Abstract
We previously showed that novel filterable bacteria remain in “sterile” (<0.2 μm filtered) terrestrial environmental samples from Japan, China, and Arctic Norway. Here, we characterized the novel filterable strain IZ6T, a representative strain of a widely distributed lineage. Phylogenetic analysis showed that this strain was affiliated with the Rhizobiales (now proposed as Hyphomicrobiales) of Alphaproteobacteria, but distinct from any other type strains. Strain IZ6T shared the following chemotaxonomic features with the closest (but distantly) related type strain, Flaviflagellibacter deserti SYSU D60017T: ubiquinone-10 as the major quinone; phosphatidylethanolamine, phosphatidylcholine, and phosphatidylglycerol as major polar lipids; and slightly high G+C content of 62.2 mol%. However, the cellular fatty acid composition differed between them, and the unsaturated fatty acid (C18:1ω7c/C18:1ω6c) was predominantly found in our strain. Moreover, unlike methyrotrophs and nitrogen-fixers of the neighboring genera of Hyphomicrobiales (Rhizobiales), strain IZ6T cannot utilize a one-carbon compound (e.g., methanol) and fix atmospheric nitrogen gas. These findings were consistent with the genome-inferred physiological potential. Based on the phylogenetic, physiological, and chemotaxonomic traits, we propose that strain IZ6T represents a novel genus and species with the name Terrihabitans soli gen. nov., sp. nov. (=NBRC 106741T = NCIMB 15058T). The findings will provide deeper insight into the eco-physiology of filterable microorganisms.
Keywords:
filter sterilization; filterable bacteria; soil; phylogeny; taxonomy; Hyphomicrobiales; Rhizobiales 1. Introduction
Removal of microorganisms using a micropore filter (e.g., 0.2 μm pore size) is a common procedure in research laboratories as well as medical and industrial processes [1]. However, some bacteria in nature are known to pass through a 0.2 μm-pore-size filter and to persist in “sterile” (<0.2 μm filtered) environmental samples (reviewed in [2,3,4,5]). Culturing efforts targeting filterable microorganisms have uncovered novel filterable bacteria, including obligate and facultative ultramicrobacteria as well as slender bacteria [3,5]. Such filterable microorganisms have been explored and mined mainly in water samples collected in aquatic systems [5]. On the other hand, culture-independent analyses have also revealed that novel and/or rarely cultivated microorganisms are present in small cell size fractions (<0.8 μm) of terrestrial soil systems [6]. Nonetheless, there are only a few successes of isolating pure cultures from filtered fractions in soils (e.g., Verrucomicrobia isolates [7]).
During the course of mining ultra-small microorganisms, we isolated and cultivated a novel 0.2 μm-filterable Rhizobiales bacterium, strain IZ6T, from a filtered suspension of forest soil [8]. We also obtained its phylogenetically closely-related isolates from other filtered samples (i.e., bark soil, sand, and travertine) collected in geographically remote areas such as Japan, China, and Arctic Norway [8]. These isolates formed a novel genus-level cluster in the 16S rRNA gene sequence-based phylogenetic tree [9]. Previous investigations also showed that strain IZ6T was not a typical ultramicrobacterium (cell volume, <0.1 μm3 [3,5]) under the optimal culture condition, but it does exhibit cell plasticity resulting in some filterable cells under a colder temperature condition [9]. This study characterized a cultivated representative strain IZ6T of a widely distributed terrestrial lineage by using polyphasic approaches including morphological, physiological, biochemical, and chemotaxonomic analyses in addition to phylogenomic analysis. Based on the characterizations, we propose the novel genus and species, Terrihabitans soli gen. nov., sp. nov., for this strain. Note that the order Hyphomicrobiales of the class Alphaproteobacteria was recently proposed as a replacement for Rhizobiales [10] and is herein tentatively described as Hyphomicrobiales (Rhizobiales) in reference to previous research [11].
2. Materials and Methods
2.1. Phylogenetic and Phylogenomic Analysis
The full-length 16S rRNA gene sequence of strain IZ6T was retrieved from the genome sequence (AP023361 [9]). The sequence was BLASTn-searched against the NCBI nt/nr database (accessed, June 2021) for the identification of the closest type strains, taxonomically undescribed strains, and uncultivated clones/phylotypes. To evaluate genome-scale similarities, the whole-genome-based average nucleotide identity (gANI) of strain IZ6T against the closest type strains identified using BLASTn was calculated using the ANI calculator [12]. The two-way averaged amino acid identity (AAI) and alignment fractions (AF) values were also calculated by using the AAI and AF calculators [13,14]. For the whole genome-based taxonomic analysis, the genome sequence data were analyzed by the Type (Strain) Genome Server (TYGS [15]) (accessed, June 2021). For the phylogenomic inference, all pairwise comparisons among the set of genomes were conducted using the Genome BLAST Distance Phylogeny approach (GBDP), and accurate intergenomic distances were inferred under the algorithm “trimming” and distance formula d5 [16]. A total of 100 distance replicates were calculated for each genome. Digital DNA-DNA hybridization (DDH) values and confidence intervals were calculated using the recommended settings of the GGDC 2.1 [16]. The resulting intergenomic distances were used to infer a balanced minimum-evolution tree with branch support via FASTME 2.1.6.1, including SPR postprocessing [17]. Branch support was inferred from 100 pseudo-bootstrap replicates. The trees were rooted at the midpoint [18] and visualized with PhyD3 [19].
