1. Introduction
Gastropods (snails and slugs) occur in saltwater, freshwater, and terrestrial environments. They are the most diverse class in the phylum Mollusca, containing about 476 families with 65,000–80,000 living species. The class is thought to be second only to the insects in overall species number [
1].
The fertilized egg of gastropods hatches directly into a spherical or pear-shaped free-swimming larval stage called the trochophore, carrying a ring of cilia [
2]. The ciliary girdle then expands into large, heavily ciliated lobes, giving rise to a larval stage called the veliger. Later the larva undergoes torsion, a 180° twisting that brings the posterior part of the body to an anterior position behind the head. Torsion is unique to the gastropods.
The veliger has a shell (secreted by the dorsal shell gland), a foot, and a velum, which is a lobed, ciliated structure used for swimming and feeding [
2]. In most cases, the veliger eventually sinks to the seabed, loses its velum, and completes its metamorphosis into a juvenile or adult with typical snail-like morphology (a heteroplanktonic life cycle). In some cases, the adult is also planktonic, making the life cycle holoplanktonic
Analyzing the development of gastropod larvae can be done by rearing and documenting individuals, but here we show that a simpler and more efficient method is to gather individuals and identify them by sequencing their DNA barcode, which is a portion of the cytochrome
c oxidase I (COI or COX1) gene, found in mitochondrial DNA [
3,
4]. We have previously used this approach to document the life cycles of cnidarians [
5] and crustaceans [
6].
The class Gastropoda includes both shelled and unshelled species. The marine shelled species include whelks, abalone, conches, periwinkles, turbans, cowries, limpets, chitons, and others. In most cases the one-piece shell is coiled in both larval and adult stages, but in the limpets it is coiled only in the larval stage. Gastropods are distinguished by their asymmetrical anatomy, in which most of the organs are more developed on one side of the body than the other. They typically have a distinct head carrying two or four sensory tentacles bearing eyes. Their ventral foot is the basis for the name gastropod (“stomach-foot”). Many species have an operculum which allows closing of the shell. In the following we use the taxonomy established by the World Register of Marine Species [
7].
2. Materials and Methods
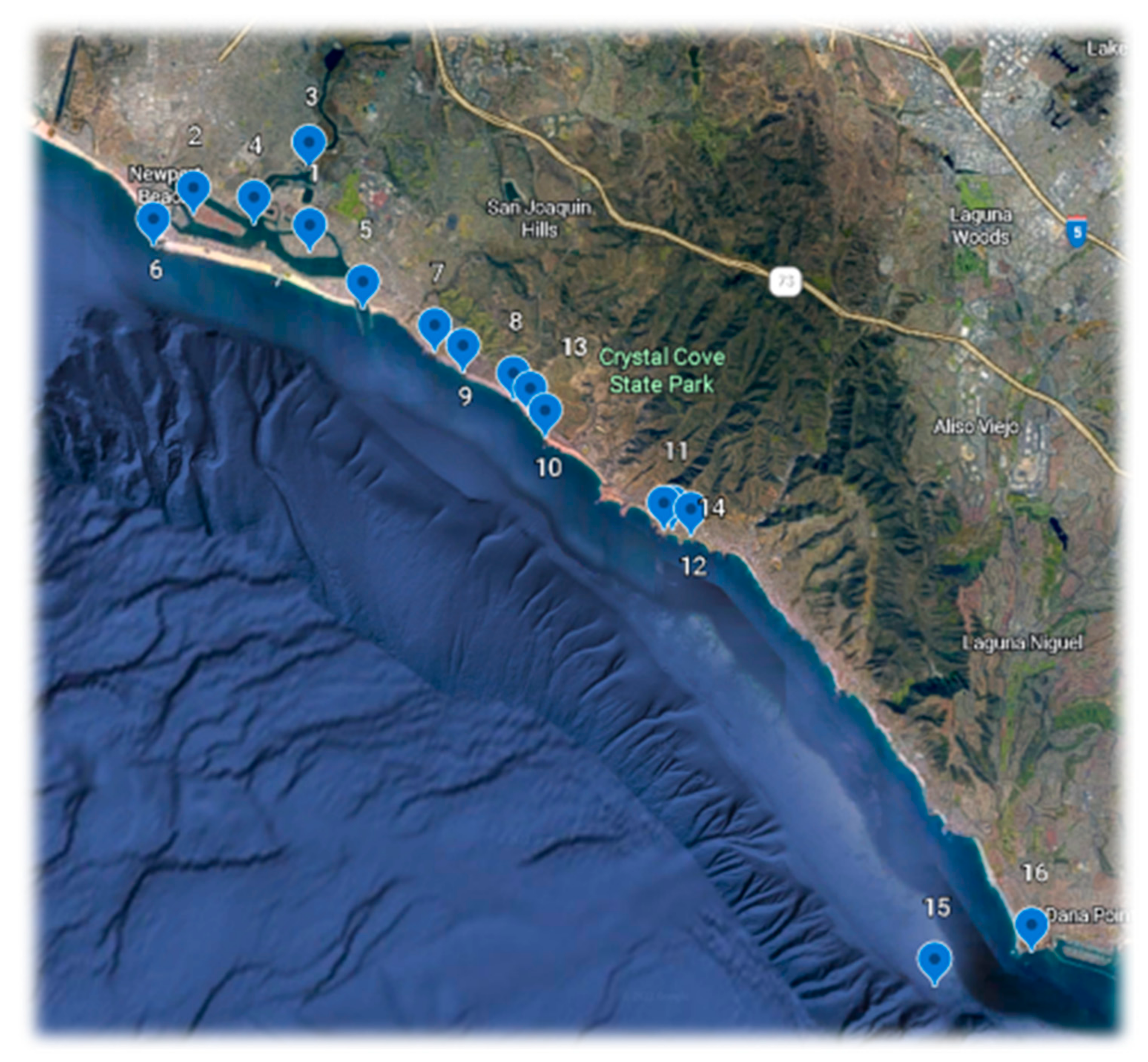
