2. History of Previous Research
The earliest report of a fossil ostrich in the Late Tertiary Siwalik beds of India apparently was in a letter sent by the British palaeontologist Hugh Falconer to his French colleague Henri Marie Ducrotay de Blainville on 4 October 1847 ([
2], p. xxi). In it, Falconer “designated certain remains of the Sewalik birds, ‘
Struthio palaeindica’”. This name was never officially published and is therefore invalid. Falconer did not publish a description of the avian material from the Siwaliks at his disposal, but, as noted by Murchison [
2] when he edited Falconer’s works, he had a lithographic plate prepared showing various bird remains. Only a few prints were made, however, before the plate was destroyed, and they were not published. Murchison [
2] published the captions of the ill-fated plate (‘plate R’), in which no systematic identifications were provided for the specimens, although they included ostrich bones, as noted later by Davies [
3]. In his biographical sketch of Falconer, Murchison ([
2], p. xxviii) mentioned the ostrich among the fossil forms discovered by Falconer in the Siwaliks deposits. This seems to be the first published mention of fossil ostriches in the Indian subcontinent (and worldwide as well).
The name
Struthio asiaticus was coined by Alphonse Milne-Edwards in the second volume of his great work
Recherches anatomiques et paléontologiques pour servir à l’histoire des oiseaux fossiles de la France. The book is dated 1869–1871, and the name
Struthio asiaticus appears at the end of the second volume, which may explain why some authors (e.g., Lambrecht [
4]) give 1871 as the date for the taxon. Milne-Edwards’s mention of the Indian ostrich ([
1], vol. 2, p. 587), in a short section about the fossil birds from the Tertiary “subhimalayan” deposits, is very brief (my translation):
“One of the most remarkable species belonged to the brevipennate group, and is very close to the African ostrich by the conformation of its foot, which bore only two toes; but it was of smaller size than the latter: to distinguish it, one could call it Struthio asiaticus”.
Milne-Edwards gave no details about the material on which he based his observations. However, in the introduction to his book ([
1], vol. 1, p. 5), he mentioned that he had visited museums in London, including the British Museum, and it seems clear that this is where he saw the fossil material on which be based
Struthio asiaticus.
When Lydekker [
5] described avian remains from the Siwaliks, he mentioned Milne-Edwards’s
Struthio asiaticus. Although he did not understand on what material the latter had based his conclusion that this fossil ostrich was two-toed, he nevertheless accepted the conclusion of the French expert.
Davies [
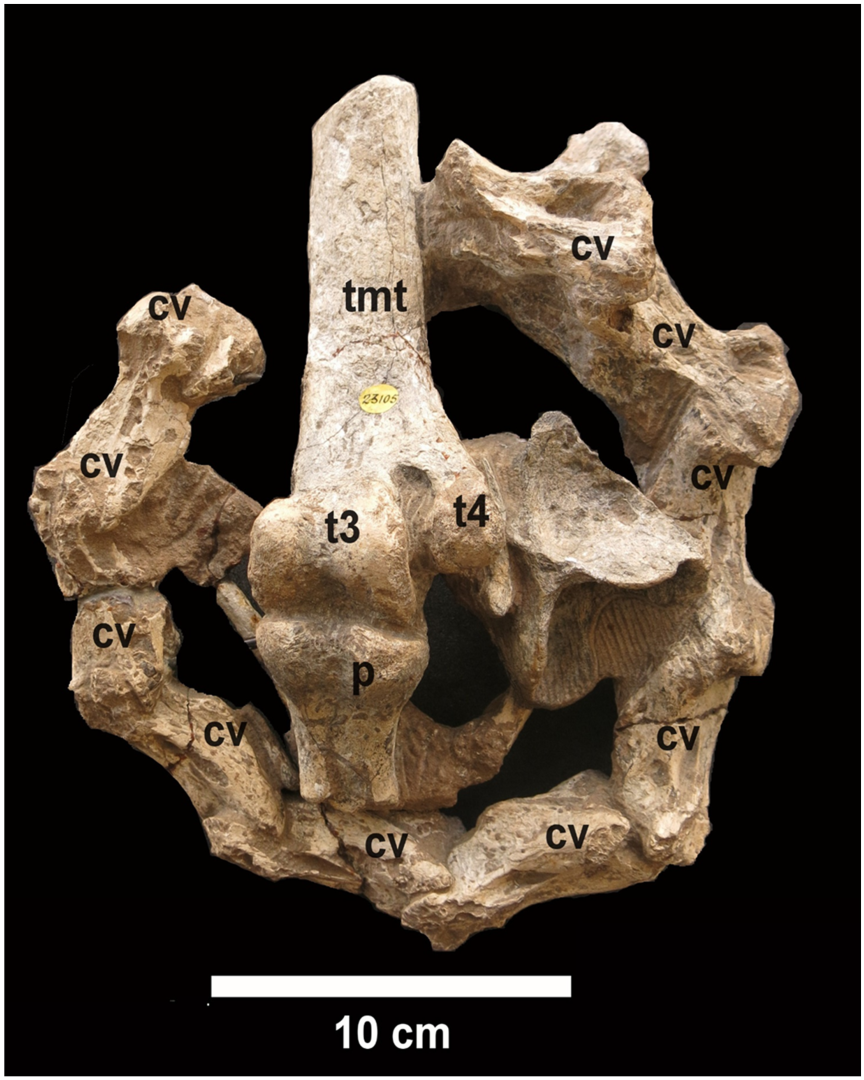
3] clarified the question when he described a specimen from the Siwaliks presented to the British Museum by Colonel Colvin in 1848, now kept at the Natural History Museum, London, under number NHMUK PV OR 23105. It consists of a block containing various avian bones, including a series of 10 articulated cervical vertebrae plus two detached vertebrae, the distal end of a left tarsometatarsus with the attached proximal end of the first phalanx of the third digit, and some elements of the wing; the recognizable wing elements are metacarpals, plus poorly preserved bones which may be the distal end of a radius and carpals, which are attached by matrix to the caudal face of the metatarsus and pedal phalanx (
Figure 1). Although Davies [
3] remarked that associations of bones of different taxa were not uncommon in the Siwaliks deposits, and despite the fact that Lydekker [
