
Characterization and Gene Expression of Vitellogenesis-Related Transcripts in the Hepatopancreas and Ovary of the Red Swamp Crayfish, Procambarus clarkii (Girard, 1852), during Reproductive Cycle

 ,
, 


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Note
2.2. Transcripts Identification
2.3. Comparative Studies
2.4. Transcript Expression by qRT-PCR
3. Results
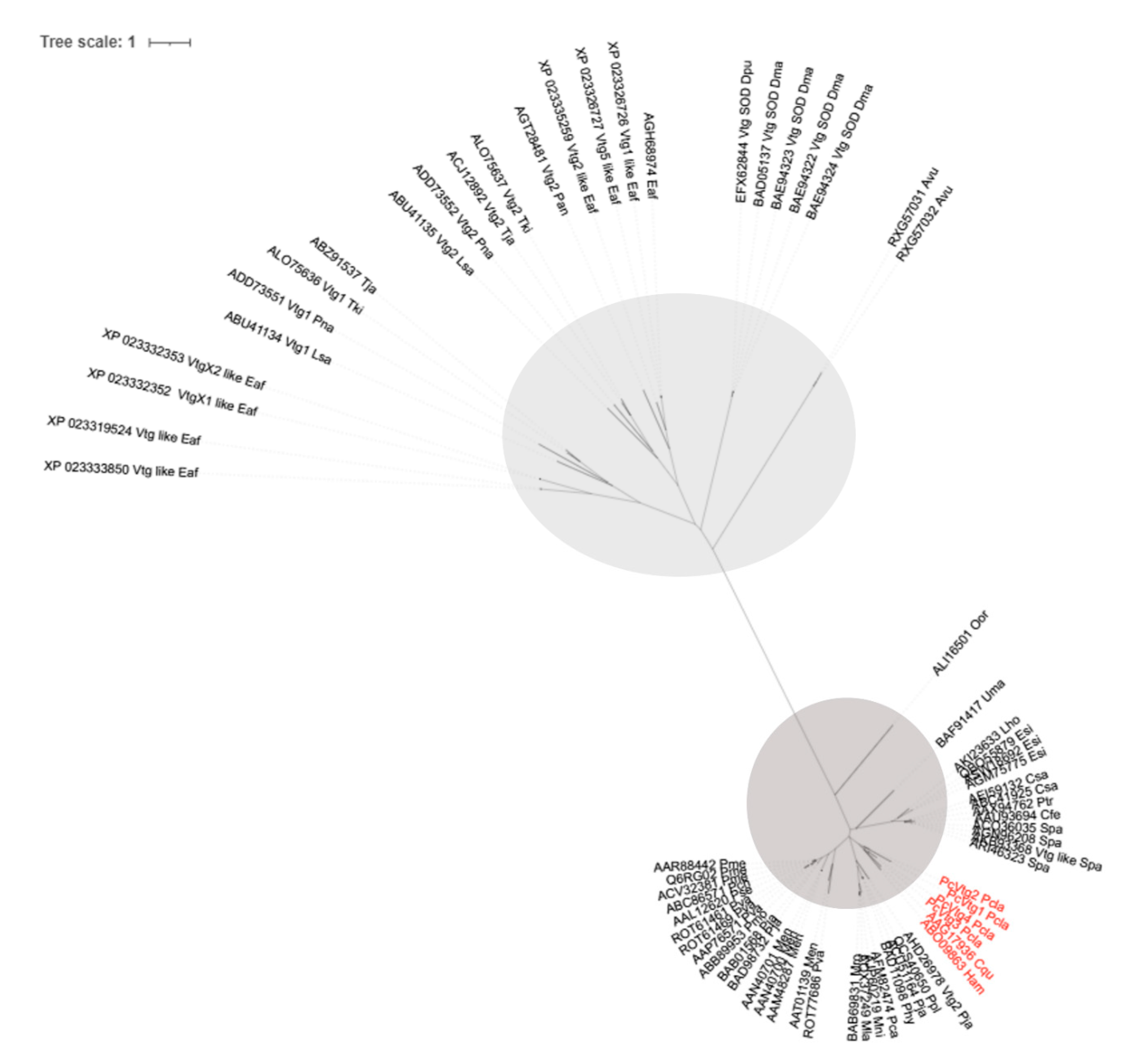
3.1. Characterization of VTG-like Transcripts
3.2. Expression of VTG-Like Transcripts during Ovarian Development
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Busson-Mabillot, S. Données récentes sur la vitellogenèse. Ann. Biol. 1969, 8, 199–228. [Google Scholar]
- Dhainaut, A.; De Leersnyder, M. Etude cytochimique et ultrastructurale de l’évolution ovocytaire du crabe Eriocheir sinensis. I. Ovogenèse naturelle. Arch. Biol. 1976, 87, 261–282. [Google Scholar]
- Meusy, J.J. Vitellogenin, the extraovarian precursor of the protein yolk in Crustacea: A review. Reprod. Nutr. Dévelop. 1980, 20, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, M.S. The hemolymph proteins of the blue crab, Callinectes sapidus. II. A lipoprotein serologically identical to oocyte lipovitellin. Develop. Biol. 1969, 20, 1–17. [Google Scholar] [CrossRef]
- Lee, C.Y.; Watson, D. In Vitro study of vitellogenesis in the blue crab (Callinectes sapidus): Site and control of vitellin synthesis. J. Exp. Zool. 1995, 271, 364–372. [Google Scholar] [CrossRef]
- Browdy, C.L.; Fainzilber, M.; Tom, M.; Loya, Y.; Lubzens, E. Vitellin synthesis in relation to oogenesis in vitro incubated ovaries of Penaeus semisulcatus. J. Exp. Zool. 1990, 255, 205–215. [Google Scholar] [CrossRef]
- Mak, A.S.; Choi, C.L.; Tiu, S.H.; Hui, J.H.; He, J.G.; Tobe, S.S.; Chan, S.M. Vitellogenesis in the red crab Charybdis feriatus: Hepatopancreas-specific expression and farnesoic acid stimulation of vitellogenin gene expression. Mol. Reprod. Dev. 2005, 70, 288–300. [Google Scholar] [CrossRef]
- Yang, W.J.; Ohira, T.; Tsutsui, N.; Subramoniam, T.; Huong, D.T.T.; Aida, K.; Wilder, M.N. Determination of amino acid sequence and site of mRNA expression of four vitellins in the giant freshwater prawn, Macrobrachium rosenbergii. J. Exp. Zool. 2000, 287, 413–422. [Google Scholar] [CrossRef]
- Girish, B.P.; Swetha, C.; Reddy, P.S. Hepatopancreas but not ovary is the site of vitellogenin synthesis in female fresh water crab, Oziothelphusa senex. Biochem. Biophy. Res. Commun. 2014, 447, 323–327. [Google Scholar] [CrossRef]
- Tsutsui, N.; Saido-Sakanaka, H.; Yang, W.J.; Jayasankar, V.; Jasmani, S.; Okuno, A.; Ohira, T.; Okumura, T.; Aida, K.; Wilder, M.N. Molecular characterization of a cDNA encoding vitellogenin in the coonstriped shrimp, Pandalus hypsinotus and site of vitellogenin mRNA expression. J. Exp. Zool. 2004, 301, 802–814. [Google Scholar] [CrossRef]
