
Seagrass Meadows Provide a Significant Resource in Support of Avifauna



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
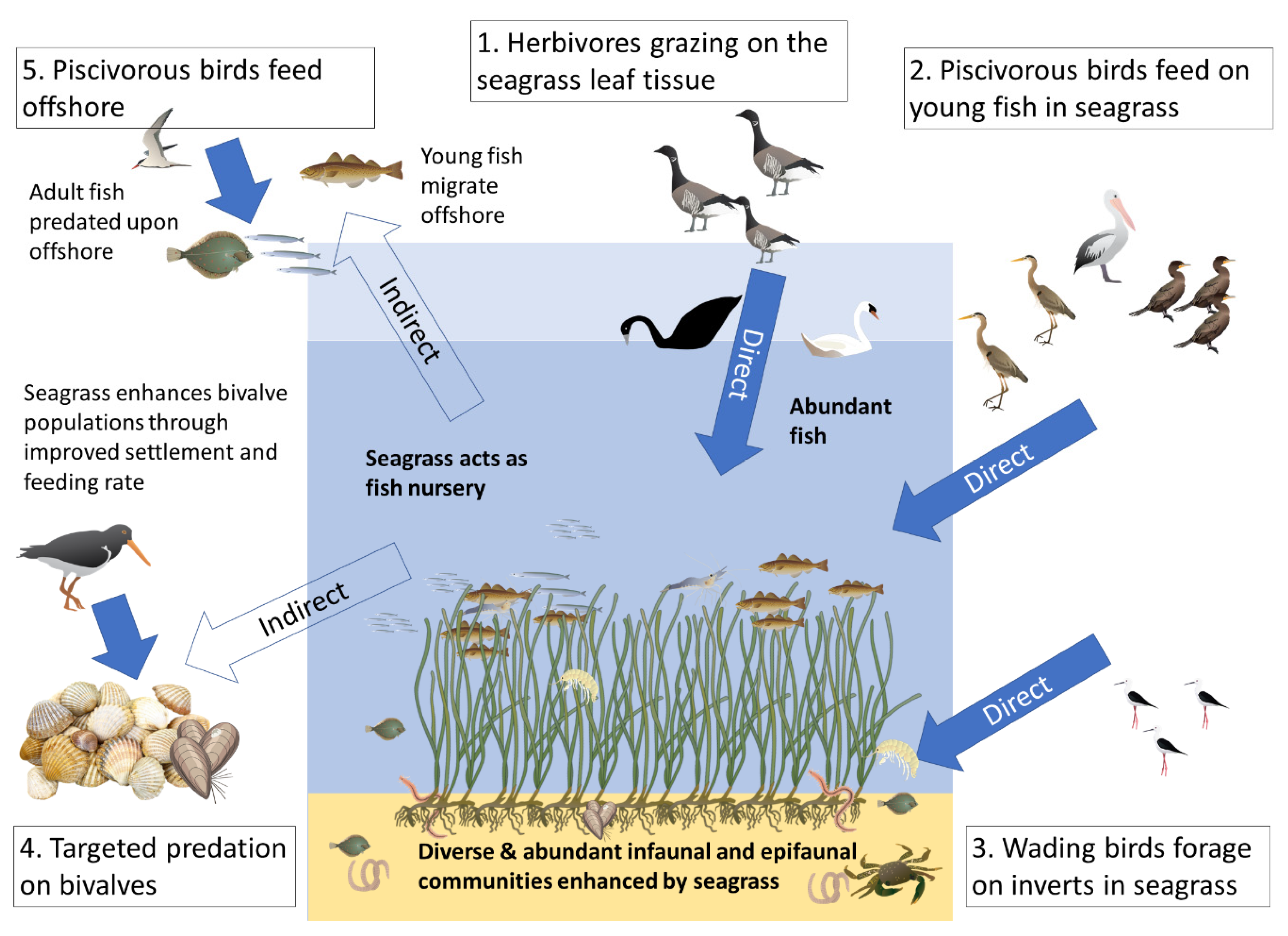
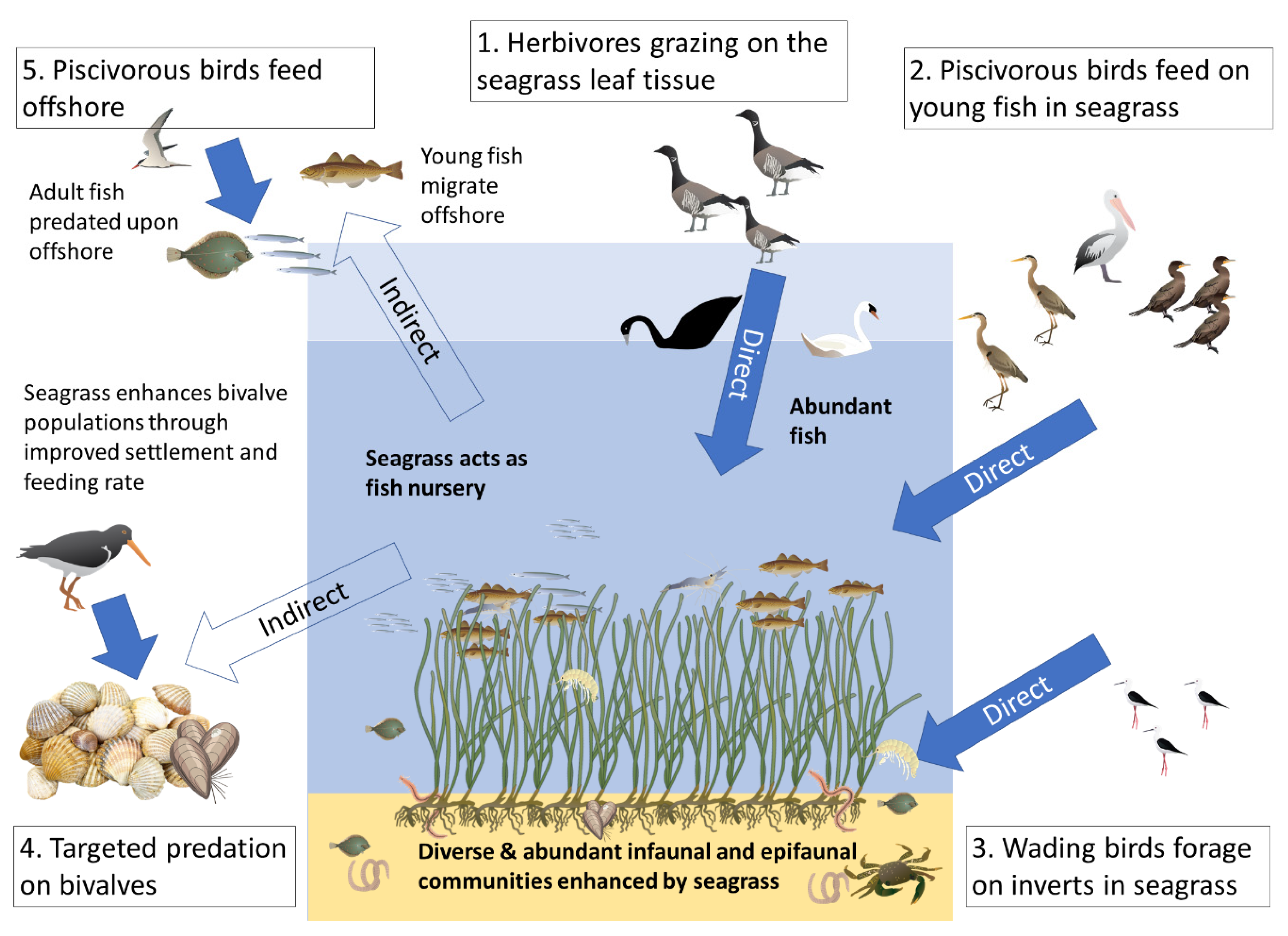
2. Direct Support: Grazing Birds
3. Direct Support: Coastal Birds Foraging on Seagrass Invertebrates
4. Direct Support: Piscivorous Bird Foraging within Seagrass
5. Indirect Support: Seabirds Benefitting from Seagrass Fish Nursery Function
6. Seagrass Loss and Shifting Baselines
7. Seabirds Support Seagrass
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McKenzie, L.; Nordlund, L.M.; Jones, B.L.; Cullen-Unsworth, L.C.; Roelfsema, C.M.; Unsworth, R.K.F. The global distribution of seagrass meadows. Environ. Res. Lett. 2020, 15, 074041. [Google Scholar] [CrossRef]
- Heck, K.L.; Carruthers, T.J.B.; Duarte, C.M.; Hughes, A.R.; Kendrick, G.; Orth, R.; Williams, S.W. Trophic Transfers from Seagrass Meadows Subsidize Diverse Marine and Terrestrial Consumers. Ecosystems 2008, 11, 1198–1210. [Google Scholar] [CrossRef]
- Nordlund, L.M.; Koch, E.W.; Barbier, E.B.; Creed, J. Seagrass Ecosystem Services and Their Variability across Genera and Geographical Regions. PLoS ONE 2016, 11, e0163091. [Google Scholar] [CrossRef] [Green Version]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunic, J.C.; Brown, C.J.; Connolly, R.M.; Turschwell, M.P.; Côté, I.M. Long-term declines and recovery of meadow area across the world’s seagrass bioregions. Glob. Chang. Biol. 2021, 27, 4096–4109. [Google Scholar] [CrossRef] [PubMed]
- Röhr, M.E.; Holmer, M.; Baum, J.K.; Björk, M.; Boyer, K.; Chin, D.; Chalifour, L.; Cimon, S.; Cusson, M.; Dahl, M.; et al. Blue Carbon Storage Capacity of Temperate Eelgrass (Zostera marina) Meadows. Glob. Biogeochem. Cycles 2018, 32, 1457–1475. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, P.S.; Eklöf, J.; van Katwijk, M.; O’Brien, K.; De La Torre-Castro, M.; Boström, C.; Bouma, T.J.; Krause-Jensen, D.; Unsworth, R.; van Tussenbroek, B.I.; et al. The fundamental role of ecological feedback mechanisms for the adaptive management of seagrass ecosystems—A review. Biol. Rev. 2016, 92, 1521–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayraktarov, E.; Saunders, M.I.; Abdullah, S.; Mills, M.; Beher, J.; Possingham, H.; Mumby, P.J.; Lovelock, C. The cost and feasibility of marine coastal restoration. Ecol. Appl. 2016, 26, 1055–1074. [Google Scholar] [CrossRef]
- Green, A.J.; Elmberg, J. Ecosystem services provided by waterbirds. Biol. Rev. 2013, 89, 105–122. [Google Scholar] [CrossRef] [Green Version]
- Kollars, N.M.; Henry, A.K.; Whalen, M.A.; Boyer, K.; Cusson, M.; Eklöf, J.S.; Hereu, C.M.; Jorgensen, P.; Kiriakopolos, S.L.; Reynolds, P.L.; et al. Meta-Analysis of Reciprocal Linkages between Temperate Seagrasses and Waterfowl with Implications for Conservation. Front. Plant Sci. 2017, 8, 2119. [Google Scholar] [CrossRef]
- Cury, P.M.; Boyd, I.L.; Bonhommeau, S.; Anker-Nilssen, T.; Crawford, R.J.M.; Furness, R.W.; Mills, J.A.; Murphy, E.J.; Österblom, H.; Paleczny, M.; et al. Global Seabird Response to Forage Fish Depletion—One-Third for the Birds. Science 2011, 334, 1703–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouwehand, J.; Leopold, M.F.; Camphuysen, K. A comparative study of the diet of Guillemots Uria aalge and Razorbills Alca torda killed during the Tricolor oil incident in the south-eastern North Sea in January 2003. Atl. Seab. 2004, 6, 147–164. [Google Scholar]
- Percival, S.; Sutherland, W.; Evans, P. Intertidal habitat loss and wildfowl numbers: Applications of a spatial depletion model. J. Appl. Ecol. 1998, 35, 57–63. [Google Scholar] [CrossRef]
- Frederiksen, M.; Edwards, M.; Richardson, A.; Halliday, N.C.; Wanless, S. From plankton to top predators: Bottom-up control of a marine food web across four trophic levels. J. Anim. Ecol. 2006, 75, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Valentine, J.; Heck, K. Seagrass herbivory:evidence for the continued grazing of marine grasses. Mar. Ecol. Prog. Ser. 1999, 176, 291–302. [Google Scholar] [CrossRef]
