
Leaf Epidermal Traits of Selected Euphorbiaceae and Phyllanthaceae Taxa of Hainan Island and Their Taxonomic Relevance



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Collection and Identification
2.3. Light Microscopy (LM)
2.4. Scanning Electron Microscopy (SEM)
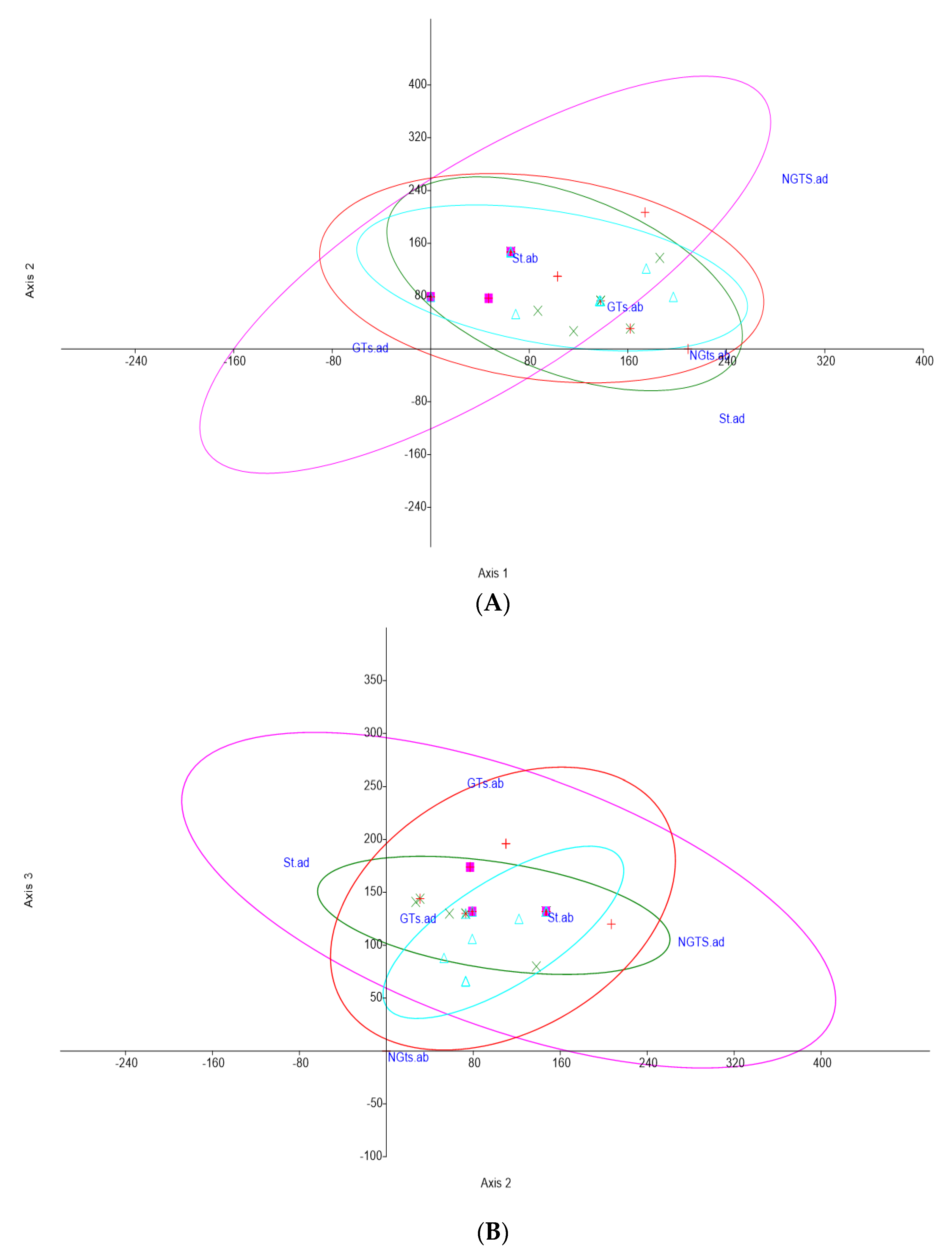
2.5. Statistical Analysis
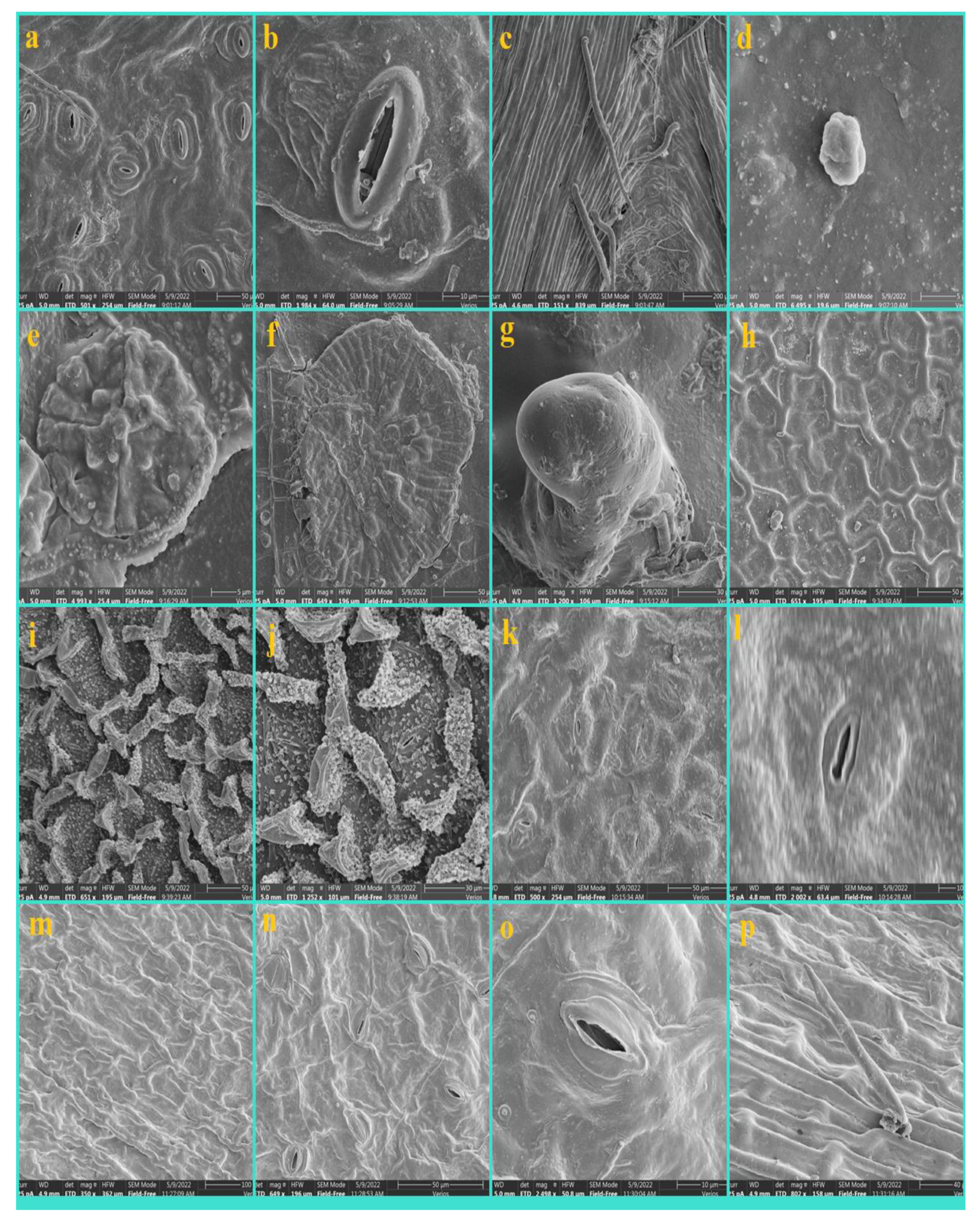
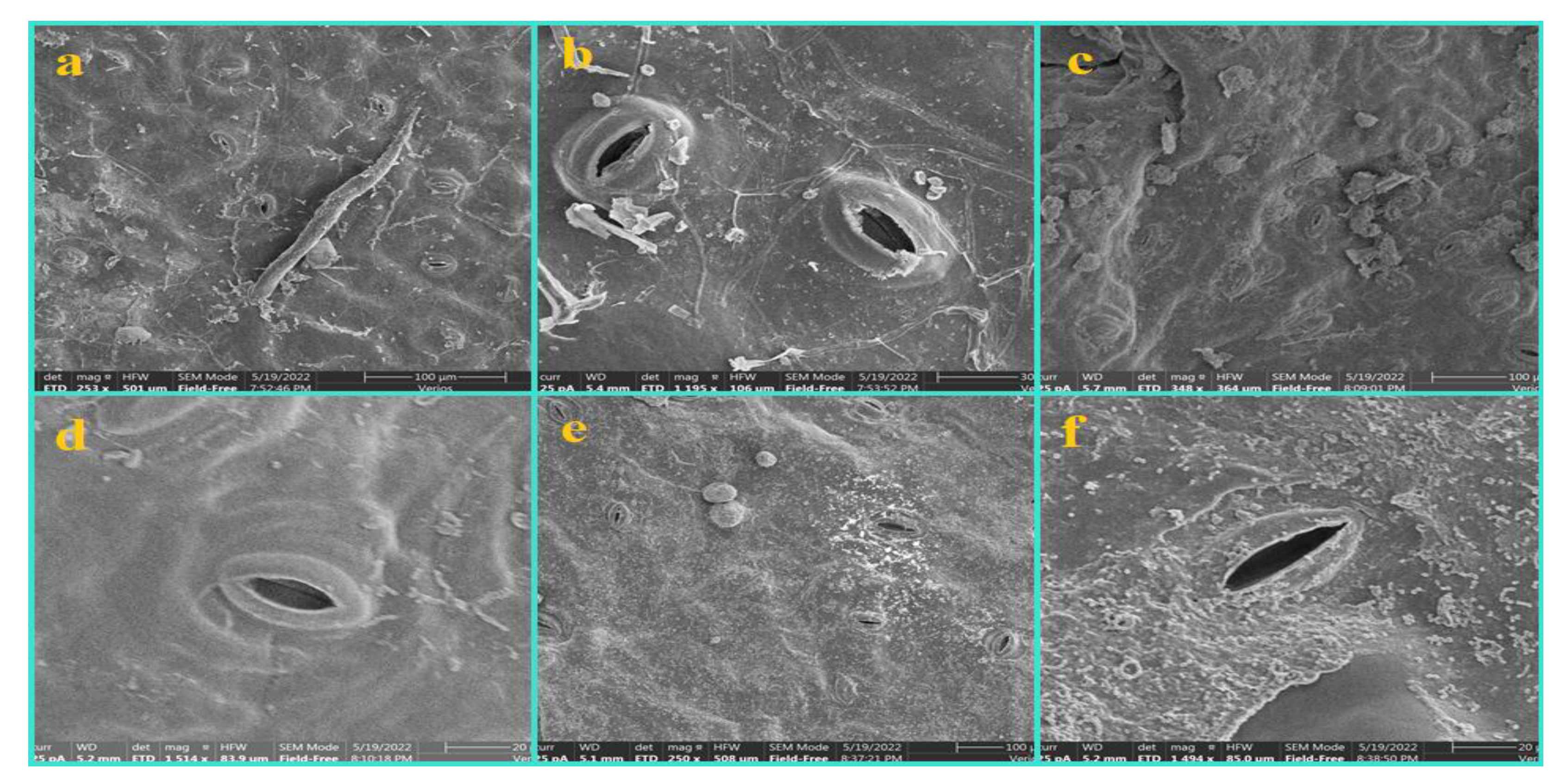
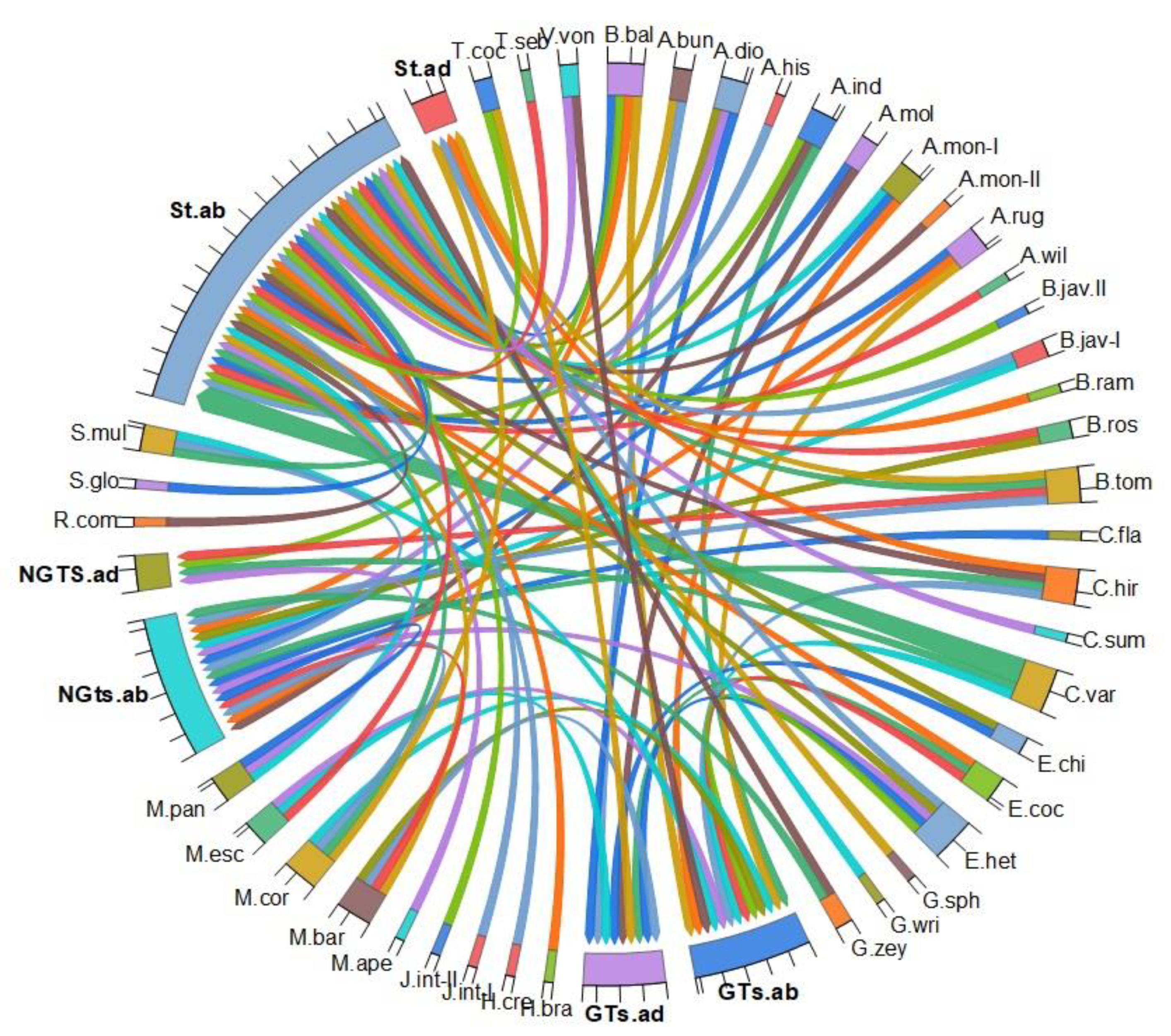
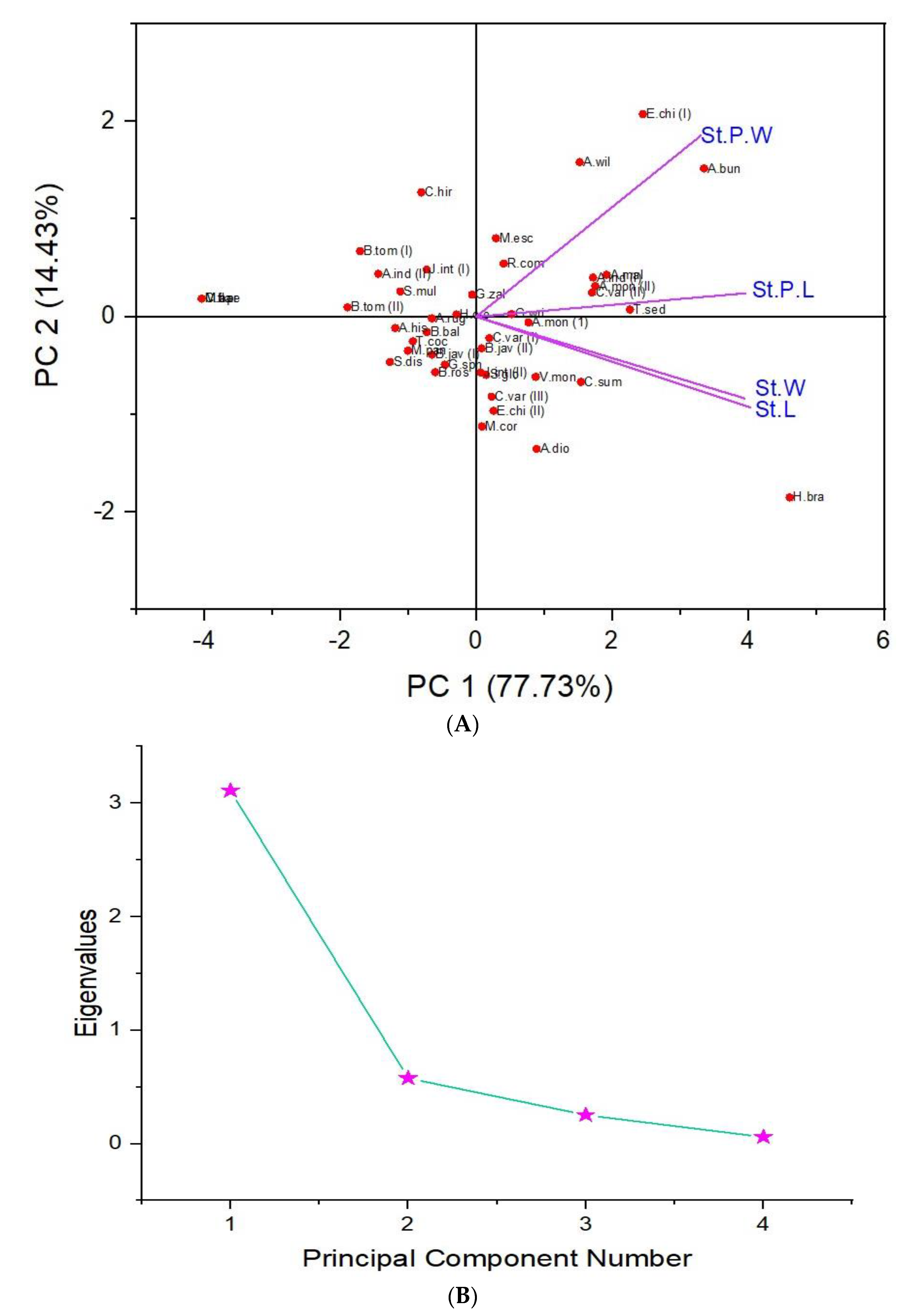
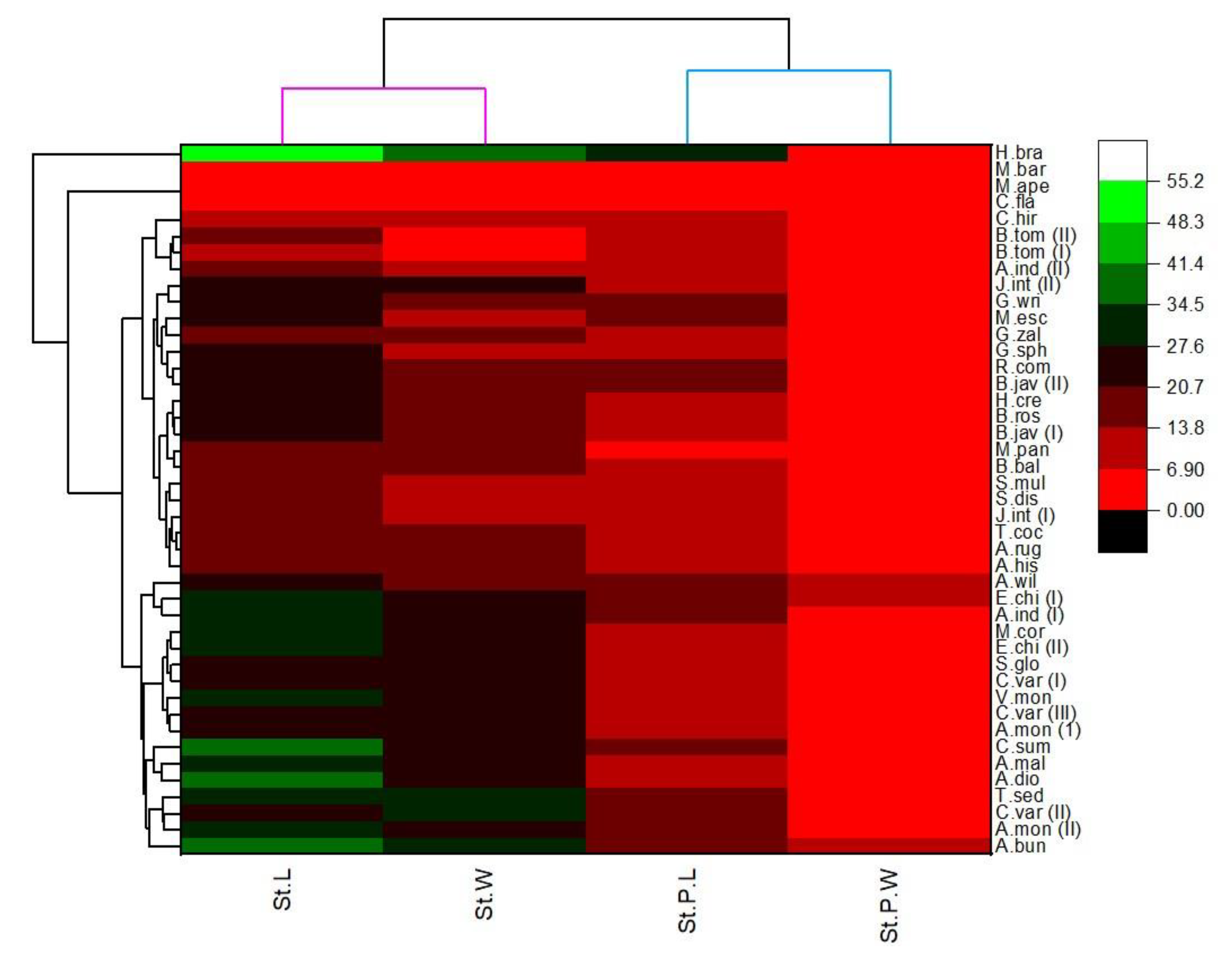
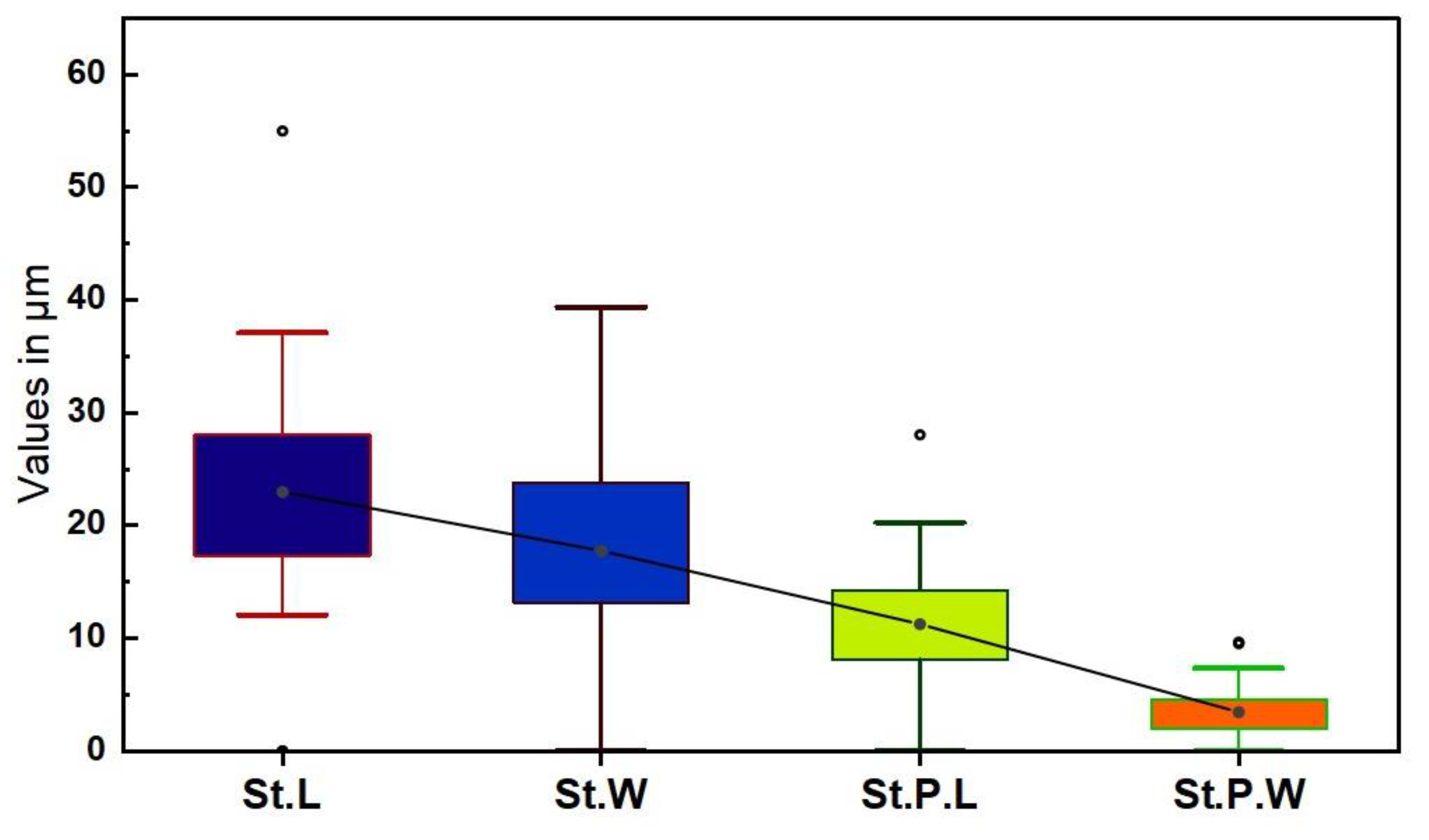
3. Results
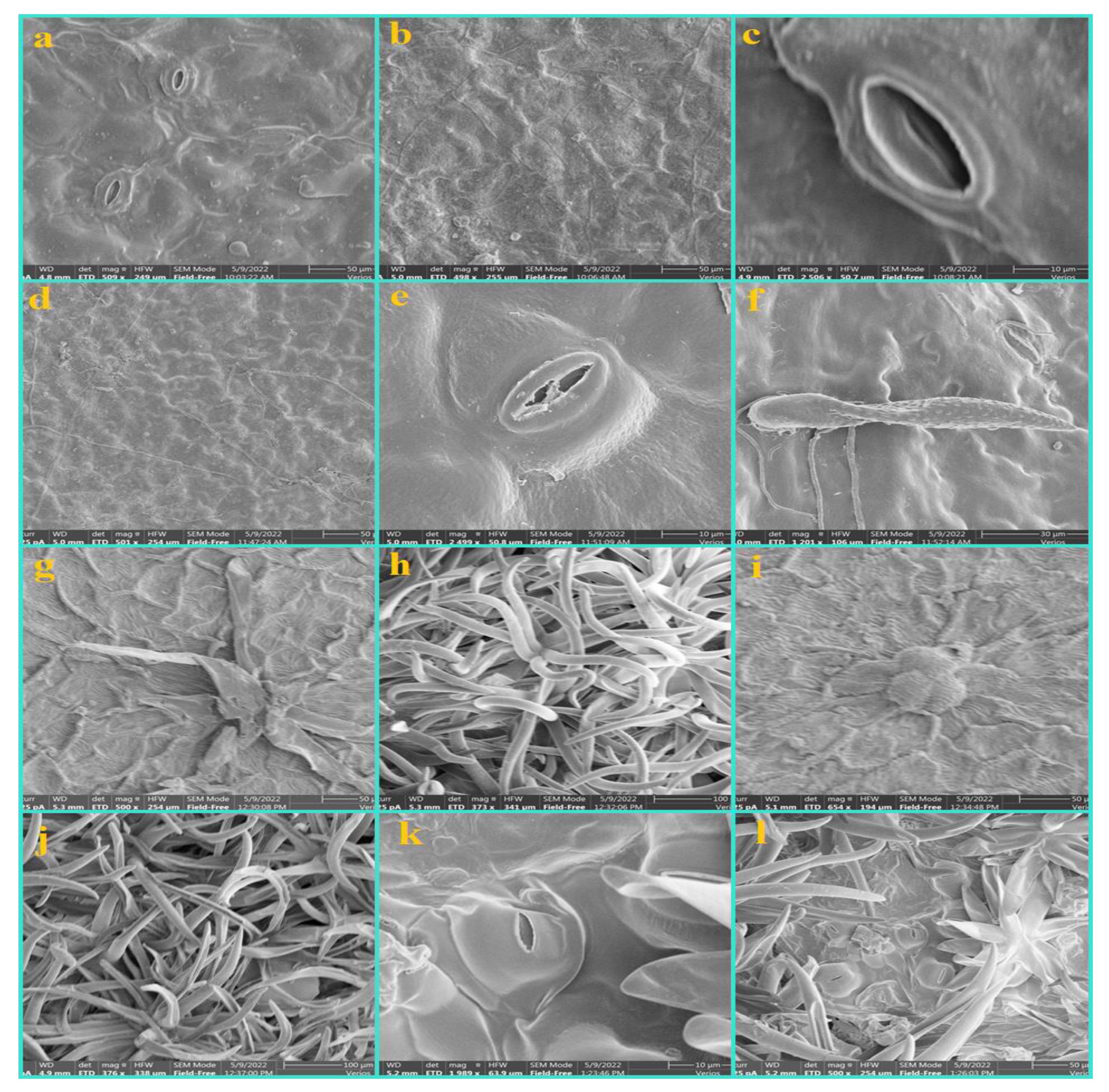
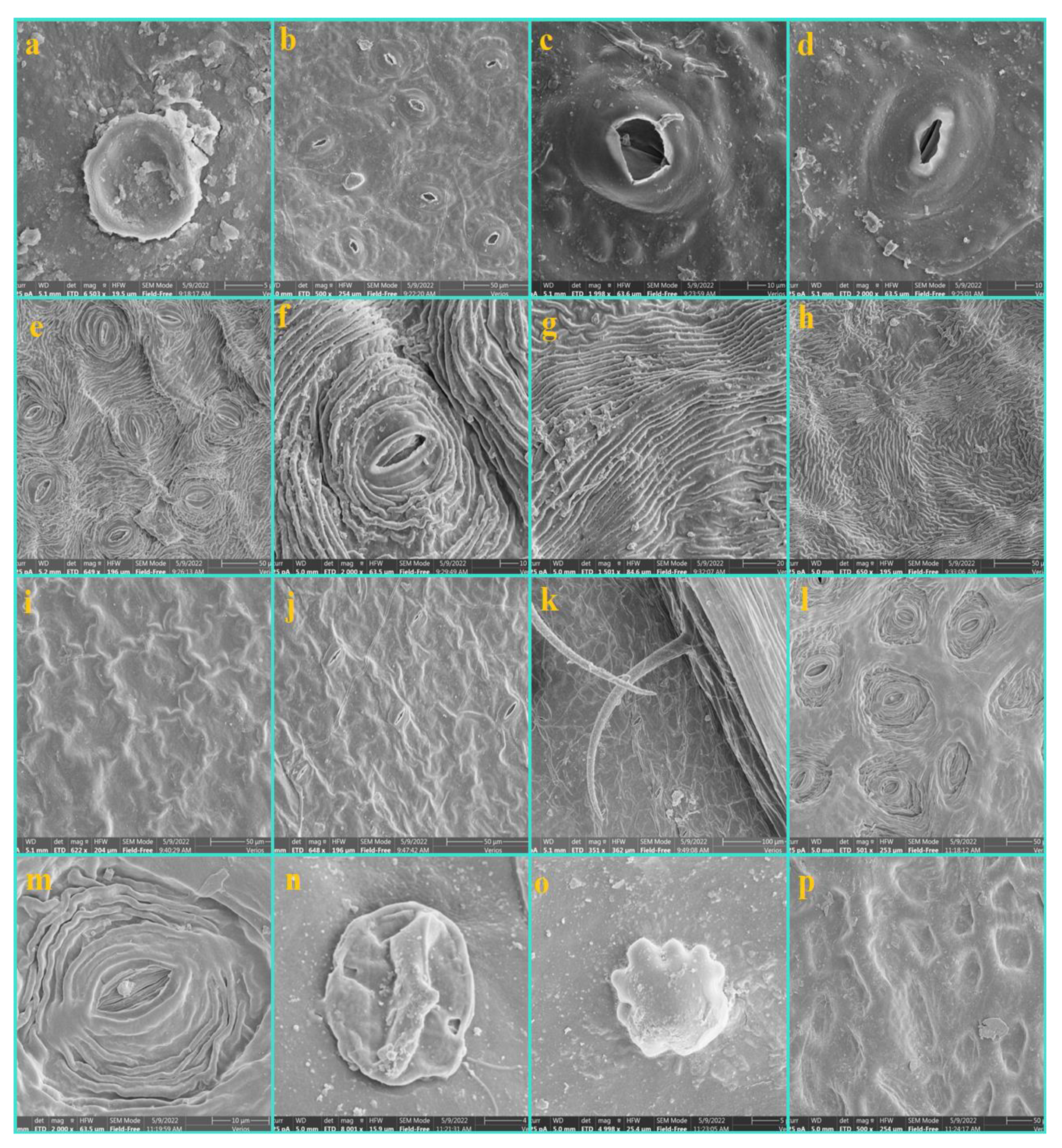
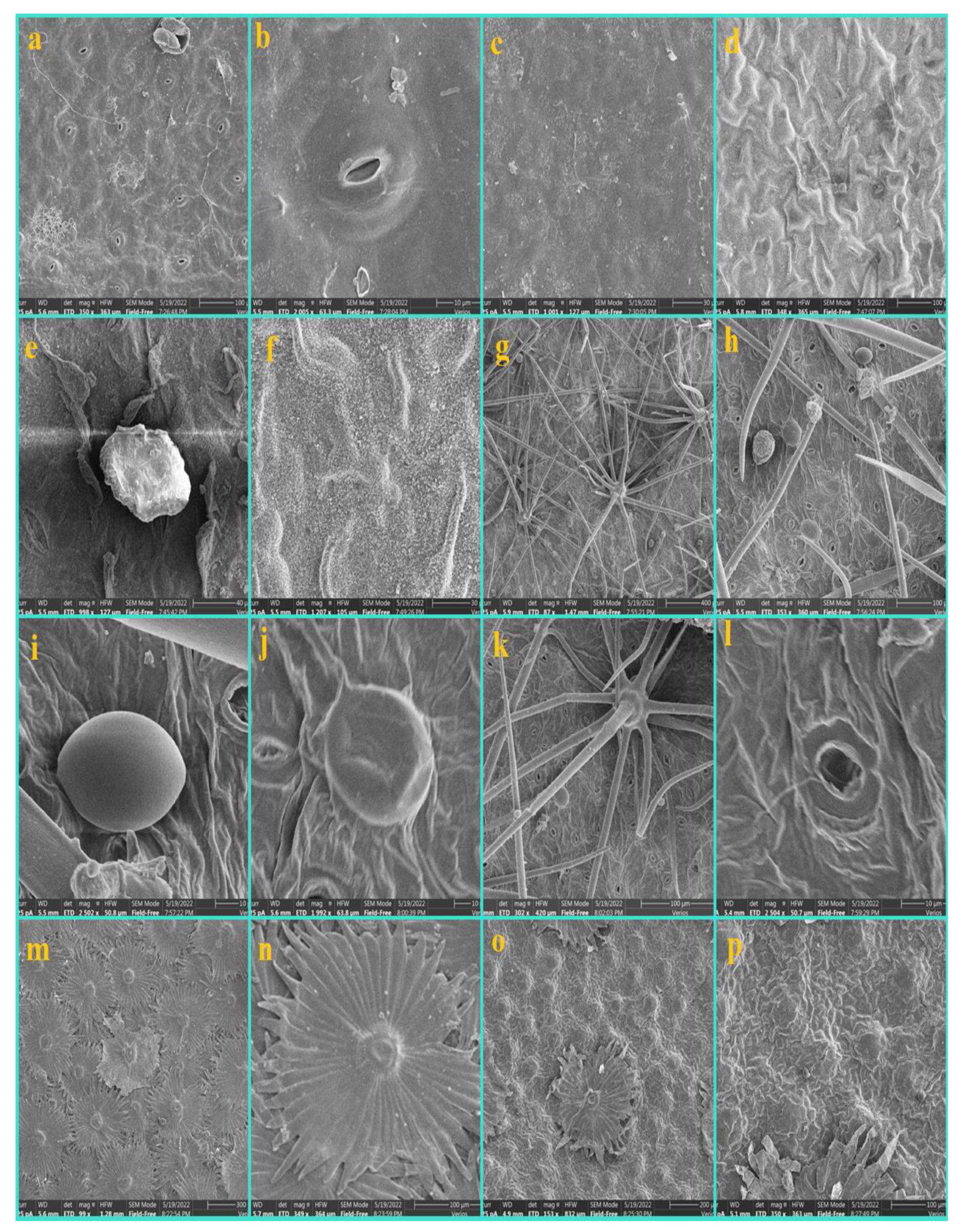
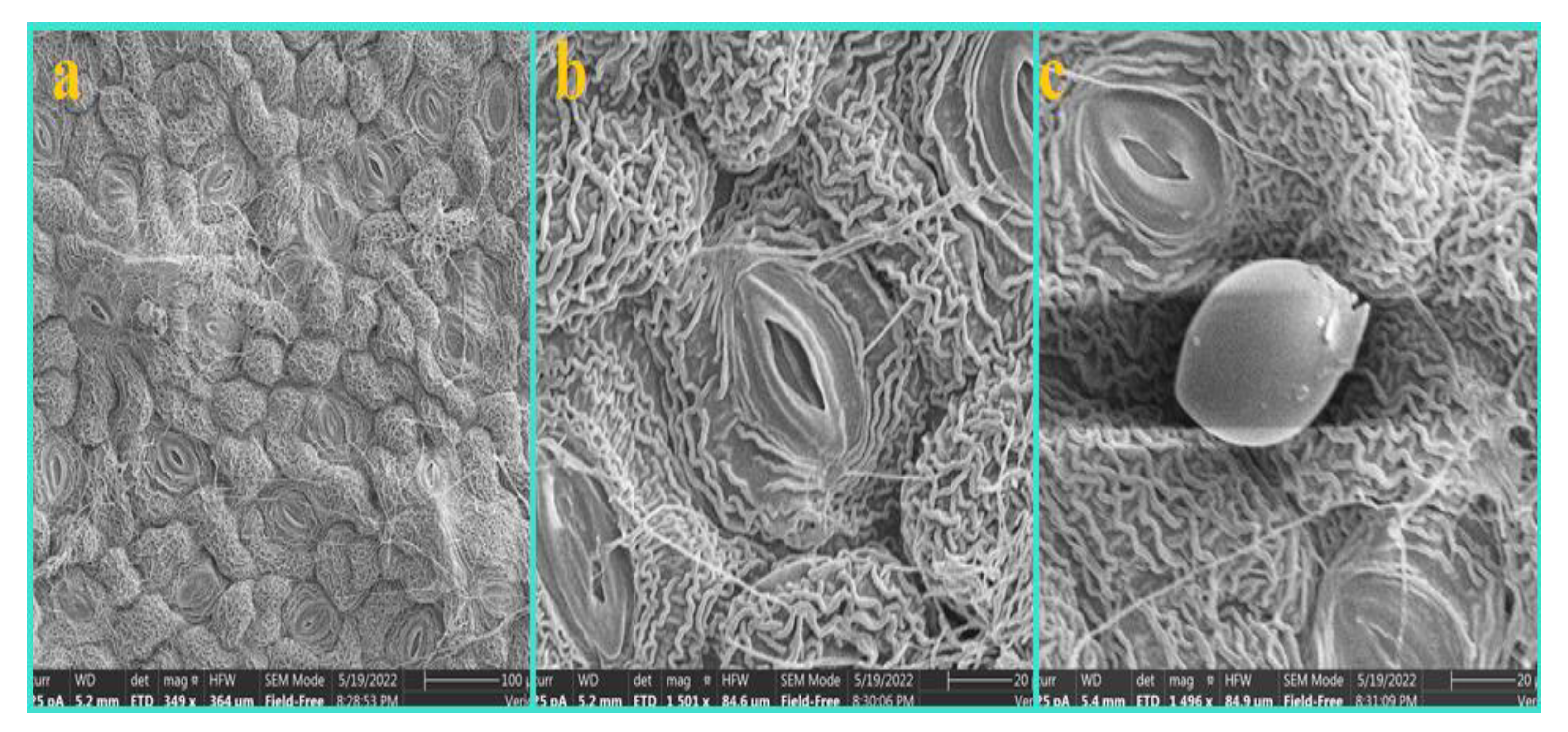
3.1. Epidermis
3.2. Nonglandular Trichomes (NGTs)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Stomata | NGTS | GTs | |||
|---|---|---|---|---|---|---|
| Adaxial | Abaxial | Adaxial | Abaxial | Adaxial | Abaxial | |
| Acalyphoideae | ||||||
| Acalypha hispida | 0 | 1 | 0 | 0 | 0 | 0 |
| Acalypha indica | 0 | 1 | 0 | 1 | 0 | 1 |
| Acalypha wilkesiana | 0 | 1 | 0 | 0 | 0 | 0 |
| Alchornea rugosa | 0 | 1 | 0 | 1 | 0 | 1 |
| Macaranga corymbosa | 1 | 1 | 0 | 1 | 0 | 1 |
| Mallotus apelta | 0 | 1 | 0 | 0 | 0 | 0 |
| Mallotus barbatus | 0 | 1 | 0 | 1 | 1 | 1 |
| Mallotus paniculatus | 0 | 1 | 1 | 1 | 0 | 0 |
| Ricinus communis | 0 | 1 | 0 | 0 | 0 | 0 |
| Euphorbioideae | ||||||
| Euphorbia heterophylla | 1 | 1 | 0 | 1 | 1 | 1 |
| Excoecaria cochinchinensis | 0 | 1 | 0 | 0 | 1 | 1 |
| Hura crepitans | 0 | 1 | 0 | 0 | 0 | 0 |
| Triadica cochinchinensis | 0 | 1 | 0 | 0 | 1 | 0 |
| Triadica sebifera | 0 | 1 | 0 | 0 | 0 | 0 |
| Crotonoideae | ||||||
| Aleurites moluccana | 0 | 1 | 0 | 0 | 1 | 0 |
| Codiaeum variegatum (I) | 0 | 1 | 0 | 0 | 0 | 1 |
| Codiaeum variegatum (II) | 0 | 1 | 0 | 0 | 0 | 0 |
| Codiaeum variegatum (III) | 0 | 1 | 1 | 0 | 0 | 0 |
| Croton flavens | 0 | 0 | 0 | 1 | 0 | 0 |
| Croton hirtus | 1 | 1 | 0 | 1 | 0 | 1 |
| Endospermum chinense | 0 | 1 | 0 | 0 | 1 | 0 |
| Hevea brasiliensis | 0 | 1 | 0 | 0 | 0 | 0 |
| Jatropha integerrima (I) | 0 | 1 | 0 | 0 | 0 | 0 |
| Jatropha integerrima (II) | 0 | 1 | 0 | 0 | 0 | 0 |
| Manihot esculenta | 0 | 1 | 0 | 0 | 1 | 1 |
| Suregada glomerulata | 0 | 1 | 0 | 0 | 0 | 0 |
| Suregada multiflora | 0 | 1 | 0 | 1 | 0 | 1 |
| Vernicia montana | 0 | 1 | 0 | 0 | 0 | 1 |
| Phyllanthaceae | ||||||
| Antidesma bunius | 0 | 1 | 0 | 0 | 1 | 0 |
| Antidesma montanum (I) | 0 | 1 | 0 | 1 | 0 | 1 |
| Antidesma montanum (II) | 0 | 1 | 0 | 0 | 0 | 0 |
| Aporosa dioca | 0 | 1 | 0 | 1 | 1 | 0 |
| Baccaurea ramiflora | 0 | 1 | 0 | 0 | 0 | 0 |
| Bischofia javanica | 0 | 1 | 0 | 1 | 0 | 0 |
| Bischofia javanica | 0 | 1 | 0 | 0 | 0 | 0 |
| Breynia rostrata | 0 | 1 | 0 | 1 | 0 | 0 |
| Bridelia balansae | 0 | 1 | 1 | 1 | 0 | 1 |
| Bridelia tomentosa | 1 | 1 | 1 | 1 | 0 | 0 |
| Cleistanthus sumatranus | 0 | 1 | 0 | 0 | 0 | 0 |
| Glochidion sphaerogynum | 0 | 1 | 0 | 0 | 0 | 0 |
| Glochidion wrightii | 0 | 1 | 0 | 0 | 0 | 0 |
| Glochidion zeylanicum | 0 | 1 | 0 | 1 | 0 | 0 |









3.3. Glandular Trichomes (GTs)

3.4. Shape and Anticlinal Wall Pattern of Epidermal Cells

3.5. Types of Stomata
| Species | Epidermal Cell Shape | Anticlinal Wall Pattern | Stomata Type |
|---|---|---|---|
| Acalyphoideae | |||
| Acalypha hispida | Polygonal | Straight/sinus | Anomocytic/ Anisocytic |
| Acalypha indica | Irregular | sinus | Anomocytic |
| Acalypha wilkesiana | Polygonal | Straight | Anisocytic |
| Alchornea rugosa | Irregular | Undulate | Anomocytic |
| Macaranga corymbosa | Irregular | Sinus | Anomocytic |
| Mallotus apelta | Irregular | Deeply undulate | Not visible |
| Mallotus barbatus | Irregular | Straight/sinus | Anisocytic |
| Mallotus paniculatus | Irregular | Straight/undulate | Paracytic |
| Ricinus communis | Irregular/elongated | undulate | Anisocytic |
| Euphorbioideae | |||
| Euphorbia heterophylla | Irregular | Sinus | Anomocytic/Anisocytic |
| Excoecaria cochinchinensis | Irregular | Deeply undulate | Anisocytic |
