Abstract
Coral reef resilience is greatly influenced by herbivory. There is a need to identify key fish species fulfilling this critical function in biogeographically distinct regions. This experimental in situ study investigated fish herbivory in coral reefs of the lower Gulf of Thailand characterized by a considerably low herbivorous fish biomass and diversity, but high live coral and low macroalgal cover. This provided an intriguing situation for macroalgal browsing research. Visual census techniques assessed the abundance of local herbivorous fish species, and filmed single-choice assays using the macroalga Turbinaria evaluated mass-standardized bites (ms-bites) and biomass removal. Multiple-choice assays offering four locally abundant macroalgae identified specific biomass removal and ms-bites to uncover selection and avoidance patterns of observed fish species. The rabbitfish Siganus virgatus constituted only 39% of herbivore biomass but accounted for 90% of ms-bites. In multiple-choice assays, fishes took most (61%) bites on Sargassum, followed by Padina (28%) and Turbinaria (11%), while Lobophora was avoided. S. virgatus exhibited the most generalized browsing pattern of all species observed. Coinciding with recent studies, our findings suggest that S. virgatus plays a key functional role in reefs characterized by low diversity of herbivores and low functional redundancy.
1. Introduction
Coral reefs constitute the taxonomically most diverse ecosystems on earth [1]. The widespread impact of anthropogenic stressors, compounded by natural disasters, can initiate a cascade of events favoring shifts to macroalgae dominance [1,2,3]. It is well established that herbivorous fishes play a key role ensuring the continued functioning of reef systems, particularly in the face of negative impacts, due to their ability to remove algae, one of the main competitors of corals [2,4,5,6]. Nominally herbivorous fishes have traditionally been classified into a small number of distinct functional groups, e.g., based on dietary preferences and methods of food acquisition [7]. Grazers predominantly feed on algal turfs and early life stages of macroalgae, thereby potentially preventing macroalgae establishment [2]. The removal of morphologically and chemically defended mature macroalgae is limited to the functional group of macroalgal browsers [1,7]. Chronic drivers such as overfishing and nutrient pollution can lead to transitional states, with turf-dominated reefs eventually developing into reefs dominated by macroalgae [8,9], but recent literature suggests that turf can also constitute a stable regime in itself [10,11,12]. Macroalgae can outgrow feeding pressure when grazing is low, thereby reducing coral settlement, recruitment [13,14] and growth [15], and overgrowing adult corals [16,17,18]. Similarly, turf algae can inhibit coral larvae settlement and decrease survivorship [19,20]. Increasing reports of coral reefs shifting to macroalgae dominance following large-scale bleaching events [5,21,22,23] have given rise to a growing number of studies on browsing fishes (e.g., [24,25,26]; for a full list of Indo-Pacific species see [27]), of which most have been conducted on the Great Barrier Reef (GBR, e.g., [22,25,28]). Key species have been identified based on assessments of estimated fish feeding rates (“estimated function”, [29]) or measured loss of macroalgae biomass (“realized function” [29], e.g., [30,31,32]. However, high variability in the identity of fish species removing macroalgae together with biogeographic differences prevent generalizations [6,33,34], and herbivorous fishes can display nuanced differences in their function across different locations [25,26,30]. Besides physical, chemical, and nutritional traits of the macroalgae coupled with herbivore gut physiology that already exclude most species as potential feeders [24,35,36,37], numerous factors can influence the feeding selectivity of browsing species, such as temporal and spatial patterns of algal availability [25,28].
On the GBR, studies have repeatedly identified major browsing species to exhibit a strong selection for the brown alga Sargassum [26,38,39]. Some of the most consistently important macroalgal browsers in the Indo-Pacific region include Naso unicornis [26,39,40], Kyphosus vaigiensis [26,36], Siganus canaliculatus [38,41] and Siganus doliatus [25,42] (see [6]). However, contrasting results exist regarding the two rabbitfish species having an impact functionally equivalent to N. unicornis and K. vaigiensis in their efficiency of macroalgae removal [38] (but see [43]). Species-specific feeding patterns can lead to varying extents of dietary overlap across a herbivore community [44], resulting in different levels of functional redundancy or complementarity within a reef system [24,30,39]. Pronounced variations in feeding selectivity may be concealed by classifying species into broad functional groups, and assumptions of functional redundancy may be challenged once finer degrees of niche partitioning are considered [42,43,45].
While high diversity systems provide the potential for functional redundancy, the redundancy can be limited in some functional groups [1]. Few species or even a single species can drive macroalgal removal irrespective of the level of herbivore diversity [22,30,39]. Some evidence suggests that species might express greater behavioral flexibility in lower-diversity assemblages, being able to support the function of macroalgal browsing to a similar extent compared to study sites richer in species and thereby increasing resilience [6].
Coral reefs located in the lower Gulf of Thailand such as the ones of the Samui Archipelago provide an interesting research ground for macroalgal browsing, as they exhibit low fish taxonomic diversity and face serious disturbances caused by extensive tourism, overfishing and water pollution [46,47]. To date, little is known about key browsing species including their feeding ecology and functional roles in this region [27]. All study sites exhibited low diversity of herbivorous species and, most strikingly, these reefs are lacking a major component of the herbivorous fish guild–acanthurids [26,39,40]. Based on initial information, herbivorous biomass as well as abundance of invertebrate macroalgae grazers seem to be particularly low as well [48].
The use of remote underwater video cameras has emerged as a viable tool to assess the identity of key species and feeding behavior using transplanted macroalgae in the field [24,26,39], and multiple-choice feeding assays offering different macroalgae species proved a suitable method to assess feeding selectivity of fish species [24,30]. Here, we used visual census techniques to (i) assess the abundance of locally occurring herbivorous fish species, and (ii) filmed macroalgal single-choice assays using the macroalga Turbinaria to identify mass-standardized bites (ms-bites) and biomass removal by grazing and browsing fish species. This was supplemented by multiple-choice assays offering locally abundant macroalgae (Sargassum, Turbinaria, Padina and Lobophora) to (iii) evaluate specific biomass removal and ms-bites to uncover selection and avoidance patterns of observed fish species. The findings of this study contribute to the limited body of information for reefs with low herbivorous fish diversity, particularly in the lower Gulf of Thailand.
