

Genetic Diversity in Marginal Populations of Nitraria schoberi L. from Romania


Abstract

1. Introduction
2. Materials and Methods
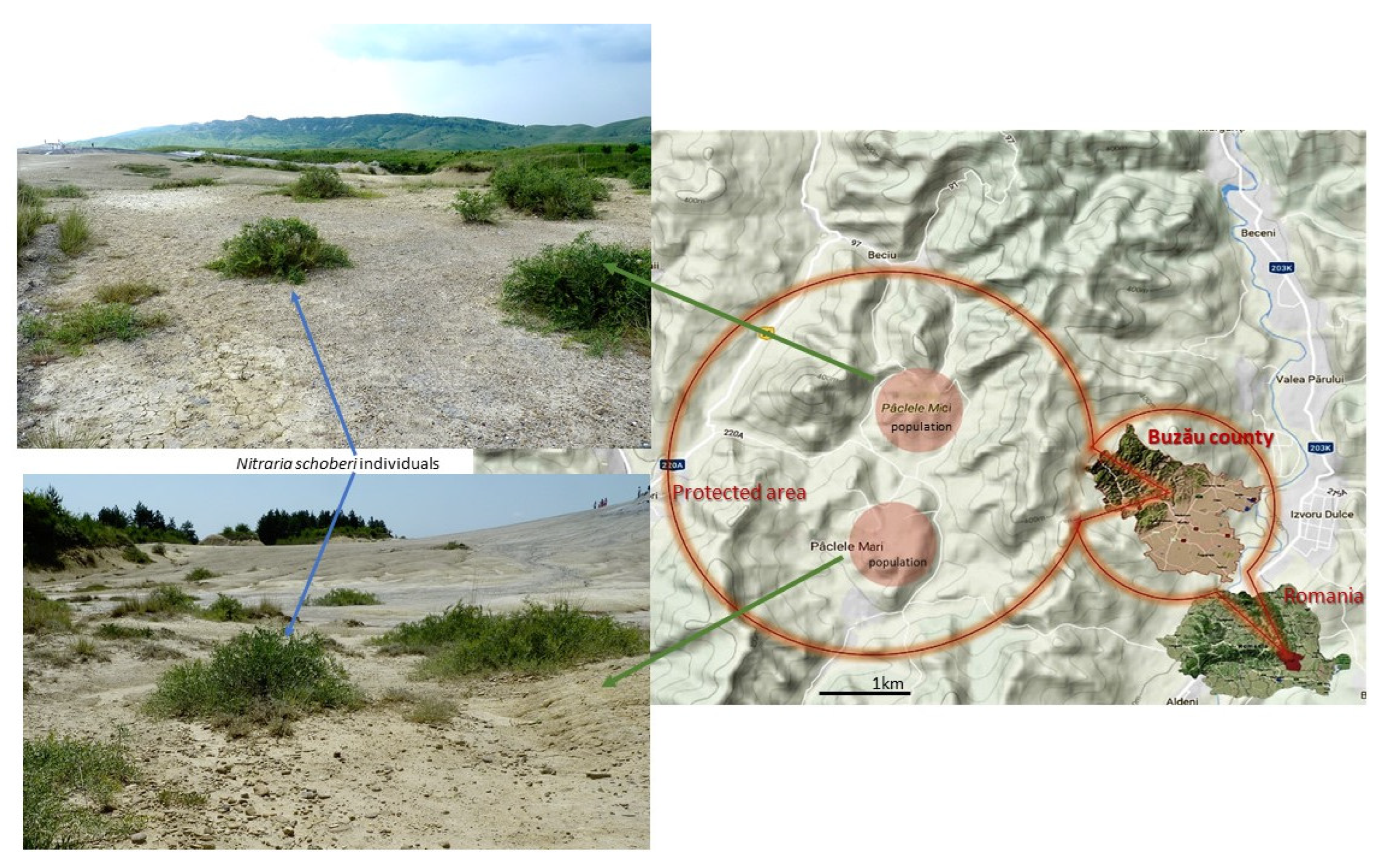
2.1. Plant Material
2.2. DNA Extraction
2.3. PCR Amplification and Electrophoresis