- Ding, X.; Nagaraju, G.P.C.; Novotney, D.; Lovett, D.L.; Borst, D.W. Yolk protein expression in the green crab, Carcinus maenas. Aquaculture 2010, 298, 325–331. [Google Scholar] [CrossRef]
- Li, K.; Chen, L.; Zhou, Z.; Li, E.; Zhao, X.; Guo, H. The site of vitellogenin synthesis in Chinese mitten-handed crab Eriocheir sinensis. Comp. Biochem. Physiol. B 2006, 143, 453–458. [Google Scholar] [CrossRef]
- Bai, H.; Qiao, H.; Li, F.; Fu, H.; Sun, S.; Zhang, W.; Jin, S.; Gong, Y.; Jiang, S.; Xiong, Y. Molecular characterization and developmental expression of vitellogenin in the oriental river prawn Macrobrachium nipponense and the effects of RNA interference and eyestalk ablation on ovarian maturation. Gene 2015, 562, 22–31. [Google Scholar] [CrossRef]
- Okumura, T.; Yamano, K.; Sakiyama, K. Vitellogenin gene expression and hemolymph vitellogenin during vitellogenesis, final maturation, and oviposition in female kuruma prawn, Marsupenaeus japonicus. Comp. Biochem. Physiol. A 2007, 147, 1028–1037. [Google Scholar] [CrossRef]
- Hiransuchalert, R.; Thamniemdee, N.; Khamnamtong, B.; Yamano, K.; Klinbunga, S. Expression profiles and localization of vitellogenin mRNA and protein during ovarian development of the giant tiger shrimp Penaeus monodon. Aquaculture 2013, 412–413, 193–201. [Google Scholar] [CrossRef]
- Jia, X.; Chen, Y.; Zou, Z.; Lin, P.; Wang, Y.; Zhang, Z. Characterization and expression profile of Vitellogenin gene from Scylla paramamosain. Gene 2013, 520, 119–130. [Google Scholar] [CrossRef]
- Salmela, H.; Stark, T.; Stucki, D.; Fuchs, S.; Freitak, D.; Dey, A.; Kent, C.F.; Zayed, A.; Dhaygude, K.; Hokkanen, H.; et al. Ancient Duplications Have Led to Functional Divergence of Vitellogenin-Like Genes Potentially Involved in Inflammation and Oxidative Stress in Honey Bees. Genome Biol. Evol. 2016, 8, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Wurm, Y.; Wang, J.; Riba-Grognuz, O.; Corona, M.; Nygaard, S.; Hunt, B.G.; Ingram, K.K.; Falquet, L.; Nipitwattanaphon, M.; Gotzek, D.; et al. The genome of the fire ant Solenopsis invicta. Proc. Natl. Acad. Sci. USA 2011, 108, 5679–5684. [Google Scholar] [CrossRef] [Green Version]
- Abdu, U.; Davis, C.; Khalaila, I.; Sagi, A. The vitellogenin cDNA of Cherax quadricarinatus encodes a lipoprotein with calcium binding ability, and its expression is induced following the removal of the androgenic gland in a sexually plastic system. Gen. Comp. Endocrinol. 2002, 127, 263–272. [Google Scholar] [CrossRef]
- Avarre, J.-C.; Michelis, R.; Tietz, A.; Lubzens, E. Relationship between vitellogenin and vitellin in a marine shrimp (Penaeus semisulcatus) and molecular characterization of vitellogenin complementary DNAs. Biol. Reprod. 2003, 69, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Okuno, A.; Yang, W.J.; Jayasankar, V.; Saido-Sakanaka, H.; Huong, D.T.T.; Jasmani, S.; Atmomarsono, M.; Subramoniam, T.; Tsutsui, N.; Ohira, T.; et al. Deduced Primary Structure of Vitellogenin in the Giant Freshwater Prawn, Macrobrachium rosenbergii, and Yolk Processing During Ovarian Maturation. J. Exp. Zool. 2002, 292, 417–429. [Google Scholar] [CrossRef]