| Hura crepitans | Irregular | Undulate | Anomocytic |
| Triadica cochinchinensis | Irregular | Sinus | Anomocytic |
| Triadica sebifera | Polygonal/irregular | Straight | Anomocytic |
| Crotonoideae | |||
| Aleurites moluccana | Polygonal/irregular | Straight/sinus | Paracytic |
| Codiaeum variegatum (I) | Irregular | Undulate | Paracytic |
| Codiaeum variegatum (II) | Irregular | Undulate | Paracytic |
| Codiaeum variegatum (III) | Irregular | Deeply undulate | Paracytic |
| Croton flavens | Irregular | Undulate | Not visible |
| Croton hirtus | Irregular | Slightly wavy | Anomocytic/Anisocytic/ Paracytic |
| Endospermum chinense | Polygonal/Irregular | Straight/Sinus | Paracytic |
| Hevea brasiliensis | Irregular | Sinus | Anomocytic |
| Jatropha integerrima (I) | Polygonal | Straight | Paracytic |
| Jatropha integerrima (II) | Polygonal | Straight | Paracytic |
| Manihot esculenta | Irregular | Deeply undulate | Anomocytic |
| Suregada glomerulata | Polygonal | Sinus | Anomocytic/Paracytic |
| Suregada multiflora | Irregular | Undulate | Anomocytic/Paracytic |
| Vernicia montana | Irregular | Sinus | Paracytic |
| Phyllanthaceae | |||
| Antidesma bunius | Irregular | Undulate | Anisocytic/Paracytic |
| Antidesma montanum (I) | Irregular | Sinus | Paracytic |
| Antidesma montanum (II) | Irregular | Undulate | Anomocytic/Paracytic |
| Aporosa dioca | Irregular | Sinus | Paracytic |
| Baccaurea ramiflora | Irregular | Deeply undulate | Anisocytic |
| Bischofia javanica (I) | Irregular | Undulate/Sinus | Diacytic/Anomocytic |
| Bischofia javanica (II) | Irregular | Undulate | Anomocytic |
| Breynia rostrata | Polygonal/irregular | Straight/Sinus | Paracytic |
| Bridelia balansae | Polygonal | Straight | Paracytic |
| Bridellia tomentosa | Irregular | Sinus | Paracytic |
| Cleistanthus sumatranus | Irregular | Undulate | Paracytic |
| Glochidion sphaerogynum | Polygonal/Irregular | Straight/Undulate | Anomocytic |
| Glochidion zeylanicum | Irregular | Undulate | Anomocytic |
| Glochidion wrightii | Irregular | Undulate | Paracytic |



| PCs | Eigenvalue | Percentage of Variance | Cumulative |
|---|---|---|---|
| 1 | 3.10932 | 77.73% | 77.73% |
| 2 | 0.57706 | 14.43% | 92.16% |
| 3 | 0.25408 | 6.35% | 98.51% |
| 4 | 0.05955 | 1.49% | 100.00% |
| Species | PC-1 (77.73%) | PC-2 (14.43%) | PC-3 (6.35%) | PC-4 (1.49%) |
|---|---|---|---|---|
| A.wil | 1.52312 | 1.58072 | −0.66743 | 0.21249 |
| A.rug | −0.65164 | −0.01798 | −0.25137 | −0.26137 |
| M.ape | −4.03681 | 0.18294 | 0.04889 | −0.05892 |
| M.pan | −1.00382 | −0.3487 | 0.35307 | −0.05137 |
| R.com | 0.40321 | 0.54186 | −0.40497 | 0.2036 |
| A.ind (I) | 1.71856 | 0.39953 | −0.45165 | 0.10073 |
| A.his | −1.19026 | −0.11769 | −0.17539 | −0.37122 |
| M.cor | 0.08357 | −1.1263 | 0.37629 | 0.16677 |
| M.bar | −4.03681 | 0.18294 | 0.04889 | −0.05892 |
| A.ind (II) | −1.43538 | 0.43737 | −0.17877 | 0.06437 |
| C.var (I) | 0.19273 | −0.21967 | 0.76075 | 0.37488 |
| J.int (I) | −0.73004 | 0.48071 | −0.13033 | −0.08692 |
| S.mul | −1.11482 | 0.25724 | −0.26761 | 0.09307 |
| E.chi (I) | 2.45294 | 2.0738 | 0.2234 | 0.08971 |
| C.var (II) | 1.70031 | 0.24545 | 0.68709 | −0.48597 |
| C.var (III) | 0.22842 | −0.82149 | 0.87356 | −0.17897 |
| H.bra | 4.61258 | −1.85297 | −1.12377 | −0.14421 |
| J.int (II) | 0.06966 | −0.57359 | 0.00296 | −0.28261 |
| E.chi (II) | 0.25567 | −0.96533 | −0.01172 | −0.05042 |
| A.mal | 1.91674 | 0.42808 | 0.92577 | 0.44469 |
| S.glo | 0.14414 | −0.59591 | 0.92228 | 0.16958 |
| M.esc | 0.29184 | 0.80224 | −0.74276 | −0.01195 |
| C.hir | −0.80823 | 1.27168 | 0.63454 | −0.12447 |
| C.fla | −4.03681 | 0.18294 | 0.04889 | −0.05892 |
| V.mon | 0.87495 | −0.61808 | 0.75408 | −0.08145 |
| S.dis | −1.26875 | −0.46528 | −0.42473 | 0.16515 |
| H.cre | −0.29005 | 0.02119 | −0.10468 | 0.16743 |
| T.sed | 2.26342 | 0.0724 | 0.15401 | −0.66746 |
| T.coc | −0.93156 | −0.25398 | −0.30908 | −0.24724 |
| A.dio | 0.8871 | −1.35552 | 0.40362 | 0.47264 |
| B.ros | −0.60293 | −0.57018 | −0.1199 | −0.07701 |
| C.sum | 1.54006 | −0.67051 | −0.64211 | 0.35876 |