2.4. Data Analysis
3. Results
3.1. Scored Bands
3.2. Genetic Diversity
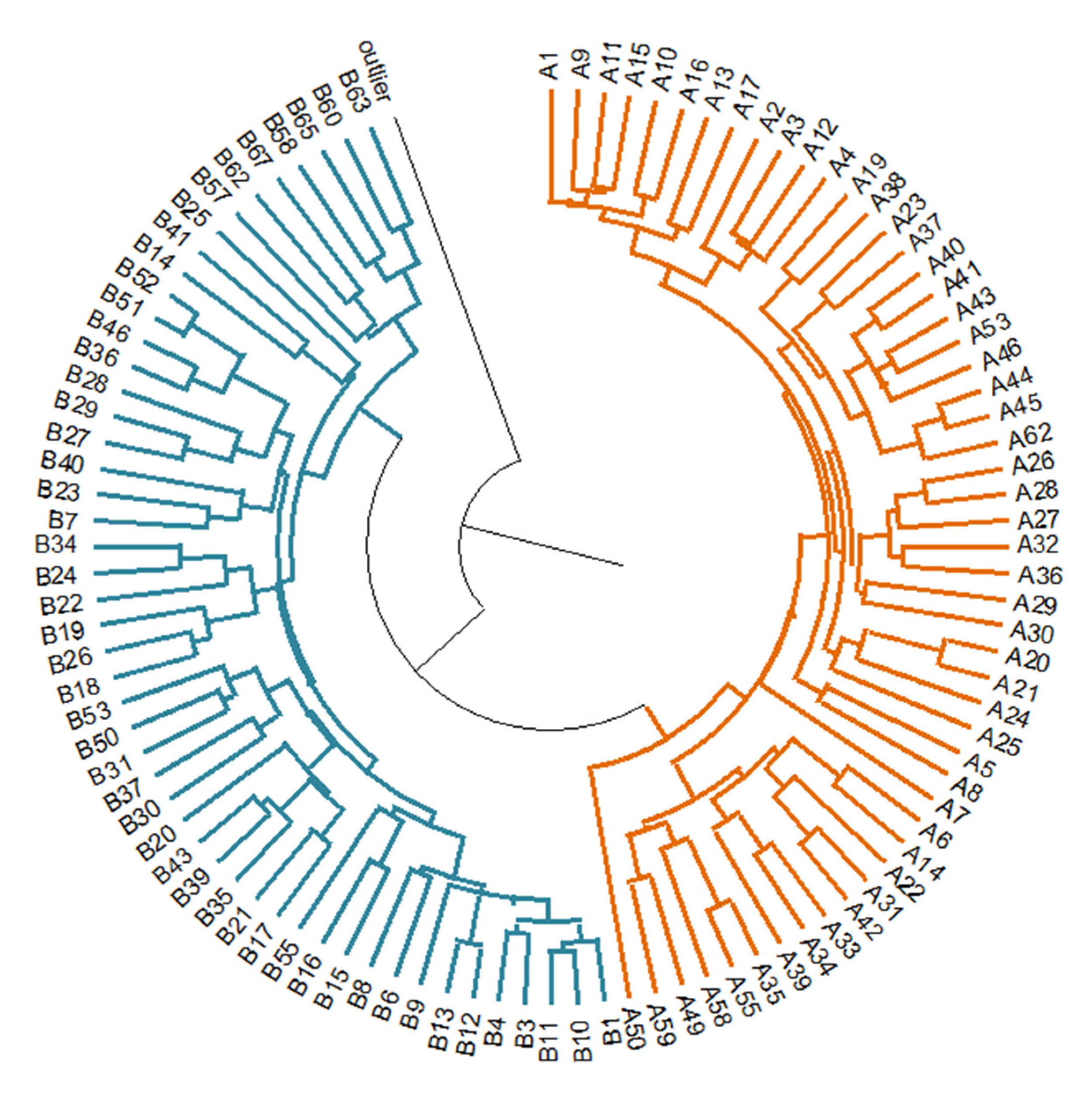
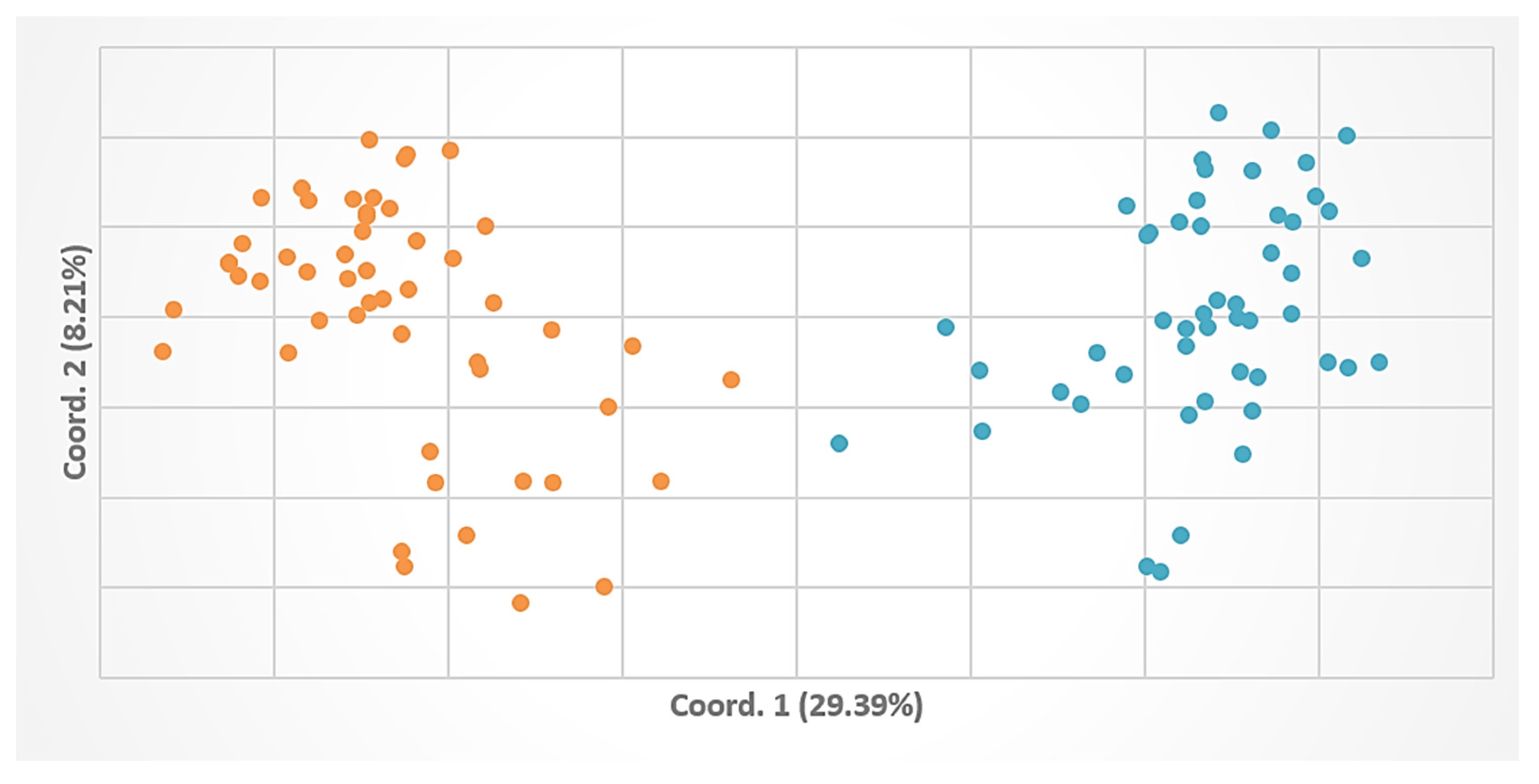
3.3. Genetic Relatedness among Populations
4. Discussion
4.1. Analysis of Genetic Diversity and Structure
