
Cultivar-Groups in Cucurbita maxima Duchesne: Diversity and Possible Domestication Pathways


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
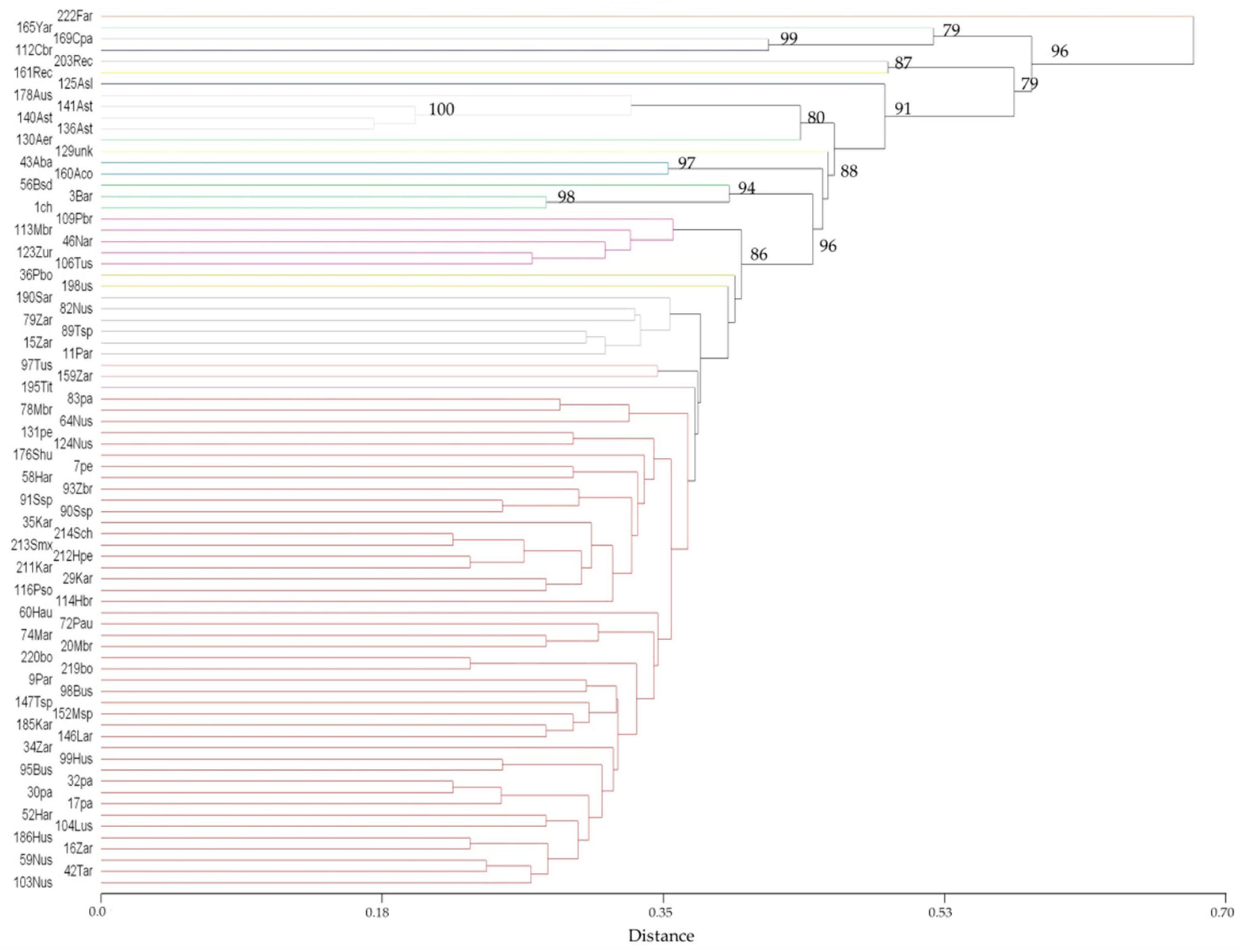{kind=link}
{kind=link}
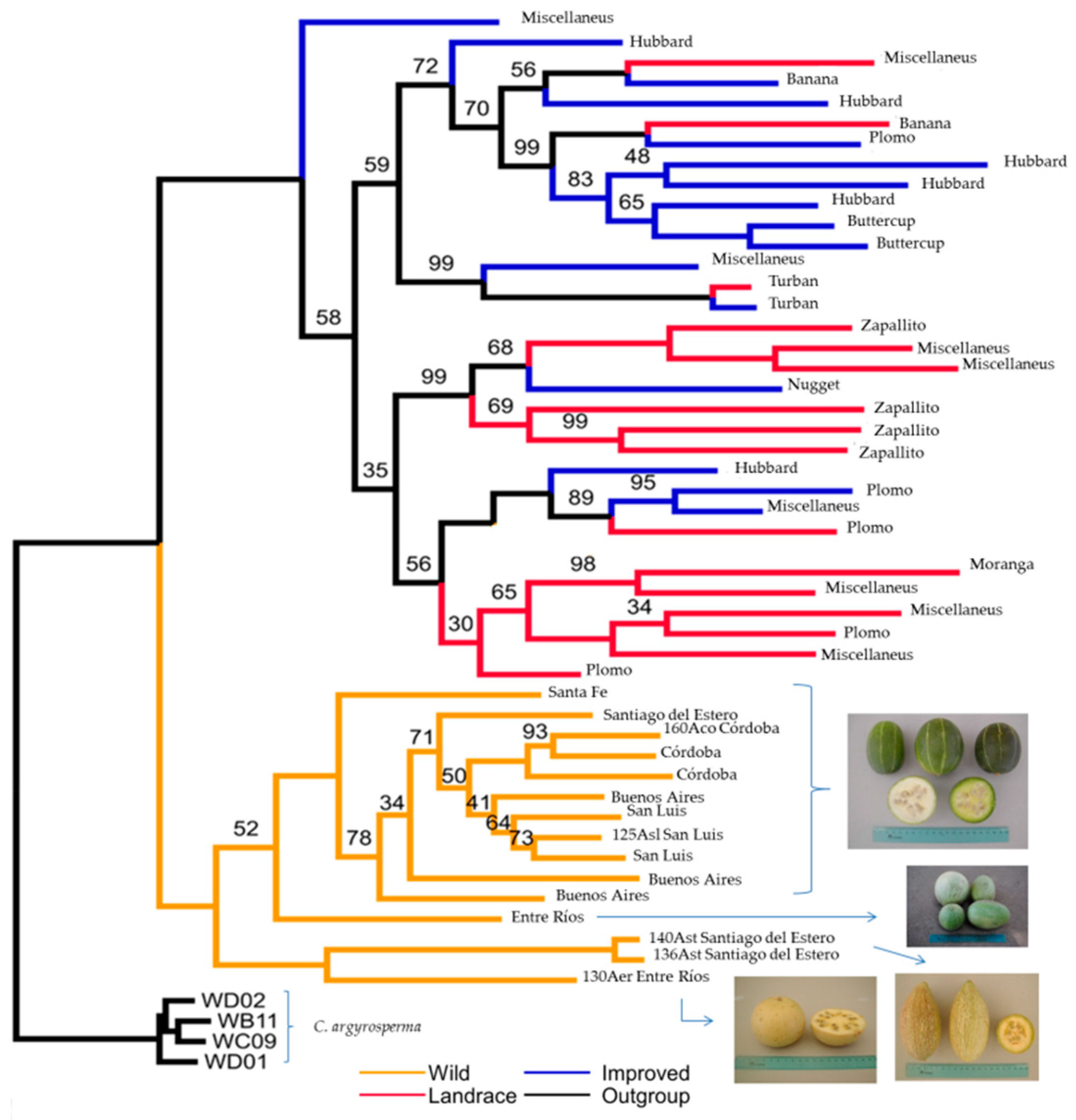{kind=link}
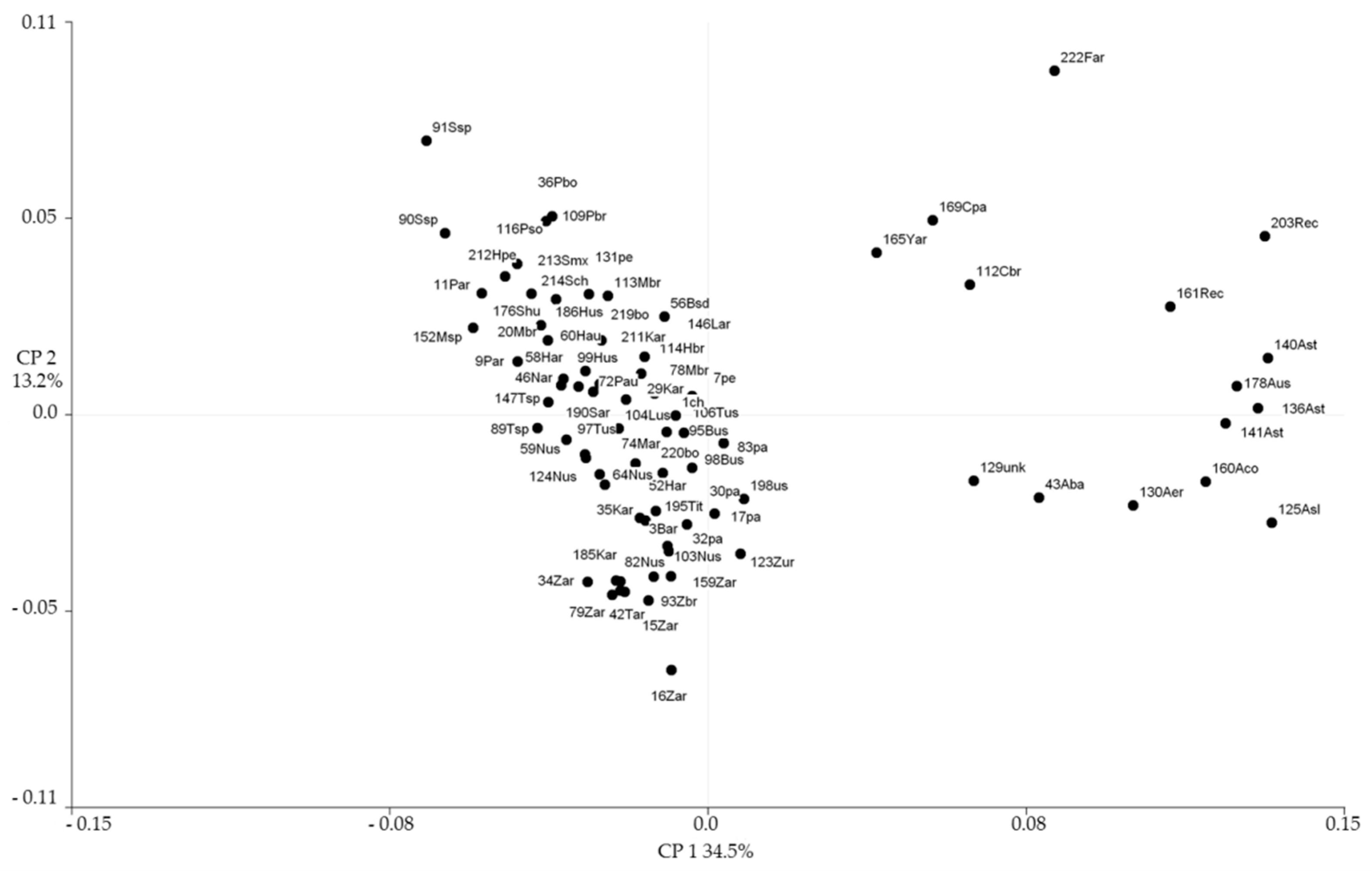{kind=link}
1. Cucurbita
1.1. Extant Wild Species
1.2. Seed Dispersal
1.3. Anthropogenic Seed Dispersal
1.4. Domestication
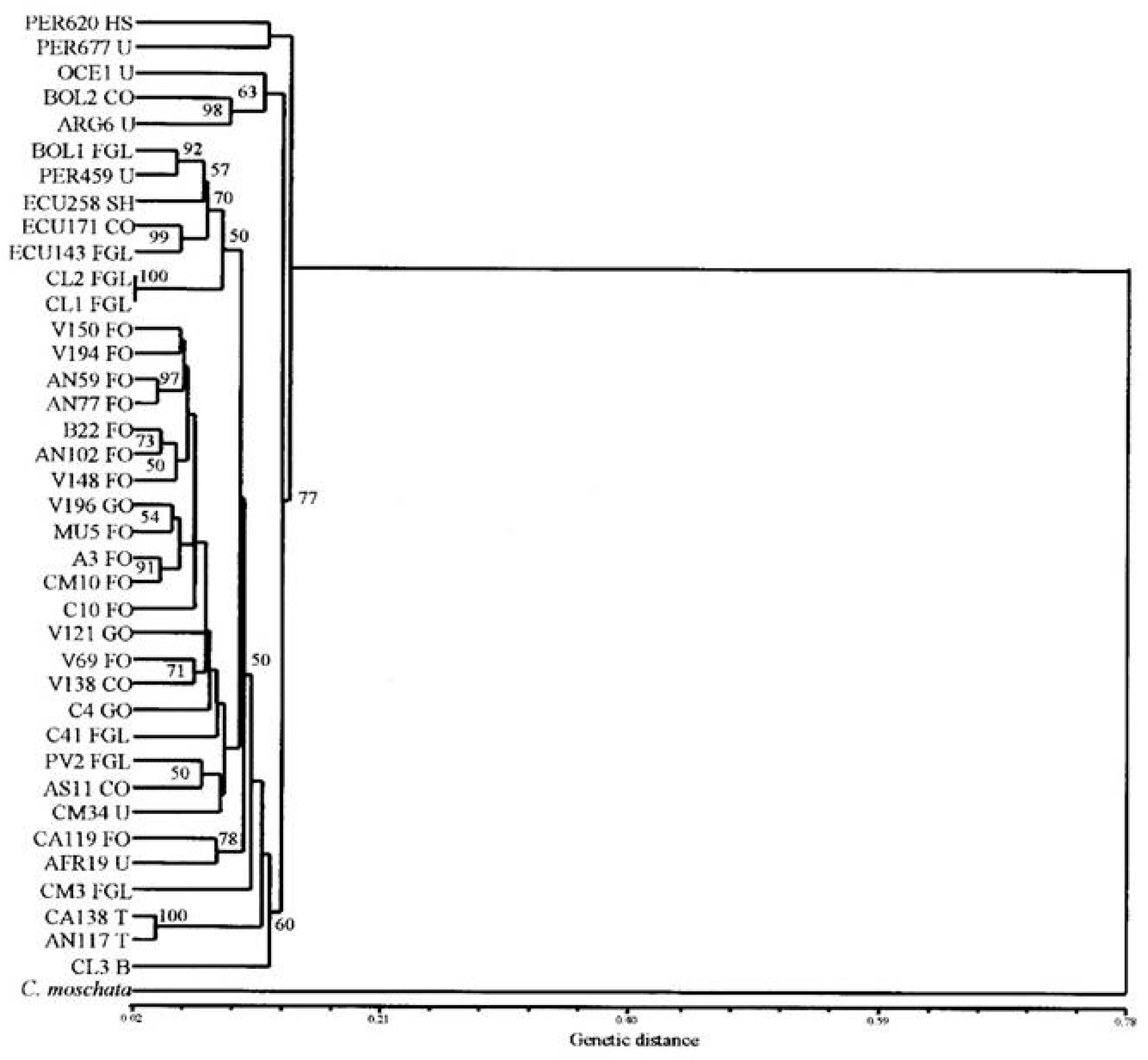
1.5. Cucurbita in South America
1.6. Natural Cucurbita Pollinators in South America
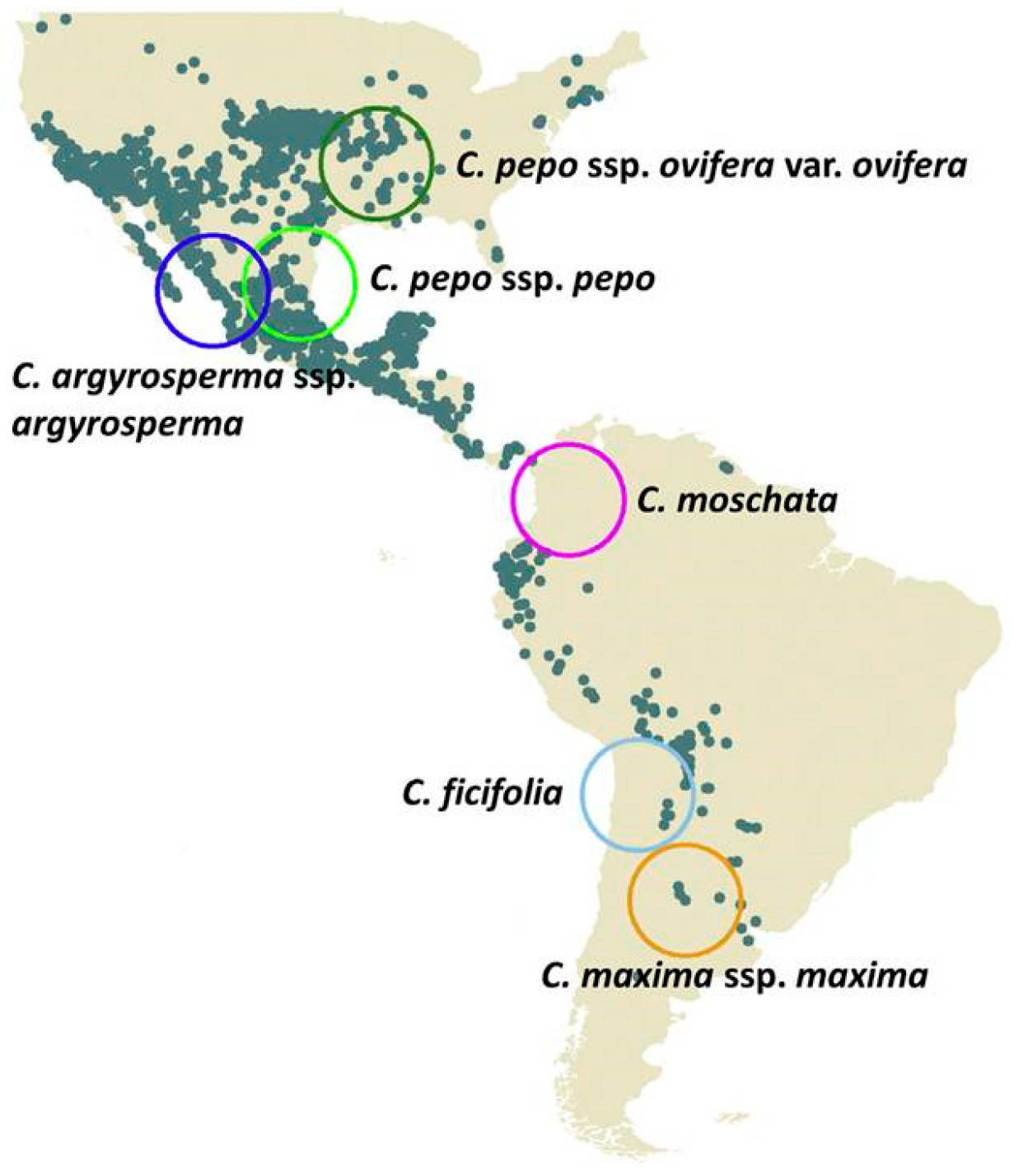
1.7. Domesticated Cucurbita Species
2. Cucurbita Archeobotanics