2.2. Metagenomic Database Search for Potential Habitat Prediction
The potential distribution and habitability of strain IZ6T and its close relatives were predicted by the IMNGS platform [20], which is a database search against metagenome-derived 16S rRNA gene amplicon datasets, as well as the ProkAtlas search, which contains multiple 16S rRNA gene sequences labeled by one environmental category [21]. Both tools were performed with the sequence similarity threshold of 99%, using the sequence of IZ6T as the query.
2.3. Morphological, Physiological, and Biochemical Characterization
Strain IZ6T was originally isolated from a 0.2 μm filtrate of a suspension of forest soil collected in March 2009 from western Japan, as previously described [8]. IZ6T is routinely cultivated and maintained in R2A agar “DAIGO” (Nihon Pharmaceutical, Tokyo, Japan) at 25 °C. For colony observation, strain IZ6T was grown on R2A agar at 25 °C for 7 days. The cell morphology was observed in a previous study [9]. The cell motility was examined by microscopy (Olympus BX-50F4; Olympus Optical, Tokyo, Japan) after cultivation at 25 °C in R2A liquid medium (R2A broth “DAIGO”; Nihon Pharmaceutical). Gram staining was carried out using a Favor G Nissui kit (Nissui Pharmaceutical, Tokyo, Japan). Growth at a range of temperatures (4 °C, 10 °C, 15 °C, 20 °C, 25 °C, 30 °C, and 37 °C) was assessed on R2A agar. Growth at various pH levels (pH 4–12) and salt concentrations (0–2% [w/v] NaCl) was also determined on R2A agar; the pH was adjusted by adding 1M or 6M HCl or NaOH prior to sterilization. Anaerobic growth was tested using a chamber with an Anaero Pack system (Mitsubishi Gas Chemical, Tokyo, Japan). Catalase and oxidase activities were assessed following the previously reported method [22]. Physiological and biochemical characteristics and enzyme activities were tested at 25 °C using the API 20NE and APIZYM test strips (bioMérieux, Tokyo, Japan) according to the manufacturer’s instructions. Growth on methanol was tested with a minimal medium K containing methanol (0.5%, v/v) as reported in a previous study [23]. Nitrogen fixation potential was assessed based on the growth at 25 °C for 3–4 weeks on a nitrogen-free MS2N medium following the method of Bao et al. [24]. To account for the effect of nitrogen carryover derived from the pre-cultivation medium (R2A medium) before inoculation, cells grown in MS2N were inoculated into a fresh MS2N medium, and further growth was confirmed.
2.4. Chemotaxonomic Analyses
The chemotaxonomic properties of strain IZ6T were characterized by data on the cellular fatty acid composition, major respiratory quinone, polar lipids, and genomic GC content. For the fatty acid analysis, cells grown on R2A agar at 25 °C for 5 days were harvested and used. Fatty acids methyl esters were prepared and analyzed using the protocol of the Sherlock Microbial Identification system version 6.0 (Microbial ID; MIDI Inc., Newark, DE, USA). The fatty acid profile was compared and determined with the TSBA library (TSBA6 6.20). For the quinone determination, cells were grown on R2A agar at 25 °C for 5 days and were harvested and freeze-dried. The total lipids of the cells were extracted by a modification of the previously reported method [25], and the quinones in the crude extract were purified using a Sep-Pak plus silica column (Waters, Milford, MA, USA). The molecular type and concentration of each quinone extracted were analyzed by an ultra-performance liquid chromatography (UPLC) system (Acquity UPLC system; Waters) equipped with a photodiode array detector (UPPDA-E; Waters), an Eclipse plus C18 column (3.0 × 150 mm; pore size, 3.5 μm; Agilent Technologies, Tokyo, Japan), and Empower 2 software (Waters). The quinone species was finally determined based on the linear relationship between the logarithm of the UPLC retention time and the number of isoprene units according to the equivalent number of isoprene units of quinone components as reported by Tamaoka et al. [26]. For the characterization of polar lipids, extracted lipids were also separated by two-dimensional HPTLC silica gel 60 (Merck, Tokyo, Japan) following the previously reported experimental method and condition [27]. The genomic G + C content was calculated from the complete genome sequence of this strain (AP023361 [9]).