2. Materials and Methods
2.1. Study Site
Data were collected at four different sites off the island Ko Pha Ngan, namely Haad Salad (A; 9°47′22.0″ N 99°58′17.1″ E), Mae Haad IV (D; 9°47′32.2″ N 99°58′32.6″ E), Mae Haad II (B; 9°47′47.6″ N 99°58′41.9″ E) and Haad Khom (C; 9°47′53.1″ N 100°00′52.0″ E) (Figure 1), and from a fifth site in the nearby marine park Ang Thong (E; 9°47′55.6″ N 99°42′44.8″ E) located in the lower Gulf of Thailand.
Figure 1.
Map of study sites. (A) Haad Salad, (D) Mae Haad IV, (B) Mae Haad II, (C) Haad Khom and (E) Ang Thong Marine Park.
Macroalgae single-choice experiments were conducted between February and June 2015 to assess potential differences on the reef crest of one openly fished site (loc D) and the marine park site (loc E). Macroalgal assays in 2018 were conducted between February and June on the reef flat in three local fished reefs for single-choice assays (loc A–C) and in one fished reef for multiple-choice assays (loc D). The data collection periods covered the dry and part of a transitional season, with southeast monsoons starting from May onwards [49]. The four Ko Pha Ngan study sites (loc A–D) were located on fringing reefs off the north coast of the island subjected to frequent anthropogenic activity [48] and were chosen to investigate macroalgae browsing around the island. At those sites, the reef flat emerges from a sandy beach at a water depth of 1–2 m and stretches into fields of macroalgae composed by the four macroalgal genera Turbinaria, Sargassum, Padina and Lobophora, coral patches dominated by Porites sp. [48], and rubble. Then, it evolves into the reef crest at a water depth of 3–6 m and slopes down to the fore reef at a water depth of 10 m. Lobophora was the only macroalga present beyond a water depth of 3 m. The fifth study site (loc E) was situated within a marine protected area, which is a no-take zone by law.
2.2. Fish and Benthic Surveys
For all study sites and years, fish and benthic surveys were carried out every two weeks on the reef crest during the study period in less than 50 m from macroalgae assay deployment sites. The herbivorous fish community was assessed by visual census [7] on SCUBA along three 5 m × 50 m belt transects, yielding a total of 250 m2 per transect. Each survey consisted of two divers trained in fish identification swimming left and right from the transect tape recording all fishes identified to species level. Nominally herbivorous fish species were considered in the surveys, including rabbitfishes (Siganidae), parrotfishes (Labridae: Scarinae), surgeonfishes (Acanthuridae), chubs (Kyphosidae), and pomacanthids (genus Pomacanthus) [7]. Individual fish size was estimated and categorized into nine different size classes (0.1–5 cm, 5.1–10 cm, 10.1–15 cm, 15.1–20 cm, 20.1–25 cm, 25.1–30 cm, 30.1–35 cm, 35.1–40 cm and >40 cm). Herbivorous fish abundance and published length-weight data [50] were used to calculate fish biomass as g m−2. The benthic community composition was measured using line-point-intercept transects [51] along the same transects used to assess the herbivore community in 0.5 m steps. Benthos was grouped into “live coral cover”, “turf algae”, “macroalgae“, “rubble”, “sand” and “other life”. Replicates per transect for each surveyed location ranged from 8 to 15 in 2018 and from 9 to 12 in 2015. A period of heavy weather in the beginning of April 2018, with wind speeds of up to 15 knots at the local sites, and difficult accessibility of the marine park resulted in a variation in replicate numbers. A table with replicate numbers of all surveys and macroalgae assays can be found in the Supplementary Material (Table S1).
2.3. Algae Feeding Assays
In this study, all reported values are stated as mean ± SE. In 2015, Turbinaria appeared to be the most abundant macroalga (Puk, personal observations), which provided the reason for working with this genus in single-choice assays. For consistency, single-choice assays in 2018 also used the then second-most abundant genus Turbinaria. As a preparatory assessment, benthic surveys were conducted in February 2018 on the reef flat of the study sites with a replicate number of 3. Lobophora was the most abundant macroalga on the reef flat (6 ± 1%), followed by Turbinaria (5 ± 1%), Sargassum (2 ± 1%) and Padina (1 ± 1%) (see results section). Sargassum has been used in various feeding experiments investigating macroalgae browsing (e.g., [26,34]). However, Sargassum abundance was very low in both 2015 and 2018 experiments with a seasonal decline until almost completely absent, making the use in assays not feasible [52]. While Lobophora was the most abundant alga, its foliose and encrusting morphology makes it difficult to handle in assays [53].
An initial assessment of the diel activity pattern of herbivorous fishes was conducted in February 2018 to determine the most suitable time for macroalgae assays. An underwater video camera of the type GoPro Hero 3+ was deployed after sunrise (7:00) and recorded until sunset (18:30), with battery changes every 2 h. Recordings were analyzed counting the number of herbivorous fishes within each 2 h-block. A Kruskal–Wallis test detected no significant differences in the abundance of fishes between the observed 2 h-blocks (Table S2). Subsequently, assays were performed between 9:00 and 17:00 for 2 h without interruption. Longer recording times would have required battery changes leading to disturbance of potential feeding events, and previous studies using filmed assays have shown the highest bite rate within the first hour of deployment [24,54].
Macroalgae thalli for all assays were collected in adjacent shallow water habitats and kept in a saltwater tank with surface current no longer than 24 h until deployment. Suitable sizes of thalli were chosen, covering the natural variety of different sizes of the respective macroalga found in the study area (see Table S3 for initial weights). Macroalgae thalli were tied onto pieces of coral rubble with fishing line and deployed at the study sites within an area of 1 m2. Per assay, one camera was mounted on a tripod built out of PVC and placed in approximately 1 m distance from the used macroalgae to record feeding. To account for biomass loss during 2 h deployment, wet weight of algae was determined to the nearest 0.01 g before and after the assays with an Ohaus Pioneer digital scale. Excess water was removed by spinning algae in a salad spinner for 10 s before weighing. One alga for each genus was covered with a mesh bag and deployed next to each assay, acting as a control accounting for growth and biomass loss not attributed to herbivory (handling loss). Deployment spots were chosen haphazardly at the respective water depth, with at least 10 m distance to previous spots. A size reference held into the view frame at the beginning of each recording allowed length estimations of feeding fish. Proportionate and absolute biomass loss due to herbivory were calculated accounting for the biomass loss of the control, which simulated biomass loss due to handling: Tloss = 1 − Aa/(Ai × [1 − Hloss]), where Tloss is the proportion of biomass lost, Aa is the algae biomass left after 2 h, Ai is the initial biomass measured before deployment and Hloss is the mean percentage of handling loss, calculated based on the controls [25].