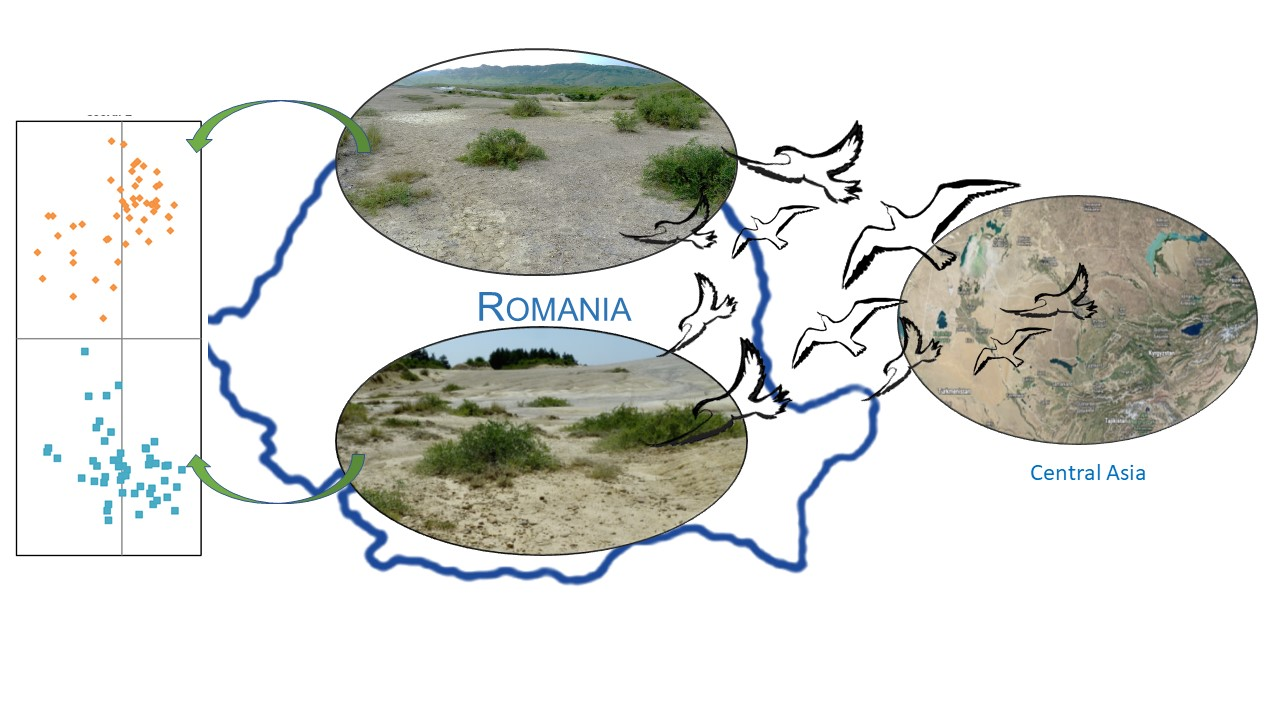
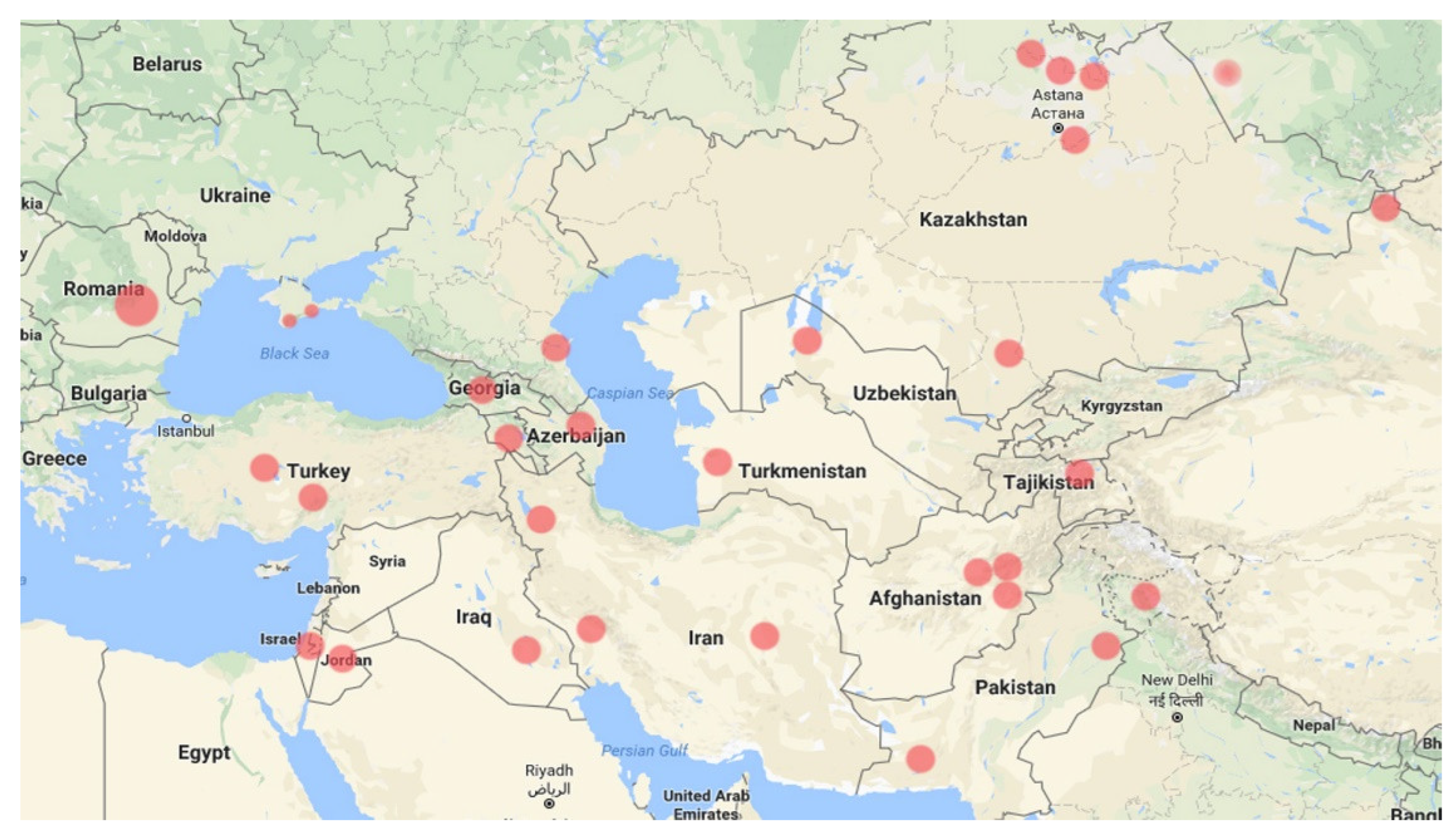
4.2. Nitre-Bush Colonization in Eastern Europe
4.3. Conservation Status
4.4. Further Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sokoloff, D.D.; Balandin, S.A.; Gubanov, A.I.; Jarvis, C.E.; Majorov, S.R.; Simonov, S.S. The history of botany in Moscow and Russia in the 18th and early 19th centuries in the context of the Linnaean Collection at Moscow University (MW). Huntia 2002, 11, 129–191. [Google Scholar] [PubMed]
- DG Environment. Reporting under Article 17 of the Habitats Directive: Explanatory Notes and Guidelines for the Period 2013–2018; DG Environment: Brussels, Belgium, 2017; p. 188. Available online: https://circabc.europa.eu/ (accessed on 7 January 2020).