| B.jav (I) | −0.64835 | −0.38895 | 0.04582 | 0.16382 |
| B.bal | −0.72521 | −0.15805 | 0.38216 | 0.12298 |
| A.bun | 3.35165 | 1.51595 | 0.22418 | 0.16267 |
| G.wri | 0.51657 | 0.02581 | −0.33469 | −0.32235 |
| G.sph | −0.45826 | −0.49411 | −0.62705 | 0.3222 |
| A.mon (1) | 0.77023 | −0.06024 | 0.72106 | −0.11255 |
| B.jav (II) | 0.07897 | −0.32685 | −0.70857 | 0.04106 |
| B.tom (I) | −1.7074 | 0.66884 | −0.26058 | 0.13338 |
| B.tom (II) | −1.89203 | 0.09449 | −0.39259 | 0.17135 |
| G.zal | −0.06004 | 0.22395 | −0.38608 | −0.25997 |
| A.mon (II) | 1.75277 | 0.31122 | 0.12455 | −0.20706 |


4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Webster, G.L. The genera of Euphorbiaceae in the southeastern United States. J. Arnold Arbor. 1967, 48, 363–430. [Google Scholar] [CrossRef]
- Webster, G.L. The saga of the spurges. A review of classification and relationships in the Euphorbiales. Bot. J. Linn. Soc. 1987, 94, 3–46. [Google Scholar] [CrossRef]
- Hans, A.S. Chromosomal conspectus of the Euphorbiaceae. Taxon 1973, 22, 591–636. [Google Scholar] [CrossRef]
- Perveen, A.; Qaiser, M. Pollen flora of Pakistan-xlvii. Euphorbiaceae. Pak. J. Bot. 2005, 37, 785. [Google Scholar]
- Lai, X.Z.; Yang, Y.B.; Shan, X.L. The investigation of Euphorbiaceous medicinal plants in Southern China. Econ. Bot. 2004, 58, 307–320. [Google Scholar] [CrossRef]
- Editorial Committee of Flora of China. Flora of China: Euphorbiaceae; China Science Press: Beijing, China, 1994; Volume 44, pp. 1–3. [Google Scholar]
- Webster, G.L. Synopsis of the genera and suprageneric taxa of Euphorbiaceae. Ann. Mo. Bot. Gard. 1994, 81, 33–144. [Google Scholar] [CrossRef]
- Angiosperm Phylogeny Group; Chase, M.W.; Christenhusz, M.J.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Wurdack, K.J.; Hoffmann, P.; Samuel, R.; de Bruijn, A.; van der Bank, M.; Chase, M.W. Molecular phylogenetic analysis of Phyllanthaceae (Phyllanthoideae pro parte, Euphorbiaceae sensu lato) using plastid rbcL DNA sequences. Am. J. Bot. 2004, 91, 1882–1900. [Google Scholar] [CrossRef]
- Wurdack, K.J.; Chase, M.W. Molecular systematics of Euphorbiaceae sensu lato using rbcL sequence data. Am. J. Bot. 1996, 83, 203. [Google Scholar]
- Soltis, D.E.; Soltis, P.S.; Nickrent, D.L.; Johnson, L.A.; Hahn, W.J.; Hoot, S.B.; Sweere, J.A.; Kuzoff, R.K.; Kron, K.A.; Chase, M.W.; et al. Angiosperm phylogeny inferred from 18S ribosomal DNA sequences. Ann. Mo. Bot. Gard. 1997, 84, 1–49. [Google Scholar] [CrossRef]
- Savolainen, V.; Chase, M.W.; Hoot, S.B.; Morton, C.M.; Soltis, D.E.; Bayer, C.; Fay, M.F.; de Bruijn, A.Y.; Sullivan, S.; Qiu, Y.L. Phylogenetics of flowering plants based on combined analysis of plastid atpB and rbcL gene sequences. Syst. Biol. 2000, 49, 306–362. [Google Scholar] [CrossRef]
- Savolainen, V.; Fay, M.F.; Albach, D.C.; Backlund, A.; van der Bank, M.; Cameron, K.M.; Johnson, S.A.; Lledó, M.D.; Pintaud, J.C.; Powell, M.; et al. Phylogeny of the eudicots: A nearly complete familial analysis based on rbcL gene sequences. Kew Bull. 2000, 55, 257–309. [Google Scholar] [CrossRef]
- Tokuoka, T.; Tobe, H. Molecular phylogeny of subfamily Phyllanthoideae (Euphorbiaceae s.L.) and its evolution of the seed coat structure. J. Plant Res. 2002, 115, 42. [Google Scholar]
- Kathriarachchi, H.; Hoffmann, P.; Samuel, R.; Wurdack, K.J.; Chase, M.W. Molecular phylogenetics of Phyllanthaceae inferred from five genes (plastid atpB, matK, 3′ ndhF, rbcL, and nuclear PHYC). Mol. Phylogenetic Evol. 2005, 36, 112–134. [Google Scholar] [CrossRef]
- Kathriarachchi, H.; Samuel, R.; Hoffmann, P.; Mlinarec, J.; Wurdack, K.J.; Ralimanana, H.; Stuessy, T.F.; Chase, M.W. Phylogenetics of tribe Phyllantheae (Phyllanthaceae; Euphorbiaceae sensu lato) based on nrITS and plastid matK DNA sequence data. Am. J. Bot. 2006, 93, 637–655. [Google Scholar] [CrossRef]
- Samuel, R.; Kathriarachchi, H.; Hoffmann, P.; Barfuss, M.H.; Wurdack, K.J.; Davis, C.C.; Chase, M.W. Molecular phylogenetics of Phyllanthaceae: Evidence from plastid matK and nuclear PHYC sequences. Am. J. Bot. 2005, 92, 132–141. [Google Scholar] [CrossRef]