Single-choice Turbinaria assays conducted in 2015 on the reef crest (3–6 m water depth) were used to compare feeding on Ko Pha Ngan (D; 44 assays and 88 h of video recordings) with the marine park (E; 18 assays and 36 h of video recordings).
In 2018, single-choice assays were placed on the reef flat (1–2 m water depth) in locations A–C (total of 93 assays; 186 h of video recordings) to assess the rate of herbivory in the reef zone in which all four macroalgal genera naturally occurred during the time of the experiment.
Additionally, multiple-choice assays at location D used all four genera of macroalgae (Turbinaria, Sargassum, Padina and Lobophora) in one setup to investigate selection and avoidance of deployed macroalgae. As Lobophora was the only macroalga occurring in water depths beyond 3 m, multiple-choice assays were also conducted on the reef flat (39 assays; 78 h of video recordings). Feeding selectivity was defined by the likelihood that a fish chooses an alga of one genus over another macroalgae genus, if offered on an equal basis [55]. “Proportional-sized” specimens of algae were selected to reflect variation in growth form and size between species at the study site D as required for measures of preference [24] rather than “equal-sized” specimens in terms of biomass and size [54].
2.4. Data Analyses
Video recordings were analyzed for fish species feeding on deployed macroalgae, the size of consumers, and the number of bites taken. Mass-standardized bites (ms-bites) were calculated by multiplying biomass (estimated using fish size and published length-weight relationships, [50]) with observed number of bites (ms-bites = biomass of fish (in kg) × number of bites) following Hoey and Bellwood [39] to account for the herbivory impact of each fish species. Statistical analyses were performed using the software R [56]. Significance level α was 0.05 for all tests. In advance, data were tested for normal distribution using the Kolmogorov–Smirnov test, and homogeneity of variances was tested using Levene’s test. Statistical tests were chosen accordingly. Differences in absolute algae biomass before and after feeding assays associated with ms-bites were tested for single-choice assays at site E in 2015 and for multiple-choice assays at location D in 2018 using a Wilcoxon signed-rank test for dependent samples. Remaining locations were not tested due to insufficient sample sizes. A Wilcoxon rank sum test was performed to compare the number of total ms-bites from all fish species together and per fish species separately on single-choice Turbinaria assays at location D with location E in 2015 to test for differences in feeding activity between a local reef and the marine protected area. Correlation between ms-bites from all assays and years and associated proportionate biomass loss was tested using a Spearman correlation test.
Feeding selectivity of individual fish species in multiple-choice assays at location D in 2018 was evaluated according to selection and/or avoidance of offered macroalgae. Positive selection, avoidance, or random selection of offered macroalgae in multiple-choice assays was tested using the Strauss’ Linear Selection Index L = ri − pi, where ri is the proportion of the i-th food type consumed and pi is the proportion of the i-th food type available in the environment [57]. This electivity index was chosen because it is more robust to sampling errors at low r and p values compared to other indices [58]. Due to an overall low number of observed bites, L values could only be calculated for the most dominant browser S. virgatus. In addition, the non-parametric Friedman test was performed to test for a difference in the number of ms-bites taken from the four offered macroalgae Turbinaria, Sargassum, Padina and Lobophora. The non-parametric Friedman test for dependent samples was chosen because the selection of one macroalgal genus may be dependent on the presence of other algal genera during a trial [59,60]. Further, the Friedman test was used to evaluate the feeding selectivity of the most dominant browser S. virgatus according to ms-bites. A Friedman post hoc multiple comparison test according to the Nemenyi method [61] was conducted to perform pairwise comparisons of ms-bites taken by S. virgatus on Turbinaria, Sargassum and Padina.
3. Results
Six rabbitfish species (Siganus virgatus, Siganus javus, Siganus corallinus, Siganus guttatus, Siganus vermiculatus and Siganus stellatus) and two parrotfish species (Scarus rivulatus and Scarus ghobban) were observed in all herbivore surveys across years and sites. Sightings of Naso unicornis, Kyphosus vaigiensis and Pomacanthus sexstriatus occurred rarely (three individuals or less in all assays, ≤2% of biomass).
3.1. Key Browsers and Macroalgae Removal in the Fished vs. Protected Reef
In 2015, the marine park showed a herbivorous fish biomass of 14.1 ± 1.4 g m−2. S. rivulatus made up 36% of herbivorous fish biomass at that site. It was followed by S. virgatus (31%), S. javus (14%), S. ghobban (8%), S. stellatus (4%), S. corallinus and S. guttatus (each 2%) (Figure 2, full details in Table S4). The marine park had a high live coral cover (49 ± 4%), 8 ± 2% of turf algae and no macroalgae (Table 1). The Ko Pha Ngan site D showed less than half of the herbivorous fish biomass of the marine park (6.3 ± 0.6 g m−2). Additionally, S. rivulatus (59%) and S. virgatus (30%) made up most of the herbivore biomass. Benthic community composition showed high live coral cover (43 ± 3%), but also high turf algae cover (32 ± 2%). Lobophora was the only macroalga present (12 ± 4%).
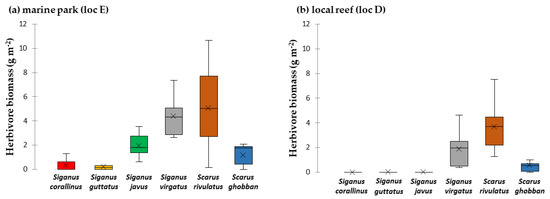
Figure 2.
Herbivore biomass surveyed on the reef crest in the marine park (loc E) (a) and in a local reef (loc D) (b) in 2015. Values are given in mean g m−2. The boxplot shows the median and 25% quantile, the black cross indicates the mean.

Table 1.