6] mentioned that the block also contained vertebrae and an incisor of an antelope, it seems very likely that the ostrich limb bones and cervical vertebrae belong together. Be that as it may, the tarsometatarsus clearly bears only two trochleae and the bird was definitely two-toed. As noted by Davies [
3], there can be no doubt that it is the specimen on the basis of which Milne-Edwards erected the species
Struthio asiaticus. This seems to have been taken for granted by various authors, including Lydekker [
6] and Brodkorb [
7]. Zelenkov and Kurochkin [
8] designated the carpometacarpal elements alone as the lectotype of
Struthio asiaticus. However, there is every reason to believe that all the ostrich bones catalogued as NHMUK PV OR 23105 belong to a single individual. Specimen NHMUK PV OR 23105, consisting of an incomplete tarsometatarsus and associated incomplete first phalanx, together with other ostrich bones including a series of cervical vertebrae and forelimb elements, is therefore designated here as the lectotype of
Struthio asiaticus Milne-Edwards, 1869–1871.
3. Other Purported or Real Ratites from the Siwaliks
In addition to
Struthio asiaticus, other fossils from the Siwaliks have been referred to ratite birds. Lydekker [
5] thus referred four phalanges from the Siwaliks of the western Punjab (in today’s Pakistan) to a fossil emu which he named
Dromaeus sivalensis. Davies [
3] accepted this identification and considered that an isolated phalanx (already mentioned by Lydekker) indicated the presence of a third ratite, different from both the ostrich and the emu, in the Siwaliks beds. Lydekker [
9] concurred with Davies’s opinion. However he rapidly changed his mind after examining mammalian remains from the Siwaliks and published a very brief note [
10] in which he explained that the purported ratite phalanges he had previously described and named
Dromaeus sivalensis actually belonged to a
Hippopotamus-like artiodactyl, concluding that ‘the genus
Dromaeus must therefore be expunged from the Siwalik fauna’ ([
10], p. 238).
The phalanx referred to a third ratite by Davies [
3] and Lydekker [
9] was considered by Lydekker [
6] as belonging to a cassowary-like bird for which he coined the name
Hypselornis sivalensis. The specimen was re-examined by Lowe [
11] who, after detailed comparisons, concluded that it belonged to a large crocodile.
Lambrecht [
4] duly noted that both
Dromaeus sivalensis and
Hypselornis sivalensis were based on non-avian material.
In this connection, it may be mentioned that the taxon
Megaloscelornis sivalensis, erected by Lydekker [
5] and initially considered by him as a gigantic wader-like carinate bird, turned out to be based on a fragment of a turtle shell (originally mistaken for an avian sternum) and on a tibiotarsus belonging to
Struthio asiaticus (according to Lydekker [
9]).
It should be noted that several ostrich or ostrich-like bones from other Siwalik localities than that of the type specimen of
Struthio asiaticus were described by Lydekker [
5,
9] and Davies [
3]. Some of them later turned out to be non-avian (see above), but the distal end of a right tibiotarsus (NHMUK PV OR 39732) described by Davies [
3] and Lydekker [
9], as well as a right tibiotarsus missing both ends, with a fragment of the fibula, described by Lydekker [
9] and kept in the Indian Museum (Calcutta), did belong to ostriches. This material was illustrated by Lydekker ([
9], plate 15), according to whom they came from the “typical Siwalik hills”, not from the Punjab, which suggests that they came from present-day India, not from Pakistan.
To sum up, all the genuine ratite bones from the Siwaliks can apparently be referred to ostriches. Whether they all belong to Struthio asiaticus, as defined on the basis of the type specimen, is a moot point because they cannot be directly compared with it and come from different localities of uncertain age.
