
Large-Scale Patterns of Soil Nematodes across Grasslands on the Tibetan Plateau: Relationships with Climate, Soil and Plants

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
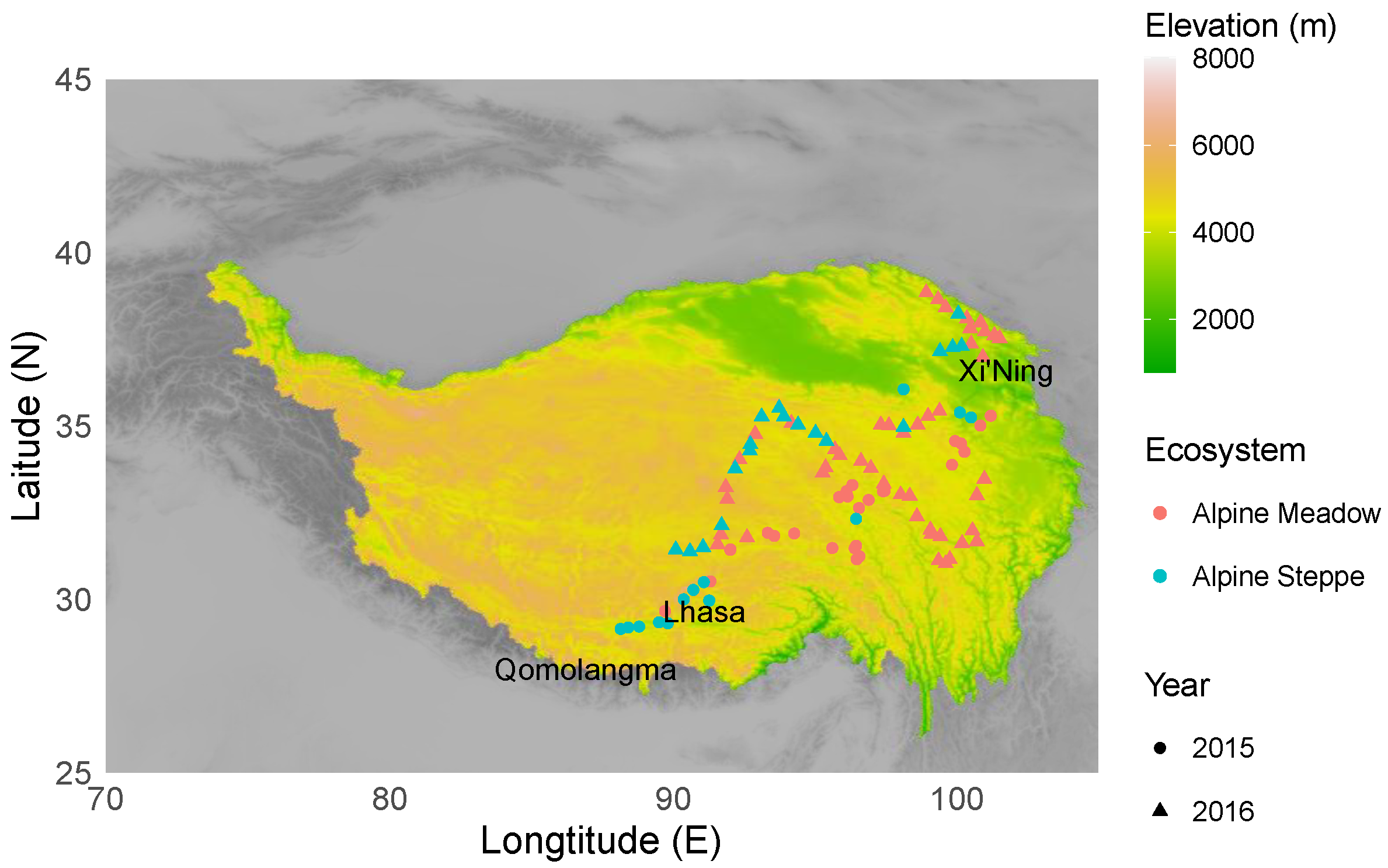
2.1. Sampling Locations
2.2. Aboveground Plant and Soil Sampling
2.3. Nematode Extraction and Identification
2.4. Measurement of Soil Physicochemical Properties
2.5. High-Resolution Gridded Dataset of Temperature and Precipitation
2.6. Data Analyses
3. Results
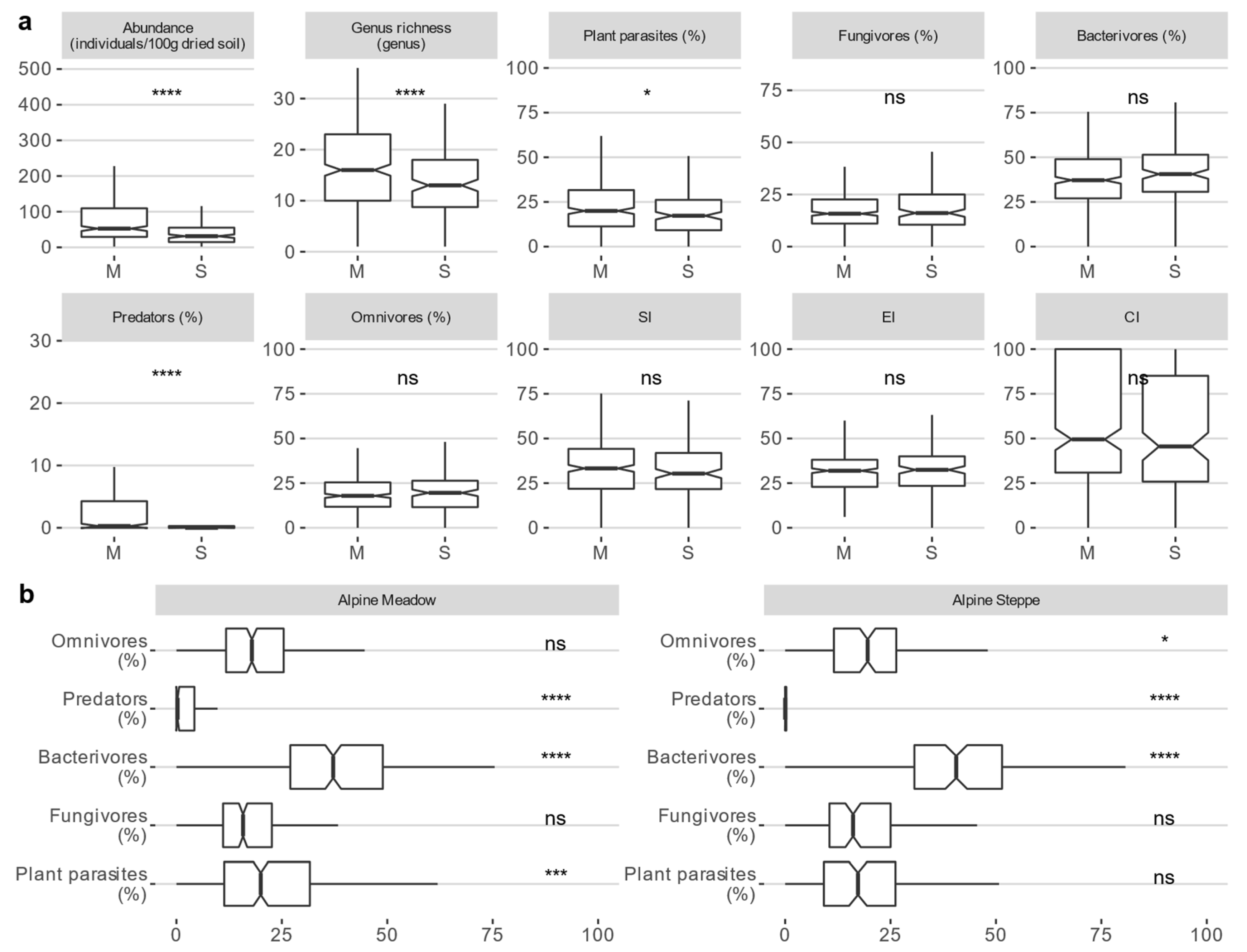
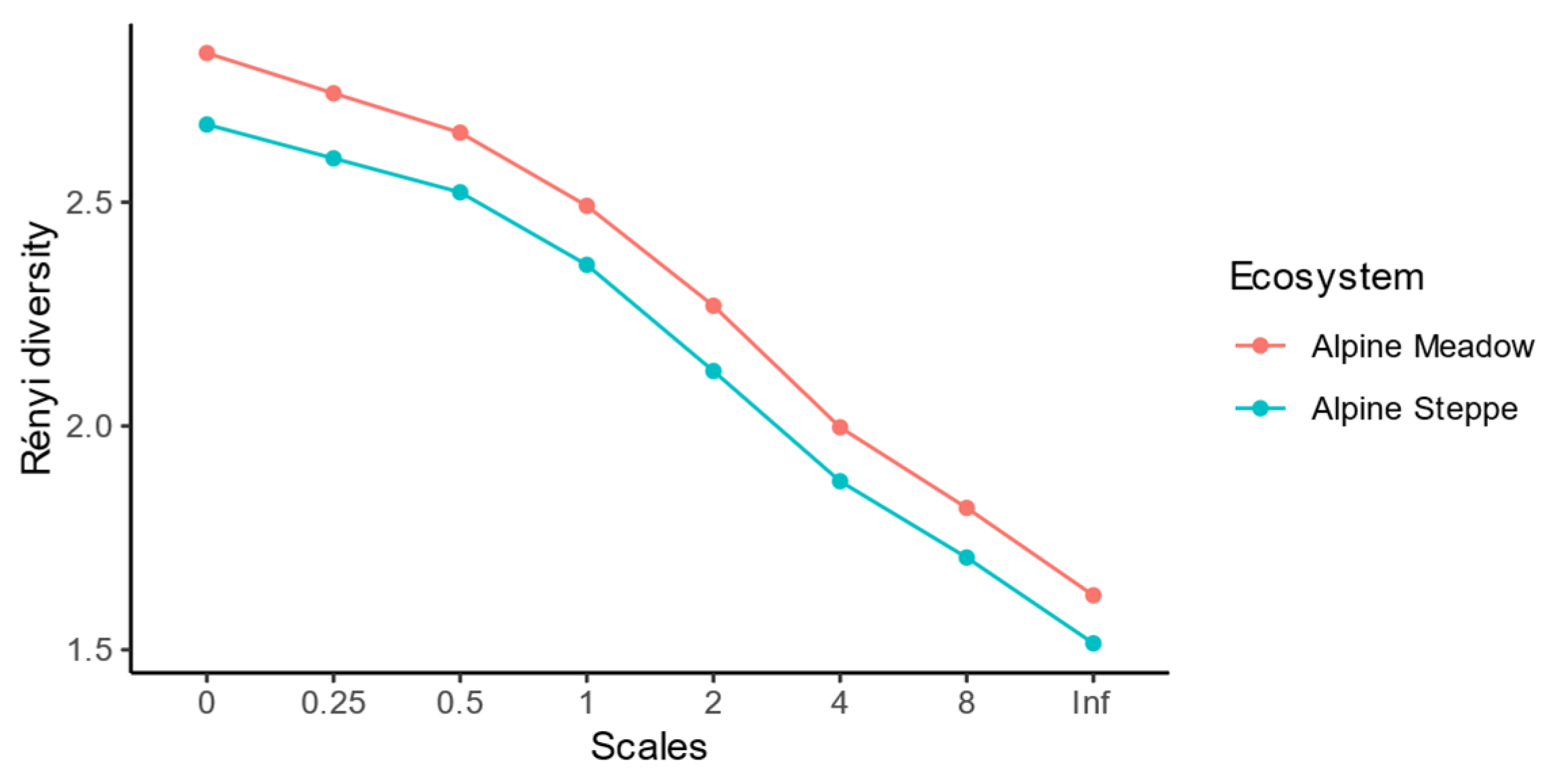
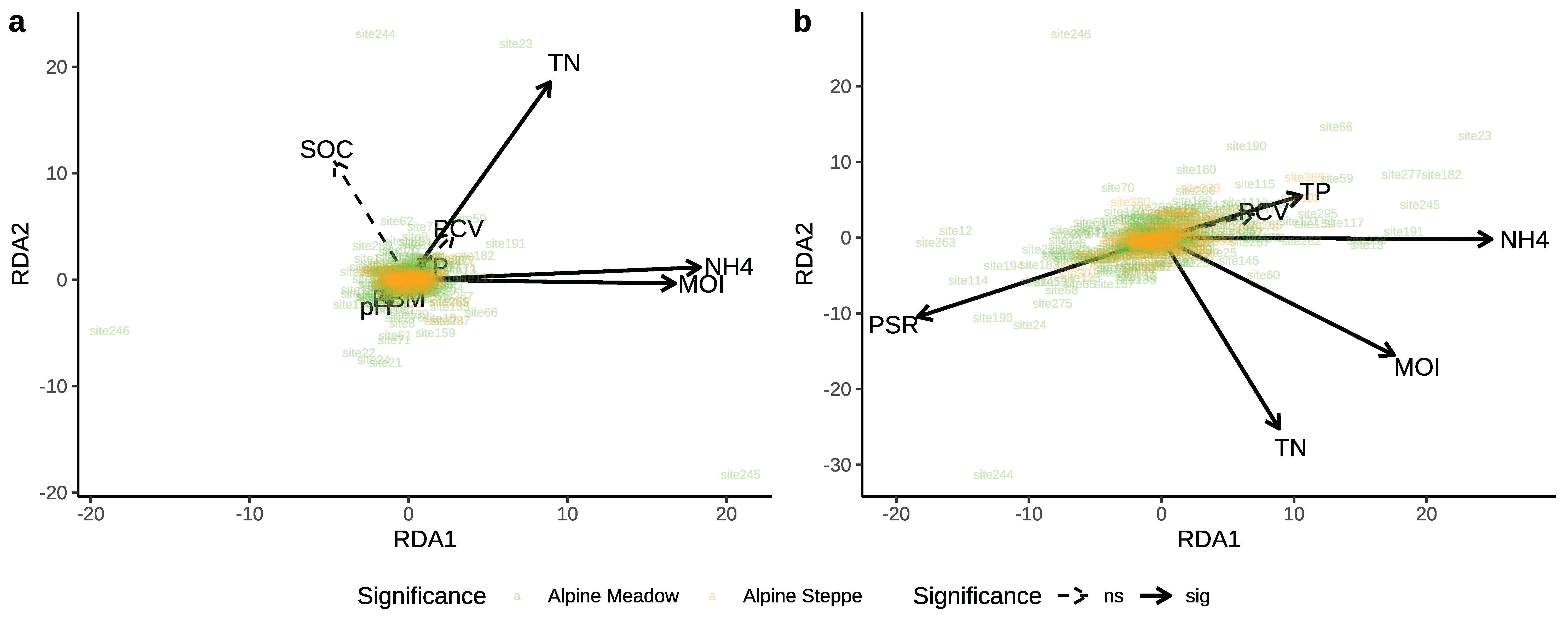
3.1. Soil Nematode Community in the Alpine Ecosystem on the Tibetan Plateau
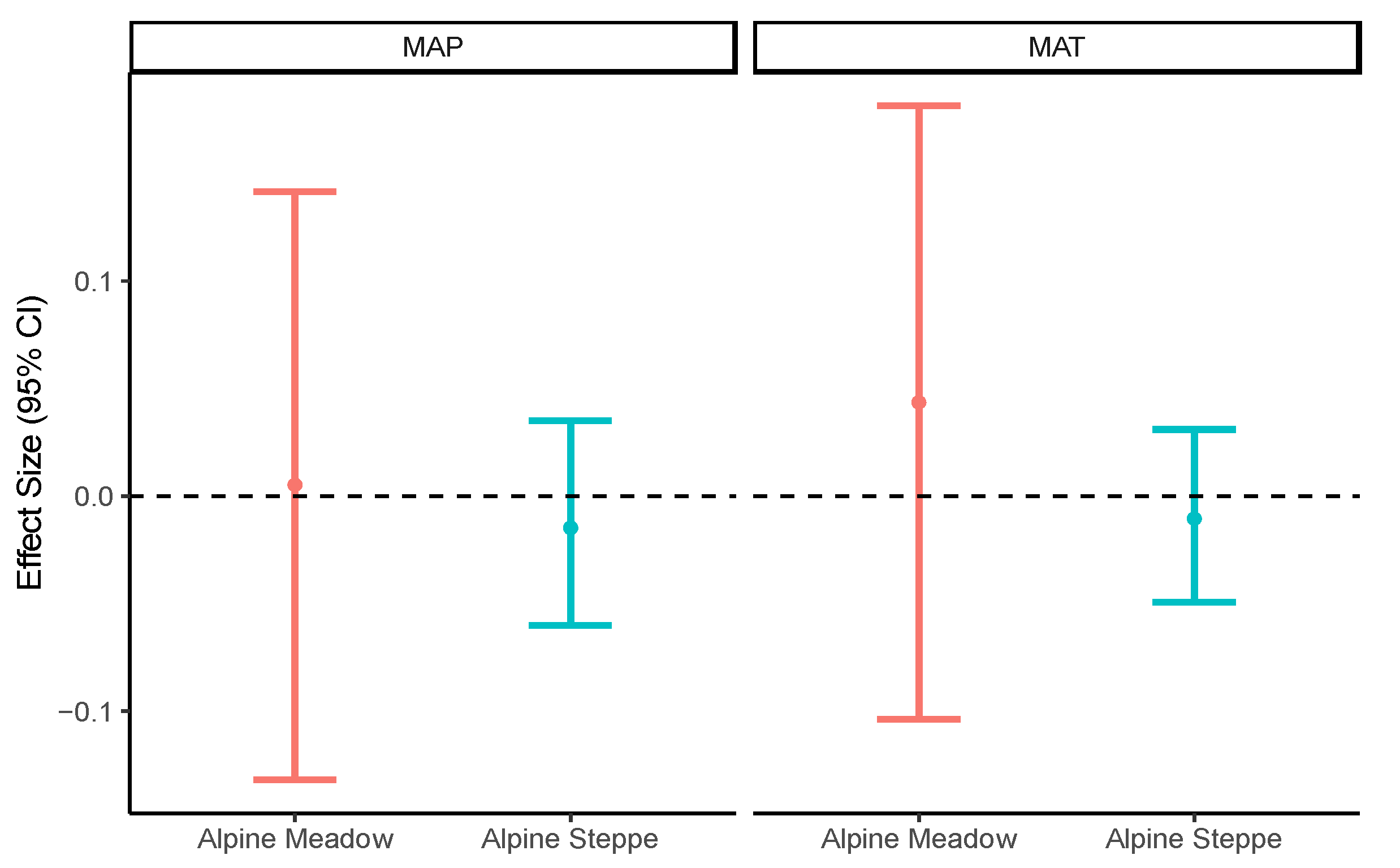
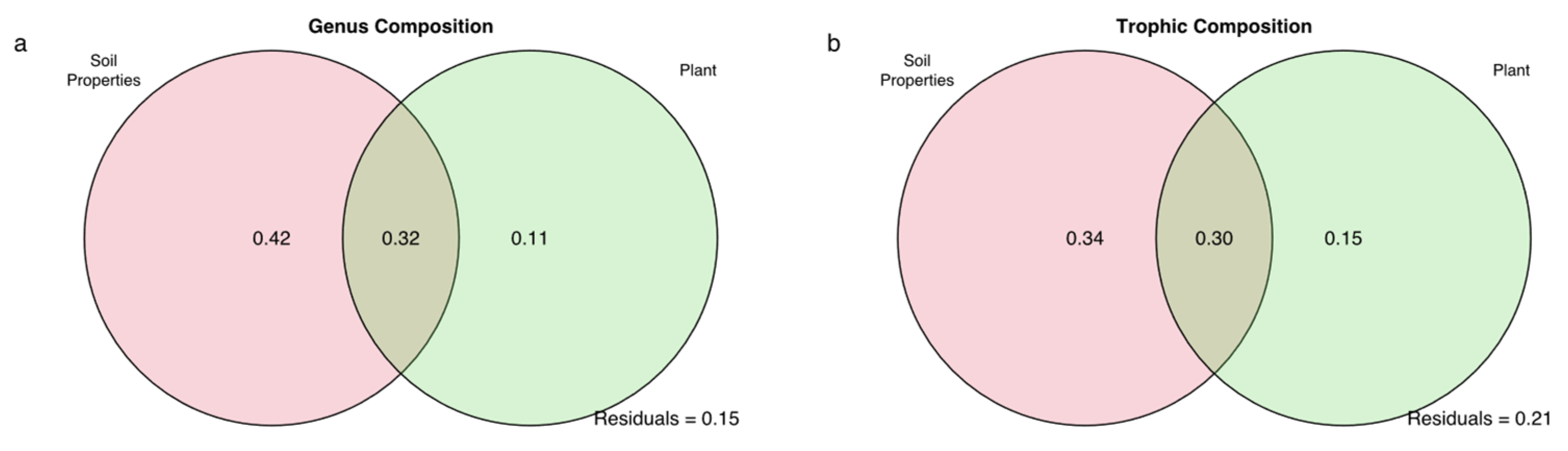
3.2. Effects of Biotic and Abiotic Factors on Nematode Communities
4. Discussion
4.1. Difference in Nematode Diversity and Characteristics between Alpine Meadow and Steppe Ecosystems
4.2. Roles of Biotic and Abiotic Factors in Relation to the Nematode Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lambshead, P.J.D. Recent Developments in Marine Benthic Biodiversity Reserch. Oceanis 1993, 19, 5–24. [Google Scholar]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding Habits in Soil Nematode Families and Genera-an Outline for Soil Ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Bongers, T.; Bongers, M. Functional Diversity of Nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Laakso, J.; Setälä, H. Population- and Ecosystem-Level Effects of Predation on Microbial-Feeding Nematodes. Oecologia 1999, 120, 279–286. [Google Scholar] [CrossRef]
- Scheu, S. The Soil Food Web: Structure and Perspectives. Eur. J. Soil Biol. 2002, 38, 11–20. [Google Scholar] [CrossRef]
- Zhao, J.; Neher, D.A. Soil Energy Pathways of Different Ecosystems Using Nematode Trophic Group Analysis: A Meta Analysis. Nematology 2014, 16, 379–385. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T.; de Goede, R.G.M. A Framework for Soil Food Web Diagnostics: Extension of the Nematode Faunal Analysis Concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Freckman, D.W.; Ettema, C.H. Assessing Nematode Communities in Agroecosystems of Varying Human Intervention. Agric. Ecosyst. Environ. 1993, 45, 239–261. [Google Scholar] [CrossRef]
- Yeates, G.W. Nematodes as Soil Indicators: Functional and Biodiversity Aspects. Biol. Fertil. Soils 2003, 37, 199–210. [Google Scholar] [CrossRef]
- Ritz, K.; Trudgill, D.L. Utility of Nematode Community Analysis as an Integrated Measure of the Functional State of Soils: Perspectives and Challenges. Plant Soil 1999, 212, 1–11. [Google Scholar] [CrossRef]
