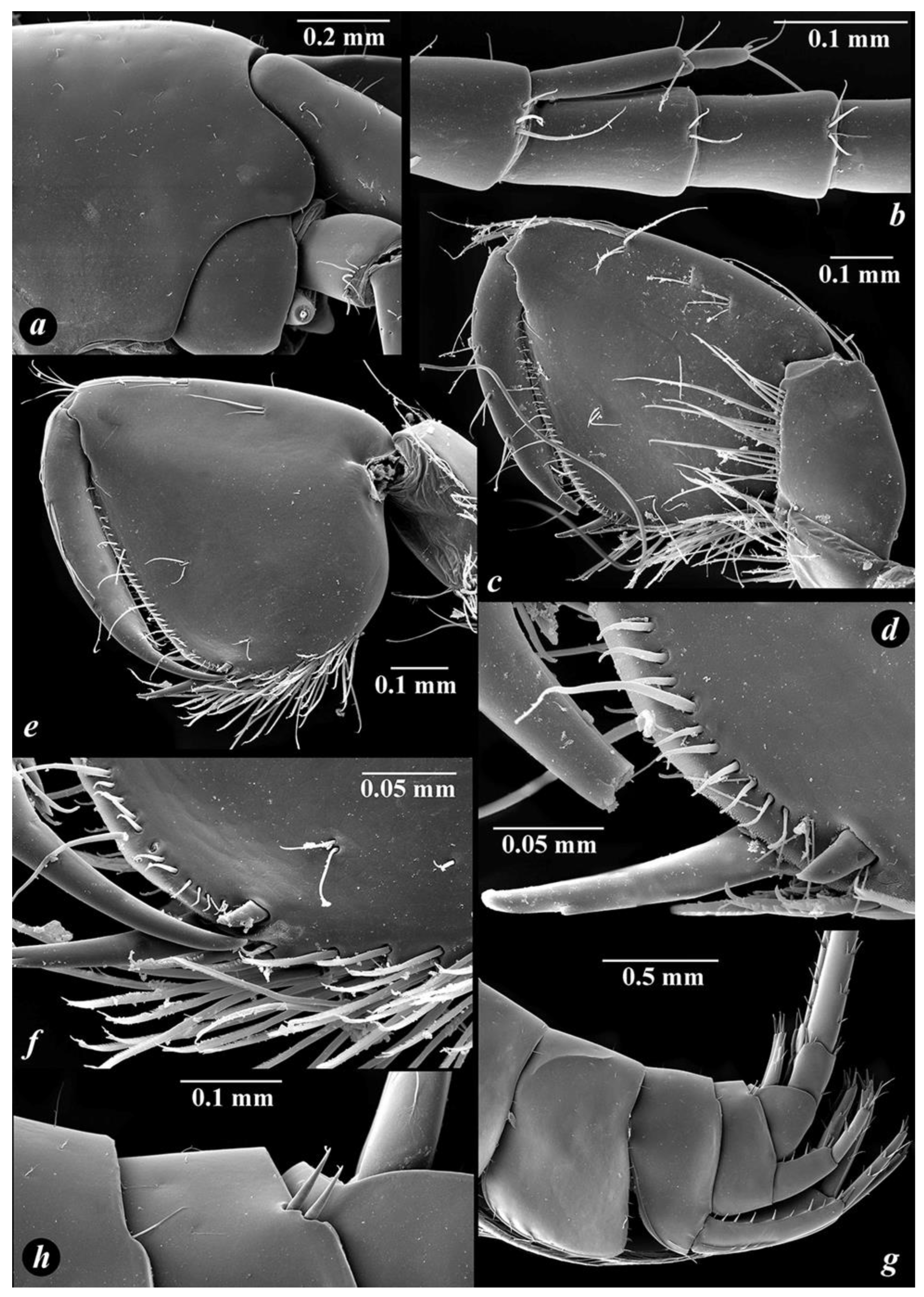
3.4. Taxonomic Part
- Phylum Arthropoda von Siebold, 1848
- Class Malacostraca Latreille, 1802
- Order Amphipoda Latreille, 1816
- Family Niphargidae Bousfield, 1977
- Genus Niphargus Schiödte, 1849
- Niphargus tarkhankuticus sp. nov.