4. Geographical and Stratigraphical Origin of the Specimen
Very little information is available about the exact provenance of the type of
Struthio asiaticus, beyond the fact that it was found in the Siwalik hills of India (bearing in mind that at the time of the discovery, “India” included present-day Pakistan). Lydekker [
9] noted that the specimens of
Struthio asiaticus came from the “typical Siwalik Hills”, as opposed to the “Siwaliks of the Punjab”. This suggests that the type specimen came from present-day northern India rather than Pakistan. Brodkorb [
7] noted that it was found “probably near Hardwar” (Haridwar, in Uttarakhand State, northern India); this, however, is based on a remark in Murchison’s biography of Falconer [
2], in which “Hurdwar” is mentioned as an area where Falconer and his associates discovered abundant Siwaliks fossils. However, the type of
Struthio asiaticus was not found by Falconer and his group, but by Colonel John Colvin [
3]. Colvin [
12] provided details about the geographical origin of some of the fossils in his collection: they were found south of the “Chur” (Churdhar) Peak, in present-day Himachal Pradesh (India), the western limit of the prospections being between the cities of Nahan and Pinjor (Pinjor is the type locality of the Pinjor Formation, part of the Siwalik Group, of Pleistocene age). Whether the type specimen of
Struthio asiaticus came from that area is likely. It is not possible to provide more accurate geographical information. As Colvin [
12] explained, the fossils were found by native collectors sent by him into the hills, who picked up whatever they found and heaped all together. Therefore, because of the lack of accurate information, it seems very difficult to ascertain the exact geographical provenance of the specimen beyond the fact that in all likelihood it came from present-day India rather than Pakistan.
As shown by numerous finds of eggshell fragments, ostriches were present on the Indian subcontinent over a long period of time, from the Miocene to the Late Pleistocene [
13,
14,
15,
16]. Ostrich eggshell fragments, for instance, are known from a 10.1 Ma old site, referred to the Dhok Pathan Formation, at Haritalyangar in India [
15]. At the other end of the stratigraphical range, eggshell fragments have been widely used in India by Late Paleolithic men for decorative purposes and ostriches are apparently represented in Indian Paleolithic rock art [
17]. An accurate placement of the type specimen of
Struthio asiaticus would therefore be important for a reliable assessment of its evolutionary and biogeographical significance. However, the precise stratigraphical origin of the specimen is difficult to ascertain. When describing it, Davies [
3] and Lydekker [
9] noted that it came from the Siwalik hills, without specifying any geological formation. Davies mentioned the “Upper Miocene or Lower Pliocene period”, reflecting the general opinion about the age of the Siwaliks formations at that time. Lydekker [
6] was more specific, indicating the “Pliocene of the Siwalik Hills”, but again this does not imply that the specimen really is Pliocene in age, since age attributions of Siwalik fossil-bearing formations suggested in the 1890s are now outdated. Following Pilgrim’s pioneering work [
18], the fossil-bearing deposits of the Siwaliks (
sensu lato) have been subdivided into a succession of formations and many attempts have been made to correlate them with the standard stratigraphical scale, using both their faunal content and (more recently) magnetostratigraphy (see [
19] for a review).
Harrison and Walker [
20] stated that the fossil bird material from the Siwaliks, including the ostrich specimens, ‘was originally said to be Lower Pliocene in age but is now regarded as Upper Pliocene to Early Pleistocene’. However, as noted by Mikhailov and Zelenkov [
21], they did not give any reason for this age attribution.
Some authors [
15,
22,
23] have claimed that
Struthio asiaticus is from the Dhok Pathan Formation (upper Miocene). According to Patnaik et al. ([
15], p. 1486), “as early as 1884, Lydekker described skeletal remains (phalangeal bones, fibula, tibiotarsus and about 12 vertebrae) of
Struthio asiaticus Milne-Edward 1871 from an unspecified locality in the Upper Siwaliks (late Miocene Dhok Pathan Formation)”. In fact, as noted above, Lydekker [
9] did not specify that the specimens were from the Upper Siwaliks. Moreover, contrary to the opinion of Patnaik et al. [
15], to judge from Lydekker’s 1884 paper (see above), it seems that the type material of
Struthio asiaticus came from the Siwaliks of northern India, not from present-day Pakistan. Although this stratigraphical assignment was accepted by Mikhailov and Zelenkov [
21], the reason why
Struthio asiaticus was considered as coming from the Dhok Pathan Formation is therefore unclear and not solidly documented—all the more so that the formation names (including the Dhok Pathan Formation) used in Pakistan are not necessarily used in India [
19]. Eggshell fragments referred to
Struthio? have indeed been reported from the Dhok Pathan Formation at Hasnot, Pakistan [
24], but this of course does not demonstrate that the type of
Struthio asiaticus also came from that formation, since, as noted above, fragments of ostrich eggs are known from formations of widely different ages in India and Pakistan (the meaning of differences in the eggshell surface is beyond the scope of this paper and will not be discussed here—see Mikhailov and Zelenkov [
21] for in-depth discussions of fossil ostrich eggshells). Widrig and Field [
25] were clearly on the safe side when they mentioned the Dhok Pathan Formation with a question mark as the geological unit having yielded
Struthio asiaticus.