Benthic community composition on the reef crest in the local reef (loc D) and the marine park (loc E) in 2015. Given are % of total cover for live coral cover (LCC), turf algae, sand, rubble, other life and macroalgae, whereas macroalgae are split into the genera Turbinara, Lobophora, Padina and Sargassum. Replicate number n = 12 at loc D and n = 9 at loc E.
In the 2015 single-choice experiments on the reef crest, a total of 438.74 ms-bites during 124 h of recording (3.54 ± 0.68 ms-bites h−1) were observed (Figure 3). S. virgatus took by far most bites from deployed Turbinaria assays, accounting for 93% of all ms-bites (3.29 ± 0.37 ms-bites h−1), followed by S. corallinus (5% of all ms-bites, 0.16 ± 0.09 ms-bites h−1) and S. guttatus, S. rivulatus, S. ghobban and S. javus which took less than 5% of all ms-bites, respectively. The bite rate by all fish species combined was 17-times lower at location D (0.63 ± 0.36 ms-bites h−1) than at the site E (10.64 ± 1.14 ms-bites h−1; p = 0.002, Table S5). This difference was mainly driven by S. virgatus, which took 10.37 ± 0.71 ms-bites h−1 at location E and 0.39 ± 0.26 ms-bites h−1 at location D (p < 0.001, Table S5).
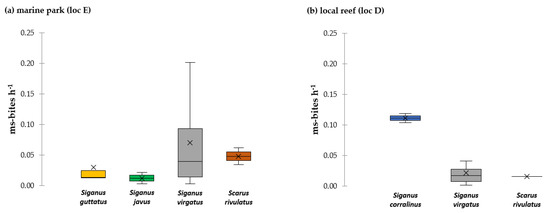
Figure 3.
Bite rate (ms-bites h−1) for recorded fish species in single-choice assays on the reef crest in the marine park (loc E) (a) and in a local reef (loc D) (b) in 2015. Fish species are color-coded. The boxplot shows the median and 25% quantile, the black cross indicates the mean.
3.2. Key Browsers and Macroalgae Removal in Local Reefs
Across the four Ko Pha Ngan sites in 2018, the average herbivorous fish biomass was 10.8 ± 2 g m−2 and ranged from 3.9 ± 0.2 g m−2 to 13.8 ± 0.6 g m−2. S. virgatus made up 49% of herbivorous fish biomass. It was followed by S. rivulatus (32%), S. ghobban (16%), S. guttatus (3%), and less than 2% for S. corallinus, S. stellatus and S. javus, respectively (Figure 4, full details in Table S6). Benthos community composition on the reef flat showed high turf algae cover (39 ± 4%), moderate live coral cover (22 ± 7%) and 12 ± 3% macroalgae (Table 2). Benthic data for sites A–D on the reef crest in 2018 was within a similar range as in 2015 for the fished reef (Table S7).
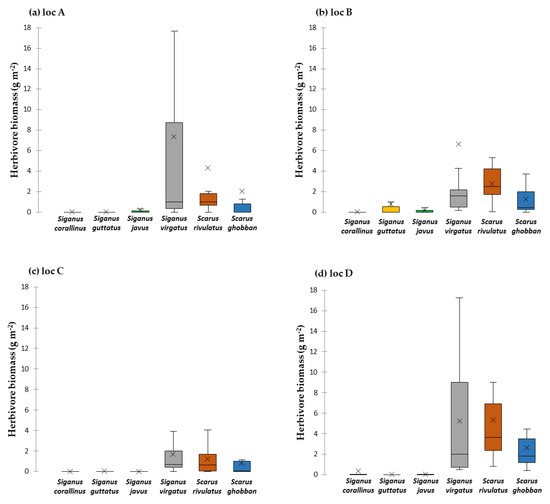
Figure 4.
Herbivore biomass surveyed on the reef crest near locations of single-choice assays loc A (a), loc B (b) and loc C (c) and multiple-choice assays loc D (d) in 2018. Values are given in mean g m−2. The boxplot shows the median and 25% quantile, the black cross indicates the mean.
Table 2.
Benthic community composition surveyed on the reef flat at locations of single-choice (loc A–C) (a) and multiple-choice assays (loc D) (b) in 2018. Given are % of total cover for live coral cover (LCC), turf algae, sand, other life and macroalgae, whereas macroalgae are split into the genera Turbinara, Lobophora, Padina and Sargassum. Replicate number n = 3 at each site.
3.2.1. Single-Choice Assays
While no bites were recorded at locations A and C, a single feeding event of S. virgatus was recorded at location B (3.60 ms-bites in 186 h of assays).
3.2.2. Multiple-Choice Assays
Overall, 31.48 ms-bites in 78 h of assays (0.40 ± 0.73 ms-bites h−1) were recorded in multiple-choice assays on the reef flat in 2018. The most important macroalgal consumer in all conducted assays was S. virgatus, which took 52% of all ms-bites (0.21 ± 0.09 ms-bites h−1), followed by S. guttatus (19% of all ms-bites, 0.08 ± 0.32 ms-bites h−1), S. rivulatus (12% of all ms-bites, 0.05 ± 0.17 ms-bites h−1) and S. corallinus (11% of all ms-bites, 0.05 ± 0.14 ms-bites h−1). S. ghobban and S. javus took less than 4% of all ms-bites (Figure 5).
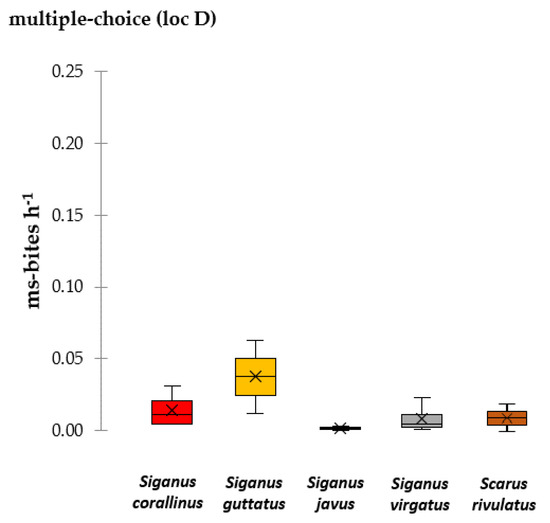
Figure 5.