- Wurdack, K.J.; Davis, C.C. Malpighiales phylogenetics: Gaining ground on one of the most recalcitrant clades in the angiosperm tree of life. Am. J. Bot. 2009, 96, 1551–1570. [Google Scholar] [CrossRef]
- Huber, H. Angiospermen-Leitfaden Durch Die Ordnungen Und Familien Der Bedecksamer; Gustav Fischer: Stuttgart, Germany, 1991; 160p. [Google Scholar]
- Jensen, U.; Vogel-Bauer, I.; Nitschke, M. Legumin like proteins and thesystematics of the Euphorbiaceae. Ann. Mo. Bot. Gard. 1994, 81, 60–179. [Google Scholar] [CrossRef]
- Vogel, C. Phytoserologische Untersuchungen zur Systematik der Euphorbiaceae; Beitrage zur infrafamiliaren Gliederung und zu Beziehungen im extrafamiliaren Bereich. Dissert Bot. 1986, 98, 1–124. [Google Scholar]
- Beutler, J.A.; Lindner, A.B.A.; McCloud, T.G. Further studies on phorbol ester bioactivity in the Euphorbiaceae. Ann. Mo. Bot. 1996, 83, 530–533. [Google Scholar] [CrossRef]
- Seigler, D.S. Phytochemistry and systematics of the Euphorbiaceae. Ann. Mo. Bot. Gard. 1994, 81, 380–401. [Google Scholar] [CrossRef]
- Moawed, M.M.; Saaid, S.; Abdelsamie, Z.; Tantawy, M. Phenetic Analysis of Certain Taxa of Euphorbiaceae Grown in Egypt. Egypt. J. Bot. 2015, 55, 247–267. [Google Scholar]
- Ahmad, K.; Khan, M.A.; Ahmad, M.; Shaheen, N.; Nazir, A. Taxonomic diversity in epidermal cells of some sub-tropical plant species. Int. J. Agric. Biol. 2010, 12, 115–118. [Google Scholar]
- Zahra, N.B.; Ahmad, M.; Shinwari, Z.K.; Zafar, M.; Sultana, S. Systematic significance of anatomical characterization in some Euphorbiaceous species. Pak. J. Bot. 2014, 46, 1653–1661. [Google Scholar]
- Raju, V.S.; Rao, P.N. Variation in the structure and development of foliar stomata in the Euphorbiaceae. Bot. J. Linn. Soc. 1977, 75, 69–97. [Google Scholar] [CrossRef]
- Bahadur, S.; Taj, S.; Long, W.; Ahmad, M. Pollen morphology and its implication in the taxonomy of some selected taxa of the bi and tri-ovulate Euphorbiaceae of the Hainan Island by using multiple microscopic techniques. Microsc. Res. Tech. 2022, 85, 2045–2060. [Google Scholar] [CrossRef]
- Solereder, H. Systematic Anatomy of the Dicotyledons: A Handbook for Laboratories of Pure and Applied Botany; Clarendon Press: Oxford, UK, 1908; Volume 2. [Google Scholar]
- Thakur, H.A.; Patil, D.A. The foliar epidermal studies in some hitherto unstudied Euphorbiaceae. Curr. Bot. 2011, 2, 22–30. [Google Scholar]
- Vitarelli, N.C.; Riina, R.; Caruzo, M.B.R.; Cordeiro, I.; Fuertes-Aguilar, J.; Meira, R.M. Foliar secretory structures in Crotoneae (Euphorbiaceae): Diversity, anatomy, and evolutionary significance. Am. J. Bot. 2015, 102, 833–847. [Google Scholar] [CrossRef]
- Ahmad, M.; Zafar, M.; Bahadur, S.; Sultana, S.; Taj, S.; Celep, F.; Majeed, S. Palynomorphological diversity among the Asteraceous honeybee flora: An aid to the correct taxonomic identification using multiple microscopic techniques. Microsc. Res. Tech. 2022, 85, 570–590. [Google Scholar]
- Bahadur, S.; Taj, S.; Ahmad, M.; Zafar, M.; Gul, S.; Shuaib, M.; Romman, M. Authentication of the therapeutic Lamiaceae taxa by using pollen traits observed under scanning electron microscopy. Microsc. Res. Tech. 2022, 85, 2026–2044. [Google Scholar] [CrossRef]
- Bahadur, S.; Taj, S.; Long, W.; Hanif, U. Pollen Morphological Peculiarities of Selected Mimosoideae Taxa of Hainan Island and Their Taxonomic Relevance. Agronomy 2022, 12, 1122. [Google Scholar] [CrossRef]
- Bahadur, S.; Taj, S.; Long, W.; Ahmad, M. Pollen morphology and its implication in the taxonomy of some selected tribes of the Asteraceae of Hainan Island South China. Bot. Rev. 2022, 88, 1–28. [Google Scholar] [CrossRef]
- Celep, A.; Tugay, O.; Dural, H.; Ulukuş, D.; Bahadur, S.; Celep, F. Comparative root and leaf anatomy of the Turkish Iris subgenus Scorpiris and their taxonomic and phylogenetic utility. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2022, 1–11. Available online: https://www.tandfonline.com/doi/abs/10.1080/11263504.2022.2048275 (accessed on 5 September 2022). [CrossRef]