3. Results and Discussion
3.1. Phylogenetic Affiliation and Phylogenomic Placement of Strain IZ6T
The full-length 16S rRNA gene sequence of strain IZ6T was moderately related to Hyphomicrobiales (Rhizobiales) members of Alphaproteobacteria, but showed no more than 97% sequence identity to those of known type strains as of June 2021. The closest type strain was Flaviflagellibacter deserti SYSU D60017T, which was isolated from desert soil [28], and shared 96.9% sequence identity with strain IZ6T. Type strain SYSU D60017T is assigned to the family Rhizobiaceae in the NCBI taxonomy and to Ca. Methylopilaceae in the Genome Taxonomy Database (GTDB), although the family-level classification is not assigned in the List of Prokaryotic names with Standing in Nomenclature (LPSN). Note that this taxonomic issue regarding the placement of the family of strains IZ6T and SYSU D60017T will be discussed later. As of June 2021, the taxonomically undescribed isolates most closely related (16S rRNA gene sequence identities, 98.7–100%) to strain IZ6T were five filterable strains (DDBJ/ENA/NCBI accession nos. AB539978, AB539979, AB539981, AB539982, and AB540022) isolated from 0.2 μm-filtered fractions of terrestrial samples collected in Japan, China, and Arctic Norway, as reported in our previous study [8]. These data indicate that strain IZ6T and its close relatives are taxonomically novel candidates belonging to the Hyphomicrobiales (Rhizobiales).
The whole-genome-based average nucleotide identity (gANI) and the alignment fractions (AF) value of strain IZ6T against the closest type strain F. deserti were 75.4% and 0.50–0.59, respectively. These values were somewhat higher than the threshold for genus-level differentiation [29], while the value (69.9%) of the averaged amino acid identity (AAI) falls within the genus-level threshold (60–80% [30]). Moreover, a phylogenomic tree based on the Genome BLAST Distance Phylogeny (GBDP) approach indicated that although this strain was relatively close to F. deserti, this represents a novel genus-level lineage (Figure 1). This result was consistent with the phylogenetic novelty inferred from the previous 16S rRNA gene-based phylogenetic tree [9].
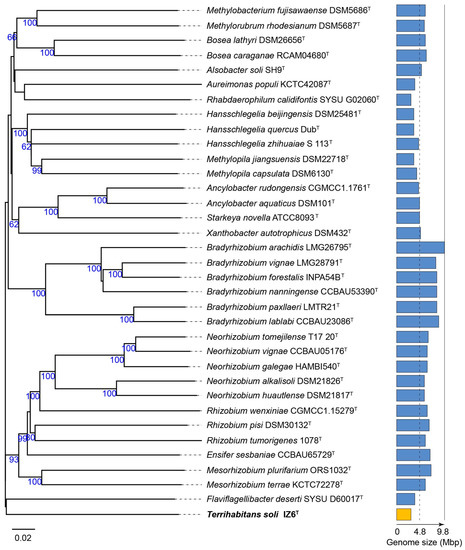
Figure 1.
Phylogenomic tree of Terrihabitans soli IZ6T and its relatives as inferred by FastME 2.1.6.1 [17] from the Genome BLAST Distance Phylogeny (GBDP) distances calculated from genome sequences. The branch lengths are scaled in terms of GBDP distance formula d5. The numbers above branches are GBDP pseudo-bootstrap support values of >60% from 100 replications, with average branch support of 71.8%. The tree was rooted at the midpoint [18]. The distribution of the genome size is shown as a bar graph on the right side; the genomes ranged from 3.1 Mbp to 9.8 Mbp. The data for strain IZ6T are colored orange, and all other data are colored blue. The scale bar indicates the branch length value.
It should be noted that the genome size (3.11 Mb) of IZ6T is smaller than that (3.72 Mb) of F. deserti and was one of the smallest sizes among the free-living members in Hyphomicrobiales (Rhizobiales) (Figure 2). The smaller genomes all belong to host-associated (e.g., parasitic) members of the genera Bartonella, Ca. Tokpelaia, Ca. Liberibacter, Ca. Hodgkinia. In general, planktonic and filterable ultramicrobacteria in aquatic systems tend to have small genomes, known as so-called “genome streamlining” (1–2 Mb [5,31,32]). Although strain IZ6T does not exhibit the typical cell size of ultramicrobacteria, as described below, its genome size is at the boundary between those of free-living and host-associated bacteria. This suggests that genome reduction and/or streamlining of IZ6T is currently underway, but this will require further validation, including genome analysis of other filterable relatives.
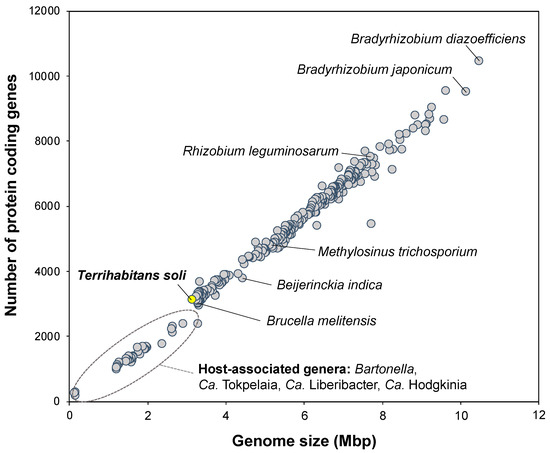
Figure 2.