Bite rates (ms-bites h−1) for recorded fish species in multiple choice-assays (loc D) in 2018. Fish species are color-coded. The boxplot shows the median and 25% quantile, the black cross indicates the mean.
3.2.3. Feeding Selection and Avoidance
During the 78 h of recordings, 65% of all ms-bites (0.26 ± 0.26 ms-bites h−1) were observed on Sargassum, 25% of all ms-bites were taken on Padina (0.10 ± 0.07 ms-bites h−1), followed by Turbinaria with 10% of all ms-bites (0.04 ± 0.48 ms-bites h−1), and Lobophora was avoided in all assays (Figure 6). The Strauss’ Linear Selection Index (L) showed a nominal positive selection for Sargassum and Padina, as well as a slight avoidance of Turbinaria, by the most dominant browser S. virgatus (Table 3). The Friedman test revealed a significant selection/avoidance pattern for S. virgatus (p < 0.001, Table S8). Sargassum (55% of all ms-bites) and Padina (41% of all ms-bites) were selected with a similar frequency by S. virgatus, while Turbinaria seemed to be less attractive (4% of all ms-bites; Sargassum − Turbinaria p = 0.008 and Padina − Turbinaria p = 0.001, Table S9) and Lobophora was always avoided. Due to an overall low number of observed bites, statistical tests could only be performed for the most dominant browser S. virgatus. Based on observations in the field, the siganids were highly selective for Sargassum. S. javus took 86% of all its ms-bites on Sargassum, while S. guttatus and S. corallinus exclusively selected Sargassum. The parrotfish S. rivulatus fed mainly on Turbinaria (73% of all ms-bites exerted by S. rivulatus) but was also observed to take bites on Padina (15% of ms-bites) and Sargassum (12% of ms-bites). S. ghobban was observed to feed on Padina only.
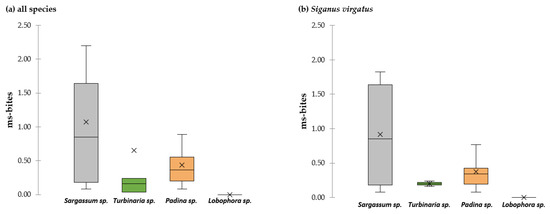
Figure 6.
Ms-bites in multiple-choice assays (loc D) in the reef flat in 2018 taken on the four offered macroalgae genera (Sargassum, Padina, Turbinaria and Lobophora) for all fish species (a) and for the most dominant browser S. virgatus (b). No bites on Lobophora were recorded. The boxplot shows the median and 25% quantile, the black cross indicates the mean.
Table 3.
Strauss’ Linear Selection Index (L) tested for positive or negative selection of the four offered algae genera Sargassum, Padina, Turbinaria and Lobophora by S. virgatus in multiple-choice assays in the reef flat (loc D) in 2018. Ri is the proportion of the i-th food type consumed and Pi is the proportion of the i-th food type available in the environment.
3.3. Biomass Removal
The difference in biomass for single-choice assays in the marine park (loc E) in 2015 was significant (0.15 ± 0.05 g h−1 [0.2 ± 0.01% h−1]; p < 0.001, Table S10).
Further, multiple-choice assays conducted on the reef flat (loc D) in 2018 showed a significant difference in absolute biomass loss of 0.04 ± 0.07 g h−1 (0.38 ± 0.07% h−1) (p < 0.01, Table S10), whereby Padina had the highest removal rate (0.21 ± 0.03% h−1), followed by Sargassum (0.15 ± 0.04% h−1) and Turbinaria (0.03 ± 0.01%h−1). For Lobophora, no biomass loss associated with herbivory was detected. Values for biomass loss are given in Table 4 (2015) and Table 5 (2018).
Table 4.
Biomass loss of macroalgae in single-choice assays in a local reef (loc D) (a) and in the marine park (loc E) (b) on the reef crest in 2015. Given are absolute biomass loss in g h−1 ± SE as well as biomass loss in % h−1 ± SE per algae genus. Locations with significant biomass loss are marked with *.
Table 5.
Biomass loss of macroalgae in single-choice assays at loc C (a) and multiple-choice assays at loc D (b) on the reef flat in 2018. Given are absolute biomass loss in g h−1 ± SE as well as biomass loss in % h−1 ± SE per algae genus. Locations with significant biomass loss are marked with *.
No correlation was found between ms-bites and measured biomass loss for all assays (Table S11).
4. Discussion
4.1. Identification of Key Browsers and Macroalgae Removal
The herbivorous fish biomass at all sites of this study was lower than the average herbivore biomass in locations accessible to fisheries throughout the Indo-Pacific (10.8 ± 2 g m−2 vs. 29.0 g m−2, [62,63]). Fishing has been observed at all sites including the marine park as the fishing ban is typically not enforced. The group of rabbitfishes made up 53% of total herbivore biomass on average, and S. virgatus alone constituted 49% of that biomass. Parrotfishes comprised 47% of the herbivore community, whereas surgeonfishes, which constitute an important group of herbivores in other reef systems [33,39,64], were absent. S. virgatus was the single most important browser observed at all study locations, exerting 90% of all ms-bites at all locations combined, ranging from 97% at location E and 62% at location D in single-choice assays in 2015 to 52% in multiple-choice assays in 2018.
The single-choice experiments aimed to gain insight into key herbivores and to assess rates of herbivory. The bite rate on the reef crest of location D of Ko Pha Ngan was 17 times lower than at the marine park site and was driven largely by S. virgatus. Additionally, higher background densities of algae at the Ko Pha Ngan sites may have deterred feeding [65,66]. In turn, the deployed Turbinaria at the protected marine park site E, which harbored turf algae in low abundance (<10%) and no macroalgae, appeared to have been an attractive subsidy. Feeding on the Turbinaria algae from the assays may be beneficial compared to the day-to-day foraging strategy of herbivorous fish, as the effort needed to crop small algae in complex reef habitats entails a significant risk of predation and/or a lower feeding efficiency [67,68]. Transplanted assays are also likely to be more apparent and may attract browsers from beyond their normal foraging range [64]. Biomass removal at location E was significant (0.2 ± 0.01% h−1).