- Attrill, M.J.; Strong, J.A.; Rowden, A.A. Are macroinvertebrate communities influenced by seagrass structural complexity? Ecography 2000, 23, 114–121. [Google Scholar] [CrossRef]
- Lee, S.Y.; Fong, C.; Wu, R. The effects of seagrass (Zostera japonica) canopy structure on associated fauna: A study using artificial seagrass units and sampling of natural beds. J. Exp. Mar. Biol. Ecol. 2001, 259, 23–50. [Google Scholar] [CrossRef]
- Barnes, R. Patterns of benthic invertebrate biodiversity in intertidal seagrass in Moreton Bay, Queensland. Reg. Stud. Mar. Sci. 2017, 15, 17–25. [Google Scholar] [CrossRef]
- Reynolds, P.L.; Stachowicz, J.J.; Hovel, K.; Boström, C.; Boyer, K.; Cusson, M.; Eklöf, J.S.; Engel, F.G.; Engelen, A.H.; Eriksson, B.K.; et al. Latitude, temperature, and habitat complexity predict predation pressure in eelgrass beds across the Northern Hemisphere. Ecology 2018, 99, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Boere, G.C.; Piersma, T. Flyway protection and the predicament of our migrant birds: A critical look at international conservation policies and the Dutch Wadden Sea. Ocean Coast. Manag. 2012, 68, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Horn, S.; Schwemmer, P.; Mercker, M.; Enners, L.; Asmus, R.; Garthe, S.; Asmus, H. Species composition of foraging birds in association with benthic fauna in four intertidal habitats of the Wadden Sea. Estuar. Coast. Shelf Sci. 2020, 233, 106537. [Google Scholar] [CrossRef]
- Harrison, P.G.; Dunn, M. Fraser River delta seagrass ecosystems, their distributions and importance to migratory birds. In Fraser River Delta, British Columbia: Issues of an Urban Estuary; Groulx, R.I., Mosher, D.C., Luternauer, I., Bilderback, D.E., Eds.; Geological Survey of Canada, Bulletin 567: Ottawa, ON, Canada, 2004; pp. 173–188. [Google Scholar]