To sum up, the original papers about the type specimen of Struthio asiaticus were rather vague about both its geographical and its stratigraphical provenance. However, it seems clear that it came from the Siwaliks of present-day northern India rather than from Pakistan. Whether it originated from the Dhok Pathan Formation (or an Indian equivalent), as assumed by various recent authors, is in fact difficult to demonstrate. The fossil-bearing formations of the Siwaliks range in age from Miocene to Pleistocene, A close examination of the matrix (a hard brownish sandstone) of the type specimen by experts on the lithology of the Siwaliks formations might help to elucidate the question.
5. Systematic Palaeontology
Aves Linnaeus, 1758
Palaeognathae Pycraft, 1900
Struthionidae Vigors, 1825
Genus Struthio Linnaeus, 1758
Struthio asiaticus Milne-Edwards, 1869–1871
Type specimen: NHMUK PV OR 23105, a group of bones comprising the distal end of a left tarsometatarsus with the articulated proximal end of the first phalanx of the third toe, twelve partly articulated cervical vertebrae and poorly preserved bones of the forelimb.
Locus typicus: Siwalik hills of northern India.
Stratum typicum: uncertain, probably Neogene.
Diagnosis: a species of Struthio similar in size to Struthio camelus but differing from the living species by the greater robustness of its cervical vertebrae and apparently by shorter toes.
Description
A generally accurate description of the type specimen was provided by Davies [
3] and only a brief redescription is needed here. The most significant elements are the articulated distal end of the tarsometatarsus and the incomplete first phalanx of the third toe (
Figure 1). The tarsometatarsus fragment bears only two, fairly well-preserved, trochleae and is generally similar to the corresponding part of the tarsometatarsus of living
Struthio. Only the proximal half of the phalanx is preserved. It does not differ significantly from the corresponding part of a modern
Struthio phalanx; however, there is a hint of a distal expansion at a relatively short distance distalwards, which suggests that the phalanx was relatively shorter than in extant ostriches.
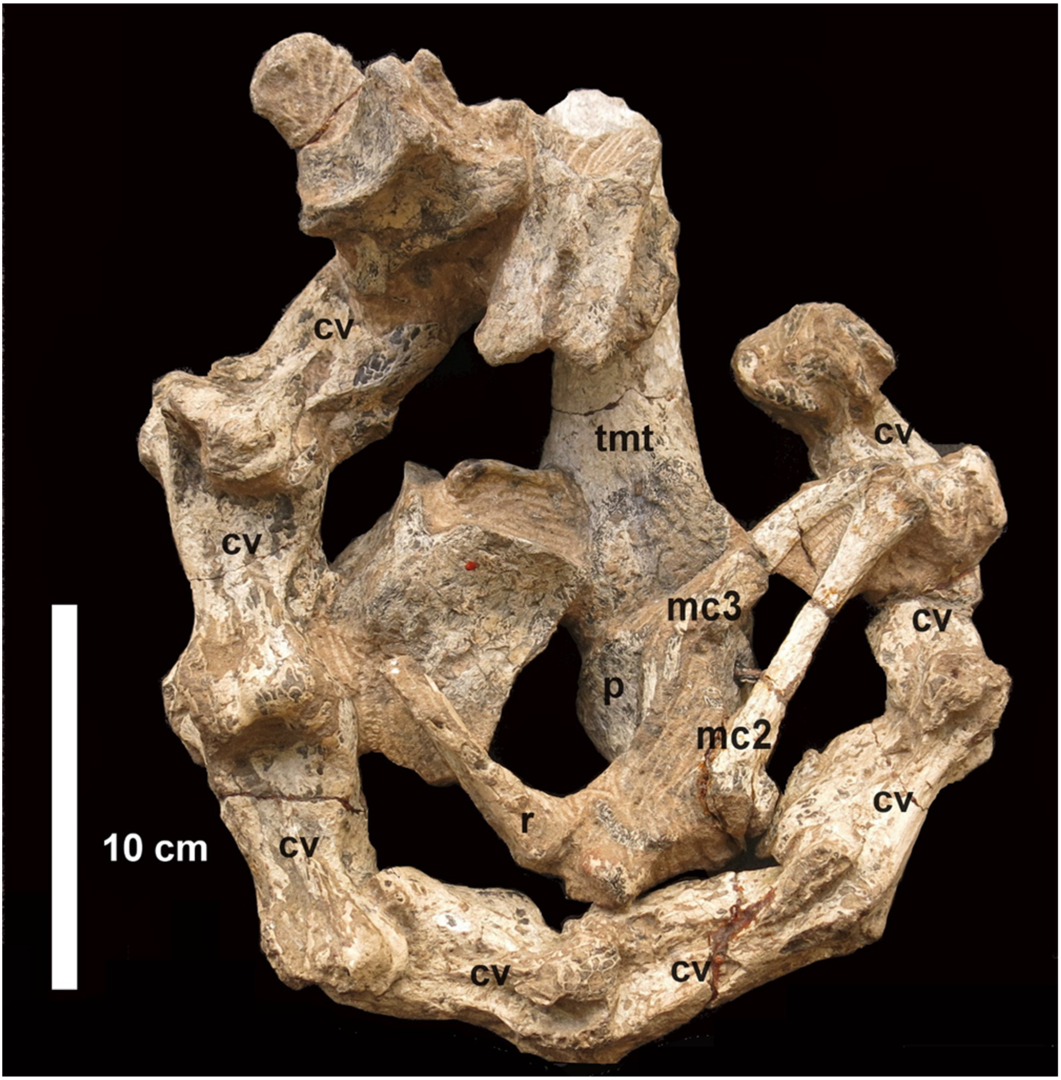
Posterior to the tarsometatarsus and phalanx, some rather poorly preserved elements of the wing are preserved, partly obscured by matrix (
Figure 2). They include the proximally fused first, second and third metacarpals, possibly carpal elements and what appears to be the distal end of the radius. These bones are generally similar to those of the living ostrich. Davies [
3] claimed that they were more robust than in the living ostrich, and this may be correct by comparison with the carpus of
Struthio camelus [
26], although the poor preservation of the bones makes accurate measurements difficult.