Relationship between the genome size and the number of protein-coding genes in Hyphomicrobiales (Rhizobiales) genomes. A total of 373 genomic datasets registered with the genome completion status “finished” and the alphaproteobacterial orders “Hyphomicrobiales” and “Rhizobiales” were downloaded from the Integrated Microbial Genomes and Microbiomes (IMG/M) [33,34] (accessed on June 2021). The scientific names of some representative members, including our strain indicated by a yellow-filled circle, are shown; the host-associated (e.g., parasitic) members are circled by a gray dashed line.
3.2. Morphological, Physiological, and Biochemical Characteristics of Strain IZ6T
Cells of strain IZ6T were rods with a size of 1.5–2.0 × 0.4–0.5 µm (cell volume, ~0.36 μm3) under the 25 °C culture condition exhibiting good growth. This is not the size of typical ultramicrobacteria, which generally have a cell volume of <0.1 μm3 [3,5]. However, our previous study showed that smaller cells, as well as some ultra-small cells and cell-like particles (cell/particle volume, ~0.06 μm3), were observed when the temperature condition was changed from 25 °C to 15 °C [9]. This cell plasticity may be related to the ability to pass through 0.2 μm-pore-sized filters. The strain was Gram-stain negative, catalase-positive, and oxidase-positive. Growth was observed at 10–30 °C, but not at 4 °C or 37 °C, and at pH 6–10 with weak growth at pH 5 or 11; it should be noted that potential changes in pH during cultivation can affect the pH growth range data. Growth was also observed at 0–1.0% NaCl, with no growth at 1.5% NaCl or above. In the API 20NE and API ZYM tests, alkaline phosphatase, esterase, esterase lipase, acid phosphatase, and naphthol-AS-BI-phosphohydrolase were positive, and all others were negative.
The Hyphomicrobiales (Rhizobiales) group is phenotypically diverse and contains methanotrophs, methylotrophs, and nitrogen fixers [10,35]. In contrast, strain IZ6T exhibited no growth on methanol-amended medium and nitrogen-free medium. This result is consistent with the absence of the gene sequences encoding the key enzymes (i.e., methanol dehydrogenase and methane monooxygenase for one-carbon compound metabolism as well as nitrogenase for nitrogen fixation) in the IZ6T genome [9]. The previous genomic annotation also identified the presence of a nitrate reductase gene (nasA) in the genome [9], but the test for nitrate reduction by using API 20NE was negative under the condition tested. Further verification of this aspect of the characterization will be needed.
The characteristics of strain IZ6T lacking methanol utilization and nitrogen fixation capacity were shared with the closest type strain F. deserti [28]. However, in addition to differences in motility and growth temperature, pH, and salinity, several enzymatic activities also differed between IZ6T and F. deserti, indicating clear phenotype differences (Table 1). Moreover, strain IZ6T also showed a different trend from the phenotypic characteristics of strains of two other relatively related genera (Table 1).

Table 1.
Differential phenotypic characteristics of strain IZ6T, its closest type strain (Flaviflagellibacter deserti SYSU D60017T), and two other related strains of the Hyphomicrobiales (Rhizobiales).
3.3. Chemotaxonomic Characteristics of Strain IZ6T
The dominant cellular fatty acids (>5% of the total cellular fatty acids) were C18:1ω7c and/or C18:1ω6c (71.6%), C16:1ω7c and/or C16:1ω6c (5.9%), C19:0 cyclo ω8c (5.48%), and C16:0 (5.26%). Other minor fatty acids (0.5%~5%) were C17:0 (2.94%), C18:0 (2.38%), C16:0 3-OH (1.99%), C17:1ω6c (0.86), C12:0 3-OH (0.72%), C17:0 cyclo (0.62), and C14:0 (0.5%). The respiratory quinone of strain IZ6T was determined to be ubiquinone-10 (Q-10) that is a typical trait of Hyphomicrobiales (Rhizobiales) members [36]. The polar lipids detected were diphosphatidylglycerol, phosphatidylcholine, phosphatidylethanolamine, phosphatidylglycerol, and two unidentified polar lipids (Figure S1). The genomic G + C content was 62.2 mol%, as calculated from the IZ6T genome sequence (accession no. AP023361). The results of comparing these chemotaxonomic features with those of other relatives in the Hyphomicrobiales (Rhizobiales) are shown in Table 2. In particular, the unsaturated fatty acid(s) (C18:1ω7c/C18:1ω6c) was predominantly (>70% of the total cellular fatty acids) found in our strain, compared to the proportions (18–52%) of members of two related genera (Flaviflagellibacter and Pinisolibacter). This suggests that the unsaturated fatty acid-rich characteristic is one of the key features of strain IZ6T.