- Ashfaq, S.; Zafar, M.; Ahmad, M.; Sultana, S.; Bahadur, S.; Khan, A.; Shah, A. Microscopic investigations of palynological features of convolvulaceous species from arid zone of Pakistan. Microsc. Res. Tech. 2018, 81, 228–239. [Google Scholar] [CrossRef]
- Senanayake, Y.D.A.; Samarnayake, P. Intraspecific variation of stomatal density in Hevea brasiliensis Muell. Q. J. Rubber Res. Inst. Ceylon 1970, 46, 61–68. [Google Scholar]
- Williams, C.N. Growth and productivity of tapioca (Manihot utilissimu). Stomatal functioning and yield. Exp. Agric. 1971, 7, 49–62. [Google Scholar] [CrossRef]
- Aworinde, D.O.; Nwoye, D.U.; Jayeola, A.A.; Olagoke, A.O.; Ogundele, A.A. Taxonomic significance of foliar epidermis in some members of euphorbiaceae family in Nigeria. Res. J. Bot. 2009, 4, 17–28. [Google Scholar] [CrossRef]
- James, O.E.; Green, B.O.; Ajuru, M.G.; Wilson, V. Anatomical Characterization of Roots Of Some Medicinal Plants Of The Family Euphorbiaceae in Nigeria. Asian J. Plant Soil Sci. 2022, 7, 51–55. [Google Scholar]
- Haegens, R.M.A.P. Taxonomy, phylogeny, and biogeography of Baccaurea, Distichirhops, and Nothobaccaurea (Euphorbiaceae). Blumea. Suppl. 2000, 12, 1–218. [Google Scholar]
- Elumalai, R.; Selvaraj, R.; Arunadevi, R.; Chidambaram, A.L.A. Foliar anatomical studies of some taxa of Euphorbiaceae. Afr. J. Plant Sci. 2014, 8, 271–277. [Google Scholar]
- Barthlott, W. Epidermal and seed surface characters of plants: Systematic applicability and some evolutionary aspects. Nord. J. Bot. 1981, 1, 345–355. [Google Scholar] [CrossRef]
- Verdus, M.-C. Contribution i l’ttude des plantules d’EuphorbiacCes. Trav. Du Lab. For. De Toulouse 1970, 8, 1–65. [Google Scholar]
- Alyas, T.; Shaheen, S.; Amber, U.; Harun, N.; Khalid, S.; Hussain, K.; Khan, F. Applications of scanning electron microscopy in taxonomy with special reference to family Euphorbiaceae. Microsc. Res. Tech. 2020, 83, 1066–1078. [Google Scholar] [CrossRef]
- Kakkar, L.; Paliwal, G.S. Studies on the leaf anatomy of Euphorbia: II. Venation patterns. Bot. J. Linn. Soc. 1974, 68, 173–208. [Google Scholar]
- Bercu, R.; Popoviciu, D.R. Comparative anatomical study on leaves of three Euphorbia L. species. Wulfenia 2015, 22, 271–276. [Google Scholar]
- Talebi, S.M.; Noori, M.; Naniz, H.A. A study of epidermal leaf anatomy of 18 Euphorbia taxa from Kerman Province, Iran. Biologija 2017, 63, 2. [Google Scholar] [CrossRef][Green Version]
- Cardiel, J.M.; Montero-Muñoz, I.; Ortúñez, E.; Dorda, E.; Sancho-García, I. Epidermal crystals in Acalypha (Euphorbiaceae, Acalyphoideae) as a new taxonomic trait of the genus. Plant Syst. Evol. 2020, 306, 1–18. [Google Scholar] [CrossRef]
- Bhavana, R.; Ramya, R.; Binu, T. Comparative studies on morphology, anatomy and phytochemistry of selected species of Croton L. (Euphorbiaceae). Plant Arch. 2020, 20, 639–656. [Google Scholar]
- Han, W. Comparative Observation on Leaf Anatomy and Pollen of Vernicia Lour. and Aleurites JR et G. Forst. J. Syst. Evol. 1985, 23, 188. [Google Scholar]
- Olowokudejo, J.D. Comparative epidermal morphology of West African species of Jatropha L. (Euphorbiaceae). Bot. J. Linn. Soc. 1993, 111, 139–154. [Google Scholar] [CrossRef]
- Rao, A.N. Reticulate Cuticle on Leaf Epidermis in Hevea brasiliensis Muell. Nature 1963, 197, 1125–1126. [Google Scholar] [CrossRef]
- Mamoucha, S.; Tsafantakis, N.; Fokialakis, N.; Christodoulakis, N.S. Structural and phytochemical investigation of the leaves of Ricinus communis. Aust. J. Bot. 2016, 65, 58–66. [Google Scholar] [CrossRef]
- Suwal, B.M.S.; Gautam, R.S.; Manandhar, D. Environmental Impact on Morphological and Anatomical Structure of Ricinus communis L. Leaves Growing in Kathmandu, Nepal. Int. J. Appl. Sci. Biotechnol. 2019, 7, 274–278. [Google Scholar] [CrossRef][Green Version]
- Metcalfe, C.R.; Chalk, L. Anatomy of Dicotyledons; Clarendon Press: Oxford, UK, 1950; Volume 1–2, p. 1500. [Google Scholar]
- Wu, H.; Li, X.; Qian, H.; Chen, J. Improved partial trend method to detect rainfall trends in Hainan Island. Theor. Appl. Climatol. 2019, 137, 2539–2547. [Google Scholar] [CrossRef]