- Yeates, G.; Boag, B. Background for nematode ecology in the 21st century. In Nematology: Advances and Perspectives; CABI Publishing: Wallingford, UK, 2004; Volume 1, pp. 406–437. [Google Scholar]
- Nielsen, U.N.; Ayres, E.; Wall, D.H.; Li, G.; Bardgett, R.D.; Wu, T.; Garey, J.R. Global-scale Patterns of Assemblage Structure of Soil Nematodes in Relation to Climate and Ecosystem Properties. Glob. Ecol. Biogeogr. 2014, 23, 968–978. [Google Scholar] [CrossRef]
- Pey, B.; Nahmani, J.; Auclerc, A.; Capowiez, Y.; Cluzeau, D.; Cortet, J.; Decaëns, T.; Deharveng, L.; Dubs, F.; Joimel, S.; et al. Current Use of and Future Needs for Soil Invertebrate Functional Traits in Community Ecology. Basic Appl. Ecol. 2014, 15, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Geisen, S.; Briones, M.J.I.; Gan, H.; Behan-Pelletier, V.M.; Friman, V.-P.; de Groot, G.A.; Hannula, S.E.; Lindo, Z.; Philippot, L.; Tiunov, A.V.; et al. A Methodological Framework to Embrace Soil Biodiversity. Soil Biol. Biochem. 2019, 136, 107536. [Google Scholar] [CrossRef]
- Fierer, N.; Strickland, M.S.; Liptzin, D.; Bradford, M.A.; Cleveland, C.C. Global Patterns in Belowground Communities. Ecol. Lett. 2009, 12, 1238–1249. [Google Scholar] [CrossRef]
- Chen, D.; Cheng, J.; Chu, P.; Hu, S.; Xie, Y.; Tuvshintogtokh, I.; Bai, Y. Regional-scale Patterns of Soil Microbes and Nematodes across Grasslands on the Mongolian Plateau: Relationships with Climate, Soil, and Plants. Ecography 2015, 38, 622–631. [Google Scholar] [CrossRef]
- Zhao, J.; Xun, R.; He, X.; Zhang, W.; Fu, W.; Wang, K. Size Spectra of Soil Nematode Assemblages under Different Land Use Types. Soil Biol. Biochem. 2015, 85, 130–136. [Google Scholar] [CrossRef]
- Jing, X.; Sanders, N.J.; Shi, Y.; Chu, H.; Classen, A.T.; Zhao, K.; Chen, L.; Shi, Y.; Jiang, Y.; He, J.-S. The Links between Ecosystem Multifunctionality and Above- and Belowground Biodiversity Are Mediated by Climate. Nat. Commun. 2015, 6, 8159. [Google Scholar] [CrossRef] [PubMed]
- Neher, D.A. Nematode Communities in Organically and Conventionally Managed Agricultural Soils. J. Nematol. 1999, 31, 142–154. [Google Scholar]
- Ruan, W.; Sang, Y.; Chen, Q.; Zhu, X.; Lin, S.; Gao, Y. The Response of Soil Nematode Community to Nitrogen, Water, and Grazing History in the Inner Mongolian Steppe, China. Ecosystems 2012, 15, 1121–1133. [Google Scholar] [CrossRef]
- Ruess, L.; Michelsen, A.; Schmidt, I.K.; Jonasson, S. Simulated Climate Change Affecting Microorganisms, Nematode Density and Biodiversity in Subarctic Soils. Plant Soil 1999, 212, 63–73. [Google Scholar] [CrossRef]