Table 2.
Differential chemotaxonomic characteristics of strain IZ6T and its three related genera of the Hyphomicrobiales (Rhizobiales).
3.4. Potential Distribution and Habitability of Close Relatives of Strain IZ6T
Comparative 16S rRNA gene sequence analysis revealed that strain IZ6T showed high sequence similarities to uncultured environmental sequences, as shown below. Environmentally derived sequences closely related (98.5–99.2%) to strain IZ6T were detected in a low-level-radioactive-waste site (accession no. GQ264001 [39]) and loamy soil (HQ119195 [40]) in the United States. Further, based on a search using the IMNGS platform [20], which is a database search against metagenome-derived 16S rRNA gene amplicon datasets, we also found that the IZ6T 16S rRNA gene sequence matched 3718 datasets, comprising mainly 2078 soil, 443 rhizosphere, and 219 plant metagenome datasets with a sequence similarity threshold of 99%. In agreement with this, the top three potential habitats inferred by using the ProkAtlas (which contains multiple 16S rRNA gene sequences labeled by one environmental category [21]) with a threshold of 99% were soil (habitat preference score, 45.4%), rhizosphere (31.0%), and plant (15.9%). These results suggested that this filterable bacterium and its close relatives inhabit different habitats in the terrestrial environment.
3.5. Proposal of a Novel Genus and Species for Strain IZ6T
Based on the clear phenotypic differences as well as the relatively low 16S rRNA gene sequence identity (<97%) and low AAI (<70%), strain IZ6T can be distinguished from the closest genus and species of F. deserti. F. deserti was first described as a novel bacterium closely clustered with Methylocystaceae clade II on the phylogenetic tree but with low sequence identity (<94%) to known neighbor genera [28]. Members of the Methylocystaceae lineage are known not to be a monophyletic group in the phylogenetic tree and are separated into four different clades [28,35]: clade I (type II methanotrophs: e.g., Methylocystis and Methylosinus), clade II (methylotrophs: e.g., Methylosulfonomonas, and Methylopila), clade III (non-methanotrophs: e.g., Chthonobacter and Pleomorphomonas), and clade IV (non-methanotrophs such as Terasakiella). In this context, it has been pointed out that the classification of members of the class Alphaproteobacteria is difficult because it relies on the interpretation of insufficiently resolved 16S rRNA gene-based trees for diverse members [10]. On the other hand, recent genome-scale phylogenetic analysis has improved the taxonomic classification of the alphaproteobacterial members, including the aforementioned genera related to Methylocystaceae [10]. Future studies will need to include strain IZ6T and F. deserti, as well as their relatives (e.g., Pinisolibacter; note that, as of June 2021, no genome information is available in public databases), in such genome-based taxonomic analyses to revisit their phylogenetic systematics.
On the basis of the phylogenetic, morphological, physiological, and chemotaxonomic traits, we propose the novel name Terrihabitans soli gen. nov., sp. nov. for strain IZ6T.
Description of Terrihabitans gen. nov.
Terrihabitans (Ter.ri.ha’bi.tans. L. fem. n. terra, earth; L. pres. part. habitans, inhabiting; N.L. part. adj. used as a masc. n. Terrihabitans, earth [soil] dweller, referring to the type of ecosystem inhabited by the bacteria).
Cells are rod-shaped and grow chemoorganotrophically and aerobically. Gram-stain negative, non-motile, and non-spore-forming. The cellular fatty acids are C18:1ω7c and/or C18:1ω6c, C16:1ω7c and/or C16:1ω6c, C19:0 cyclo ω8c, and C16:0. The respiratory quinone was ubiquinone-10 (Q-10). The polar lipids contain diphosphatidylglycerol, phosphatidylcholine, phosphatidylethanolamine, phosphatidylglycerol, and unknown polar lipids. The DNA G + C content is circa 62 mol%. The genome size is circa 3.1 Mbp.
The genus is a member of the class Alphaproteobacteria [41] of the phylum Proteobacteria [42]. The type species is Terrihabitans soli.
Description of Terrihabitans soli sp. nov.
Terrihabitans soli (so’li. L. gen. n. soli, of soil, the source of the type strain)
General descriptions of morphological and chemotaxonomic features are as given in the genus description. Cells grown in R2A liquid medium are rod-shaped with a size of about 1.5–2.0 × 0.4–0.5 µm (cell volume, ~0.36 μm3). Colonies on R2A agar medium are circular, light white, smooth, <1.0 mm in diameter, and slightly raised. Grows on R2A medium aerobically but not under anaerobic conditions. The temperature range for growth on R2A is 10 °C to 30 °C. Cells of reduced size, as well as some ultra-small cells and cell-like particles (cell/particle volume, ~0.06 μm3), are observed when the temperature condition is changed from 25 °C to 15 °C. The pH range for growth on R2A is pH 6–10 (note that potential changes in pH during cultivation can affect the pH growth range data). NaCl concentrations of 0–0.5% in R2A medium are tolerated, and weak growth occurs at 1%. Catalase-positive and oxidase-positive. Enzymatic activities are positive for alkaline phosphatase, esterase, esterase lipase, acid phosphatase, and naphthol-AS-BI-phosphohydrolase.