- Long, W.; Zang, R.; Wang, X.; Bahadur, S. Environmental Characteristics in Tropical Cloud Forests. In Tropical Cloud Forest Ecology in Hainan Island; Springer: Singapore, 2022; pp. 3–12. [Google Scholar]
- Yaseen, M.; Long, W.; Khalid, F.; Bahadur, S.; Noushahi, H.A. Shifts in Community Vegetative Organs and Their Dissimilar Trade-Off Patterns in a Tropical Coastal Secondary Forest, Hainan Island, Southern China. Diversity 2022, 14, 823. [Google Scholar] [CrossRef]
- Long, W.X.; Ding, Y.; Zang, R.G.; Yang, M.; Chen, S.W. Environmental characteristics of tropical cloud forests in the rainy season in Bawangling National Nature Reserve on Hainan Island, South China. Chin. J. Plant Ecol. 2011, 35, 137. [Google Scholar] [CrossRef]
- Long, W.; Yang, X.; Li, D. Patterns of species diversity and soil nutrients along a chrono sequence of vegetation recovery in Hainan Island, South China. Ecol. Res. 2012, 27, 561–568. [Google Scholar] [CrossRef]
- Wang, X.X.; Long, W.X.; Yang, X.B.; Xiong, M.H.; Kang, Y.; Huang, J.; Li, S.X. Patterns of plant diversity within and among three tropical cloud forest communities in Hainan Island. Chin. J. Plant Ecol. 2016, 40, 469. [Google Scholar]
- Ali, M.; Bahadur, S.; Hussain, A.; Saeed, S.; Khuram, I.; Ullah, M.; Akhtar, N. Foliar epidermal micromorphology and its taxonomic significance in Polygonatum (Asparagaceae) using scanning electron microscopy. Microsc. Res. Tech. 2020, 83, 1381–1390. [Google Scholar] [CrossRef]
- Nichodemus, C.O.; Ekeke, C. Morpho-anatomical and histological characters in the systematics of the Croton species (Euphorbiaceae: Crotonoideae) in Southern Nigeria. Hospital 2020, 29, 5. [Google Scholar]
- Webster, G.L. A provisional synopsis of the sections of the genus Croton (Euphorbiaceae). Taxon 1993, 42, 793–823. [Google Scholar] [CrossRef]
- Van Ee, B.W.; Berry, P.E. A phylogenetic and taxonomic review of Croton (Euphorbiaceae ss) on Jamaica including the description of Croton jamaicensis, a new species of section Eluteria. Syst. Bot. 2009, 34, 129–140. [Google Scholar] [CrossRef]
- Van Ee, B.W.; Berry, P.E. Taxonomy of Croton flavens (Euphorbiaceae), a Caribbean Endemic, and a Preliminary Subsectional Classification of Croton section Adenophylli. Caribb. J. Sci. 2021, 51, 314–357. [Google Scholar] [CrossRef]
- Webster, G.L.; Del-Arco-Aguilar, M.J.; Smith, B.A. Systematic distribution of foliar trichome types in Croton (Euphorbiaceae). Bot. J. Linn. Soc. 1996, 121, 41–57. [Google Scholar] [CrossRef]
- Senakun, C.; Chantaranothai, P. A morphological survey of foliar trichomes of Croton L.(Euphorbiaceae) in Thailand. Thai For. Bull. 2010, 38, 167–172. [Google Scholar]
- De Sá-Haiad, B.; Serpa-Ribeiro, A.C.C.; Barbosa, C.N.; Pizzini, D.; de O Leal, D.; de Senna-Valle, L.; de Santiago-Fernandes, L.D.R. Leaf structure of species from three closely related genera from tribe Crotoneae Dumort.(Euphorbiaceae ss, Malpighiales). Plant Syst. Evol. 2009, 283, 179–202. [Google Scholar] [CrossRef]
- Feio, A.C.; Meira, R.M.; Riina, R. Leaf anatomical features and their implications for the systematics of dragon’s blood, Croton section Cyclostigma (Euphorbiaceae). Bot. J. Linn. Soc. 2018, 187, 614–632. [Google Scholar] [CrossRef]
- Martins, M.B.G.; Zieri, R. Leaf anatomy of rubber-tree clones. Sci. Agric. 2003, 60, 709–713. [Google Scholar] [CrossRef][Green Version]
- Shaw, H.A. Notes on Malaysian and Other Asiatic. Euphorbiaceae. Kew Bull. 1966, 20, 379–415. [Google Scholar] [CrossRef]
- Kolawole, O.S.; Jimoh, M.A.; Yakubu, F.; Chukwuma, E.C. Taxonomic Value of the Leaf Micro-Morphology and Quantitative Phytochemistry of Jatropha Integerrima Jacq. and Jatropha podagrica Hook. (Euphorbiaceae)-Known Horticultural Plants in Nigeria. In Anales de Biología; Publications Service of the University of Murcia: Murcia, Spain, 2017; Volume 39, pp. 55–62. [Google Scholar]
- Norfaizal, G.M.; Khalijah, H.; Ruzi, A.M. Leaf anatomical study of five Macaranga species (Euphorbiaceae). J. Trop. Agric. Food Sci. 2012, 40, 289–296. [Google Scholar]