- urn:lsid:zoobank.org:pub:878BE10A-27B7-45B2-A9A6-0362E12FA552





4. Discussion

Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muratov, V.V. A Brief Sketch of the Geological Structure of the Crimean Peninsula; State Scientific and Technical Publishing House of Literature on the Geology and Conservation of Mineral Resources: Moscow, Russia, 1960; pp. 1–208. (In Russian) [Google Scholar]
- Kostrytsky, M.E. Physical and geographical description. In Geology of the USSR. Vol. 8. Crimea. Part 1. Geological Description; Nedra: Moscow, Russia, 1969; pp. 18–31. (In Russian) [Google Scholar]
- Ryan, W.B.F.; Pitman, W.C.; Major, C.O.; Shimkus, K.; Moskalenko, V.; Jones, G.A.; Dimitrov, P.; Gorür, N.; Sakinç, M. An abrupt drowning of the Black Sea shelf. Mar. Geol. 1997, 138, 119–126. [Google Scholar] [CrossRef]
- Ryan, W.B.; Major, C.O.; Lericolais, G.; Goldstein, S.L. Catastrophic flooding of the Black Sea. Ann. Rev. Earth Planet. Sci. 2003, 31, 525–554. [Google Scholar] [CrossRef]
- Dublyansky, V.N.; Dublyanskaya, G.N. Karst of Crimea and Its Problems; Crimean Academy of Sciences: Simferopol, Russia, 1996; pp. 1–88. (In Russian) [Google Scholar]
- Vakhrushev, B.A. Regionalization of karst of the Crimean Peninsula. Speleol. Karstol. 2009, 3, 39–46. (In Russian) [Google Scholar]
- Ved, I.P. Climate Atlas of Crimea; Tavria Plus: Simferopol, Ukraine, 2000; p. 120. (In Russian) [Google Scholar]
- Lvova, E.V. Plain Crimea. Geological Structure, Hydrogeology, Nature Conservation; Naukova Dumka: Kyiv, Russia, 1978; p. 188. (In Russian) [Google Scholar]
- Turbanov, I.S.; Palatov, D.M.; Golovatch, S.I. The present state of the art of biospeleology in Russia and the countries of the former Soviet Union: A review of the cave (endogean) invertebrate fauna. 1. Introduction—Crustacea. Zool. Zh. 2016, 95, 1136–1159. (In Russian) [Google Scholar] [CrossRef]
- Turbanov, I.S.; Palatov, D.M.; Golovatch, S.I. The present state of the art of biospeleology in Russia and the countries of the former Soviet Union: A review of the cave (endogean) invertebrate fauna. 2. Arachnida—Acknowledgments. Zool. Zh. 2016, 95, 1283–1304. (In Russian) [Google Scholar] [CrossRef]
- Turbanov, I.S.; Palatov, D.M.; Golovatch, S.I. The present state of the art of biospeleology in Russia and the countries of the former Soviet Union: A review of the cave (endogean) invertebrate fauna. 3. References. Zool. Zh. 2016, 95, 1399–1426. (In Russian) [Google Scholar] [CrossRef]
- Martynov, A.B. Zur Kenntnis der Amphipoden der Krim. Zool. Jahrb. Abt. Syst. Ökol. Geog. Tiere. 1931, 60, 573–606. [Google Scholar]
- Birštein, J.A. The subterranean amphipods of the Crimea. Bull. Mosc. Soc. Nat. Biol. Ser. 1961, 66, 126–144. (In Russian) [Google Scholar]
- Birštein, J.A. A third species of the underground genus Niphargus (Crustacea Amphipoda) from the Crimea. Bull. Mosc. Soc. Nat. Biol. Ser. 1964, 69, 119–121. (In Russian) [Google Scholar]
- Vargovich, R. Crustaceans (Crustacea). In Cave fauna of Ukraine; Zagorodniuk, I., Ed.; National Science and Natural History Museum of the National Academy of Sciences of Ukraine: Kiev, Ukraine, 2004; pp. 47–52. (In Ukrainian) [Google Scholar]
- Zagorodniuk, I.; Vargovich, R. Checklist of cave fauna of Ukraine. In Cave Fauna of Ukraine; Zagorodniuk, I., Ed.; National Science and Natural History Museum of the National Academy of Sciences of Ukraine: Kiev, Ukraine, 2004; pp. 191–216. (In Ukrainian) [Google Scholar]
- Horton, T.; Lowry, J.; De Broyer, C.; Bellan-Santini, D.; Coleman, C.O.; Corbari, L.; Costello, M.J.; Daneliya, M.; Dauvin, J.-C.; Fišer, C.; et al. World Amphipoda Database. Niphargus Schiödte. 1849. Available online: https://www.marinespecies.org/amphipoda/aphia.php?p=taxdetails&id=545672 (accessed on 3 October 2022).