The type strain, strain IZ6T (=NBRC 106741T = NCIMB 15058T), was isolated from the 0.2 μm-filtered filtrates of a suspension of forest soil from Okuizumo, Shimane, Japan. The G + C content of the genomic DNA is 62.2 mol%, and the genome size is 3.11Mb. The genome sequence has been deposited in DDBJ/ENA/NCBI under accession no. AP023361.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/d13090422/s1. Figure S1: Polar lipids of strain IZ6T after separation by two-dimensional HPTLC. DPG, diphosphatidylglycerol; PC, phosphatidylcholine; PE, phosphatidylethanolamine; PG, phosphatidylglycerol; UL1 and UL2, unidentified polar lipids.
Author Contributions
Conceptualization, R.N.; methodology, experiments, and data analysis, R.N., N.T., T.K. (Tadao Kunihiro), S.M. and T.K. (Tomomi Koide); writing and visualization, R.N.; discussion, review, and editing, all authors; project supervision, R.N.; and funding acquisition, R.N., T.N. (Takeshi Naganuma), H.K. and H.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a Grant-in-Aid for JSPS Fellows (JSPS KAKENHI no. JP13J03441 to R.N.), a Grant-in-Aid for Challenging Exploratory Research (no. JP15K14424 to R.N.), a Grant-in-Aid for Young Scientists (A) (no. JP15H05620 to R.N.), and a Grant-in-Aid for Scientific Research (C) (no. JP23570117 to T. Naganuma). This study was also partially funded by a Grant-in-Aid for Challenging Exploratory Research (JSPS KAKENHI no. JP21K19394 to R.N.) and a Grant-in-Aid for Scientific Research on the Innovative Area “Post-Koch Ecology” (MEXT KAKENHI nos. JP19H05683 to R.N., H.K., H.T., and JP19H05679 to R.N.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Strain IZ6T is deposited at the Biological Resource Center, NITE (NBRC) in Japan and the National Collection of Industrial, Food, and Marine Bacteria (NCIMB) in the United Kingdom under accession nos. NBRC 106741T and NCIMB 15058T, respectively.
Acknowledgments
We thank Aharon Oren of the Hebrew University of Jerusalem (Jerusalem, Israel) for helpful comments and suggestions on nomenclature.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Levy, R.V.; Jornitz, M.W. Types of filtration. In Sterile Filtration; Jornitz, M.W., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 1–26. [Google Scholar] [CrossRef]
- Hahn, M.W. Broad Diversity of Viable Bacteria in ‘Sterile’ (0.2 μm) Filtered Water. Res. Microbiol. 2004, 155, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Duda, V.I.; Suzina, N.E.; Polivtseva, V.N.; Boronin, A.M. Ultramicrobacteria: Formation of the Concept and Contribution of Ultramicrobacteria to Biology. Microbiology 2012, 81, 379–390. [Google Scholar] [CrossRef]
- Ghuneim, L.-A.J.; Jones, D.L.; Golyshin, P.N.; Golyshina, O.V. Nano-Sized and Filterable Bacteria and Archaea: Biodiversity and Function. Front. Microbiol. 2018, 9, 1971. [Google Scholar] [CrossRef]
- Nakai, R. Size Matters: Ultra-Small and Filterable Microorganisms in the Environment. Microbes Environ. 2020, 35, ME20025. [Google Scholar] [CrossRef] [PubMed]
- Portillo, M.C.; Leff, J.W.; Lauber, C.L.; Fierer, N. Cell Size Distributions of Soil Bacterial and Archaeal Taxa. Appl. Environ. Microbiol. 2013, 79, 7610–7617. [Google Scholar] [CrossRef] [PubMed]
- Janssen, P.H.; Schuhmann, A.; Mörschel, E.; Rainey, F.A. Novel Anaerobic Ultramicrobacteria Belonging to the Verrucomicrobiales Lineage of Bacterial Descent Isolated by Dilution Culture from Anoxic Rice Paddy Soil. Appl. Environ. Microbiol. 1997, 63, 1382–1388. [Google Scholar] [CrossRef]
- Nakai, R.; Shibuya, E.; Justel, A.; Rico, E.; Quesada, A.; Kobayashi, F.; Iwasaka, Y.; Shi, G.-Y.; Amano, Y.; Iwatsuki, T.; et al. Phylogeographic Analysis of Filterable Bacteria with Special Reference to Rhizobiales Strains That Occur in Cryospheric Habitats. Antarct. Sci. 2013, 25, 219–228. [Google Scholar] [CrossRef]