- Sutherland, T.F.; Shepherd, P.C.F.; Elner, R.W. Predation on meiofaunal and macrofaunal invertebrates by western sandpipers (Calidris mauri): Evidence for dual foraging modes. Mar. Biol. 2000, 137, 983–993. [Google Scholar] [CrossRef]
- Spruzen, F.L.; Richardson, A.M.; Woehler, E.J. Influence of environmental and prey variables on low tide shorebird habitat use within the Robbins Passage wetlands, Northwest Tasmania. Estuar. Coast. Shelf Sci. 2008, 78, 122–134. [Google Scholar] [CrossRef]
- Seymour, N.R.; Miller, A.G.; Garbary, D.J. Decline of Canada geese (Branta canadensis) and common goldeneye (Bucephala clangula) associated with a collapse of eelgrass (Zostera marina) in a Nova Scotia estuary. Helgol. Mar. Res. 2002, 56, 198–202. [Google Scholar] [CrossRef]
- Eadie, J.M.; Mallory, M.L.; Lumsden, H.G. Common Goldeneye (Bucephala clangula), version 2.0. In The Birds of North America; Rodewald, P.G., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 1995. [Google Scholar]
- Pehrsson, O. Food and Feeding Grounds of the Goldeneye Bucephala clangula (L.) on the Swedish West Coast. Ornis Scand. 1976, 7, 91. [Google Scholar] [CrossRef]
- Brun, F.G.; Van Zetten, E.; Cacabelos, E.; Bouma, T.J. Role of two contrasting ecosystem engineers (Zostera noltii and Cymodocea nodosa) on the food intake rate of Cerastoderma edule. Helgol. Mar. Res. 2008, 63, 19–25. [Google Scholar] [CrossRef]
- Norris, K.; Bannister, R.; Walker, P.; Walker, W.D. Changes in the number of oystercatchers Haematopus ostralegus wintering in the Burry Inlet in relation to the biomass of cockles Cerastoderma edule and its commercial exploitation. J. Appl. Ecol. 1998, 35, 75–85. [Google Scholar] [CrossRef]
- Bologna, P.A.; Fetzer, M.L.; McDonnell, S.; Moody, E.M. Assessing the potential benthic–pelagic coupling in episodic blue mussel (Mytilus edulis) settlement events within eelgrass (Zostera marina) communities. J. Exp. Mar. Biol. Ecol. 2005, 316, 117–131. [Google Scholar] [CrossRef]