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Copilaş-Ciocianu, D.; Fišer, C.; Borza, P.; Petrusek, A. Is subterranean lifestyle reversible? Independent and recent large-scale dispersal into surface waters by two species of the groundwater amphipod genus Niphargus. Mol. Phylogen. Evol. 2018, 119, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Fišer, C.; Alther, R.; Zakšek, V.; Borko, Š.; Fuchs, A. Altermatt, F. Translating Niphargus barcodes from Switzerland into taxonomy with a description of two new species (Amphipoda, Niphargidae). Zookeys 2018, 760, 113–141. [Google Scholar] [CrossRef] [PubMed]
- Fišer, C.; Delić, T.; Luštrik, R.; Zagmajster, M.; Altermatt, F. Niches within a niche: Ecological differentiation of subterranean amphipods across Europe’s interstitial waters. Ecography 2019, 42, 1212–1223. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. Beast 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef]
- McInerney, C.E.; Maurice, L.; Robertson, A.L.; Knight, L.R.F.D.; Arnscheidt, J.; Venditti, C.; Dooley, J.S.G.; Mathers, T.; Matthijs, S.; Eriksson, K.; et al. The ancient Britons: Groundwater fauna survived extreme climate change over tens of millions of years across NW Europe. Mol. Ecol. 2014, 23, 1153–1166. [Google Scholar] [CrossRef]
- Lefébure, T.; Douady, C.J.; Gouy, M.; Trontelj, P.; Briolay, J.; Gibert, J. Phylogeography of a subterranean amphipod reveals cryptic diversity and dynamic evolution in extreme environments. Mol. Ecol. 2006, 15, 1797–1806. [Google Scholar] [CrossRef]
- Lefébure, T.; Douady, C.J.; Malard, F.; Gibert, J. Testing dispersal and cryptic diversity in a widely distributed groundwater amphipod (Niphargus rhenorhodanensis). Mol. Phylogen. Evol. 2007, 42, 676–686. [Google Scholar] [CrossRef]
- Copilaş-Ciocianu, D.; Petrusek, A. The southwestern Carpathians as an ancient centre of diversity of freshwater gammarid amphipods: Insights from the Gammarus fossarum species complex. Mol. Ecol. 2015, 24, 3980–3992. [Google Scholar] [CrossRef]
- Copilaș-Ciocianu, D.; Fišer, C.; Borza, P.; Balázs, G.; Angyal, D.; Petrusek, A. Low intraspecific genetic divergence and weak niche differentiation despite wide ranges and extensive sympatry in two epigean Niphargus species (Crustacea: Amphipoda). Zool. J. Linn. Soc. 2017, 181, 485–499. [Google Scholar] [CrossRef]
- Marin, I.; Krylenko, S.; Palatov, D. The Caucasian relicts: A new species of the genus Niphargus (Crustacea: Amphipoda: Niphargidae) from the Gelendzhik–Tuapse area of the Russian southwestern Caucasus. Zootaxa 2021, 4963, 483–504. [Google Scholar] [CrossRef] [PubMed]
- Karaman, G.S. Redescription of subterranean gammaridean species Niphargus longicaudatus (Costa 1851) based on topotypic material (Contribution to the Knowledge of the Amphipoda 161). Frag. Balc. 1986, 13, 27–42. [Google Scholar]