- Nakai, R.; Naganuma, T.; Tazato, N.; Morohoshi, S.; Koide, T. Cell Plasticity and Genomic Structure of a Novel Filterable Rhizobiales Bacterium That Belongs to a Widely Distributed Lineage. Microorganisms 2020, 8, 1373. [Google Scholar] [CrossRef]
- Hördt, A.; López, M.G.; Meier-Kolthoff, J.P.; Schleuning, M.; Weinhold, L.-M.; Tindall, B.J.; Gronow, S.; Kyrpides, N.C.; Woyke, T.; Göker, M. Analysis of 1,000+ Type-Strain Genomes Substantially Improves Taxonomic Classification of Alphaproteobacteria. Front. Microbiol. 2020, 11, 468. [Google Scholar] [CrossRef]
- Volpiano, C.G.; Sant’Anna, F.H.; Ambrosini, A.; de São José, J.F.B.; Beneduzi, A.; Whitman, W.B.; de Souza, E.M.; Lisboa, B.B.; Vargas, L.K.; Passaglia, L.M.P. Genomic Metrics Applied to Rhizobiales (Hyphomicrobiales): Species Reclassification, Identification of Unauthentic Genomes and False Type Strains. Front. Microbiol. 2021, 12, 661. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.; Lim, J.; Kwon, S.; Chun, J. A Large-Scale Evaluation of Algorithms to Calculate Average Nucleotide Identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Varghese, N.J.; Mukherjee, S.; Ivanova, N.; Konstantinidis, K.T.; Mavrommatis, K.; Kyrpides, N.C.; Pati, A. Microbial Species Delineation Using Whole Genome Sequences. Nucleic Acids Res. 2015, 43, 6761–6771. [Google Scholar] [CrossRef]
- Rodriguez-R., L.M.; Konstantinidis, K.T. The Enveomics Collection: A Toolbox for Specialized Analyses of Microbial Genomes and Metagenomes. PeerJ Prepr. 2016, 4, e1900v1. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS Is an Automated High-Throughput Platform for State-of-the-Art Genome-Based Taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome Sequence-Based Species Delimitation with Confidence Intervals and Improved Distance Functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A Comprehensive, Accurate, and Fast Distance-Based Phylogeny Inference Program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef] [PubMed]
- Farris, J.S. Estimating Phylogenetic Trees from Distance Matrices. Am. Nat. 1972, 106, 645–668. [Google Scholar] [CrossRef]
- Kreft, Ł.; Botzki, A.; Coppens, F.; Vandepoele, K.; Van Bel, M. PhyD3: A Phylogenetic Tree Viewer with Extended PhyloXML Support for Functional Genomics Data Visualization. Bioinformatics 2017, 33, 2946–2947. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Joseph, D.; Kapfhammer, M.; Giritli, S.; Horn, M.; Haller, D.; Clavel, T. IMNGS: A Comprehensive Open Resource of Processed 16S rRNA Microbial Profiles for Ecology and Diversity Studies. Sci. Rep. 2016, 6, 33721. [Google Scholar] [CrossRef]
- Mise, K.; Iwasaki, W. Environmental Atlas of Prokaryotes Enables Powerful and Intuitive Habitat-Based Analysis of Community Structures. iScience 2020, 23, 101624. [Google Scholar] [CrossRef]
- Barrow, G.I.; Feltham, R.K.A. Cowan and Steel’s Manual for the Identification of Medical Bacteria, 3rd ed.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar] [CrossRef]
- Kim, D.; Kang, K.; Ahn, T.-Y. Chthonobacter Albigriseus Gen. Nov., Sp. Nov., Isolated from Grass-Field Soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 883–888. [Google Scholar] [CrossRef]
- Bao, Z.; Sato, Y.; Fujimura, R.; Ohta, H. Alsobacter Metallidurans Gen. Nov., Sp. Nov., a Thallium-Tolerant Soil Bacterium in the Order Rhizobiales. Int. J. Syst. Evol. Microbiol. 2014, 64, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Tamaoka, J.; Katayama-Fujimura, Y.; Kuraishi, H. Analysis of Bacterial Menaquinone Mixtures by High Performance Liquid Chromatography. J. Appl. Bacteriol. 1983, 54, 31–36. [Google Scholar] [CrossRef]
- Nakai, R.; Baba, T.; Niki, H.; Nishijima, M.; Naganuma, T. Aurantimicrobium Minutum Gen. Nov., Sp. Nov., a Novel Ultramicrobacterium of the Family Microbacteriaceae, Isolated from River Water. Int. J. Syst. Evol. Microbiol. 2015, 65, 4072–4079. [Google Scholar] [CrossRef]