Zooplankton was collected under Scientific Collecting Permit SC-12162 from the California Department of Fish and Wildlife. Collections were made from 16 sites between Newport Beach and Dana Point, Orange County California, as well as one off Santa Barbara and two from Baja California (
Table 1 and
Figure 1).
Field work. In the following account, only the localities outside of Orange County are identified specifically.
Shore-based collections were made with a 150 μm mesh net (aperture 30 cm) attached to a rope, with a 50 mL collection tube at the base. They were made from public docks using repeated horizontal sweeps near the surface and diagonal sweeps down to about 5 m. depth. About 5–10 sweeps of a total of about 35 m. usually yielded sufficient specimens, but no attempt was made to monitor collections quantitatively.
Ocean collection #15 was made with a 250 μm. mesh net attached to a 35 m. rope. The net (aperture 30 cm) was towed behind the vessel, just below the surface, for a period of 7 min at the slowest possible speed. Deployment and retrieval extended the total tow period to 10 min.
Laboratory analysis. Plankton collections were brought to the laboratory at the University of California, Irvine and examined under a dissecting microscope with lateral light and a dark background. Each specimen of interest was removed using a Pasteur Pipette, transferred to a depression slide, and recorded by video microscopy using a Zeiss microscope with a dark-field condenser, fitted with a phototube attached to a Nikon D5100 single-lens reflex camera. The most informative frames were taken from the videos and used in the figures for this paper. Each plankton specimen was preserved in 90% ethanol in a well of a 96-well microplate.
Each adult specimen was photographed in place, removed physically from its location, brought to the laboratory, and examined under the dissecting microscope. Using the microwave method [
8] live adults were quickly heated sufficient to kill the animals and firm up their tissues; the steam from inside the shell forcing the body from the shell for easy removal. Multiple tissue samples were then removed using dissecting tools and transferred to the microplates. If available, typically three individuals per species were sacrificed, though occasionally more were used due to our inability to positively identify some species using field characteristics. Many of the species that also have a benthic stage in the life cycle have already been listed by the Southern California Association of Marine Invertebrate Taxonomists (SCAMIT) [
9].