- Jeon, J.M.; Lee, S.O.; Kim, K.S.; Baek, H.J.; Kim, S.; Kim, I.K.; Mykles, D.L.; Kim, H.W. Characterization of two vitellogenin cDNAs from a Pandalus shrimp (Pandalopsis japonica): Expression in hepatopancreas is down-regulated by endosulfan exposure. Comp. Biochem. Physiol. B 2010, 157, 102–112. [Google Scholar] [CrossRef]
- Phiriyangkul, P.; Utarabhand, P. Molecular Characterization of a cDNA Encoding vitellogenin in the banana shrimp, Penaeus (Litopenaeus) merguiensis and sites of vitellogenin mRNA Expression. Molec. Reprod. Dev. 2006, 73, 410–423. [Google Scholar] [CrossRef]
- Raviv, S.; Parnes, S.; Segall, C.; Davis, C.; Sagi, A. Complete sequence of Litopenaeus vannamei (Crustacea: Decapoda) vitellogenin cDNA and its expression in endocrinologically induced sub-adult females. Gen. Comp. Endocrinol. 2006, 145, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Tiu, S.H.K.; Hui, J.H.L.; Mak, A.S.C.; He, J.G.; Chan, S.M. Equal contribution of hepatopancreas and ovary to the production of vitellogenin (PmVg1) transcripts in the tiger shrimp, Penaeus monodon. Aquaculture 2006, 254, 666–674. [Google Scholar] [CrossRef]
- Tsang, W.S.; Quackenbush, S.L.; Chow, B.K.C.; Tiu, S.H.K.; He, J.G.; Chan, S.M. Organization of the shrimp vitellogenin gene: Evidence of multiple genes and tissue specific expression by the ovary and hepatopancreas. Gene 2003, 303, 99–109. [Google Scholar] [CrossRef]
- Tsutsui, N.; Kawazoe, I.; Ohira, T.; Jasmani, S.; Yang, W.J.; Wilder, M.N.; Aida, K. Molecular characterization of a cDNA encoding vitellogenin and its expression in the hepatopancreas and ovary during vitellogenesis in the kuruma prawn, Penaeus japonicus. Zool. Sci. 2000, 17, 651–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.; Sun, L.; Liu, F.; Dong, B. Molecular characterization and mRNA transcript profile of vitellogenin in Chinese shrimp, Fenneropenaeus chinensis. Molec. Biol. Rep. 2009, 36, 389–397. [Google Scholar] [CrossRef]
- Yang, F.; Xu, H.T.; Dai, Z.M.; Yang, W.J. Molecular characterization and expression analysis of vitellogenin in the marine crab Portunus trituberculatus. Comp. Biochem. Physiol. B 2005, 142, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cheng, L.; Liu, J.; Li, Z.; Xie, S.; Silva, S.S.D. Freshwater aquaculture in PR China: Trends and prospects. Rev. Aquac. 2015, 7, 283–302. [Google Scholar] [CrossRef]
- Putra, M.D.; Bláha, M.; Wardiatno, Y.; Krisanti, M.; Yonvitner; Jerikho, R.; Kamal, M.M.; Mojžišová, M.; Bystřický, P.K.; Kouba, A.; et al. Procambarus clarkii (Girard, 1852) and crayfish plague as new threats for biodiversity in Indonesia. Aquat. Conserv. 2018, 28, 1434–1440. [Google Scholar] [CrossRef]
- Haubrock, P.J.; Oficialdegui, F.J.; Zeng, Y.; Patoka, J.; Yeo, D.C.J.; Kouba, A. The red claw crayfish: A prominent aquaculture species with invasive potential in tropical and subtropical biodiversity hotspots. Rev. Aquac. 2021, 13, 1488–1530. [Google Scholar] [CrossRef]