- Van den Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil Nematode Abundance and Functional Group Composition at a Global Scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Chen, H.; Zhang, Y. Latitudinal Variation in Nematode Diversity and Ecological Roles along the Chinese Coast. Ecol. Evol. 2016, 6, 8018–8027. [Google Scholar] [CrossRef] [Green Version]
- Traunspurger, W.; Reiff, N.; Krashevska, V.; Majdi, N.; Scheu, S. Diversity and Distribution of Soil Micro-Invertebrates across an Altitudinal Gradient in a Tropical Montane Rainforest of Ecuador, with Focus on Free-Living Nematodes. Pedobiologia 2017, 62, 28–35. [Google Scholar] [CrossRef]
- Liu, T.; Hu, F.; Li, H. Spatial Ecology of Soil Nematodes: Perspectives from Global to Micro Scales. Soil Biol. Biochem. 2019, 137, 107565. [Google Scholar] [CrossRef]
- Viketoft, M. Determinants of Small-Scale Spatial Patterns: Importance of Space, Plants and Abiotics for Soil Nematodes. Soil Biol. Biochem. 2013, 62, 92–98. [Google Scholar] [CrossRef]
- Li, X.; Zhu, H.; Geisen, S.; Bellard, C.; Hu, F.; Li, H.; Chen, X.; Liu, M. Agriculture Erases Climate Constraints on Soil Nematode Communities across Large Spatial Scales. Glob. Chang. Biol. 2020, 26, 919–930. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Luo, Y. Meta-Analysis of the Impacts of Global Change Factors on Soil Microbial Diversity and Functionality. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Li, W.; Zhou, X. Ecosystems of Qinghai-Xizang (Tibetan) Plateau and Approach for Their Sustainable Manag; The series of studies on Qinghai-Xizang (Tibetan) Plateau; Guang-dong Science & Technology Press: Guangzhou, China, 1998; ISBN 978-7-5359-1945-8. [Google Scholar]
- Li, X.-L.; Gao, J.; Brierley, G.; Qiao, Y.-M.; Zhang, J.; Yang, Y.-W. Rangeland degradation on the oinghai-tibet plateau: Implications for rehabilitation. Land Degrad. Develop. 2013, 24, 72–80. [Google Scholar] [CrossRef]
- Qi, W.; Zhou, X.; Ma, M.; Knops, J.M.H.; Li, W.; Du, G. Elevation, Moisture and Shade Drive the Functional and Phylogenetic Meadow Communities’ Assembly in the Northeastern Tibetan Plateau. Community Ecol. 2015, 16, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Dalling, J.W.; Ma, Z.; Zhou, X. Soil Environmental Factors Drive Seed Density across Vegetation Types on the Tibetan Plateau. Plant. Soil 2017, 374, 1–13. [Google Scholar] [CrossRef]
- Wu, H.; Guo, Z.; Peng, C. Distribution and Storage of Soil Organic Carbon in China: Soil organic carbon in China. Glob. Biogeochem. Cycles. 2003, 17, 1043. [Google Scholar] [CrossRef]
- Han, L. Viewing the Sustainable Development of the Qinghai-Tibet Plateau from Water Resources System. IOP Conf. Ser. Earth Environ. Sci. 2018, 170, 032083. [Google Scholar] [CrossRef] [Green Version]