A series of ten articulated cervical vertebrae forms a semi-circle around the limb elements. The two vertebrae at the ends of the series are incomplete. Two additional cervicals are not articulated with the rest. The vertebrae are not very well preserved and show few details (
Figure 3), but their approximate position in the vertebral column can be estimated. By comparison with the cervical series of
Struthio camelus [
27], they probably correspond to the middle to posterior section of the neck (5th to 14th according to Davies [
3]). They generally resemble vertebrae of the living
Struthio (
Figure 3), although Davies [
3] noted some differences in proportions (see below).
The most striking character of the type specimen of
Struthio asiaticus is that the tarsometatarsus bears only two trochleae for articulation with the third and fourth toes. This didactyly was the reason why Milne-Edwards [
1] referred the fossil to the genus
Struthio, an attribution followed by subsequent authors. The only reason he provided for distinguishing the fossil species
Struthio asiaticus from the living
S. camelus was its supposedly smaller size.
Davies [
3] gave an accurate description of the Indian specimen and showed that it was comparable in size with a large male specimen of the living
Struthio camelus, thus questioning Milne-Edwards’s assertion about its smaller size. Comparison with the measurements of extant ostrich specimens published by Elzanowski and Louchart [
28] confirms that the distal width of the trochlea III (41 mm) of the tarsometatarsus is within the range of modern ostriches. The same applies to the proximal width of the first phalanx of the third toe (42 mm). There is therefore no reason to accept Milne-Edwards’s assertion that
Struthio asiaticus was smaller than living ostriches. Davies ([
3], p. 19) wrote: “but if we eliminate the conditions of occurrence and locality, the fragment possesses in itself no distinctive characters by which it could be separated from the existing African Ostrich”. Lydekker ([
9], p. 144) echoed Davies’s remark when he wrote about the tarsometatarsus and phalanx: “These bones are indistinguishable, both as regards form and size, from those of the existing ostrich”. Nevertheless, Davies [
3] noted that the cervical vertebrae, although their lengths were similar to those of the living ostrich, had somewhat taller and broader centra; he concluded that the Siwalik ostrich was as tall as the living species but had a more robust neck. Similarly, according to Davies, the poorly preserved metacarpals were stouter than in the African ostrich. Lydekker [
9] agreed with Davies’s observations, but considered that the small differences in the robustness of the cervical vertebrae could be the result of individual or varietal variation and concluded that the name
Struthio asiaticus should be regarded as provisional, because of the considerable similarities with
Struthio camelus.