Filled plates were sent to the Canadian Centre for DNA Barcoding at the University of Guelph, 50 Stone Road East, Guelph, ON, N1G2W1, Canada for DNA extraction using an in-house protocol (
http://ccdb.ca/resources/ accessed on 1 October 2022), and bidirectional sequencing of the standard 648-bp “DNA barcode” [
3,
4] in the COI mitochondrial gene. All samples except three were run with cocktail primers C_GasF1_t1 + GasR1_t1; Gast14_A01, Gast14_A02, and Gast14_A03 were run with both C_GasF1_t1 + GasR1_t1 and ZplankF1_t1 + ZplankR1_t1.
The procedure usually produced a DNA barcode of 658 nucleotides, and only those containing > 300 nucleotides were included in the sequence analysis. Groups of specimens with identical or almost identical DNA barcodes were assigned BIN numbers. They were compared with all barcode records on BOLD (10,580,183 Sequences) including the Public Record Barcode Database (2,529,561 Sequences/153,565 Species/66,474 Interim Species) using the Bold Aligner (Amino Acid Based HMM). The identification system on BOLD delivers a species identification if the query sequence shows less than 1% divergence to a reference sequence.
Conceptualization, Project administration, Data curation, Formal analysis: Peter Bryant; Investigation, Methodology: Timothy Arehart, Peter Bryant; Writing—original draft, Peter Bryant; Writing—review and editing, Peter Bryant and Timothy Arehart. Collections were made and analyzed with the assistance of Undergraduate students Taylor Sais, Alicia Navarro, Debbie Chung, Lesly Ortiz, and Bita Rostam.