- Mu, C.; Abbott, B.W.; Norris, A.J.; Mu, M.; Fan, C.; Chen, X.; Jia, L.; Yang, R.; Zhang, T.; Wang, K.; et al. The Status and Stability of Permafrost Carbon on the Tibetan Plateau. Earth-Sci. Rev. 2020, 211, 103433. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Wardle, D.A. Oxford Series in Ecology and Evolution. In Aboveground-Belowground Linkages; Oxford University Press: Oxford, UK, 2010; ISBN 978-0-19-954688-6. [Google Scholar]
- De Vries, F.T.; Manning, P.; Tallowin, J.R.B.; Mortimer, S.R.; Pilgrim, E.S.; Harrison, K.A.; Hobbs, P.J.; Quirk, H.; Shipley, B.; Cornelissen, J.H.C.; et al. Abiotic Drivers and Plant Traits Explain Landscape-Scale Patterns in Soil Microbial Communities. Ecol. Lett. 2012, 15, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-P.; Huang, M.-T.; Zhai, P.-M. Change in Drought Conditions and Its Impacts on Vegetation Growth over the Tibetan Plateau. Adv. Clim. Chang. Res. 2021, 12, 333–341. [Google Scholar] [CrossRef]
- Piao, S.; Zhang, X.; Wang, T.; Liang, E.; Wang, S.; Zhu, J.; Niu, B. Responses and Feedback of the Tibetan Plateau’s Alpine Ecosystem to Climate Change. Chin. Sci. Bull. 2019, 64, 2842–2855. [Google Scholar] [CrossRef]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and Its Drivers. Nat. Clim. Chang. 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, S.; Niu, B.; Chen, Q.; Wang, J.; Zhao, J.; Luo, T.; Zhang, G. Effect of Increasing Precipitation and Warming on Microbial Community in Tibetan Alpine Steppe. Environ. Res. 2020, 189, 109917. [Google Scholar] [CrossRef]
- Zhao, J.; Luo, T.; Wei, H.; Deng, Z.; Li, X.; Li, R.; Tang, Y. Increased Precipitation Offsets the Negative Effect of Warming on Plant Biomass and Ecosystem Respiration in a Tibetan Alpine Steppe. Agric. For. Meteorol. 2019, 279, 107761. [Google Scholar] [CrossRef]
- Zhao, K.; Jing, X.; Sanders, N.J.; Chen, L.; Shi, Y.; Flynn, D.F.B.; Wang, Y.; Chu, H.; Liang, W.; He, J.-S. On the Controls of Abundance for Soil-Dwelling Organisms on the Tibetan Plateau. Ecosphere 2017, 8, e01901. [Google Scholar] [CrossRef] [Green Version]
- Bell, C.W.; Acosta-Martinez, V.; McIntyre, N.E.; Cox, S.; Tissue, D.T.; Zak, J.C. Linking Microbial Community Structure and Function to Seasonal Differences in Soil Moisture and Temperature in a Chihuahuan Desert Grassland. Microb Ecol. 2009, 58, 827–842. [Google Scholar] [CrossRef]
- Geng, Y.; Wang, Y.; Yang, K.; Wang, S.; Zeng, H.; Baumann, F.; Kuehn, P.; Scholten, T.; He, J.-S. Soil Respiration in Tibetan Alpine Grasslands: Belowground Biomass and Soil Moisture, but Not Soil Temperature, Best Explain the Large-Scale Patterns. PLOS ONE 2012, 7, e34968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, M.; Collins, S.L.; Du, G. Direct and Indirect Effects of Temperature and Precipitation on Alpine Seed Banks in the Tibetan Plateau. Ecol. Appl. 2020, 30. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Campbell, C.D.; Chapman, S.J.; Freitag, T.E.; Nicol, G.W.; Singh, B.K. Multi-Factorial Drivers of Ammonia Oxidizer Communities: Evidence from a National Soil Survey: Multi-Factorial Drivers of Ammonia Oxidizer Communities. Environ. Microbiol. 2013, 15, 2545–2556. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Mi, J.; Chu, P.; Cheng, J.; Zhang, L.; Pan, Q.; Xie, Y.; Bai, Y. Patterns and Drivers of Soil Microbial Communities along a Precipitation Gradient on the Mongolian Plateau. Landsc. Ecol. 2014, 30, 1669–1682. [Google Scholar] [CrossRef]
- Southey, J.F. Laboratory Methods for Work with Plant. and Soil Nematodes, 6th ed.; H.M.S.O. Books: London, UK, 1986; ISBN 0-11-242754-5. [Google Scholar]
- Goodey, T.; Goodey, J.B. Soil and Freshwater Nematodes, 2nd ed.; Methuen, Wiley: London, UK, 1963; ISBN B0019Y180M. [Google Scholar]
- Jairajpuri, M.S.; Ahmad, W. Dorylaimida: Free-Living, Predaceous and Plant.-Parasitic Nematodes; Brill: Leiden, The Netherlands; New York, NY, USA, 1992; ISBN 978-90-04-09229-7. [Google Scholar]
- Warwick, R.M.; Platt, H.M.; Somerfield, P.J. Freeliving Marine Nematodes: Part. III. Monhysterida. Synopses of the British Fauna No. 53; Field Studies Council: Shrewsbury, UK, 1998. [Google Scholar]
- Tóthmérész, B. Comparison of Different Methods for Diversity Ordering. J. Veg. Sci. 1995, 6, 283–290. [Google Scholar] [CrossRef]
- Kindt, R.; Van Damme, P.; Simons, A.J. Tree diversity in western Kenya: Using profiles to characterise richness and evenness. In Forest Diversity and Management; Hawksworth, D.L., Bull, A.T., Eds.; Topics in Biodiversity and Conservation; Springer: Dordrecht, The Netherland, 2006; pp. 193–210. ISBN 978-1-4020-5208-8. [Google Scholar]
- Gardner, W.H. Water Content. In Methods of Soil Analysis; Klute, A., Ed.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 82–187. ISBN 978-0-89118-811-7. [Google Scholar]
- Allen, W.; Armstrong, B.I. An Examination of the Degtjareff Method for Determining Soil Organic Matter, and a Proposed Modification of the Chromic Acid Titration Method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Bray, B.; Kurtz, L.T. Determination of Total, Organic, and Available Forms of Phosphorus in Soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Bao, S. Soil Agro-Chemistrical Analysis (in Chinese), 1st ed.; Textbook Series for 21st Century; Chinese Agricultural Press: Beijing, China, 2000; ISBN 978-7-109-06644-1. [Google Scholar]
- Cort J., W.; Kenji, M. University of Delaware Air Temperature & Precipitation (V5.01). Available online: https://www.esrl.noaa.gov/psd/data/gridded/data.UDel_AirT_Precip.html (accessed on 10 December 2019).