The single-choice experiments on the reef flat, the reef zone with the highest macroalgae cover and in which all genera were naturally occurring, showed virtually no feeding. This is in line with previous studies, in which significantly lower rates of herbivory compared to other reef habitats were recorded on the reef flat (e.g., [69]). The reef crest generally has the highest diversity of fishes [70,71], extensive territoriality with fishes protecting preferred feeding locations [72], the highest rates of primary productivity [73,74], and it often is the preferred feeding habitat for macroalgae browsers [42,69]. The overall high availability of algae in the Ko Pha Ngan reef flats likely led to a low attractiveness of the algae assays compared to the surrounding, resulting in diluted feeding pressure as assays are likely to be less apparent [25,64,75]. Similarly, macroalgal removal may decrease with higher macroalgal density due to higher predation risk and/or the reduced ability of herbivores to detect predators [65,69,76]. However, variation in initial algae biomass in single-choice assays may have influenced feeding rates as resource quantity can influence herbivory [77], hence leading to a difference in observed patterns between the single-choice assays at different sites.
Turbinaria was selected for the single-choice assays due to its higher relative abundance compared to other macroalgae and because of the low seasonal abundance of Sargassum, which is regularly used in feeding assays. However, the use of Turbinaria could be an explanation for the rather low feeding rates in our study, since it is usually not preferred by herbivores such as S. virgatus as it has a rather low palatability due to several morphological and chemical defenses [78,79,80] and is mainly consumed by large herbivores such as N. unicornis and K. vaigiensis not present in this study [30,36,42]. We therefore suggest further feeding assessments using single-choice assays of Sargassum as well as of Turbinaria during different times of the year to uncover possible variations in herbivory linked to macroalgae genus and algae condition in the studied reefs [52,81,82].
The capability to digest macroalgae varies between herbivores and may determine the benefit gained by feeding on them. The analysis of short chain fatty acid (SCFA) levels of different herbivores places siganids in between browsers, such as kyphosids and acanthurids feeding on large macroscopic algae including fucoids, and scarine parrotfishes which feed on detritus, indicating a mixed diet with algal components [83]. The three siganid species observed at the study sites, S. guttatus, S. corallinus and S. javus, may not have been capitalizing on the subsidies due to different feeding preferences, as they are considered omnivorous [41,84,85] or primarily feed on turf [86]. All assays tested the consumption of macroalgae only. Since turf algae is a major component of coral reef systems, further investigation to elucidate grazing on turf algae on the flat and crest of the studied reefs would yield interesting results.
The browsing rabbitfish S. virgatus contributed 97% of mass-standardized bites at site E. S. virgatus is a mixed herbivore feeding on both macroalgae and the epilithic algal matrix [87]. In recent years several studies have emerged on the role of S. virgatus contributing towards macroalgal (i.e., Sargassum) removal within various ecosystems in the central Indo-Pacific region, and it has been concluded that S. virgatus is the most functionally important macroalgal remover along a gradient of reef degradation, under varying environmental conditions and over different spatial and temporal scales [34,76,82,88].
Our results clearly show that S. virgatus thrives in surroundings with virtually no edible macroalgae present, such as the reef flat of this study, feeding on turf algae and small thalli of macroalgae species, but is also capable of consuming adult macroalgae stands. S. virgatus is closely related to S. doliatus, and their division into separate species is considered incipient [89]. Although S. doliatus has been previously identified as an algal cropper [41,70,90] and its functional impact has been questioned [41,43], S. doliatus is capable of consuming adult macroalgal stands and its abundance in many inshore regions of the GBR has led to the suggestion that macroalgal browsing in those reefs might be reliant on S. doliatus as a single species [91].Like its sister species S. doliatus, S. virgatus is a mixed herbivore with high functional plasticity, and its prevalence and feeding habits may prove useful in low-diversity systems to sustain important functions in the absence of more specialized herbivores such as K. vaigiensis and N. unicornis [41,42].
4.2. Selection and Avoidance of Macroalgae in Multiple-Choice Assays
S. virgatus and S. rivulatus displayed a broader feeding pattern in multiple-choice assays at location D, consuming three out of four offered macroalgae, while the other herbivores observed were highly selective. Biomass loss in multiple-choice assays (0.4 ± 0.01% h−1) was considerably low [54]. Only a few studies have been able to find a relationship between feeding intensity and biomass loss of macroalgae ([39], but see [38]). However, the variation of initial weights in algae assays needs to be considered as well, as it can influence the rate of herbivory and lead to lower biomass removal [77].
In line with previous observations [24,34,41], the brown alga Sargassum was most readily consumed in this study (65% of all ms-bites and 0.15 ± 0.04% h−1 biomass loss) and selected for by five out of six species, including all siganids. S. virgatus fed on Sargassum (55% of ms-bites), Padina (41% of ms-bites) and to a smaller extent on Turbinaria (4% of ms-bites), while the remaining siganid species almost exclusively selected Sargassum. Only 10% of all ms-bites in multiple-choice assays and a biomass loss of 0.03 ± 0.01% h−1 were observed on Turbinaria, which was targeted mainly by S. rivulatus (73% of ms-bites). Parrotfishes such as S. rivulatus and S. ghobban can readily consume macroalgae with strong structural defenses [36,92], and they consume various substrates to obtain their main food source endo- and epilithic microbial resources. Despite few reports of parrotfish species feeding on macroalgae on the GBR [24,54], macroalgae removal is considered incidental [37,83,93]. Epibionts living on Turbinaria constitute a secondary food source for browsing species [94], and epiphyte loads are known to shape feeding patterns [28]. Since epibionts were observed in the assays, there is a high likelihood that both S. rivulatus and S. virgatus were targeting epibionts in our assays, rather than Turbinaria itself, leading to incidental low biomass removal and explaining why no relationship between ms-bites and biomass loss could be found [22,28,38,90].
S. ghobban fed on Padina only, which was the second most targeted alga (25% of all ms-bites) and showed the highest biomass loss (0.2 ± 0.03% h−1). Lobophora was highly abundant at the study sites and no herbivory was observed in our assays. Indeed, most herbivore species are not able to derive nutrients from Lobophora [36,83,95] and a recent study showed different grazing fishes removing Lobophora recruits to varying degrees, while removal of adult Lobophora was very low [96].