3. Results
This project GASSC (Gastropoda of Southern California) included 1238 specimens of which 1235 provided images. 680 specimens provided a COI-5P sequence and 589 of these were Barcode-compliant, falling into 143 BINs containing 127 species (See
Supplementary Materials). Our data (
Figure 2) show a much larger range of interspecific divergences (seen in the graph of ”Distance to Nearest Neighbour”) compared to intraspecific divergences in this DNA sequence for gastropods suggesting the existence of a “DNA barcode gap”. Within species in this set of samples, the mean% divergence in sequence was 0.66 +/− 0.0 S.E. Within the largest set of conspecifics (
Crepidula onyx, n = 54) the mean% divergence in sequence was 0.69 +/− 0.0 S.E., maximum 2.49%, minimum 0.0%.
Subclass caenogastropoda
Order Littorinimorpha: Sea Snails
Family Atlantidae Wiegmann and Ruthe, 1832
A family of microscopic (<1 cm shell diameter), holoplanktonic gastropods [
1]. They have a transparent, coiled shell into which their bodies can be retracted, and an operculum that is used to close off the opening. The larval stage is a veliger in which the velum is initially small and bilobed, but with growth it develops three lobes (
Figure 3). The larval shell and operculum are retained in the adult.
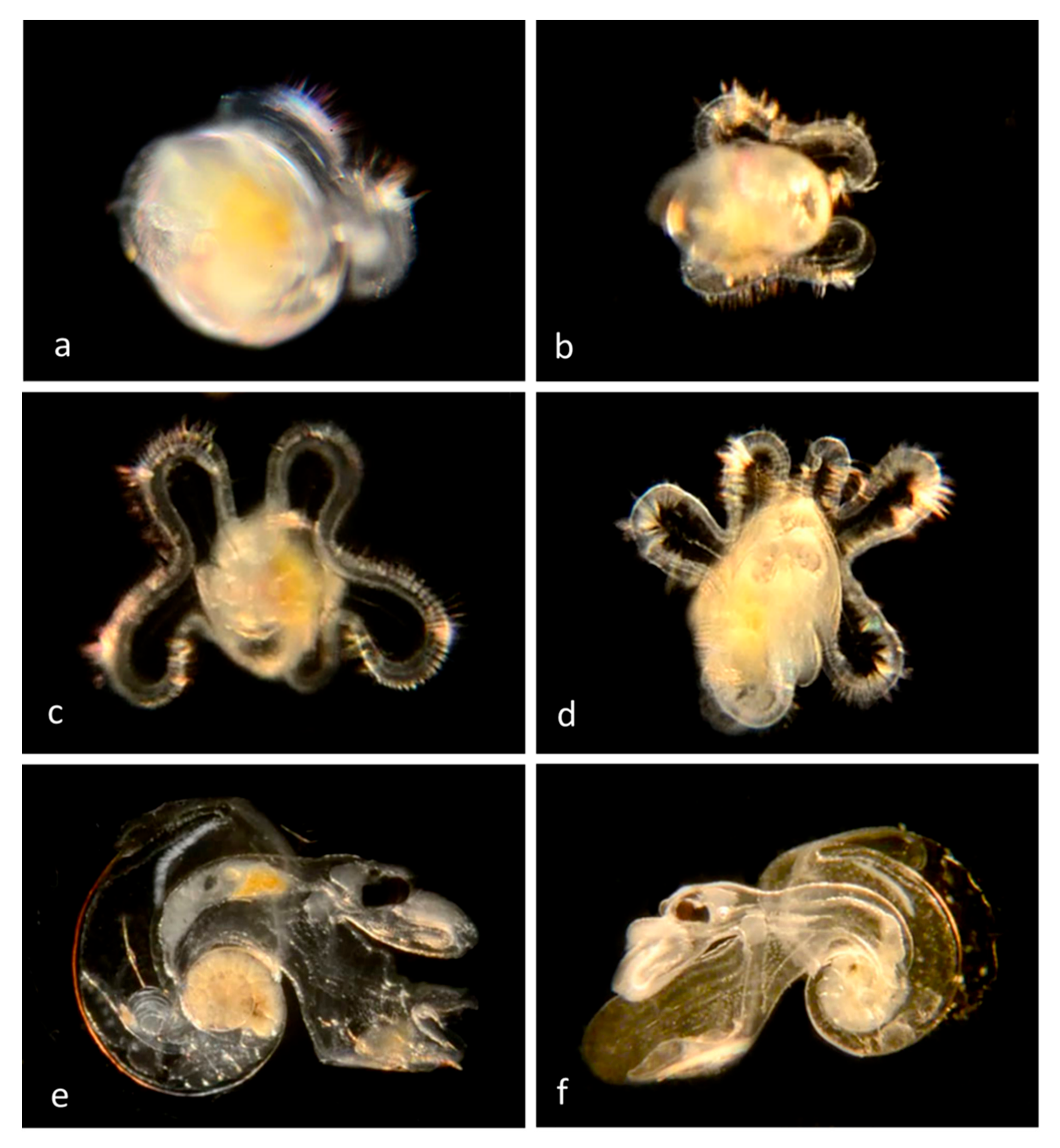
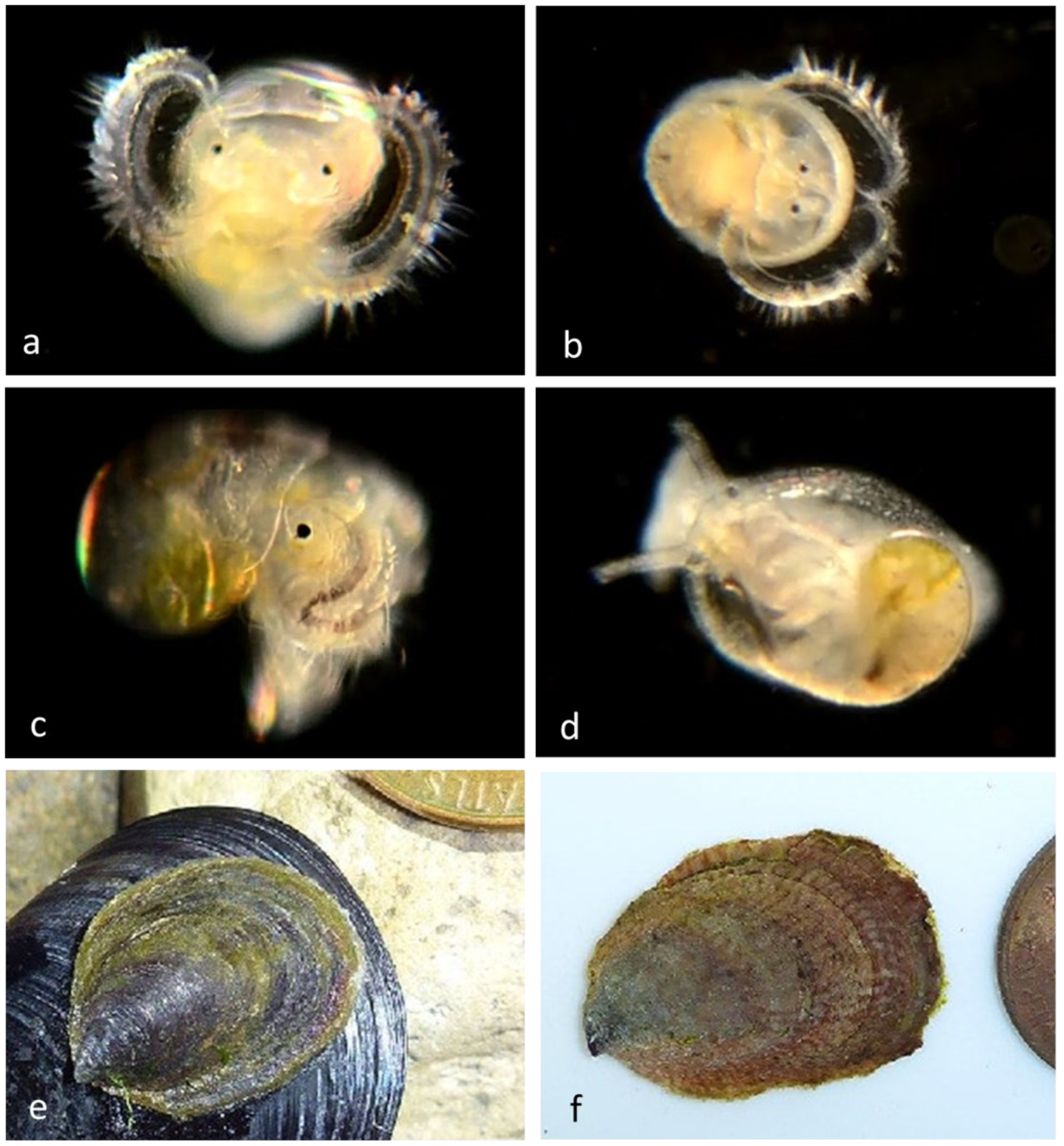
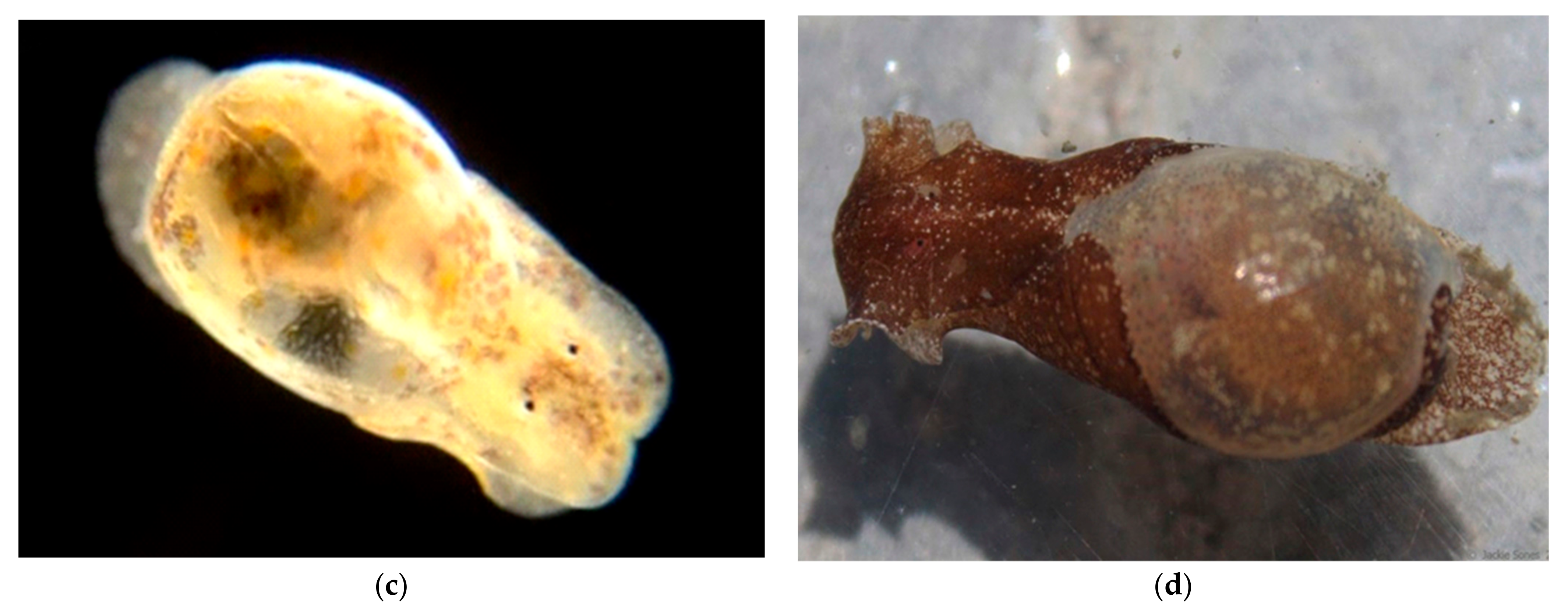
Family Calyptraeidae Lamarck, 1809, the Slipper Shells
The shell is quite flat and has an internal shelf-like half shell, so that it resembles a traditional bedroom slipper; hence the name “slipper shell” or “slipper limpet” (although these are not true limpets). During mating they pile on top of each other to form a tower called a mating chain, in which all individuals start as males but the basal member transforms into a female. Females produce eggs that are fertilized internally, and developing embryos are held beneath the mother’s shell until they hatch into microscopic trochophore larvae before developing into swimming larvae (veligers) carrying shells, thousands of which disperse and later metamorphose into juveniles on the ocean floor.We have examined individuals of
Crepidula onyx (
Figure 4)
Crepidula naticarum (
Figure 5) and
Crepidula huerta (
Figure 6). In some species (e.g.,
Figure 6a and
Figure 7b) the vela are decorated with yellow spots, the nature and function of which is unknown.
Our adult specimens of
Crepidula huertae, (
Figure 6) found inside the empty shell of a Bubble Snail (
Bulla gouldiana) from Balboa at Coral (Newport Harbor), supported by the presence of DNA-sequenced veligers at the Harbor Entrance, extend the host and geographic range of this species, which was otherwise known from hermit crab shells at Naples Reef, near Santa Barbara, Santa Barbara County, California [