- Legates, D.R.; Willmott, C.J. Mean Seasonal and Spatial Variability in Global Surface Air Temperature. Appl. Clim. 1990, 41, 11–21. [Google Scholar] [CrossRef]
- Willmott, C.J.; Matsuura, K. Smart Interpolation of Annually Averaged Air Temperature in the United States. J. Appl. Meteor. 1995, 34, 2577–2586. [Google Scholar] [CrossRef]
- Willmott, C.J.; Rowe, C.M.; Philpot, W.D. Small-Scale Climate Maps: A Sensitivity Analysis of Some Common Assumptions Associated with Grid-Point Interpolation and Contouring. Am. Cartogr. 1985, 12, 5–16. [Google Scholar] [CrossRef]
- Willmott, C.J.; Robeson, S.M. Climatologically aided interpolation (CAI) of terrestrial air temperature. Int. J. Climatol. 1995, 15, 221–229. [Google Scholar] [CrossRef]
- Koutsouris, A.J.; Chen, D.; Lyon, S.W. Comparing Global Precipitation Data Sets in Eastern Africa: A Case Study of Kilombero Valley, Tanzania: Comparing Global Precipitation Data Sets in Tanzania, East Africa. Int. J. Climatol. 2016, 36, 2000–2014. [Google Scholar] [CrossRef]
- Hosseini-Moghari, S.-M.; Araghinejad, S.; Ebrahimi, K. Spatio-Temporal Evaluation of Global Gridded Precipitation Datasets across Iran. Hydrol. Sci. J. 2018, 63, 1669–1688. [Google Scholar] [CrossRef]
- Kanda, N.; Negi, H.S.; Rishi, M.; Kumar, A. Performance of Various Gridded Temperature and Precipitation Datasets over Northwest Himalayan Region. Environ. Res. Commun. 2020, 2, 085002. [Google Scholar] [CrossRef]
- Oksanen, J.; Kindt, R.; Legendre, P.; Hara, B.; Henry, M.; Stevens, H. The Vegan Package. Community Ecol. Package 2007, 10, 719. [Google Scholar]
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.L.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H.; RStudio. Ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics. 2019. Available online: https://CRAN.R-project.org/package=ggplot2 (accessed on 15 June 2021).
- Douglas, C.E.; Michael, F.A. On Distribution-Free Multiple Comparisons in the One-Way Analysis of Variance. Commun. Stat. Theory Methods 1991, 20, 127–139. [Google Scholar] [CrossRef]
- Hsu, J. Multiple Comparisons: Theory and Methods, 1st ed.; Chapman and Hall/CRC: London, UK, 1996; ISBN 978-0-412-98281-1. [Google Scholar]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the Spatial Component of Ecological Variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Legendre, L.; Legendre, L.; Legendre, L. Developments in environmental modelling. In Numerical Ecology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 1998; ISBN 978-0-444-89249-2. [Google Scholar]
- Legendre, P.; Oksanen, J.; Braak, C.J.F. ter Testing the Significance of Canonical Axes in Redundancy Analysis. Methods Ecol. Evol. 2011, 2, 269–277. [Google Scholar] [CrossRef]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation Partitioning of Species Data Matrices: Estimation and Comparison of Fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Lindeman, R.H.; Merenda, P.F.; Gold, R.Z. Introduction to Bivariate and Multivariate Analysis; Scott Foresman: Brook, IL, USA, 1980; ISBN 978-0-673-15099-8. [Google Scholar]
- Groemping, U.; Matthias, L. Relaimpo: Relative Importance of Regressors in Linear Models. 2018. Available online: https://CRAN.R-project.org/package=relaimpo (accessed on 15 June 2021).
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, J.; Fisher, D.N.; Goodwin, C.E.D.; Robinson, B.S.; Hodgson, D.J.; Inger, R. A Brief Introduction to Mixed Effects Modelling and Multi-Model Inference in Ecology. PeerJ 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Berkelmans, R.; Ferris, H.; Tenuta, M.; van Bruggen, A.H.C. Effects of Long-Term Crop Management on Nematode Trophic Levels Other than Plant Feeders Disappear after 1 Year of Disruptive Soil Management. Appl. Soil Ecol. 2003, 23, 223–235. [Google Scholar] [CrossRef]
- Anderson, M.J.; Willis, T.J. Canonical Analysis of Principal Coordinates: A Useful Method of Constrained Ordination for Ecology. Ecology 2003, 84, 511–525. [Google Scholar] [CrossRef]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate Dispersion as a Measure of Beta Diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Moreno, S.; Minoshima, H.; Ferris, H.; Jackson, L.E. Linking Soil Properties and Nematode Community Composition: Effects of Soil Management on Soil Food Webs. Nematology 2006, 8, 703–715. [Google Scholar] [CrossRef]
- Nielsen, U.N.; Osler, G.H.R.; Campbell, C.D.; Burslem, D.F.R.P.; van der Wal, R. The Influence of Vegetation Type, Soil Properties and Precipitation on the Composition of Soil Mite and Microbial Communities at the Landscape Scale. J. Biogeogr. 2010, 37, 1317–1328. [Google Scholar] [CrossRef]
- Ingham, E.R.; Moldenke, A.R.; Edwards, C.A. Soil Biology Primer; Tugel, A.J., Lewandowski, A.M., Happe-vonArb, D., Eds.; Soil and Water Conservation Society: Ankeny, IA, USA, 2000. [Google Scholar]
- Yeates, G.W.; Wardle, D.A.; Watson, R.N. Responses of Soil Nematode Populations, Community Structure, Diversity and Temporal Variability to Agricultural Intensification over a Seven-Year Period. Soil Biol. Biochem. 1999, 31, 1721–1733. [Google Scholar] [CrossRef]
- Freestone, A.L.; Inouye, B.D. Dispersal Limitation and Environmental Heterogeneity Shape Scale-Dependent Diversity Patterns in Plant Communities. Ecology 2006, 87, 2425–2432. [Google Scholar] [CrossRef]
- Horner-Devine, M.C.; Carney, K.M.; Bohannan, B.J.M. An Ecological Perspective on Bacterial Biodiversity. Proc. R. Soc. London Ser. B Biol. Sci. 2004, 271, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Ranjard, L.; Dequiedt, S.; Chemidlin Prévost-Bouré, N.; Thioulouse, J.; Saby, N.P.A.; Lelievre, M.; Maron, P.A.; Morin, F.E.R.; Bispo, A.; Jolivet, C.; et al. Turnover of Soil Bacterial Diversity Driven by Wide-Scale Environmental Heterogeneity. Nat. Commun. 2013, 4, 1434. [Google Scholar] [CrossRef]