2.1. South American Archeological Sites
2.2. Diagnostic Macroremains
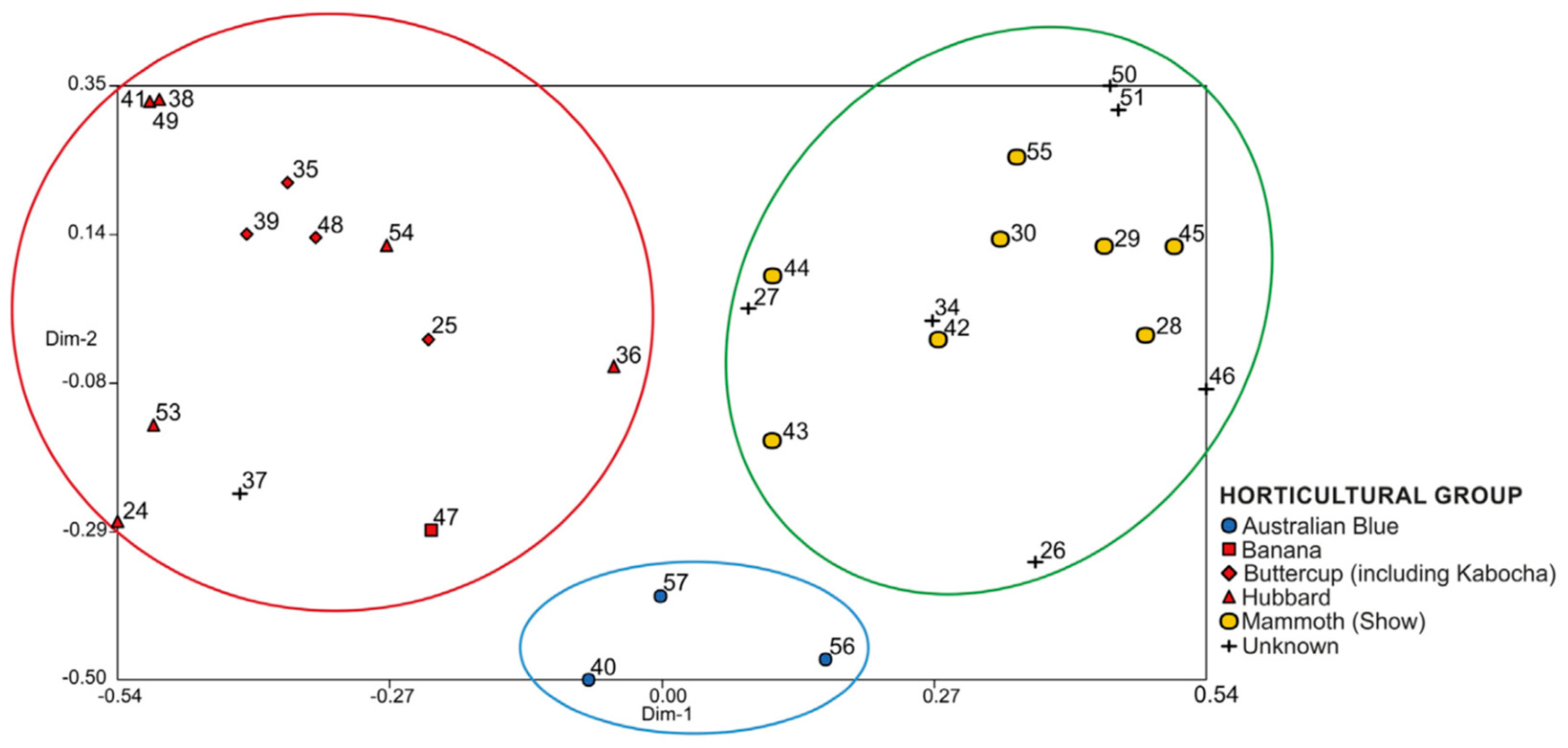
3. Cultivar-Groups
3.1. Hubbard
3.2. Turban
3.3. Banana
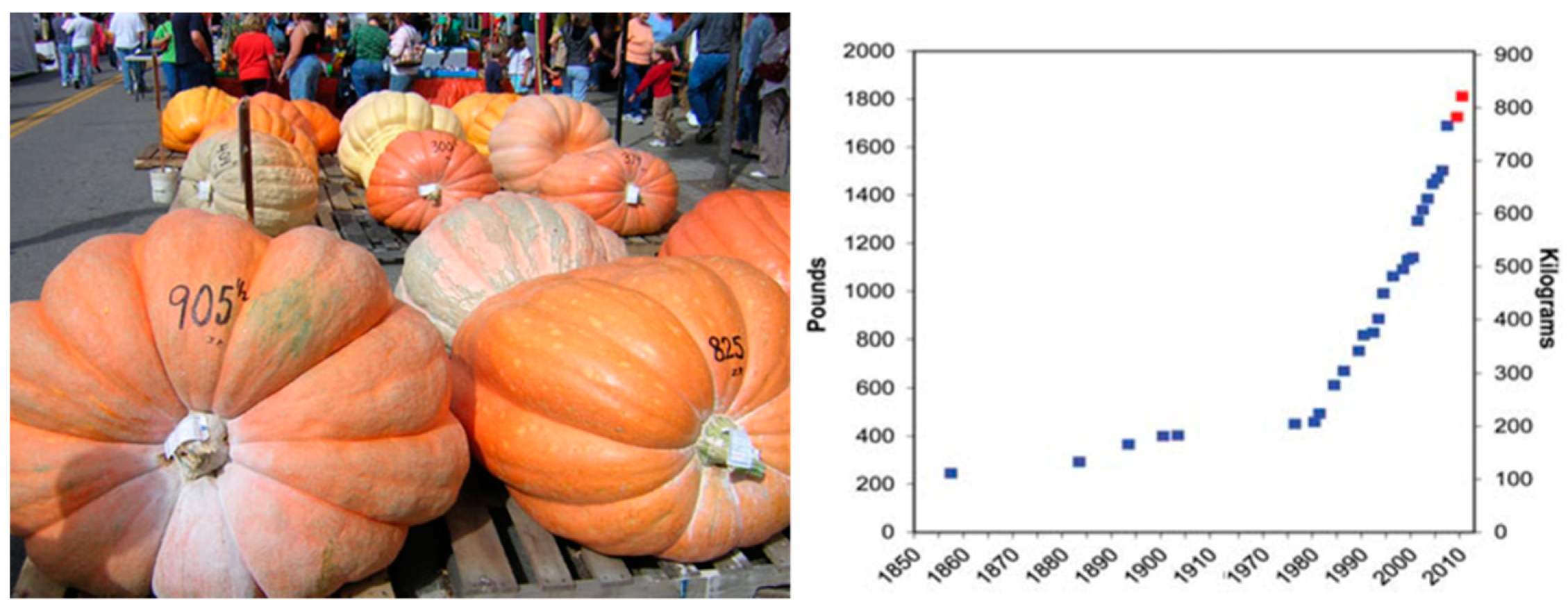
3.4. Show
3.5. Buttercup
3.6. Nugget
3.7. Zapallito
3.8. Plomo
3.9. Moranga
3.10. Zipinka
4. Diversity and Domestication Pathways
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Paris, H.S. Genetic Resources of Pumpkins and Squash, Cucurbita spp. In Genetics and Genomics of Cucurbitaceae. Plant Genetics and Genomics: Crops and Models; Grumet, R., Katzir, N., Garcia-Mas, J., Eds.; Springer: Cham, Switzerland, 2016; Volume 20, pp. 11–154. [Google Scholar] [CrossRef]