- Dong, L.; Han, M.-X.; Wang, D.; Liu, F.; Asem, M.D.; Jiao, J.-Y.; Xiao, M.; Salam, N.; Li, W.-J. Flaviflagellibacter Deserti Gen. Nov., Sp. Nov., a Novel Member of the Order Rhizobiales Isolated from a Desert Soil. Antonie Van Leeuwenhoek 2019, 112, 947–954. [Google Scholar] [CrossRef]
- Barco, R.A.; Garrity, G.M.; Scott, J.J.; Amend, J.P.; Nealson, K.H.; Emerson, D.; Giovannoni Stephen, J. A Genus Definition for Bacteria and Archaea Based on a Standard Genome Relatedness Index. mBio 2020, 11, e02475-19. [Google Scholar] [CrossRef]
- Luo, C.; Rodriguez-R., L.M.; Konstantinidis, K.T. MyTaxa: An Advanced Taxonomic Classifier for Genomic and Metagenomic Sequences. Nucleic Acids Res. 2014, 42, e73. [Google Scholar] [CrossRef]
- Giovannoni, S.J. SAR11 Bacteria: The Most Abundant Plankton in the Oceans. Annu. Rev. Mar. Sci. 2017, 9, 231–255. [Google Scholar] [CrossRef]
- Hahn, M.W.; Scheuerl, T.; Jezberová, J.; Koll, U.; Jezbera, J.; Šimek, K.; Vannini, C.; Petroni, G.; Wu, Q.L. The Passive Yet Successful Way of Planktonic Life: Genomic and Experimental Analysis of the Ecology of a Free-Living Polynucleobacter Population. PLoS ONE 2012, 7, e32772. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-M.A.; Chu, K.; Palaniappan, K.; Ratner, A.; Huang, J.; Huntemann, M.; Hajek, P.; Ritter, S.; Varghese, N.; Seshadri, R.; et al. The IMG/M Data Management and Analysis System v.6.0: New Tools and Advanced Capabilities. Nucleic Acids Res. 2020, 49, D751–D763. [Google Scholar] [CrossRef]
- Mukherjee, S.; Stamatis, D.; Bertsch, J.; Ovchinnikova, G.; Sundaramurthi, J.C.; Lee, J.; Kandimalla, M.; Chen, I.-M.A.; Kyrpides, N.C.; Reddy, T.B.K. Genomes OnLine Database (GOLD) v.8: Overview and Updates. Nucleic Acids Res. 2020, 49, D723–D733. [Google Scholar] [CrossRef]
- Webb, H.K.; Ng, H.J.; Ivanova, E.P. The Family Methylocystaceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 341–347. [Google Scholar] [CrossRef]
- Dahal, R.H.; Chaudhary, D.K.; Kim, J. Pinisolibacter Ravus Gen. Nov., Sp. Nov., Isolated from Pine Forest Soil and Allocation of the Genera Ancalomicrobium and Pinisolibacter to the Family Ancalomicrobiaceae Fam. Nov., and Emendation of the Genus Ancalomicrobium Staley 1968. Int. J. Syst. Evol. Microbiol. 2018, 68, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-N.; Tian, W.-Y.; He, W.-H.; Chen, G.-C.; An, M.-L.; Jia, B.; Liu, L.; Zhou, Y.; Liu, S.-J. Methylopila Henanense Sp. Nov., a Novel Methylotrophic Bacterium Isolated from Tribenuron Methyl-Contaminated Wheat Soil. Antonie Van Leeuwenhoek 2015, 107, 329–336. [Google Scholar] [CrossRef]
- Doronina, N.V.; Trotsenko, Y.A. Incertae Sedis IV. Methylopila. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Rainey, F., Kämpfer, P., Trujillo, M., Chun, J., DeVos, P., Hedlund, B., Dedysh, S., Eds.; Wiley: Hoboken, NJ, USA, 2015; pp. 1–4. [Google Scholar] [CrossRef]
- Field, E.K.; D’Imperio, S.; Miller, A.R.; VanEngelen, M.R.; Gerlach, R.; Lee, B.D.; Apel, W.A.; Peyton, B.M. Application of Molecular Techniques to Elucidate the Influence of Cellulosic Waste on the Bacterial Community Structure at a Simulated Low-Level-Radioactive-Waste Site. Appl. Environ. Microbiol. 2010, 76, 3106–3115. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Williamson, K.E.; Kan, J.; Polson, S.W.; Williamson, S.J. Optimizing the Indirect Extraction of Prokaryotic DNA from Soils. Soil Biol. Biochem. 2011, 43, 736–748. [Google Scholar] [CrossRef]
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Alphaproteobacteria class. nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Rainey, F., Kämpfer, P., Trujillo, M., Chun, J., DeVos, P., Hedlund, B., Dedysh, S., Eds.; Wiley: Hoboken, NJ, USA, 2015; p. 1. [Google Scholar] [CrossRef]
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Proteobacteria phyl. nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Rainey, F., Kämpfer, P., Trujillo, M., Chun, J., DeVos, P., Hedlund, B., Dedysh, S., Eds.; Wiley: Hoboken, NJ, USA, 2015; p. 1. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).