10].
Family Cerithiopsidae H. Adams & A. Adams, 1853: The Cerithiopsids
A family of very small gastropods (
Figure 8) with high spires and multiple whorls.
Family Strombidae Rafinesque, 1815: The true Conchs
Medium to very large snails (
Figure 9), with eyes on long stalks. The shell has a long, narrow aperture with an anterior indentation that accommodates one of the eye stalks.
Family Vermetidae Rafinesque, 1815: Worm Snails
Small to medium-sized snails (
Figure 10), with very irregular elongated tubular shells often forming large clumps. Some species have opercula at the ends of the tubes, while others do not.
Subclass heterobranchia
The veliger has a shell, but this is lost during metamorphosis into the adult.
Family Hermaeidae H. Adams & A. Adams 1854
Small sea slugs (
Figure 11) with cerata containing branches of the digestive gland, and rhinophores with a recessed tip.
Family Onchidorididae Gray, 1827
Dorid nudibranchs (
Figure 12), of which we have found only one pelagic larva.
Order Neogastropoda: Sea Snails
Characterized by a long incurrent siphon and accompanying siphonal structure on the base of the shell.
Family Columbellidae Swainson, 1840: The Dove Snails
Minute to small snails (
Figure 13) with a thick shell with a narrow opening. The foot is narrow, and the siphon is very long.