- Nee, M. The domestication of Cucurbita (Cucurbitaceae). Econ. Bot. 1990, 44, 56–68. [Google Scholar] [CrossRef]
- Whitaker, T.W.; Cutler, H.C. Cucurbits and cultures in the Americas. Econ. Bot. 1965, 19, 344–349. [Google Scholar] [CrossRef]
- Khoury, C.K.; Carver, D.; Kates, H.R.; Achicanoy, H.A.; van Zonneveld, M.; Thomas, E.; Heinitz, C.; Jarret, R.; Labate, J.; Reitsma, K.; et al. Distributions, conservation status, and abiotic stress tolerance potential of wild cucurbits (Cucurbita L.). Plants People Planet. 2020, 2, 269–283. [Google Scholar] [CrossRef]
- Millán, R. Variaciones el zapallito amargo Cucurbita andreana y el origen de Cucurbita maxima. Rev. Arg. Agron. 1945, 12, 86–93. [Google Scholar]
- Newsom, L.A.; Mihlbachler, M.C. Mastodons (Mammut americanum) Diet Foraging Patterns Based on Analysis of Dung Deposits. In First Floridians and Last Mastodons: The Page-Ladson Site in The Aucilla River; Webb, S.D., Ed.; Springer: Dordrecht, The Netherlands, 2006; pp. 263–332. [Google Scholar]
- Barlow, C. The Ghost of Evolution, Nonsensical Fruit, Missing Partners, and Other Ecological Anachronisms; Basic Books: New York, NY, USA, 2000; pp. 51–69. [Google Scholar]
- Janzen, D.H.; Martin, P.S. Neotropical Anachronisms: The Fruits the Gomphotheres Ate. Science 1982, 215, 19–27. [Google Scholar] [CrossRef]
- Johnson, C.N. Ecological consequences of Late Quaternary extinctions of megafauna. Proc. R. Soc. B 2009, 276, 2509–2519. [Google Scholar] [CrossRef]
- Kistler, L.; Newsom, L.A.; Ryan, T.M.; Clark, A.C.; Smith, B.D.; Perry, G.H. Gourds and squashes (Cucurbita spp.) adapted to megafaunal extinction and ecological anachronism through domestication. Proc. Natl. Acad. Sci. USA 2015, 112, 15107–15112. [Google Scholar] [CrossRef]
- Tropicos Connecting the World to Botanical Data Since 1982. Available online: http://legacy.tropicos.org/Name/50172465?projectid=13 (accessed on 8 July 2021).