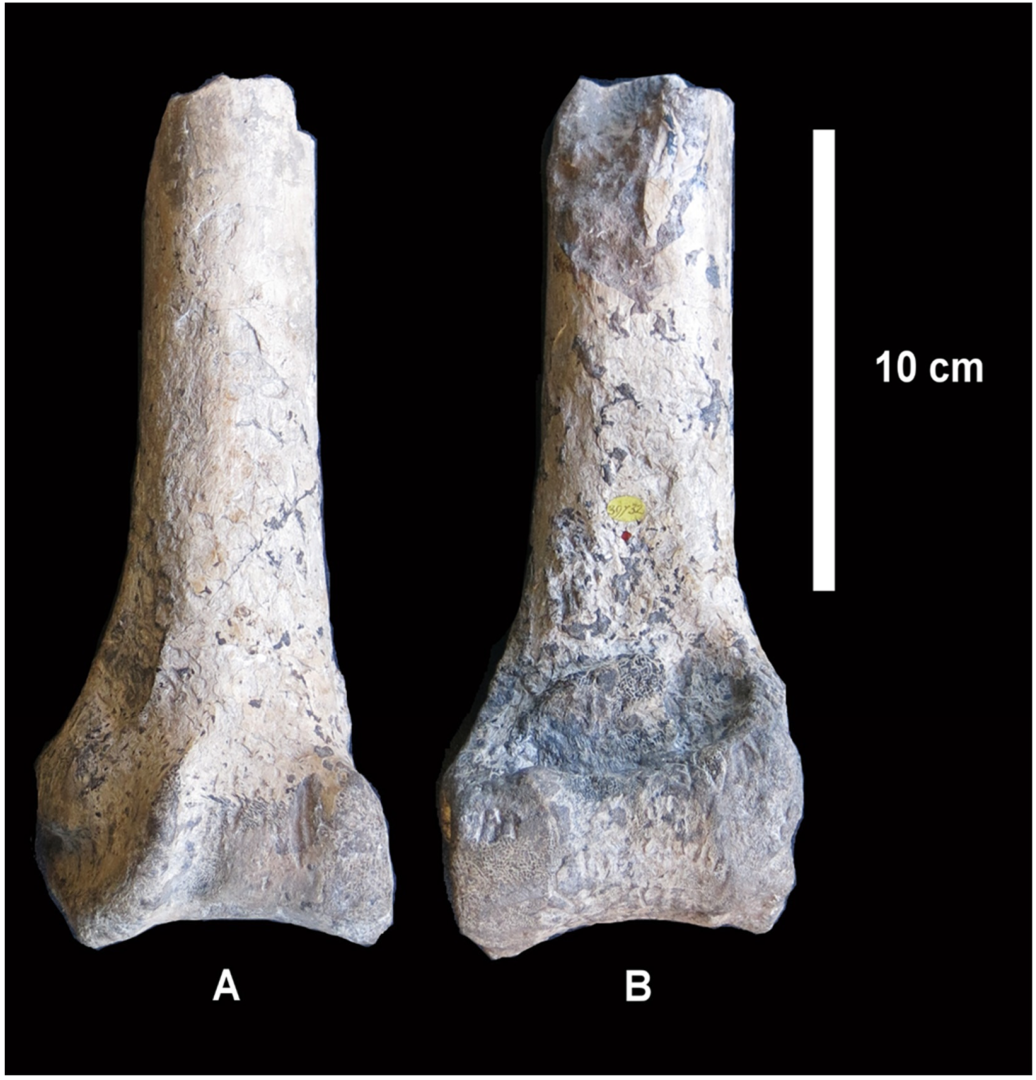
It is worth noting that, according to Davies [
3], the above-mentioned distal end of a right tibiotarsus (NHMUK PV OR 39732;
Figure 4) possibly belonged to the same individual as the group of bones constituting the type specimen of
Struthio asiaticus. Lydekker [
6,
9] accepted the identification as
S. asiaticus.The type of fossilisation is indeed very similar; however, the label accompanying NHMUK PV OR 39732, indicates that it was collected and presented by P.J. Cautley (in 1842 according to Lydekker [
6]), which suggests that its origin is different from that of the type specimen presented by J. Colvin in 1848. As noted by Davies [
3], this bone is very similar to the corresponding element in
Struthio camelus in its morphology and measurements. Lydekker ([
9], p. 144) found it “indistinguishable in all respects from the tibia of
S. camelus”.
Similarly, both Davies and Lydekker found the other genuine ostrich specimens from the Siwaliks (see above) very similar to the corresponding bones of the living ostrich.
Since the above-mentioned ostrich bones from the Siwaliks have no counterparts in the type specimen of Struthio asiaticus and therefore cannot really be compared with it, it seems difficult to confidently refer them to the latter taxon, all the more so that there is no solid evidence to show that they all come from the same geological horizon. At most they could be referred to as Struthio cf. asiaticus, to indicate that at least on the basis of size they are comparable with that species.
This raises the question of the distinguishing characters of
Struthio asiaticus. As shown by Davies [
3], Milne-Edwards’s assumption that it was smaller than
Struthio camelus was unfounded. Davies noted the greater robustness of the cervical vertebrae and metacarpals, and despite Lydekker’s doubts, it seems that this robustness is one of the main distinguishing characters that can be found to separate
Struthio asiaticus from
Struthio camelus. Marked robustness of the cervical vertebrae has also been reported in late Miocene specimens from Greece referred to
Struthio karatheodoris [
29]. However, it should be admitted that comparisons with the dimensions of more specimens of living ostriches would be welcome. Mourer-Chauviré and Geraads [
30], when comparing fossil ostrich remains from Morocco with
Struthio asiaticus, considered the latter as “massive” on the basis of the proportions of the phalanx as compared with those observed in recent ostriches, but the ratio (proximal width/minimum width of shaft) they provide for
S. asiaticus (1.67) is erroneous, the correct ratio being 1.75, well within the range of living
Struthio. Moreover, massiveness as such may not always be a reliable distinguishing specific character. Elzanowski and Louchart [
28], for instance, have noted the existence in living ostriches of a sexual dimorphism resulting in more robust limb bones in females.