Family Nassariidae Iredale, 1916 (1835): The Dog Whelks
Snails with rounded shells (
Figure 14), a high spire, an oval aperture, and a siphonal notch.
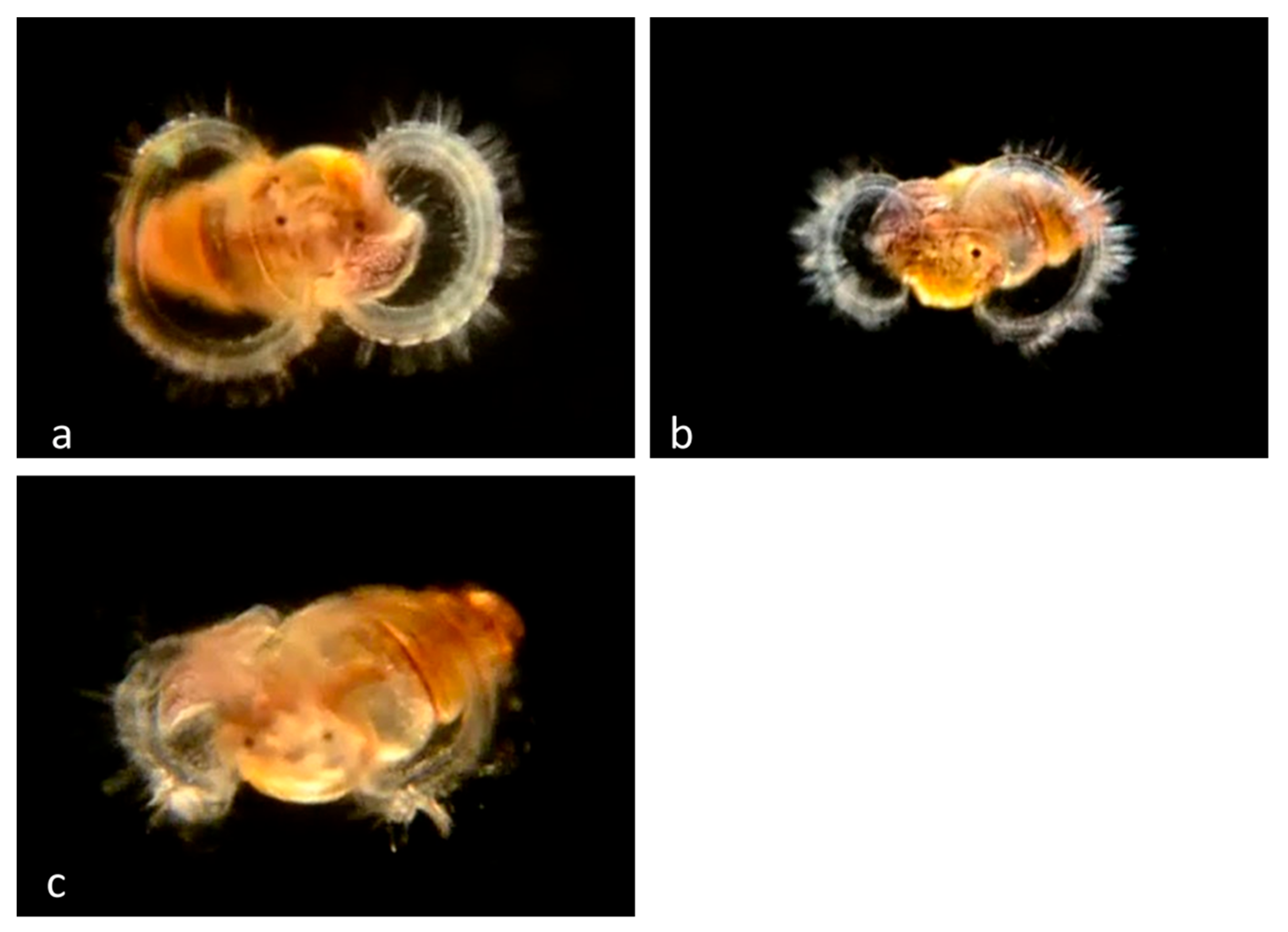
Family Muricidae Rafinesque, 1815
Within the family Muricidae, our collections include 27 adults but no larvae of
Nucella ostrina (BIN AAA4209), 26 adults but no larvae of
Acanthinucella spirata, 18 adults but no larvae of
Ceratostoma nuttalli and 2 adults but no larvae of
Mexacanthina lugubris. This is consistent with the finding that in these species of Muricidae the equivalent of the veliger stage occurs within the egg capsule, and the individuals hatch as juveniles which are not represented in our collections because of the limitations of our collection methods. In other family members Poulson’s Dwarf Triton,
Roperia poulsoni, and Festive Murex,
Pteropurpura festiva, individuals escape as veligers (
Figure 15 and
Figure 16).