- Ashwoth, L.; Galetto, L. Pollinators and Reproductive Success of the Wild Cucurbit Cucurbita maxima ssp. andreana (Cucurbitaceae). Plant. Biol. 2001, 3, 398–404. [Google Scholar] [CrossRef]
- Erickson, D.L.; Smith, B.D.; Clarke, A.C.; Sandweiss, D.H.; Tuross, N. An Asian origin for a 10,000-year-old domesticated plant in the Americas. Proc. Natl. Acad. Sci. USA 2005, 102, 18315–18320. [Google Scholar] [CrossRef] [PubMed]
- Spengler, R.N. Anthropogenic Seed Dispersal: Rethinking the Origins of Plant Domestication. Trends Plant. Sci. 2020, 25, 340–348. [Google Scholar] [CrossRef]
- Perugganan, M.D. Evolutionary Insights into the Nature of Plant Domestication. Curr. Biol. 2019, 29, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Fuller, D.Q.; Allaby, R.G.; Stevens, C. Domestication as innovation: The entanglement of techniques, technology and chance in the domestication of cereal crops. World Archaeol. 2010, 42, 13–28. [Google Scholar] [CrossRef]
- Millán, R. Los zapallitos de tronco de Sudamérica extratropical. Darwiniana 1947, 7, 333–345. [Google Scholar]
- Wu, T.; Zhou, J.; Zhang, Y.; Cao, J. Characterization and inheritance of a bush-type in tropical pumpkin (Cucurbita moschata Duchesne). Sci. Hortic. 2007, 114, 1–4. [Google Scholar] [CrossRef]
- Paris, H.S.; Lebeda, A.; Křistkova, E.; Andres, T.C.; Nee, M.H. Parallel Evolution Under Domestication and Phenotypic Differentiation of the Cultivated Subspecies of Cucurbita pepo (Cucurbitaceae). Econ. Bot. 2012, 66, 71–90. [Google Scholar] [CrossRef]
- Heiser, C.B. Aspects of unconscious selection and the evolution of domesticated plants. Euphytica 1988, 37, 77–81. [Google Scholar] [CrossRef]
- Marshall, L.G. Land Mammals and the Great American Interchange. Am. Sci. 1988, 76, 381–388. [Google Scholar]
- Sánchez, B.; Prado, J.L.; Alberdi, M.T. Feeding ecology, dispersal, and extinction of South American Pleistocene gomphotheres (Gomphotheriidae, Proboscidea). Paleobiology 2004, 30, 146–161. [Google Scholar] [CrossRef]
- Giannini, T.C.; Saraiva, A.M.; Alves-dos-Santos, I. Ecological niche modeling and geographical distribution of pollinator and plants: A case study of Peponapis fervens (Smith, 1879) (Eucerini: Apidae) and Cucurbita species (Cucurbitaceae). Ecol. Inform. 2010, 5, 59–66. [Google Scholar] [CrossRef]
- Ferriol, M.; Picó, B. Pumpkin and Winter Squash. In Handbook of Plant Breeding Vegetables I: Asteraceae, Brassicaceae, Chenopodicaceae, and Cucurbitaceae; Prohens, J., Nuez, F., Eds.; Springer Science Business Media: New York, NY, USA, 2008; pp. 317–349. [Google Scholar]
- Smith, B.D. The initial domestication of Cucurbita pepo in the Americas 10,000 years ago. Science 1997, 276, 932–934. [Google Scholar] [CrossRef]
- Andres, T.C. Biosystematics, theories on the origin, and breeding potential of Cucurbita ficifolia. In Biology and Utilization of the Cucurbitaceae; Bates, D.V., Robinson, R.W., Jeffrey, C., Eds.; Cornell University Press: Ithaca, NY, USA, 1990; pp. 102–119. [Google Scholar]
- Sanjur, O.I.; Piperno, D.R.; Andres, T.C.; Wessel-Beaver, L. Phylogenetic relationships among domesticated and wild species of Cucurbita (Cucurbitaceae) inferred from a mitocondrial gene: Implications for crop plant evolution and areas of origin. Proc. Natl. Acad. Sci. USA 2002, 99, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Decker, D.S. Origin(s), Evolution and Systematics of Cucurbita pepo (Cucurbitaceae). Econ. Bot. 1988, 42, 4–15. [Google Scholar] [CrossRef]
- Barrera-Redondo, J.; Sánchez-de la Vega, J.; Aguirre-Liguori, J.A.; Castellanos-Morales, G.; Gutiérrez-Guerrero, Y.T.; Aguirre-Dugua, X.; Aguirre-Planter, E.; Tenaillon, M.I.; Rafael Lira-Saade, R.; Eguiarte, L.E. The domestication of Cucurbita argyrosperma as revealed by the genome of its wild relative. Hortic. Res. 2021, 8, 109–123. [Google Scholar] [CrossRef]
- Kates, H.R.; Soltis, P.S.; Soltis, D.E. Evolutionary and domestication history of Cucurbita (pumpkin and squash) species inferred from 44 nuclear loci. Mol. Phylogenetics Evol. 2017, 111, 98–109. [Google Scholar] [CrossRef]
- Gong, L.; Paris, H.S.; Nee, M.H.; Stift, G.; Pachner, N.; Vollmann, J.; Lelley, T. Genetic relationships and evolution in Cucurbita pepo (pumpkin, squash, gourd) as revealed by simple sequence repeat polymorphisms. Appl. Genet. 2012, 124, 875–891. [Google Scholar] [CrossRef]
- Castellanos-Morales, G.; Ruiz-Mondragón, K.Y.; Hernández-Rosales, H.S.; Guillermo Sánchez-de la Vega, G.; Gámez, N.; Aguirre-Planter, E.; Montes-Hernández, S.; Lira-Saade, R.; Eguiarte, L.E. Tracing back the origin of pumpkins (Cucurbita pepo ssp. pepo L.) in Mexico. Proc. R. Soc. B 2019, 286, 20191440. [Google Scholar] [CrossRef] [PubMed]
- Andres, T.C. Diversity in tropical pumpkin (Cucurbita moschata): Cultivar origin and history. In Progress in Cucurbit Genetics and Breeding Research, Proceeding of the Cucurbitaceae 2004, Olomouc, Czech Republic, 12–17 July 2004; Lebeda, A., Paris, H.S., Eds.; Palacky University: Olomouc, Czech Republic, 2004; pp. 113–119. [Google Scholar]
- Whitaker, T.W. Cucurbits in Andean prehistory. Am. Antiq. 1983, 48, 576–585. [Google Scholar] [CrossRef]
- Lema, V. Domesticación Vegetal y Grado de Dependencia ser Humano-Planta en el Desarrollo Cultural Prehispánico del Noroeste Argentino. Ph.D. Thesis, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata, Argentina, 2009. [Google Scholar]
- Parodi, L.R. La Agricultura Aborigen Argentina; Universitaria de Buenos Aires: Buenos Aires, Argentina, 1966; p. 45. [Google Scholar]
- Paucke, F. Hacia Allá y para Acá, 1st ed.; Ministerio de Innovación y Cultura: Santa Fe, Argentina, 2010; p. 168.