Our study found low removal of algal biomass by herbivores on the reef crest and almost none on the reef flat, which questions whether the essential ecosystem function of macroalgal removal (the “realized function” [29]) in the fished reefs is maintained. However, the single-choice assays investigated feeding on Turbinaria, which is strongly defended and usually not a preferred macroalga [80]. As indicated by the results of our multiple-choice assays, at least two of the other occurring macroalgae genera are likely to be effectively removed from the reef crest, as corroborated by the observation that only Lobophora occurred at depths >3 m. The local reefs in our study showed a relatively high live coral cover and low macroalgae cover, indicating that herbivores successfully prevent macroalgae establishment by grazing on algal turfs and early life stages of macroalgae. However, further investigation is crucially needed to quantify overall grazing pressure of the fish species across different habitats. Our results corroborate the role of S. virgatus as an opportunistic herbivore, thriving in environments with varying turf algae and macroalgae availability and feeding on a range of algae, and hence demonstrating functional plasticity in response to environmental and biological variations. S. virgatus displayed a generalized feeding pattern in a herbivore community characterized by low biomass and diversity and consisting mostly of specialized species. S. virgatus appears to constitute a key consumer species in the study region and likely is of great importance in herbivore-depauperate reef systems such as the investigated sites.
Previous studies found the functional role of species to be highly context dependent [24], e.g., differing between locations and depending on co-occurring species [34,86]. Potential functional groups may be revealed in experiments, but anthropogenic and natural disturbances increasingly alter coral reef systems towards macroalgae being a dominant component [93,97]. The potential of herbivore species to exert critical functional roles in benthic communities, such as macroalgae browsing, may only be fully exposed when conditions change, and might be essential for coral reef resilience in the present and future.
5. Conclusions
Currently, the ecosystem function of macroalgae control seems sufficiently supported by the herbivorous fish community in the studied reefs. Based on the important role S. virgatus appears to play on the studied reefs, it should be given particular consideration in spatial and species-specific management efforts. This could be relevant to ensure provision of ecosystem functions and to bolster the resilience of reefs harboring species differing from the most consistently identified browsing species in times of frequent overfishing and increased anthropogenic disturbances.
Supplementary Materials
The following are available online at https://www.mdpi.com/1424-2818/13/3/123/s1, Table S1: study design, Table S2: diel assays, Table S3: Initial algae weights, Table S4: fish census data 2015, Table S5: Wilcoxon Test for ms-bites, Table S6: fish census data 2018, Table S7: benthos reef crest 2018, Table S8: Friedman Test, Table S9: Friedman post hoc Nemenyi Test, Table S10: Wilcoxon Test for biomass loss, Table S11: Spearman correlation, Table S12—ms-bites and raw bites in multiple- and single-choice assays, Table S13–S16: raw data.
Author Contributions
Conceptualization and methodology: M.M., C.F.K.S., L.D.P., E.M.S., S.C.A.F. and C.W.; data collection: M.M., C.F.K.S. and L.D.P.; data analysis: M.M., C.F.K.S., L.D.P., E.M.S.; writing—original draft preparation: M.M. and C.F.K.S.; writing—review and editing: C.W., S.C.A.F., L.D.P. and E.M.S.; visualization: M.M. and C.F.K.S.; funding acquisition: L.D.P. and C.F.K.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Studienstiftung des Deutschen Volkes and Kellner-Stoll-Stiftung für Klima und Umwelt.
Institutional Review Board Statement
Ethical review or approval were waived for this study, since only fish feeding behavior observations were involved. In addition, only non-invasive methodologies were used.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the Supplementary Material.
Acknowledgments
Surveys were conducted under a Memorandum of Understanding with the Department of Marine and Coastal Resources Thailand and the Phuket Marine Biological Center. The authors wish to express their gratitude to Janina Schoenig, Victoria Fahey, Stefan Follows of COREsea (Center for Oceanic Research and Education South East Asia) for their support during field work in Thailand and their warm hospitality. This manuscript is dedicated to the memory of Eike Schoenig, who passed away during the writing of this manuscript. He offered vital support and advice to young scientists and served as a role model due to his outstanding personality and advocation of the protection of coral reefs. He will continue to play an exceptional role through his legacy.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nat. Cell Biol. 2004, 429, 827–833. [Google Scholar] [CrossRef]
- Hughes, T.P.; Rodrigues, M.J.; Bellwood, D.R.; Ceccarelli, D.; Hoegh-Guldberg, O.; McCook, L.; Moltschaniwskyj, N.; Pratchett, M.S.; Steneck, R.S.; Willis, B. Phase shifts, herbivory, and the resilience of coral reefs to climate change. Curr. Biol. 2007, 17, 360–365. [Google Scholar] [CrossRef]
- Mumby, P.J. Phase shifts and the stability of macroalgal communities on Caribbean coral reefs. Coral Reefs 2009, 28, 761–773. [Google Scholar] [CrossRef]
- Mumby, P.J.; Harborne, A.R. Marine reserves enhance the recovery of corals on Caribbean reefs. PLoS ONE 2010, 5, e8657. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Jennings, S.; MacNeil, M.A.; Mouillot, D.; Wilson, S.K. Predicting climate-driven regime shifts versus rebound potential in coral reefs. Nat. Cell Biol. 2015, 518, 94–97. [Google Scholar] [CrossRef]