Family Olivellidae Troschel, 1869: The Dwarf Olives
A family of small predatory snails (
Figure 17) with smooth, shiny, elongated shells.
Order Cephalaspidea: Sea Slugs and Bubble Snails
Family Gastropteridae Swainson, 1840 [
1] Bat-Winged Slugs
Adults have no shell, or an internal reduced shell (
Figure 18). They have outgrowths from the mantle wall called parapodia, used in swimming.
Family Haminoeidae Pilsbry, 1895
The shell (
Figure 19) is partially or completely enfolded by lateral fleshy parapodial lobes. Represented in our collection by the Blister Glassy-bubble,
Haminoea virescens (
Figure 19) and the White Bubble snail,
Haminoea vesicula (
Figure 20).
4. Discussion
Our results confirm that DNA barcoding using the COI barcode [
3,
4,
11] is a useful strategy to match life-cycle stages within species as well as to identify species and to document the level of biodiversity within a taxon, in this case the gastropods. Our data also show the expected “DNA barcode gap”; i.e., a much larger range of interspecific divergences versus intraspecific divergences in this DNA sequence for gastropods (
Figure 1). DNA barcoding has often revealed unexpected species diversity in many taxa [
11,
12] and the present study leads to the same conclusion for gastropods. The data will contribute to the development of a DNA barcode reference library, which will allow the rapid and convenient identification of individual gastropod individuals and parts collected at any developmental stage.
In this study we have begun to compile a collection of images of veligers, which are identified to the species level by matching of their DNA barcodes to those of morphologically recognized adults. These images confirm that the respective species have free-swimming veligers, and show that there is considerable morphological diversity, with some veligers having two velar lobes, some having four velar lobes, and some having yellow- or red-spotted vela. However, as expected from studies of larval stages in general, it is difficult to identify many species from the morphology of veligers.
The DNA sequence differences in the COI barcode are, of course, not responsible for the morphological differences we have observed between specimens in separate taxa. However, the DNA barcode differences that have evolved between morphologically distinct organisms can be used to examine the degree of relatedness between them. When the DNA sequence data are organized into a taxonomic tree (see Taxon Tree in
Supplementary Materials), the results are generally consistent with the taxonomic tree according to conventional morphological methods. This can be explored by cladistic analysis, in which the taxonomic tree is examined for “DNA clades”—groups of species that are uniquely and exclusively related by DNA sequence. According to this analysis, all four species of
Lottia, three species of
Crepidula, two of
Cerithidea, two of
Nassarius, three of
Littorina, two of
Nuttalina, two of
Epitonium, four of
Tegula, three of
Doriopsilla, two of
Ancula, two of
Felimare, two of
Corambe, and two of
Aplysia form clades, although the
Lottia and
Tegula clades include some different species.
DNA barcode reference libraries can lead to the development of more global sequencing strategies including metabarcoding and parallel sequencing of complex bulk samples including “environmental DNA”, which are being developed for monitoring ecosystem health. For example, in a recent study of plankton communities in the Baltic Sea, five nonindigenous species were discovered, and four of these were identified exclusively by metabarcoding [
13]. Our work illustrates the need for more larval/adult matching to build sequence libraries specifically for meta- and eDNA barcoding.

























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}