- Hussin, K.H.; Wahab, B.A.; Teh, C.P. Comparative leaf anatomical studies of some Mallotus Lour.(Euphorbiaceae) species. Bot. J. Linn. Soc. 1996, 122, 137–153. [Google Scholar] [CrossRef][Green Version]
- Kakkar, L.; Paliwal, G.S. Studies on the leaf anatomy of Euphorbia IV. Terminal idioblasts. Proc. Indian Natl. Sci. Acad. 1972, 40, 55–67. [Google Scholar]
- Jangid, P.P.; Gupta, S. Systematic wood anatomy of the tribe Phyllantheae (Phyllanthaceae, Euphorbiaceae sl) from India: Implication in reinstatement of Phyllanthus, Glochidion and allies. Nord. J. Bot. 2016, 34, 496–512. [Google Scholar] [CrossRef]
- Yao, G.; Zhang, D.X. Reinstatement of three names in Glochidion (Phyllanthaceae) based on morphological and palynological evidence. Nord. J. Bot. 2015, 33, 197–203. [Google Scholar] [CrossRef]
- Yao, G.; Li, Y.; Luo, S. New records of Glochidion species from several provinces in China. Acta Bot. Boreali-Occident. Sin. 2019, 39, 1322–1324. [Google Scholar]
- Schot, A.M. Systematics of Aporosa (Euphorbiaceae). Blumea. Suppl. 2004, 17, 1–380. [Google Scholar]
- Webster, G.L.; Ellis, J.R. Cytotaxonomic studies in the Euphorbiaceae, subtribe Phyllanthinae. Am. J. Bot. 1962, 49, 14–18. [Google Scholar] [CrossRef]
- Levin, G.A. Systematic foliar morphology of Phyllanthoideae (Euphorbiaceae). III. Cladistic analysis. Syst. Bot. 1986, 11, 515–530. [Google Scholar] [CrossRef]
- Stace, C.A. Plant Anatomy and Biosystematics; Edward Arnold Publishers: London, UK, 1980; 279p. [Google Scholar]
- Endress, P.K.; Baas, P.; Gregory, M. Systematic plant morphology and anatomy-50 years of progress. Taxon 2000, 49, 401–434. [Google Scholar] [CrossRef]
- Da Silva, M.J.; Alonso, A.A.; Dos Santos, I.S. A new Critically Endangered species of stone breaker (Phyllanthus, Phyllanthaceae) from Central Brazil, with notes on its leaf and stem anatomy. Phytotaxa 2022, 530, 53–64. [Google Scholar] [CrossRef]
- Pornpongrungrueng, P.; Parnell, J.A.; Hodkinson, T.R.; Chantaranothai, P. Phyllanthus kaweesakii (Phyllanthaceae), a new species from Thailand. Botany 2017, 95, 567–577. [Google Scholar] [CrossRef]
- Santiago, L.J.M.; Louro, R.P.; Emmerich, M. Phylloclade anatomy in Phyllanthus section Choretropsis (Phyllanthaceae). Bot. J. Linn. Soc. 2008, 157, 91–102. [Google Scholar] [CrossRef]
- Cutter, E.G. Plant Anatomy: Experiment and Interpretation: Part I: Cells and Tissues; Edward Arnold: Lincoln, UK, 1969. [Google Scholar]
- Essiett, U.A.; Illoh, H.C.; Udoh, U.E. Leaf epidermal studies of three species of Euphorbia in Akwa Ibom State. Adv. Appl. Sci. Res. 2012, 3, 2481–2491. [Google Scholar]
- Inamdar, J.A.; Gangadhara, M. Studies on the trichomes of some Euphorbiaceae. Feddes Repert. 1977, 88, 103–111. [Google Scholar] [CrossRef]
- Nurul-Aini, C.A.C.; Nur-Shuhada, T.; Rozilawati, S.; Fatin-Munirah, A.; Noraini, T.; Latiff, A. Comparative leaf anatomy of selected medicinal plants in Acanthaceae. Int. Med. J. 2018, 17, 17–23. [Google Scholar]
- Mueller, S. The taxonomic significance of cuticular patterns within the genus Vaccinium (Ericaceae). Am. J. Bot. 1966, 53, 633. [Google Scholar]
- Cutler, D.F.; Brandham, P.E. Experimental evidence for genetic control of leaf surface characters in hybrid Aloineae (Liliaceae). Kew Bull. 1977, 32, 23–32. [Google Scholar] [CrossRef]
| Species | Locality | Voucher Number | Latitude | Longitude | Meter above Sea Level |
|---|---|---|---|---|---|
| Subfamily Acalyphoideae | |||||
| Acalypha hispida Burm. f. | Haikou | SB-HN-101 | 20.062363 | 110.3186713 | 3 |
| Acalypha indica L. | Chang Jiang | SB-HN-110 | 19.33905 | 108.210 | 33 |
| Acalypha wilkesiana Müll. Arg. | Haikou, Wanning | SB-HN-115 | 18.69396 | 110.23170 | 118 |
| Alchornea rugosa (Lour.) Müll. Arg. | Wanning, Botanical Garden | SB-HN-131 | 18.69396 | 110.23170 | 90 |
| Macaranga corymbosa Müll. Arg. | SB-HN-145 | ||||
| Mallotus apelta Müll. Arg. | Wanning | SB-HN-135 | 18.69396 | 110.23170 | 118 |
| Mallotus barbatus Müll. Arg. | Sanya | SB-HN-136 | 18.26415 | 109.52084 | 119 |
| Mallotus paniculatus (Lam.) Müll. Arg. | Wanning | SB-HN-137 | 18.69396 | 110.23170 | 118 |
| Melanolepis multiglandulosa (Reinw. Ex Blume) Rchb. & Zoll. | Wanning | SB-HN-149 | 18.69396 | 110.23170 | 118 |