- Karaman, G.S. New species of the family Niphargidae and new localities of some other subterranean Gammaridean species from Yugoslavia (Contribution to the knowledge of the Amphipoda 166). Glas. Repub. Zav. Zaš. Prirod. Muz. Titograd. 1989, 19, 15–32. [Google Scholar]
- Karaman, S. Novi amfipodi podzemne faune Grčke. Rad Jugos. Akad. Znan. Umjet. 1950, 280, 106–114. [Google Scholar]
- Karaman, G.S. On the endemic subterranean amphipod Niphargus versluysi S. Karaman, 1950 (Fam. Niphargidae) in Greece (Contribution to the Knowledge of the Amphipoda 297). Biol. Serb. 2017, 39, 52–67. [Google Scholar] [CrossRef]
- Ntakis, A.; Anastasiadou, C.; Zakšek, V.; Fišer, C. Phylogeny and biogeography of three new species of Niphargus (Crustacea: Amphipoda) from Greece. Zool. Anz. 2015, 255, 32–46. [Google Scholar] [CrossRef]
- Karaman, G.S.; Borowsky, B.; Dattagupta, S. Two new species of the genus Niphargus Schiödte, 1849 (Amphipoda, fam. Niphargidae) from the Frasassi cave system in Central Italy. Zootaxa 2010, 2439, 35–52. [Google Scholar] [CrossRef]
- Karaman, S. Dve nove vrste podzemnih amfipoda Popova Polja u Hercegovini. O nekim amfipodima—izopodima Balkana i o njihovoj sistematici [CLXIII]. In Posebna Izdanja. Odjeljenje Prirodno-Matematičkih Nauka; Srpska Akademija Nauka: Belgrade, Yugoslavia, 1950; pp. 101–118. [Google Scholar]
- Vigna Taglianti, A. Sulla presenza di Niphargus del gruppo elegans nell’Italia appenninica (Amphipoda, Gammaridae). Arch. Zool. Ital. 1966, 51, 863–876. [Google Scholar]
- Karaman, G.S. Contribution to the knowledge of the Amphipoda. Description of several new Niphargus species (Gammaridea, Niphargidae) from Southern Europe. Poljop. I Sumar. Titograd. 1984, 30, 39–64. [Google Scholar]
- Karaman, S. Die Niphargiden des Slovenischen Karstes, Istriens sowie des benachb. Italiens. Acta Mus. Maced. Sci. Nat. 1954, 2, 159–180. [Google Scholar]
- Karaman, G.S. Contribution to the knowledge of the Amphipoda 145. Two subterranean species of the family Niphargidae, Niphargus lunaris, n. sp. and N. timavi S. Kar. 1954. Glas. Repub. Zav. Zaš. Prirod. Muz. Titograd. 1984, 17, 29–46. [Google Scholar]
- Marin, I.N. The Quaternary speciation in the Caucasus: A new cryptic species of stygobiotic amphipod of the genus Niphargus (Crustacea: Amphipoda: Niphargidae) from the Kumistavi (Prometheus) Cave, Western Georgia. Arth. Sel. 2020, 29, 419–432. [Google Scholar] [CrossRef]
- Marin, I.N.; Krylenko, S.V.; Palatov, D.M. Euxinian relict amphipods of the Eastern Paratethys in the subterranean fauna of coastal habitats of the Northern Black Sea region. Inverteb. Zool. 2021, 18, 247–320. [Google Scholar] [CrossRef]
- Monchenko, V.I.; Sopova, E.M. New cyclops (Crustacea, Cyclopidae) for the fauna of the Ukraine. Vest. Zool. 1984, 5, 10–16. (In Russian) [Google Scholar]
- Birštein, J.A. Freshwater Isopods (Asellota). Fauna of USSR. Crustacea. Vol. 7. No. 5; Izdatelstvo Akademii Nauk SSSR: Moskva, Russia; Leningrad, Russia, 1951; pp. 1–143. (In Russian) [Google Scholar]