- Watling, J.; Shock, M.P.; Mongelo, G.Z.; Almeida, F.O.; Kater, T.; De Oliveira, P.E.; Neves, E.G. Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLoS ONE 2018, 13, 1–28. [Google Scholar] [CrossRef]
- Bonomo, M.; Politis, G.G. Mound Building, Social Complexity and Horticulture in the Lower Paraná River. In Encyclopedia of Global Archaeology; Smith, C., Ed.; Springer International Publishing AG: Cham, Switzerland, 2018; pp. 1–22. [Google Scholar] [CrossRef]
- Kates, H.R.; López-Anido, F.; Sánchez-De La Vega, G.; Eguiarte, L.E.; Soltis, P.; Soltis, D.E. Targeted sequencing suggests wild-crop gene flow is central to different genetic consequences of two independent pumpkin domestications. Front. Ecol. Evol. 2021, 9, 405, in press. [Google Scholar] [CrossRef]
- Pearsall, D.M. Plant Domestication and the Shift to Agriculture in the Andes. In Handbook of South American Archaeology; Silverman, H., Isbell, W.H., Eds.; Springer: New York, NY, USA, 2008; pp. 105–120. [Google Scholar] [CrossRef]
- Dillehay, T.D.; Rossen, J.; Andres, T.C.; Williams, D.E. Preceramic Adoption of Peanut, Squash, and Cotton in Northern Peru. Science 2007, 316, 1890–1893. [Google Scholar] [CrossRef]
- Clement, C.R. 1492 and the Loss of Amazonian Crop Genetic Resources. I. The relation between Domestication and Human Population Decline. Econ. Bot. 1999, 53, 188–202. [Google Scholar] [CrossRef]
- Clement, C.R. 1492 and the Loss of Amazonian Crop Genetic Resources. II. Crop Biogeography at Contact. Econ. Bot. 1999, 53, 203–216. [Google Scholar] [CrossRef]
- Piperno, D.R.; Andres, T.C.; Stothert, K.E. Phytoliths in Cucurbita and other Neotropical Cucurbitaceae and their Occurrence in Early Archaeological Sites from the Lowland American Tropics. J. Archaeol. Sci. 2000, 27, 193–208. [Google Scholar] [CrossRef]
- Lema, V.S. Non-domesticated cultivation in the Andes: Plant management and nurturing in the Argentine northwest. Veg. Hist. Archaeobot. 2015, 24, 143–150. [Google Scholar] [CrossRef]
- Zeder, M.A.; Emshwille, E.; Smith, B.D.; Bradley, D.G. Documenting domestication: The intersection of genetics and archaeology. TRENDS Genet. 2006, 22, 139–156. [Google Scholar] [CrossRef]
- Martínez, A.; Lema, V.; Capparelli, A.; Bartoli, C.; López-Anido, F.; Perez, S. Multidisciplinary studies in Cucurbita maxima (squash) domestication. Veg. Hist. Archaeobot. 2018, 27, 207–217. [Google Scholar] [CrossRef]
- The Cucurbit Genetics Cooperative (CGC) Gen List of Cucurbita Species. Available online: https://cucurbit.info/wp-content/uploads/2018/10/gene14squash.pdf (accessed on 9 July 2021).