- Caldow, R.W.G.; Beadman, H.A.; McGrorty, M.J.K.S.; Goss-Custard, J.D.; Mould, K.; Wilson, A. Effects of intertidal mussel cultivation on bird assemblages. Mar. Ecol. Prog. Ser. 2003, 259, 173–183. [Google Scholar] [CrossRef]
- Spruzen, F.L.; Richardson, A.M.M.; Woehler, E.J. Spatial variation of intertidal macroinvertebrates and environmental variables in Robbins Passage wetlands, NW Tasmania. Hydrobiologia 2007, 598, 325–342. [Google Scholar] [CrossRef]
- Lilley, R.J.; Unsworth, R.K. Atlantic Cod (Gadus morhua) benefits from the availability of seagrass (Zostera marina) nursery habitat. Glob. Ecol. Conserv. 2014, 2, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Unsworth, R.K.; Nordlund, L.M.; Cullen-Unsworth, L.C. Seagrass meadows support global fisheries production. Conserv. Lett. 2018, 12, e12566. [Google Scholar] [CrossRef]
- Campbell, K.J.; Steinfurth, A.; Underhill, L.G.; Coetzee, J.; Dyer, B.M.; Ludynia, K.; Makhado, A.B.; Merkle, D.; Rademan, J.; Upfold, L.; et al. Local forage fish abundance influences foraging effort and offspring condition in an endangered marine predator. J. Appl. Ecol. 2019, 56, 1751–1760. [Google Scholar] [CrossRef]
- Bertelli, C.M.; Unsworth, R.K. Protecting the hand that feeds us: Seagrass (Zostera marina) serves as commercial juvenile fish habitat. Mar. Pollut. Bull. 2014, 83, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Lefcheck, J.S.; Hughes, B.B.; Johnson, A.; Pfirrmann, B.W.; Rasher, D.B.; Smyth, A.R.; Williams, B.L.; Beck, M.; Orth, R. Are coastal habitats important nurseries? A meta-analysis. Conserv. Lett. 2019, 12, e12645. [Google Scholar] [CrossRef]
- Henderson, C.J.; Gilby, B.L.; Lee, S.Y.; Stevens, T. Contrasting effects of habitat complexity and connectivity on biodiversity in seagrass meadows. Mar. Biol. 2017, 164, 117. [Google Scholar] [CrossRef]
- Butler, R.W. Habitat Selection and Time of Breeding in the Great Blue Heron, (Ardea herodias). University of British Columbia. Available online: https://open.library.ubc.ca/collections/ubctheses/831/items/1.01003921991 (accessed on 1 July 2021).