- Rosa, J.D.; Weber, G.G.; Cardoso, R.; Górski, F.; Da-Silva, P.R. Variability and population genetic structure in Achyrocline flaccida (Weinm.) DC., a species with high value in folk medicine in South America. PLoS ONE 2017, 12, e0183533. [Google Scholar] [CrossRef] [PubMed]
- Temirbayeva, K.; Zhang, M.L. Molecular phylogenetic and biogeographical analysis of Nitraria based on nuclear and chloroplast DNA sequences. Plant Syst. Evol. 2017, 301, 1897–1906. [Google Scholar] [CrossRef]
- Zhang, M.L.; Temirbayeva, K.; Sanderson, S.C.; Chen, X. Young dispersal of xerophil Nitraria lineages in intercontinental disjunctions of the Old World. Sci. Rep. 2015, 5, 13840. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yang, A.; Dick, W.C.; Yao, X.; Huang, H. Impacts of biogeographic history and marginal population genetics on species range limits: A case study of Liriodendron chinense. Sci. Rep. 2016, 6, 25632. [Google Scholar] [CrossRef]
- Chhatre, V.E.; Rajora, O.P. Genetic Divergence and Signatures of Natural Selection in Marginal Populations of a Keystone, Long-Lived Conifer, Eastern White Pine (Pinus strobus) from Northern Ontario. PLoS ONE 2014, 9, e97291. [Google Scholar] [CrossRef]
- Reddy, M.P.; Sarla, N.; Siddiq, E.A. Inter Sample Sequence Repeat (ISSR) polymorphism and its application in plant breeding. Euphytica 2002, 128, 9–17. [Google Scholar] [CrossRef]
- Ziekiewicz, E.; Rafalski, A.; Labuda, A. Genome fingerprinting by simple sequence repeat (SSR) anchored polymerase chain reaction amplification. Genomics 1994, 20, 178–183. [Google Scholar]
- Bornet, B.; Branchard, M. Non-anchored inter-simple sequence repeat (ISSR) markers: Reproducible and specific tools for genome fingerprinting. Plant Mol. Biol. Rep. 2001, 19, 209–215. [Google Scholar] [CrossRef]
- Rakoczy-Trojanowska, M.; Bolibok, H. Characteristics and a comparison of three classes of microsatellite-based markers and their application in plants. Cell. Mol. Biol. Lett. 2004, 9, 221–238. [Google Scholar]
- Khajeddini, M.A.; Dadpour, M.R.; Khodaverdi, M.; Naghiloo, S. The GC-MS analyses of the n-hexane extract of Nitraria schoberi L., its total phenolics and in vitro antioxidant activity. J. Med. Plants Res. 2012, 6, 4874–4878. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Hoseini-Alfatemi, S.M.; Sharifi-Rad, M.; Teixeira da Silva, J.A. Antibacterial, antioxidant, antifungal and anti-inflammatory activities of crude extract from Nitraria schoberi fruits. 3 Biotech 2015, 5, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Üstünes, L. Pharmacological activity of the extracts of Nitraria schoberi (Zygophyllaceae). J. Fac. Pharm. Ankara Univ. 1988, 18, 74–86. [Google Scholar] [CrossRef]
- Tomoshevich, M.; Banaev, E.; Khozyaykina, S.; Erst, A. Pollen Morphology of Some Species from Genus Nitraria. Plants 2022, 11, 2359. [Google Scholar] [CrossRef] [PubMed]
- Banaev, E.V.; Tomoshevich, M.A.; Voronkova, M.S. Flow cytometry analysis of the relative content of nuclear DNA in Nitraria schoberi L. seeds. Bot. Pac. 2018, 7, 89–92. [Google Scholar] [CrossRef][Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W. Effects of life history traits on genetic diversity in plant species’. Philos. Trans. R. Soc. B Biol. Sci. 1996, 351, 1291–1298. [Google Scholar] [CrossRef]
- Nybom, H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants. Mol. Ecol. 2004, 13, 1143–1155. [Google Scholar] [CrossRef]
- Vásquez, D.L.; Balslev, H.; Hansen, M.M.; Sklenář, P.; Romoleroux, K. Low genetic variation and high differentiation across sky island populations of Lupinus alopecuroides (Fabaceae) in the northern Andes. Alp. Bot. 2016, 126, 135–142. [Google Scholar] [CrossRef]