- Flynn, D.F.B.; Gogol-Prokurat, M.; Nogeire, T.; Molinari, N.; Richers, B.T.; Lin, B.B.; Simpson, N.; Mayfield, M.M.; DeClerck, F. Loss of Functional Diversity under Land Use Intensification across Multiple Taxa. Ecol. Lett. 2009, 12, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ding, J.; Peng, Y.; Li, F.; Yang, G.; Liu, L.; Qin, S.-Q.; Fang, K.; Yang, Y.-H. Patterns and Drivers of Soil Microbial Communities in Tibetan Alpine and Global Terrestrial Ecosystems. J. Biogeogr. 2016, 43, 2027–2039. [Google Scholar] [CrossRef]
- Boag, B.; Yeates, G.W. Soil Nematode Biodiversity in Terrestrial Ecosystems. Biodivers. Conserv. 1998, 7, 617–630. [Google Scholar] [CrossRef]
- Cam, E.; Nichols, J.D.; Hines, J.E.; Sauer, J.R.; Alpizar-Jara, R.; Flather, C.H. Disentangling Sampling and Ecological Explanations Underlying Species–Area Relationships. Ecology 2002, 83, 1118–1130. [Google Scholar] [CrossRef]
- Woodcock, S.; Curtis, T.P.; Head, I.M.; Lunn, M.; Sloan, W.T. Taxa-Area Relationships for Microbes: The Unsampled and the Unseen. Ecol. Lett. 2006, 9, 805–812. [Google Scholar] [CrossRef]
- Meyer, K.M.; Memiaghe, H.; Korte, L.; Kenfack, D.; Alonso, A.; Bohannan, B.J.M. Why Do Microbes Exhibit Weak Biogeographic Patterns? ISME J. 2018, 12, 1404–1413. [Google Scholar] [CrossRef]
- Kerfahi, D.; Tripathi, B.M.; Porazinska, D.L.; Park, J.; Go, R.; Adams, J.M. Do Tropical Rain Forest Soils Have Greater Nematode Diversity than High Arctic Tundra? A Metagenetic Comparison of Malaysia and Svalbard. Glob. Ecol. Biogeogr. 2016, 25, 716–728. [Google Scholar] [CrossRef]
- Kergunteuil, A.; Campos-Herrera, R.; Sánchez-Moreno, S.; Vittoz, P.; Rasmann, S. The Abundance, Diversity, and Metabolic Footprint of Soil Nematodes Is Highest in High Elevation Alpine Grasslands. Front. Ecol. Evol. 2016, 4, 84. [Google Scholar] [CrossRef] [Green Version]
- Oostenbrink, M. Estimating nematode populations by some selected methods. In Nematology; University of North Carolina Press: Chapel Hill, NC, USA, 1960; pp. 85–102. [Google Scholar]
- Bezooijen, J. van Methods and Techniques for Nematology; Wageningen University: Wageningen, The Netherlands, 2006. [Google Scholar]
- Denton, C.S.; Bardgett, R.D.; Cook, R.; Hobbs, P.J. Low Amounts of Root Herbivory Positively Influence the Rhizosphere Microbial Community in a Temperate Grassland Soil. Soil Biol. Biochem. 1998, 31, 155–165. [Google Scholar] [CrossRef]
- Sun, F.; Pan, K.; Tariq, A.; Zhang, L.; Sun, X.; Li, Z.; Wang, S.; Xiong, Q.; Song, D.; Olatunji, O.A. The Response of the Soil Microbial Food Web to Extreme Rainfall under Different Plant Systems. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Scherber, C.; Eisenhauer, N.; Weisser, W.W.; Schmid, B.; Voigt, W.; Fischer, M.; Schulze, E.-D.; Roscher, C.; Weigelt, A.; Allan, E.; et al. Bottom-up Effects of Plant Diversity on Multitrophic Interactions in a Biodiversity Experiment. Nature 2010, 468, 553–556. [Google Scholar] [CrossRef]
- Wang, C.; Bruening, G.; Williamson, V.M. Determination of Preferred PH for Root-Knot Nematode Aggregation Using Pluronic F-127 Gel. J. Chem. Ecol. 2009, 35, 1242–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matute, M.M. Soil Nematodes of Brassica Rapa: Influence of Temperature and PH. Adv. Nat. Sci. 2013, 6, 20–26. [Google Scholar] [CrossRef]
- Wei, C.; Zheng, H.; Li, Q.; Lü, X.; Yu, Q.; Zhang, H.; Chen, Q.; He, N.; Kardol, P.; Liang, W.; et al. Nitrogen Addition Regulates Soil Nematode Community Composition through Ammonium Suppression. PLOS ONE 2012, 7, e43384. [Google Scholar] [CrossRef] [Green Version]
- Bongers, T. The Maturity Index: An Ecological Measure of Environmental Disturbance Based on Nematode Species Composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef]
- Bongers, T.; Ferris, H. Nematode Community Structure as a Bioindicator in Environmental Monitoring. Trends Ecol. Evol. 1999, 14, 224–228. [Google Scholar] [CrossRef]
- Neher, D.; Weicht, T. Nematode Genera in Forest Soil Respond Differentially to Elevated CO2. J. Nematol. 2013, 45, 214–222. [Google Scholar]
- Landsman, A.P.; Ladin, Z.S.; Gardner, D.; Bowman, J.L.; Shriver, G.; D’Amico, V.; Delaney, D.A. Local Landscapes and Microhabitat Characteristics Are Important Determinants of Urban–Suburban Forest Bee Communities. Ecosphere 2019, 10, e02908. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-G.; Zhou, X.-B.; Zhang, Y.-M. Moss Patch Size and Microhabitats Influence Stoichiometry of Moss Crusts in a Temperate Desert, Central Asia. Plant. Soil. 2019, 443, 55–72. [Google Scholar] [CrossRef]
- Devetter, M.; Háněl, L.; Řeháková, K.; Doležal, J. Diversity and Feeding Strategies of Soil Microfauna along Elevation Gradients in Himalayan Cold Deserts. PLOS ONE 2017, 12, e0187646. [Google Scholar] [CrossRef] [Green Version]
- Bach, E.M.; Williams, R.J.; Hargreaves, S.K.; Yang, F.; Hofmockel, K.S. Greatest Soil Microbial Diversity Found in Micro-Habitats. Soil Biol. Biochem. 2018, 118, 217–226. [Google Scholar] [CrossRef]
- Wang, Y.; Niu, K. Effect of soil environment on functional diversity of soil nematodes in Tibetan alpine meadows. Biodivers. Sci. 2020, 28, 707–717. [Google Scholar] [CrossRef]
- Flynn, D.F.B.; Mirotchnick, N.; Jain, M.; Palmer, M.I.; Naeem, S. Functional and Phylogenetic Diversity as Predictors of Biodiversity–Ecosystem-Function Relationships. Ecology 2011, 92, 1573–1581. [Google Scholar] [CrossRef] [Green Version]
- Niu, K.; Choler, P.; de Bello, F.; Mirotchnick, N.; Du, G.; Sun, S. Fertilization Decreases Species Diversity but Increases Functional Diversity: A Three-Year Experiment in a Tibetan Alpine Meadow. Agric. Ecosyst. Environ. 2014, 182, 106–112. [Google Scholar] [CrossRef]
- Enquist, B.J.; Norberg, J.; Bonser, S.P.; Violle, C.; Webb, C.T.; Henderson, A.; Sloat, L.L.; Savage, V.M. Chapter Nine-Scaling from Traits to Ecosystems: Developing a General Trait Driver Theory via Integrating Trait-Based and Metabolic Scaling Theories. In Advances in Ecological Research; Pawar, S., Woodward, G., Dell, A.I., Eds.; Trait-Based Ecology-From Structure to Function; Academic Press: Cambridge, MA, USA, 2015; Volume 52, pp. 249–318. [Google Scholar]