- Hom, C.W. Foraging Ecology of Herons in a Southern San Francisco Bay Salt Marsh. Colon. Waterbirds 1983, 6, 37. [Google Scholar] [CrossRef]
- Huang, A.C.; Essak, M.; O’Connor, M.I. Top-down control by great blue herons Ardea herodias regulates seagrass-associated epifauna. Oikos 2015, 124, 1492–1501. [Google Scholar] [CrossRef]
- Custer, T.W. The Use of Feeding Habitat by a Colony of Herons, Egrets, and Ibises near Beaufort, North Carolina (Abstract Only). Proc. Colon. Waterbird Group 1978, 1, 154. [Google Scholar] [CrossRef]
- Dorfman, E.J.; Kingsford, M. Environmental determinants of distribution and foraging behaviour of cormorants (Phalacrocorax spp.) in temperate estuarine habitats. Mar. Biol. 2001, 138, 1–10. [Google Scholar] [CrossRef]
- Grémillet, D.; Argentin, G.; Schulte, B.; Culik, B.M. Flexible foraging techniques in breeding Cormorants Phalacrocorax carbo and Shags Phalacrocorax aristotelis: Benthic or pelagic feeding? Ibis 2008, 140, 113–119. [Google Scholar] [CrossRef]
- Heithaus, M.R. Habitat use and group size of pied cormorants (Phalacrocorax varius) in a seagrass ecosystem: Possible effects of food abundance and predation risk. Mar. Biol. 2005, 147, 27–35. [Google Scholar] [CrossRef]
- Redfern, C.P.; Bevan, R.M.; Redfern, C. A comparison of foraging behaviour in the North Sea by Black-legged Kittiwakes Rissa tridactyla from an inland and a maritime colony. Bird Study 2014, 61, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Bugge, J.; Barrett, R.T.; Pedersen, T. Optimal foraging in chick-raising Common Guillemots (Uria aalge). J. Ornithol. 2011, 152, 253–259. [Google Scholar] [CrossRef] [Green Version]
- McDevitt-Irwin, J.; Iacarella, J.; Baum, J. Reassessing the nursery role of seagrass habitats from temperate to tropical regions: A meta-analysis. Mar. Ecol. Prog. Ser. 2016, 557, 133–143. [Google Scholar] [CrossRef]
- Furness, E.; Unsworth, R.K. Demersal Fish Assemblages in NE Atlantic Seagrass and Kelp. Diversity 2020, 12, 366. [Google Scholar] [CrossRef]
- Kritzer, J.P.; DeLucia, M.-B.; Greene, E.; Shumway, C.; Topolski, M.F.; Thomas-Blate, J.; Chiarella, L.A.; Davy, K.B.; Smith, K. The Importance of Benthic Habitats for Coastal Fisheries. BioScience 2016, 66, 274–284. [Google Scholar] [CrossRef]
- Seitz, R.D.; Wennhage, H.; Bergström, U.; Lipcius, R.N.; Ysebaert, T. Ecological value of coastal habitats for commercially and ecologically important species. ICES J. Mar. Sci. J. Cons. 2013, 71, 648–665. [Google Scholar] [CrossRef] [Green Version]
- Einoder, L. A review of the use of seabirds as indicators in fisheries and ecosystem management. Fish. Res. 2009, 95, 6–13. [Google Scholar] [CrossRef]
- Polte, P.; Asmus, H. Influence of seagrass beds (Zostera noltii) on the species composition of juvenile fishes temporarily visiting the intertidal zone of the Wadden Sea. J. Sea Res. 2006, 55, 244–252. [Google Scholar] [CrossRef]
- Rönnbäck, P.; Kautsky, N.; Pihl, L.; Troell, M.; Söderqvist, T.; Wennhage, H. Ecosystem Goods and Services from Swedish Coastal Habitats: Identification, Valuation, and Implications of Ecosystem Shifts. Ambio 2007, 36, 534–544. [Google Scholar] [CrossRef]
- Anderson, H.B.; Evans, P.G.H.; Potts, J.; Harris, M.P.; Wanless, S. The diet of Common Guillemot Uria aalge chicks provides evidence of changing prey communities in the North Sea. Ibis 2013, 156, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Barrett, R.T. The diet, growth and survival of Razorbill Alca torda chicks in the southern Barents Sea. Ornis Nor. 2015, 38, 25. [Google Scholar] [CrossRef] [Green Version]
- Riordan, J.; Birkhead, T. Changes in the diet composition of Common Guillemot Uria aalge chicks on Skomer Island, Wales, between 1973 and 2017. Ibis 2018, 160, 470–474. [Google Scholar] [CrossRef]