- Gherardi, F.; Barbaresi, S.; Salvi, G. Spatial and temporal patterns in the movement of Procambarus clarkii, an invasive crayfish. Aquat. Sci. 2000, 62, 179–193. [Google Scholar] [CrossRef]
- Scalici, M.; Gherardi, F. Structure and dynamics of an invasive population of the red swamp crayfish (Procambarus clarkii) in a Mediterranean wetland. Hydrobiologia 2007, 583, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Peruzza, L.; Piazza, F.; Manfrin, C.; Bonzi, L.C.; Battistella, S.; Giulianini, P.G. Reproductive plasticity of a Procambarus clarkii population living 10 °C below its thermal optimum. Aquat. Inv. 2015, 10, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Silveyra, G.R.; Silveyra, P.; Vatnick, I.; Medesani, D.A.; Rodríguez, E.M. Effects of atrazine on vitellogenesis, steroid levels and lipid peroxidation, in female red swamp crayfish Procambarus clarkii. Aquat. Toxicol. 2018, 197, 136–142. [Google Scholar] [CrossRef]
- Guan, Z.B.; Yin, J.; Chen, K.; Shui, Y.; Cai, Y.J.; Liao, X.R. The hepatopancreas and ovary are the sites of vitellogenin synthesis in female red swamp crayfish Procambarus clarkii (Girard, 1852) (Decapoda: Astacoidea: Cambaridae). J. Crust. Biol. 2016, 36, 637–641. [Google Scholar] [CrossRef] [Green Version]
- Kang, P.F.; Mao, B.; Fan, C.; Wang, Y.F. Transcriptomic information from the ovaries of red swamp crayfish (Procambarus clarkii) provides new insights into development of ovaries and embryos. Aquaculture 2019, 505, 333–343. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhao, W.; Tang, Z.; Huang, L.; Zhu, X.; Liang, X.; Yan, A.; Lu, Z.; Yu, Y.; Tang, D.; et al. Comparative transcriptomic analysis of the different developmental stages of ovary in red swamp crayfish Procambarus clarkii. BMC Genom. 2021, 22, 199. [Google Scholar] [CrossRef]
- Duvic, B.; Söderhäll, K. Purification and partial characterization of a beta-1,3-glucanbinding-protein membrane receptor from blood cells of the crayfish Pacifastacus leniusculus. Eur. J. Biochem. 1992, 207, 223–228. [Google Scholar] [CrossRef]
- Stieb, S.; Roth, Z.; Dal Magro, C.; Fischer, S.; Butz, E.; Sagi, A.; Khalaila, I.; Lieb, B.; Schenk, S.; Hoeger, U. One precursor, three apolipoproteins: The relationship between two crustacean lipoproteins, the large discoidal lipoprotein and the high density lipoprotein/β-glucan binding protein. Biochim. Biophys. Acta 2014, 1841, 1700–1708. [Google Scholar] [CrossRef]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, 260–265. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcorlo, P.; Geiger, W.; Otero, M. Reproductive biology and life cycle of the invasive crayfish Procambarus clarkii (Crustacea: Decapoda) in diverse aquatic habitats of South-Western Spain: Implications for population. Fundam. Appl. Limnol. 2008, 173, 197–212. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.; Moorman, A.F. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [Green Version]