- Brickell, C.D.; Baum, B.R.; Hetterscheid, W.L.A.; Leslie, A.C.; McNeill, J.; Trehane, P.; Vrugtman, F.; Wiersema, J.H. International Code of Nomenclature for Cultivated Plants. Acta Hortic. 2004, 647, 17–19. [Google Scholar] [CrossRef]
- Paris, H.S. A proposed subspecific classification of Cucurbita pepo. Phytologia 1986, 61, 133–138. [Google Scholar]
- Paris, H.S. Historical records, origin, and development of the edible cultivar groups of Cucurbita pepo (Cucurbitaceae). Econ. Bot. 1989, 43, 423–443. [Google Scholar] [CrossRef]
- Jeffrey, C. Cucurbita. In Mansfeld’s Encyclopedia of Agricultural and Horticultural Crops (Except Ornamentals); Hanelt, P., Institute of Plant Genetics and Crop Plant Research, Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 1541–1552. [Google Scholar]
- Andres, T.C. Diversity in tropical pumpkin (Cucurbita moschata): A review of infraspecific classifications. In Progress in Cucurbit Genetics and Breeding Research, Proceeding of the Cucurbitaceae 2004, Olomouc, Czech Republic, 12–17 July 2004; Lebeda, A., Paris, H.S., Eds.; Palacky University: Olomouc, Czech Republic, 2004; pp. 107–112. [Google Scholar]
- Castetter, E.F. Horticultural groups of Cucurbita. Proc. Am. Soc. Hortic. Sci. 1925, 22, 338–340. [Google Scholar]
- Goldman, A. The Compleat Squash: A Passionate Grower’s Guide to Pumpkins, Squash, and Gourds; Artisan: New York, NY, USA, 2004; p. 214. [Google Scholar]
- Grebenscikov, I. Notulae Cucurbitologicae III. Die Kult. 1958, 6, 38–60. [Google Scholar] [CrossRef]
- Cummings, M.B.; Jenkins, E.W. Pure Lines Studies with Ten Generations of Hubbard Squash; Free Press Printing Co: Burlington, VT, USA, 1928; p. 29. [Google Scholar]
- López-Anido, F.S. Diversidad Morfológica y Molecular en Cucurbita maxima Duchesne ex Lam. Ph.D. Thesis, Facultad de Ciencias Agrarias, Universidad Nacional de Rosario, Zavalla, Argentina, 2017. [Google Scholar]
- López-Anido, F. Genetics of seed coat colour in winter squash. In Proceedings of the 7th International Horticulture Research Conference, College of Horticulture and State Key Laboratory of Crop Stress Biology for Arid Areas of Northwest A&F University, Nanjing, China, 1–30 July 2020. [Google Scholar]
- Formiga, A.K.; Myers, J.R. Images and Descriptions of Cucurbita maxima in Western Europe in the Sixteenth and Seventeenth Centuries. Plant. Breed. Rev. 2020, 43, 317–356. [Google Scholar] [CrossRef]
- Janick, J. Giant Pumpkins: Genetic and Cultural Breakthroughs. Chron. Hortic. 2008, 48, 16–17. [Google Scholar]
- Paris, H.S.; Nerson, H. Seed dimension in the subspecies and cultivar-groups of Cucurbita pepo. Genet. Res. Crop Evol. 2003, 50, 615–625. [Google Scholar] [CrossRef]
- Paris, H.S.; Brown, R.N. The genes of pumpkin and squash. HortScience 2005, 40, 1620–1630. [Google Scholar] [CrossRef]
- Langevin, D.; Janick, J. New World Record of Giant Pumpkin, 2010. Chron. Hortic. 2011, 51, 24. [Google Scholar]
- Cumarasamy, R.; Corrigan, V.; Hurst, P.; Bendall, M. Cultivar differences in New Zealand “Kabocha” (buttercup squash, Cucurbita maxima). N. Z. J. Crop Hortic. Sci. 2002, 30, 197–208. [Google Scholar] [CrossRef][Green Version]
- Heiden, G.; Barbieri, R.L.; Neitzke, R.S. Chave para a Identificação das Espécies de Abóboras (Cucurbita, Cucurbitaceae) Cultivadas no Brasil; Embrapa Clima Temperado: Pelotas, Brazil, 2007; p. 32. [Google Scholar]
- Herrington, M.E.; Prytz, S.; Wright, R.M.; Walker, I.O.; Brown, P.; Persley, D.M.; Greber, R.S. ‘Dulong QHI’ and ‘Redlands Trailblazer’, PRSV-W-, ZYMV-, and WMV-resistant Winter Squash Cultivars. HortSience 2001, 36, 811–812. [Google Scholar] [CrossRef]
- Ferriol, M.; Picó, B.; Nuez, F. Morphological and molecular diversity of a collection of Cucurbita maxima landraces. J. Am. Soc. Hortic. Sci. 2004, 129, 60–69. [Google Scholar] [CrossRef]
- Kazminska, K.; Sobieszek, K.; Targonska-Karasek, M.; Korzeniewska, A.; Niemirowicz-Szczytt, K.; Bartoszewski, G. Genetic diversity assessment of a winter squash and pumpkin(Cucurbita maxima Duchesne) germplasm collection based ongenomic Cucurbita-conserved SSR markers. Sci. Hortic. 2017, 219, 37–44. [Google Scholar] [CrossRef]
- Esquinas Alcazar, J.T.; Gulick, P.J. Genetic Resources of Cucurbitaceae; IBPGR: Rome, Italy, 1983; p. 100. [Google Scholar]
- UPOV. Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability, Pumpkin (Cucurbita maxima Duch.), TG/155/4; UPOV: Geneva, Switzerland, 2007; p. 29. [Google Scholar]
- Martínez, A.B.; Lema, V.S.; Capparelli, A.; López-Anido, F.; Benech-Arnold, R.L.; Bartoli, C.G. Differences in seed dormancy associated with the domestication of Cucurbita maxima: Elucidation of some mechanisms behind this response. Seed Sci. Res. 2018, 28, 1–7. [Google Scholar] [CrossRef]
- Hammer, K.; Gladis, T. Notes on infraspecific nomenclature and classifications of cultivated plants in Compositae, Cruciferae, Cucurbitaceae, Gramineae (with a remark on Triticum dicoccon Schrank) and Leguminosae. Genet. Resour. Crop Evol. 2014, 61, 1455–1467. [Google Scholar] [CrossRef]
- Gower, J.C. Generalized procrustes analysis. Psychometrika 1975, 40, 33–51. [Google Scholar] [CrossRef]
- Whitaker, T.W.; Carter, G.F. Critical Notes on the origin and Domestication of the Cultivated Species of Cucurbita. Am. J. Bot. 1946, 33, 10–15. [Google Scholar] [CrossRef]


















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Anido, F.S. Cultivar-Groups in Cucurbita maxima Duchesne: Diversity and Possible Domestication Pathways. Diversity 2021, 13, 354. https://doi.org/10.3390/d13080354
López-Anido FS. Cultivar-Groups in Cucurbita maxima Duchesne: Diversity and Possible Domestication Pathways. Diversity. 2021; 13(8):354. https://doi.org/10.3390/d13080354
Chicago/Turabian StyleLópez-Anido, Fernando Sebastián. 2021. "Cultivar-Groups in Cucurbita maxima Duchesne: Diversity and Possible Domestication Pathways" Diversity 13, no. 8: 354. https://doi.org/10.3390/d13080354
APA StyleLópez-Anido, F. S. (2021). Cultivar-Groups in Cucurbita maxima Duchesne: Diversity and Possible Domestication Pathways. Diversity, 13(8), 354. https://doi.org/10.3390/d13080354