- De Kort, H.; Prunier, J.G.; Ducatez, S.; Honnay, O.; Baguette, M.; Stevens, V.M.; Blanchet, S. Life history, climate and biogeography interactively affect worldwide genetic diversity of plant and animal populations. Nat. Commun. 2021, 12, 516. [Google Scholar] [CrossRef] [PubMed]
- Banaev, E.V.; Khozyaykina, S.A.; Dorogina, O.V.; Tomoshevich, M.A. Genetic Diversity of Nitraria sibirica Pall.(Nitrariaceae) in the Populations of Siberia and Kazakhstan Based on the Polymorphism of Intermicrosatellite Markers. Contemp. Probl. Ecol. 2022, 15, 400–408. [Google Scholar] [CrossRef]
- Kvaček, Z. Forest flora and vegetation of the European early Palaeogene—A review. B Geosci. 2010, 85, 63–76. [Google Scholar] [CrossRef]
- Brustur, T.; Stănescu, I.; Macaleţ, R.; Melinte—Dobrinescu, M. The mud volcanoes from Berca: A significant geological patrimony site of the Buzău Land Geopark (Romania). Geo-Eco-Marina 2015, 21, 1–22. [Google Scholar] [CrossRef]
- Bao, F.; Xin, Z.; Liu, M.; Li, J.; Gao, Y.; Lu, Q.; Wu, B. Preceding Phenological Events Rather than Climate Drive the Variations in Fruiting Phenology in the Desert Shrub Nitraria tangutorum. Plants 2022, 11, 1578. [Google Scholar] [CrossRef]
- Noble, J.C.; Whalley, R.D.B. The biology and autecology of Nitraria, L. in Australia. II. Seed germination, seedling establishment and response to salinity. Austral Ecol. 1978, 3, 165–177. [Google Scholar] [CrossRef]
- Galbraith, C.A.; Kirby, J.; Mundkur, T.A. Review of Migratory Bird Flyways and Priorities for Management; CMS Technical Series No. 27; UNEP/CMS Secretariat: Bonn, Germany, 2014; 164p, Available online: https://www.cms.int/sites/default/files/publication/CMS_Flyways_Reviews_Web.pdf (accessed on 4 May 2020).
- Morrison, C.A.; Robinson, R.A.; Butler, S.J.; Clark, J.A.; Gill, J.A. Demographic drivers of decline and recovery in an Afro-Palaearctic migratory bird population. Proc. Soc. B Sci. 2016, 283, 20161387. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Hao, J.; Chen, Z.; Zhang, G.; Yan, M.; Wang, J. Root Water Uptake Patterns for Nitraria during the Growth Period Differing in Time Interval from a Precipitation Event in Arid Regions. Sustainability 2022, 14, 8203. [Google Scholar] [CrossRef]
- Igea, J.; Miller, E.F.; Papadopulos, A.S.T.; Tanentzap, A.J. Seed size and its rate of evolution correlate with species diversification across angiosperms. PLOS Biol. 2017, 15, e2002792. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | No.of Loci | No.of Polymorphic Loci | % Polymorphic Loci | Size of PCR Products (bp) | ||
|---|---|---|---|---|---|---|---|
| PM | pm | PM | pm | ||||
| UBC809 | (AG)8G | 14 | 11 | 14 | 78.57% | 100.00% | 150–2000 |
| UBC824 | (TC)8G | 17 | 15 | 17 | 88.24% | 100.00% | 130–1350 |
| UBC853 | (TC)8RT | 14 | 13 | 12 | 92.86% | 85.71% | 180–1500 |
| UBC854 | (TC)8RG | 12 | 10 | 11 | 83.33% | 91.67% | 200–1500 |
| UBC888 | BDB(CA)7 | 8 | 6 | 5 | 75.00% | 62.50% | 270–1450 |
| UBC891 | HVH(TG)7 | 11 | 9 | 5 | 81.82% | 45.45% | 200–1900 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paica, I.C.; Banciu, C.; Maria, G.M.; Vladimirescu, M.; Manole, A. Genetic Diversity in Marginal Populations of Nitraria schoberi L. from Romania. Diversity 2022, 14, 882. https://doi.org/10.3390/d14100882
Paica IC, Banciu C, Maria GM, Vladimirescu M, Manole A. Genetic Diversity in Marginal Populations of Nitraria schoberi L. from Romania. Diversity. 2022; 14(10):882. https://doi.org/10.3390/d14100882
Chicago/Turabian StylePaica, Ioana C., Cristian Banciu, Gabriel M. Maria, Mihnea Vladimirescu, and Anca Manole. 2022. "Genetic Diversity in Marginal Populations of Nitraria schoberi L. from Romania" Diversity 14, no. 10: 882. https://doi.org/10.3390/d14100882
APA StylePaica, I. C., Banciu, C., Maria, G. M., Vladimirescu, M., & Manole, A. (2022). Genetic Diversity in Marginal Populations of Nitraria schoberi L. from Romania. Diversity, 14(10), 882. https://doi.org/10.3390/d14100882