- Kibbe, W.A. OligoCalc: An online oligonucleotide properties calculator. Nucleic Acids Res. 2007, 35, 43–46. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pairwise correlations. Biotech. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data set. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchler-Bauer, A.; Bryant, S.H. CD-Search: Protein domain annotations on the fly. Nucl. Acids Res. 2004, 32, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Avarre, J.C.; Lubzens, E.; Babin, P.J. Apolipocrustacein, formerly vitellogenin, is the major egg yolk precursor protein in decapod crustaceans and is homologous to insect apolipophorin II/I and vertebrate apolipoprotein B. BMC Evol. Biol. 2007, 7, 3. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer ID | Forward 5’-3’ | Reverse 5’-3’ |
|---|---|---|
| PcVTG1 | TCACCAGTCAACAGAGCAGC | TTCTCAGCACACCGAACTGC |
| PcVTG2 | GAGGGTGGAAAGTCAGCTCC | ACAGTTCATCGCTCCTTCGG |
| PcVTG3 | GTCGGACTGCAGATGAAGGG | AACAAAGCCTTCGGTTTGCG |
| PcVTG4 | TCTGTTGAGAAAGCCGAGCC | TCTAGGCGTACTAGACCCAGC |
| PcBGBP1 | CACACAAGACGAAGTGCTGC | TAAACGGTGCTAAGGGCTGG |
| PcBGBP2 | CCCCTAGCATTAGCAACCCC | ACAACTCGGCGTCTTTCTCG |
| PcEF1α | AGATCTGAAACGTGGTTTTGTT | TCAATCTTTTCCAGAAGTTCGT |
| Pcβ-Actin | AGGGCGTGATGGTTGGTAT | CCGTGCTCAATGGGATATTT |
| PcGAPDH | CTCCATCTTTGACGCTAAGGC | GCACTATCCACCTTCTGCATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manfrin, C.; Tom, M.; Avian, M.; Battistella, S.; Pallavicini, A.; Giulianini, P.G. Characterization and Gene Expression of Vitellogenesis-Related Transcripts in the Hepatopancreas and Ovary of the Red Swamp Crayfish, Procambarus clarkii (Girard, 1852), during Reproductive Cycle. Diversity 2021, 13, 445. https://doi.org/10.3390/d13090445
Manfrin C, Tom M, Avian M, Battistella S, Pallavicini A, Giulianini PG. Characterization and Gene Expression of Vitellogenesis-Related Transcripts in the Hepatopancreas and Ovary of the Red Swamp Crayfish, Procambarus clarkii (Girard, 1852), during Reproductive Cycle. Diversity. 2021; 13(9):445. https://doi.org/10.3390/d13090445
Chicago/Turabian StyleManfrin, Chiara, Moshe Tom, Massimo Avian, Silvia Battistella, Alberto Pallavicini, and Piero Giulio Giulianini. 2021. "Characterization and Gene Expression of Vitellogenesis-Related Transcripts in the Hepatopancreas and Ovary of the Red Swamp Crayfish, Procambarus clarkii (Girard, 1852), during Reproductive Cycle" Diversity 13, no. 9: 445. https://doi.org/10.3390/d13090445
APA StyleManfrin, C., Tom, M., Avian, M., Battistella, S., Pallavicini, A., & Giulianini, P. G. (2021). Characterization and Gene Expression of Vitellogenesis-Related Transcripts in the Hepatopancreas and Ovary of the Red Swamp Crayfish, Procambarus clarkii (Girard, 1852), during Reproductive Cycle. Diversity, 13(9), 445. https://doi.org/10.3390/d13090445