Perhaps more significantly, Mikhailov and Zelenkov [
21] have argued that
Struthio asiaticus was “short-toed”, by comparison with ostriches with longer toes, including the living species. However, this interpretation is based only on the first phalanx of the third toe of the type specimen, which is very incomplete, lacking the distal end and a fraction of the shaft, the length of which cannot be accurately estimated. Nevertheless, comparison with the corresponding bone in
Struthio camelus and the short-toed late Miocene
S. brachydactylus, as illustrated by Mikhailov and Zelenkov ([
21], Figure 13), does suggest that
S. asiaticus was more similar to the latter. However, Mikhailov and Zelenkov’s ([
21], p. 26) contention that “the presence of the abbreviated third toe in
S. asiaticus is in general accordance with the late Miocene age of this species” should be taken with caution, since, as noted above, the age of
S. asiaticus remains highly uncertain. If indeed
Struthio asiaticus was short-toed, and if, as accepted by Mikhailov and Zelenkov, short-toed ostriches are mainly Miocene forms, a Miocene age for
S. asiaticus may be suggested, but one should beware of circular reasoning. In the absence of strong stratigraphical evidence, the geological age of the taxon must remain uncertain.
In view of the incompleteness of the material and of the paucity of distinguishing characters, the question may be asked whether
Struthio asiaticus should be considered as a valid species. If it is not, the type specimen should be referred to as
Struthio indet. However, as indicated in the diagnosis given above, the specimen appears to show a combination of characters that seems sufficient to distinguish it from other ostriches:
Struthio asiaticus was not larger than living ostriches, but its cervical vertebrae were less slender than in
S. camelus and it had shorter toes. The latter character, especially, seems to have some functional significance (see below) and it has been considered as significant by various authors, notably Mikhailov and Zelenkov [
21]. There are apparent similarities, in the shortness of the first phalanx of the third toe, between
Struthio asiaticus and
S. brachydactylus, from the late Miocene of Ukraine [
31,
32], although the Ukrainian form may have a less expanded distal end of the tarsometatarsus than the Indian one. However, as noted by Mikhailov and Zelenkov [
21], it does not seem advisable to merge these taxa because they are of widely different geographical origins and the age of
Struthio asiaticus is highly uncertain. Moreover, direct comparisons are impossible since the type material of
Struthio brachydactylus was lost during World War II [
32]. In any case, since the name
Struthio asiaticus considerably antedates
S. brachydactylus, which was coined in 1939, it would have clear priority if the two taxa turned out to be identical. Pending the discovery of more complete material, it seems preferable to retain
Struthio asiaticus as a distinct and valid species.
The apparent robustness of the neck and wing of Struthio asiaticus and the fact that it was probably short-toed suggest that it may have been less cursorial than the living ostriches, although it was certainly not as heavily built as the giant ostriches of the Pleistocene such as Pachystruthio and Struthio anderssoni. The lack of data about the locality and possible accompanying flora and fauna makes it difficult to reconstruct the palaeoenvironment in which Struthio asiaticus may have lived and little can be said about its palaeobiology.
6. Struthio asiaticus Outside the Indian Subcontinent?
Various avian fossils, comprising both bones and eggs or eggshell fragments, found outside the Indian subcontinent have been referred to
Struthio asiaticus. This raises various geographical, stratigraphical and palaeontological problems that are briefly discussed below. The idea of a very vast geographical and stratigraphical distribution of
Struthio asiaticus was put forward by Kurochkin and Lungu [
33], who considered that various ostrich remains from the Ukraine and southern Russia, that had been described under various specific names, should in fact be considered as belonging to
S. asiaticus. They thought that
Struthio asiaticus had existed from the Meotian (late Miocene) to the “Early Anthropogene” (Early Pleistocene), and had a very extensive geographical distribution, covering “the vast expanse of the modern steppe and in part the semi-desert zone of Eurasia between Eastern Europe and Central Asia”. They went even further, noting that “there are no grounds to consider that ostriches described from other parts of the skeleton from the Pliocene of Europe, Asia and Africa, which fall entirely within the range of
S. asiaticus both geographically and stratigraphically, are a different species”. This interpretation was based on limited osteological evidence, consisting of similarities in the morphology of the tibiotarsus and tarsometatarsus. Kurochkin and Lungu also described
Struthio asiaticus as “a relatively small ostrich” in the Early Pliocene, even though Davies [
3] had already shown that it was the size of a male
Struthio camelus (but their remark may have been based on a comparison with the very large ostriches of the Early Pleistocene, such as
Pachystruthio: see [
34,
35]).