- Borko, Š.; Trontelj, P.; Seehausen, O.; Moškrič, A.; Fišer, C. A subterranean adaptive radiation of amphipods in Europe. Nat. Commun. 2022, 12, 3688. [Google Scholar] [CrossRef]
- Borko, S.; Altermatt, F.; Zagmajster, M.; Fišer, C. A hotspot of groundwater amphipod diversity on a crossroad of evolutionary radiations. Divers. Distrib. 2022, 00, 1–13. [Google Scholar] [CrossRef]
- Coleman, C.O.; Myers, A.A. New Amphipoda from Baltic amber. Polsk. Arch. Hydrobiol. 2000, 47, 457–464. [Google Scholar]
- Jażdżewski, K.; Kupryjanowicz, J. One more fossil niphargid (Malacostraca: Amphipoda) from Baltic amber. J. Crust. Biol. 2010, 30, 413–416. [Google Scholar] [CrossRef][Green Version]
- Palatov, D.M.; Marin, I.N. Epigean (pond-dwelling) species of the genus Niphargus Schiödte, 1849 (Crustacea: Amphipoda: Niphargidae) from the coastal plains of the Black and Azov seas of the north- and south-western Caucasus. Invert. Zool. 2021, 18, 105–151. [Google Scholar] [CrossRef]
- Marin, I.; Palatov, D. A new species of the genus Niphargus (Crustacea: Amphipoda: Niphargidae) from the southwestern part of the North Caucasus. Zool. Mid. East 2019, 65, 1–11. [Google Scholar] [CrossRef]
- Marin, I.; Palatov, D. Cryptic refugee on the northern slope of the Greater Caucasian Ridge: Discovery of Niphargus (Crustacea: Amphipoda: Niphargidae) in the North Ossetia-Alania, North Caucasus, separated from its relatives in the late Miocene. Zool. Anz. 2021, 292, 163–183. [Google Scholar] [CrossRef]
- Yudin, V.V. Geology of Crimea. Photoatlas; Publishing House Printing House Arial: Simferopol, Crimea, 2017; pp. 1–160. (In Russian) [Google Scholar]
- Sket, B. Prispevek k poznavanju naših amfipodov. Biol. Vest. 1958, 6, 67–75. [Google Scholar]
- Ginet, R. Écologie, éthologie et biologie de Niphargus (Amphipodes Gammaridés hypogés). Ann. Spéléo. 1960, 15, 127–237. [Google Scholar]
- Mathieu, J.; Debouzie, D.; Martin, D. Influence des conditions hydrologiques sur la dynamique dʼune population phreatique de Niphargus rhenorhodanensis (Amphipode souterrain). Vie et Milieu 1987, 37, 193–200. [Google Scholar]
- Rumm, P. Zur Ökologie des Grundwasseramphipoden Niphargus fontans (Bate 1859). In Deutsche Gesellschaft für Limnologie e. V. Tagungbericht; Deutsche Gesellschaft für Limnologie e. V.: Karlsruhe, Germany, 2000; pp. 384–388. [Google Scholar]
- Fišer, C.; Kovačec, Ž.; Pustovrh, M.; Trontelj, P. The role of predation in the diet of Niphargus (Amphipoda: Niphargidae. Speleobiol. Not. 2010, 2, 4–6. [Google Scholar]
- Knight, L.R.; Johns, T. Auto-ecological studies on Niphargus aquilex (Schiödte, 1855) and Niphargus glenniei (Spooner, 1952) (Crustacea: Amphipoda: Niphargidae). Cave Karst Sci. 2015, 42, 63–77. [Google Scholar]
- Magniez, G. Observations sur la biologie de Stenasellus virei (Crustacea Isopoda Asellota) des eaux souterraines. Int. J. Speleo. 1975, 7, 79–228. [Google Scholar] [CrossRef]
- Gibert, J.; Deharveng, L. Subterranean ecosystems: A truncated functional biodiversity. BioScience 2002, 52, 473–481. [Google Scholar] [CrossRef]