| Ricinus communis L. | Wanning | SB-HN-113 | 18.69396 | 110.23170 | 118 |
| Subfamily Euphorbioideae | |||||
| Euphorbia heterophylla L. | Chang Jiang | SB-HN-120 | 19.33905 | 108.210 | 33 |
| Excoecaria cochinchinensis Lour. | Sanya, Qiong Zhong | SB-HN-122 | 19.25343 | 109.38799 | 445 |
| Hura crepitans L. | Wanning | SB-HN-130 | 18.69396 | 110.23170 | 118 |
| Triadica cochinchinensis Lour. | Bawangling | SB-HN-140 | 19.112612 | 109.082207 | 650 |
| Triadica sebifera (L.) Small | Chang Jiang | SB-HN-142 | 19.33905 | 108.210 | 33 |
| Subfamily Crotonoideae | |||||
| Aleurites moluccanus Willd. | ChangJiang | SB-HN-10 | 19.33905 | 108.210 | 33 |
| Codiaeum variegatum (Linn.) Bl. (I) | Beihualing | SB-HN-16 | 19.00262 | 109.81523 | 471 |
| Codiaeum variegatum (Linn.) Bl. (II) | Wanning | SB-HN-23 | 18.69396 | 110.23170 | 90 |
| Codiaeum variegatum (Linn.) Bl. (III) | Sanya | SB-HN-121 | 18.27725 | 109.50888 | 12 |
| Croton flavens L. | ChangJiang | SB-HN-26 | 19.33905 | 108.210 | 33 |
| Croton hirtus L’Hér. | ChangJiang | SB-HN-27 | 19.33905 | 108.210 | 33 |
| Endospermum chinense Benth. | Haikou | SB-HN-29 | 20.062363 | 110.3186713 | 3 |
| Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg. | Wanning | SB-HN-30 | 18.69396 | 110.23170 | 90 |
| Jatropha integerrima Jacq. (I) | Haikou | SB-HN-9 | 20.062363 | 110.3186713 | 3 |
| Jatropha integerrima Jacq. (II) | Wanning | SB-HN-36 | 18.69396 | 110.23170 | 118 |
| Manihot esculenta Crants | Wanning | SB-HN-37 | 18.69396 | 110.23170 | 118 |
| Suregada glomerulata Baill. | Bawangling | SB-HN-141 | 19.112612 | 109.082207 | 650 |
| Suregada multiflora Baill. | Wanning | SB-HN-39 | 18.69396 | 110.23170 | 118 |
| Vernicia montana Lour. | Beihualing | SB-HN-17 | 19.00262 | 109.81523 | 471 |
| Family Phyllanthaceae | |||||
| Aporosa dioca (Roxb.) Müll. Arg. | Haikou | SB-HN-19 | 20.062363 | 110.3186713 | 3 |
| Antidesma bunius Wall. | Wanning | SB-HN-41 | 18.69396 | 110.23170 | 118 |
| Antidesma montanum Blume (I) | Wanning | SB-HN-49 | 18.69396 | 110.23170 | 90 |
| Antidesma montanum Blume (II) | Wanning | SB-HN-50 | 18.69396 | 110.23170 | 90 |
| Baccaurea ramiflora Lour. | Beihualing | SB-HN-51 | 19.00262 | 109.81523 | 471 |
| Bischofia javanica Blume (I) | Beihualing | SB-HN-53 | 19.00262 | 109.81523 | 471 |
| Bischofia javanica Blume (II) | Wanning | SB-HN-54 | 18.69396 | 110.23170 | 90 |
| Breynia rostrata Merr. | Wanning | SB-HN-57 | 18.69396 | 110.23170 | 90 |
| Bridelia balansae Tutcher | Wanning | SB-HN-58 | 18.69396 | 110.23170 | 90 |
| Bridelia tomentosa Blume | Wanning | SB-HN-60 | 18.69396 | 110.23170 | 90 |
| Cleistanthus sumatranus (Miq.) Müll. Arg. | Wanning | SB-HN-63 | 18.69396 | 110.23170 | 90 |
| Glochidion sphaerogynum (Müll. Arg.) Kurz | Wanning | SB-HN-64 | 18.69396 | 110.23170 | 90 |
| Glochidion wrightii Benth. | Wanning | SB-HN-65 | 18.69396 | 110.23170 | 90 |
| Glochidion zeylanicum (Gaertn.) A. Juss. | Bawangling | SB-HN-146 | 19.112612 | 109.082207 | 650 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bahadur, S.; Ahmad, M.; Long, W.; Yaseen, M.; Hanif, U. Leaf Epidermal Traits of Selected Euphorbiaceae and Phyllanthaceae Taxa of Hainan Island and Their Taxonomic Relevance. Diversity 2022, 14, 881. https://doi.org/10.3390/d14100881
Bahadur S, Ahmad M, Long W, Yaseen M, Hanif U. Leaf Epidermal Traits of Selected Euphorbiaceae and Phyllanthaceae Taxa of Hainan Island and Their Taxonomic Relevance. Diversity. 2022; 14(10):881. https://doi.org/10.3390/d14100881
Chicago/Turabian StyleBahadur, Saraj, Mushtaq Ahmad, Wenxing Long, Muhammad Yaseen, and Uzma Hanif. 2022. "Leaf Epidermal Traits of Selected Euphorbiaceae and Phyllanthaceae Taxa of Hainan Island and Their Taxonomic Relevance" Diversity 14, no. 10: 881. https://doi.org/10.3390/d14100881
APA StyleBahadur, S., Ahmad, M., Long, W., Yaseen, M., & Hanif, U. (2022). Leaf Epidermal Traits of Selected Euphorbiaceae and Phyllanthaceae Taxa of Hainan Island and Their Taxonomic Relevance. Diversity, 14(10), 881. https://doi.org/10.3390/d14100881