- Hamer, K.C.; Humphreys, E.M.; Garthe, S.; Hennicke, J.; Peters, G.; Grémillet, D.; Phillips, R.A.; Harris, M.P.; Wanless, S. Annual variation in diets, feeding locations and foraging behaviour of gannets in the North Sea: Flexibility, consistency and constraint. Mar. Ecol. Prog. Ser. 2007, 338, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Paiva, V.; Geraldes, P.; Ramirez, I.; Meirinho, A.; Garthe, S.; Ramos, J. Foraging plasticity in a pelagic seabird species along a marine productivity gradient. Mar. Ecol. Prog. Ser. 2010, 398, 259–274. [Google Scholar] [CrossRef]
- Pettex, E.; Lorentsen, S.-H.; Grémillet, D.; Gimenez, O.; Barrett, R.T.; Pons, J.-B.; Le Bohec, C.; Bonadonna, F. Multi-scale foraging variability in Northern gannet (Morus bassanus) fuels potential foraging plasticity. Mar. Biol. 2012, 159, 2743–2756. [Google Scholar] [CrossRef]
- Pauly, D. Anecdotes and the shifting baseline syndrome of fisheries. Trends Ecol. Evol. 1995, 10, 430. [Google Scholar] [CrossRef]
- Green, A.E.; Unsworth, R.K.F.; Chadwick, M.A.; Jones, P.J.S. Historical Analysis Exposes Catastrophic Seagrass Loss for the United Kingdom. Front. Plant Sci. 2021, 12, 261. [Google Scholar] [CrossRef]
- Baden, S.; Gullstrom, M.; Lunden, B.; Pihl, L.; Rosenberg, R. Vanishing seagrass (Zostera marina L.) in Swedish coastal waters. Ambio 2003, 32, 374–377. [Google Scholar] [CrossRef]
- Unsworth, R.K.F.; McKenzie, L.J.; Collier, C.J.; Cullen-Unsworth, L.C.; Duarte, C.M.; Eklöf, J.S.; Jarvis, J.C.; Jones, B.L.; Nordlund, L.M. Global challenges for seagrass conservation. Ambio 2019, 48, 801–815. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.; Wanless, S.; Wright, P.J.; Harris, M.P.; Bull, J.; Elston, D.A. Diet and breeding performance of black-legged kittiwakes Rissa tridactyla at a North Sea colony. Mar. Ecol. Prog. Ser. 2001, 221, 277–284. [Google Scholar] [CrossRef]
- Thurstan, R.H.; Hawkins, J.P.; Raby, L.; Roberts, C.M. Oyster (Ostrea edulis) extirpation and ecosystem transformation in the Firth of Forth, Scotland. J. Nat. Conserv. 2013, 21, 253–261. [Google Scholar] [CrossRef]
- Short, F.; Dennison, W.; Capone, D. Phosphorus-limited growth of the tropical seagrass Syringodium filiforme in carbonate sediments. Mar. Ecol. Prog. Ser. 1990, 62, 169–174. [Google Scholar] [CrossRef]
- Atkinson, M.J.; Smith, S.V. C:N:P ratios of benthic marine plants1. Limnol. Oceanogr. 1983, 28, 568–574. [Google Scholar] [CrossRef]
- Kenworthy, W.J.; Hall, M.O.; Hammerstrom, K.K.; Merello, M.; Schwartzschild, A. Restoration of tropical seagrass beds using wild bird fertilization and sediment regrading. Ecol. Eng. 2018, 112, 72–81. [Google Scholar] [CrossRef] [Green Version]
- McMahon, K.M.; Van Dijk, K.-J.; Ruiz-Montoya, L.; Kendrick, G.; Krauss, S.; Waycott, M.; Verduin, J.; Lowe, R.; Statton, J.; Brown, E.; et al. The movement ecology of seagrasses. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140878. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Unsworth, R.K.F.; Butterworth, E.G. Seagrass Meadows Provide a Significant Resource in Support of Avifauna. Diversity 2021, 13, 363. https://doi.org/10.3390/d13080363
Unsworth RKF, Butterworth EG. Seagrass Meadows Provide a Significant Resource in Support of Avifauna. Diversity. 2021; 13(8):363. https://doi.org/10.3390/d13080363
Chicago/Turabian StyleUnsworth, Richard K. F., and Emma G. Butterworth. 2021. "Seagrass Meadows Provide a Significant Resource in Support of Avifauna" Diversity 13, no. 8: 363. https://doi.org/10.3390/d13080363
APA StyleUnsworth, R. K. F., & Butterworth, E. G. (2021). Seagrass Meadows Provide a Significant Resource in Support of Avifauna. Diversity, 13(8), 363. https://doi.org/10.3390/d13080363