The idea of a huge geographical distribution of
Struthio asiaticus was taken up by various subsequent authors. Mourer-Chauviré and Geraads [
30], for instance, referred to
Struthio asiaticus fairly abundant but fragmentary ostrich material from the Ahl al Oughlam locality in Morocco, close to the Plio-Pleistocene boundary. The main reasons for this attribution were similarities in the dimensions of phalanx 1 of pedal digit III—although the comparison was made difficult by the fact that only the distal portion of that phalanx is known from Ahl al Oughlam, while only the proximal part is preserved in the type specimen of
Struthio asiaticus. The only measurement that can be compared between the type of
Struthio asiaticus and the phalanx from Morocco is the minimum width of the shaft ([
30], Table 1), and it is significantly larger in the Ahl al Oughlam specimen (31.2 mm) than in the Indian specimen (25 mm). Nevertheless, Mourer-Chauviré and Geraads concluded that both the Moroccan and the Indian ostriches were “massive” and this seems to be one of the reasons for their attribution, in addition to the fact that they apparently accepted Kurochkin and Lungu’s ideas about the vast distribution of
Struthio asiaticus. Mikhailov and Zelenkov [
21] are certainly right in rejecting the identification of the ostrich from Ahl al Oughlam as
Struthio asiaticus, which is based on very flimsy morphological evidence and would imply a huge geographical distribution without any safe stratigraphical correlation between the relevant localities. As noted above, the proximal width/minimum shaft width ratio for the first phalanx of the first toe used by Mourer-Chauviré and Geraads [
30] is not correct. As remarked by Mikhailov and Zelenkov [
21], the width of the distal end of the tarsometatarsus (61 mm) of
Struthio asiaticus falls within the range of living
S. camelus, as do the distal width of trochlea III and the proximal width of the first phalanx of the third toe (see above), all of which does not suggest a specially robust form.
Manegold et al. [
36] similarly referred to
Struthio asiaticus ostrich bones from the Early Pliocene of Langebaanweg (South Africa) that were described as substantially larger than those of
S. camelus. The main reason for this attribution seems to have been similarities in the size and shape of the phalanges from Langebaanweg with those of the ostrich from Ahl al Oughlam described by Mourer-Chauviré and Geraads [
30]. Manegold et al. ([
36], p. 143) noted that “
Struthio asiaticus is about 20 % larger than large
S. camelus specimens”, which is certainly not indicated by the dimensions of the type specimen of the Asian species or those of the incomplete tibiotarsus NHMUK PV OR 39732 (see [
3]).
All this suggests that attributions of African specimens to Struthio asiaticus are based on rather flimsy evidence and erroneous assumptions about the size and robustness of the Asian species, and should be taken with extreme caution. On the basis of the available material, there are no valid reasons to admit that the Indian species Struthio asiaticus was really present in Africa.
Another question worth considering is that of the purported occurrence of
Struthio asiaticus in the Late Pleistocene of north-eastern Asia (China, Mongolia, Siberia). Ostrich eggs and eggshell fragments have been known from these areas for a long time [
37,
38,
39,
40,
41,
42], while skeletal remains are extremely rare. Lowe [
40] coined the name
Struthio anderssoni for the large eggs from the loess of northern China, and a few bones from the Late Pleistocene of the Upper Cave at Zhoukoudian were later referred to it [
43,
44,
45]. Kurochkin et al. ([
46], p. 1091), however, noted that “Asiatic ostrich (
Struthio asiaticus Brodkorb, 1963) inhabited northern China, Mongolia, and the southern part of Siberia in the later half of the Cenozoic: in the Miocene, Pliocene, and the Pleistocene, and perhaps up to the Holocene” and accordingly referred the abundant eggshell material from these regions, often found in anthropic sites, to that taxon. This identification has been followed mainly by Russian archaeologists (e.g., [
47]), other researchers preferring to refer these eggshells to
Struthio sp. [
48]. However, as mentioned above, no eggshells are associated with the type of
Struthio asiaticus and it seems unlikely that this presumably Neogene ostrich from India was present during the Late Pleistocene in northeastern Asia. It should also be noted that mass estimates based on the eggs from the Chinese loess and on the femora from Zhoukoudian indicate a very large ostrich, about 1.5 times the mass of
Struthio camelus [
49], which does not fit
Struthio asiaticus, which, although possibly somewhat more robustly built, was not taller than
S. camelus. It therefore seems preferrable to refer the eggs (and the few bones) from the Late Pleistocene of northeastern Asia to
Struthio anderssoni Lowe, 1931 (the holotype of which is a complete egg from China, NHMUK A1308) rather than to
S. asiaticus. This was the conclusion reached by Mikhailov and Zelenkov [
21], although their assumption, based on eggshell evidence, that
Struthio anderssoni and
S. camelus may have belonged to the same “biological ostrich species” seems highly questionable in view of the notable differences in size and morphology indicated by the skeletal remains from Zhoukoudian attributed to
S. anderssoni [
43,
44,
45].
To sum up, in view of the morphological characters of
Struthio asiaticus, as represented by the type specimen, and of the uncertainties about its stratigraphical origin, it seems unwarranted to refer to this taxon fossil material from outside the Indian subcontinent.
Struthio asiaticus has too often been used as “somewhat of a wastebasket taxon”, to use Widrig and Field’s expression [
25].



{kind=link}
{kind=link}
{kind=link}
{kind=link}