- Haimi, J.; Laamanen, J.; Penttinen, R.; Räty, M.; Koponen, S.; Kellomäki, S.; Niemelä, P. Impacts of Elevated CO2 and Temperature on the Soil Fauna of Boreal Forests. Appl. Soil Ecol. 2005, 30, 104–112. [Google Scholar] [CrossRef]
- Darby, B.J.; Neher, D.A.; Housman, D.C.; Belnap, J. Few Apparent Short-Term Effects of Elevated Soil Temperature and Increased Frequency of Summer Precipitation on the Abundance and Taxonomic Diversity of Desert Soil Micro- and Meso-Fauna. Soil Biol. Biochem. 2011, 43, 1474–1481. [Google Scholar] [CrossRef]
- Dequiedt, S.; Saby, N.P.A.; Lelievre, M.; Jolivet, C.; Thioulouse, J.; Toutain, B.; Arrouays, D.; Bispo, A.; Lemanceau, P.; Ranjard, L. Biogeographical Patterns of Soil Molecular Microbial Biomass as Influenced by Soil Characteristics and Management. Glob. Ecol. Biogeogr. 2011, 20, 641–652. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Cesarz, S.; Koller, R.; Worm, K.; Reich, P.B. Global Change Belowground: Impacts of Elevated CO2, Nitrogen, and Summer Drought on Soil Food Webs and Biodiversity. Glob. Chang. Biol. 2011, 18, 435–447. [Google Scholar] [CrossRef]
- Jones, A.R.; Brunsell, N.A. A Scaling Analysis of Soil Moisture–Precipitation Interactions in a Regional Climate Model. Theor. Appl. Climatol. 2009, 98, 221–235. [Google Scholar] [CrossRef]
- Duerinck, H.M.; van der Ent, R.J.; van de Giesen, N.C.; Schoups, G.; Babovic, V.; Yeh, P.J.F. Observed Soil Moisture–Precipitation Feedback in Illinois: A Systematic Analysis over Different Scales. J. Hydrometeorol. 2016, 17, 1645–1660. [Google Scholar] [CrossRef]
- Wang, G.; Li, Y.; Wu, Q.; Wang, Y. Impacts of Permafrost Changes on Alpine Ecosystem in Qinghai-Tibet Plateau. Sci. China Ser. D 2006, 49, 1156–1169. [Google Scholar] [CrossRef]
- Yang, K.; Wu, H.; Qin, J.; Lin, C.; Tang, W.; Chen, Y. Recent Climate Changes over the Tibetan Plateau and Their Impacts on Energy and Water Cycle: A Review. Glob. Planet. Chang. 2014, 112, 79–91. [Google Scholar] [CrossRef]
- Zhou, J.; Wen, J.; Liu, R.; Wang, X.; Xie, Y. Late Spring Soil Moisture Variation over the Tibetan Plateau and Its Influences on the Plateau Summer Monsoon. Int. J. Climatol. 2018, 38, 4597–4609. [Google Scholar] [CrossRef]
- Cottenie, K. Integrating Environmental and Spatial Processes in Ecological Community Dynamics. Ecol. Lett. 2005, 8, 1175–1182. [Google Scholar] [CrossRef]
- Spatial Structure and Soil Properties Shape Local Community Structure of Plant-Parasitic Nematodes in Cultivated Olive Trees in Southern Spain. Agric. Ecosyst. Environ. 2020, 287, 106688. [CrossRef]
- De Goede, R.G.M.; Bongers, T. Nematode Community Structure in Relation to Soil and Vegetation Characteristics. Appl. Soil Ecol. 1994, 1, 29–44. [Google Scholar] [CrossRef]
- Deyn, G.B.; Raaijmakers, C.; Zoomer, H.; Berg, M.; Ruiter, P.; Verhoef, H.; Bezemer, T.M.; Putten, W. Soil Invertebrate Fauna Enhances Grassland Succession and Diversity. Nature 2003, 422, 711–713. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.P.; Freckman, D.W. The Spatial Distribution of Nematode Trophic Groups Across a Cultivated Ecosystem. Ecology 1995, 76, 1425–1432. [Google Scholar] [CrossRef] [Green Version]
- Rossi, J.-P.; Queneherve, P. Relating Species Density to Environmental Variables in Presence of Spatial Autocorrelation: A Study Case on Soil Nematodes Distribution. Ecography 1998, 21, 117–123. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Meadows | Steppes | p-Value | |
|---|---|---|---|
| Climate | |||
| MAT | −0.7 (3.4) | 0.5 (4.6) | 0.605 |
| MAP | 493.8 (226.6) | 376.6 (227.3) | 0.001 |
| Plants | |||
| PSR | 16.0 (10.0) | 10.0 (6.0) | 0.001 |
| PCV | 85.0 (35.0) | 30.0 (23.0) | 0.001 |
| PBM | 357.5 (151.7) | 46.00 (65.6) | 0.001 |
| Soil properties | |||
| pH | 7.2 (1.9) | 8.2 (0.5) | 0.001 |
| MOI | 23.9 (29.9) | 8.0 (8.3) | 0.001 |
| TN | 3.7 (3.4) | 1.0 (1.2) | 0.001 |
| TP | 0.7 (0.4) | 0.4 (0.2) | 0.001 |
| AP | 1.3 (1.3) | 0.4 (0.4) | 0.001 |
| NH4 | 19.3 (11.2) | 10.0 (2.9) | 0.001 |
| NO3 | 6.6 (9.2) | 3.8 (2.9) | 0.001 |
| SOC | 51.7 (42.3) | 9.7 (7.8) | 0.001 |
| Alpine Meadow | Alpine Steppe | |||
|---|---|---|---|---|
| Soil (%) | Plant (%) | Soil (%) | Plant (%) | |
| Abundance (individuals/100 g of dried soil) | 100.00 | 0 | 92.19 | 7.81 |
| Genus richness (genus) | 100.00 | 0 | 43.99 | 56.01 |
| Plant parasites (%) | 100.00 | 0 | 100.00 | |
| Fungivores (%) | 43.77 | 56.23 | ||
| Bacterivores (%) | 48.36 | 51.64 | 33.07 | 66.93 |
| Predators (%) | 95.73 | 4.27 | ||
| Omnivores (%) | 73.24 | 26.76 | 59.41 | 40.59 |
| EI | 92.97 | 7.03 | 100.00 | 0 |
| SI | 78.70 | 12.30 | 47.84 | 52.16 |
| CI | 100.00 | 0 | 68.37 | 31.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Luo, S.; Li, G.; Jiang, W.; Qi, W.; Hu, J.; Ma, M.; Du, G. Large-Scale Patterns of Soil Nematodes across Grasslands on the Tibetan Plateau: Relationships with Climate, Soil and Plants. Diversity 2021, 13, 369. https://doi.org/10.3390/d13080369
Chen H, Luo S, Li G, Jiang W, Qi W, Hu J, Ma M, Du G. Large-Scale Patterns of Soil Nematodes across Grasslands on the Tibetan Plateau: Relationships with Climate, Soil and Plants. Diversity. 2021; 13(8):369. https://doi.org/10.3390/d13080369
Chicago/Turabian StyleChen, Han, Shuaiwei Luo, Guixin Li, Wanyanhan Jiang, Wei Qi, Jing Hu, Miaojun Ma, and Guozhen Du. 2021. "Large-Scale Patterns of Soil Nematodes across Grasslands on the Tibetan Plateau: Relationships with Climate, Soil and Plants" Diversity 13, no. 8: 369. https://doi.org/10.3390/d13080369
APA StyleChen, H., Luo, S., Li, G., Jiang, W., Qi, W., Hu, J., Ma, M., & Du, G. (2021). Large-Scale Patterns of Soil Nematodes across Grasslands on the Tibetan Plateau: Relationships with Climate, Soil and Plants. Diversity, 13(8), 369. https://doi.org/10.3390/d13080369