- Hutchins, B.T.; Schwartz, B.F.; Nowlin, W.H. Morphological and trophic specialization in a subterranean amphipod assemblage. Freshwat. Biol. 2014, 59, 2447–2461. [Google Scholar] [CrossRef]
- Francois, C.M.; Mermillod-Blondin, F.; Malard, F.; Fourel, F.; Lecuyer, C.; Douady, C.J.; Simon, L. Trophic ecology of groundwater species reveals specialization in a low-productivity environment. Funct. Ecol. 2016, 30, 262–273. [Google Scholar] [CrossRef]
- Ercoli, F.; Lefebvre, F.; Delangle, M.; Godé, N.; Caillon, M.; Raimon, R.; Souty-Grosset, C. Differing trophic niches of three French stygobionts and their implications for conservation of endemic stygofauna. Aqua. Cons. Mar. Freshwat. Ecosys. 2019, 29, 2193–2203. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | δ13C | δ15N | Mean ± SD (δ13C) | Mean ± SD (δ15N) | Number |
|---|---|---|---|---|---|
| Niphargus tarkhankuticus sp. nov. | −18.63 | 12.86 | −19.31 ± 0.78 | 11.86 ± 2.05 | 5 |
| −18.36 | 11.72 | ||||
| −20.31 | 10.65 | ||||
| −19.16 | 14.71 | ||||
| −19.61 | 9.36 | ||||
| Asellus cf. aquaticus | −27.65 | 10.63 | −30.65 ± 1.92 | 9.84 ± 0.67 | 5 |
| −30.26 | 10.36 | ||||
| −31.01 | 9.51 | ||||
| −32.84 | 8.94 | ||||
| −31.48 | 9.78 |
| Species (or Locality for Undescribed Species) | p-Distances ± S.E. |
|---|---|
| Caucasus: Satanok, Ashe, Bolshaya Khosta, Agoy, Dederkoy (n = 6) | 0.135 ± 0.017 |
| Turkey: Akguvey Guze (n = 2) | 0.142 ± 0.019 |
| Crimean Peninsula: Staryi Krym (n = 2) | 0.172 ± 0.020 |
| N. frasassinus (n = 3) | 0.193 ± 0.022 |
| N. cf. longicaudatus (n = 2) | 0.199 ± 0.019 |
| N. aitolosi (n = 3) | 0.200 ± 0.021 |
| N. pasquini (n = 4) | 0.204 ± 0.021 |
| N. cvijici (n = 1) | 0.213 ± 0.024 |
| N. versuysi (n = 3) | 0.233 ± 0.021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marin, I.N.; Turbanov, I.S.; Prokopov, G.A.; Palatov, D.M. A New Species of the Genus Niphargus Schiödte, 1849 (Crustacea: Amphipoda: Niphargidae) from Groundwater Habitats of the Tarkhankut Upland, Crimean Peninsula. Diversity 2022, 14, 1010. https://doi.org/10.3390/d14121010
Marin IN, Turbanov IS, Prokopov GA, Palatov DM. A New Species of the Genus Niphargus Schiödte, 1849 (Crustacea: Amphipoda: Niphargidae) from Groundwater Habitats of the Tarkhankut Upland, Crimean Peninsula. Diversity. 2022; 14(12):1010. https://doi.org/10.3390/d14121010
Chicago/Turabian StyleMarin, Ivan N., Ilya S. Turbanov, Gregoryi A. Prokopov, and Dmitry M. Palatov. 2022. "A New Species of the Genus Niphargus Schiödte, 1849 (Crustacea: Amphipoda: Niphargidae) from Groundwater Habitats of the Tarkhankut Upland, Crimean Peninsula" Diversity 14, no. 12: 1010. https://doi.org/10.3390/d14121010
APA StyleMarin, I. N., Turbanov, I. S., Prokopov, G. A., & Palatov, D. M. (2022). A New Species of the Genus Niphargus Schiödte, 1849 (Crustacea: Amphipoda: Niphargidae) from Groundwater Habitats of the Tarkhankut Upland, Crimean Peninsula. Diversity, 14(12), 1010. https://doi.org/10.3390/d14121010