- Tebbett, S.B.; Hoey, A.S.; Depczynski, M.; Wismer, S.; Bellwood, D.R. Macroalgae removal on coral reefs: Realised ecosystem functions transcend biogeographic locations. Coral Reefs 2019, 39, 203–214. [Google Scholar] [CrossRef]
- Monitoring functional groups of herbivorous reef fishes as indicators of coral reef resilience. In A Practical Guide for Coral Reef Managers in the Asia Pacific Region, 2009 ed.; Green, A.L., Bellwood, D.R., Eds.; International Union for Conservation of Nature: Gland, Switzerland, 2009; ISBN 283171169X. [Google Scholar]
- Done, T.J. Phase shifts in coral reef communities and their ecological significance. Hydrobiologia 1992, 247, 121–132. [Google Scholar] [CrossRef]
- Hughes, T.P. Catastrophes, phase shifts, and large-scale degradation of a caribbean coral reef. Science 1994, 265, 1547–1551. [Google Scholar] [CrossRef]
- Jouffray, J.-B.; Nyström, M.; Norström, A.V.; Williams, I.D.; Wedding, L.M.; Kittinger, J.N.; Williams, G.J. Identifying multiple coral reef regimes and their drivers across the Hawaiian archipelago. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20130268. [Google Scholar] [CrossRef]
- Donovan, M.K.; Friedlander, A.M.; Lecky, J.; Jouffray, J.-B.; Williams, G.J.; Wedding, L.M.; Crowder, L.B.; Erickson, A.L.; Graham, N.A.J.; Gove, J.M.; et al. Combining fish and benthic communities into multiple regimes reveals complex reef dynamics. Sci. Rep. 2018, 8, 16943. [Google Scholar] [CrossRef]
- Jouffray, J.-B.; Wedding, L.M.; Norström, A.V.; Donovan, M.K.; Williams, G.J.; Crowder, L.B.; Erickson, A.L.; Friedlander, A.M.; Graham, N.A.J.; Gove, J.M.; et al. Parsing human and biophysical drivers of coral reef regimes. Proc. R. Soc. B Boil. Sci. 2019, 286, 20182544. [Google Scholar] [CrossRef]
- Kuffner, I.B.; Walters, L.J.; Becerro, M.A.; Paul, V.J.; Ritson-Williams, R.; Beach, K.S. Inhibition of coral recruitment by macroalgae and cyanobacteria. Mar. Ecol. Prog. Ser. 2006, 323, 107–117. [Google Scholar] [CrossRef]
- Mumby, P.J.; Steneck, R.S.; Adjeroud, M.; Arnold, S.N. High resilience masks underlying sensitivity to algal phase shifts of Pacific coral reefs. Oikos 2016, 125, 644–655. [Google Scholar] [CrossRef]
- Box, S.; Mumby, P. Effect of macroalgal competition on growth and survival of juvenile Caribbean corals. Mar. Ecol. Prog. Ser. 2007, 342, 139–149. [Google Scholar] [CrossRef]
- Lirman, D. Competition between macroalgae and corals: Effects of herbivore exclusion and increased algal biomass on coral survivorship and growth. Coral Reefs 2001, 19, 392–399. [Google Scholar] [CrossRef]
- Jompa, J.; McCook, L.J. Effects of competition and herbivory on interactions between a hard coral and a brown alga. J. Exp. Mar. Biol. Ecol. 2002, 271, 25–39. [Google Scholar] [CrossRef]
- Vieira, C.; Payri, C.; De Clerck, O. Overgrowth and killing of corals by the brown alga L obophora hederacea (Dictyotales, Phaeophyceae) on healthy reefs in New Caledonia: A new case of the epizoism syndrome. Psychol. Res. 2015, 63, 152–153. [Google Scholar]
- Birrell, C.L.; McCook, L.J.; Willis, B.L. Effects of algal turfs and sediment on coral settlement. Mar. Pollut. Bull. 2005, 51, 408–414. [Google Scholar] [CrossRef]
- Arnold, S.N.; Steneck, R.S.; Mumby, P.J. Running the gauntlet: Inhibitory effects of algal turfs on the processes of coral recruitment. Mar. Ecol. Prog. Ser. 2010, 414, 91–105. [Google Scholar] [CrossRef]
- Hughes, T.P.; Baird, A.H.; Bellwood, D.R.; Card, M.; Connolly, S.R.; Folke, C.; Grosberg, R.; Hoegh-Guldberg, O.; Jackson, J.B.C.; Kleypas, J.; et al. Climate Change, Human Impacts, and the Resilience of coral reefs. Science 2003, 301, 929–933. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hughes, T.P.; Hoey, A.S. Sleeping functional group drives coral-reef recovery. Curr. Biol. 2006, 16, 2434–2439. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Simpson, T. Large-scale bleaching of corals on the Great Barrier Reef. Ecology 2018, 99, 501. [Google Scholar] [CrossRef]
- Mantyka, C.S.; Bellwood, D.R. Macroalgal grazing selectivity among herbivorous coral reef fishes. Mar. Ecol. Prog. Ser. 2007, 352, 177–185. [Google Scholar] [CrossRef]
- Bennett, S.; Bellwood, D. Latitudinal variation in macroalgal consumption by fishes on the Great Barrier Reef. Mar. Ecol. Prog. Ser. 2011, 426, 241–252. [Google Scholar] [CrossRef][Green Version]
- Vergés, A.; Bennett, S.; Bellwood, D.R. Diversity among macroalgae-consuming fishes on coral reefs: A transcontinental comparison. PLoS ONE 2012, 7, e45543. [Google Scholar] [CrossRef]
- Puk, L.D.; Ferse, S.C.A.; Wild, C. Patterns and trends in coral reef macroalgae browsing: A review of browsing herbivorous fishes of the Indo-Pacific. Rev. Fish Biol. Fish. 2015, 26, 53–70. [Google Scholar] [CrossRef]
- Lefèvre, C.D.; Bellwood, D.R. Temporal variation in coral reef ecosystem processes: Herbivory of macroalgae by fishes. Mar. Ecol. Prog. Ser. 2011, 422, 239–251. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Streit, R.P.; Brandl, S.J.; Tebbett, S.B. The meaning of the term ‘function’in ecology: A coral reef perspective. Funct. Ecol. 2019, 33, 948–961. [Google Scholar]
- Rasher, D.B.; Hoey, A.S.; Hay, M.E. Consumer diversity interacts with prey defenses to drive ecosystem function. Ecology 2013, 94, 1347–1358. [Google Scholar] [CrossRef]
- Bonaldo, R.M.; Pires, M.M.; Guimarães, P.R.; Hoey, A.S.; Hay, M.E. Small marine protected areas in Fiji provide refuge for reef fish assemblages, feeding groups, and corals. PLoS ONE 2017, 12, e0170638. [Google Scholar] [CrossRef]
- Loffler, Z.; Hoey, A.S. Canopy-forming macroalgal beds (Sargassum) on coral reefs are resilient to physical disturbance. J. Ecol. 2018, 106, 1156–1164. [Google Scholar] [CrossRef]
- Cvitanovic, C.; Bellwood, D.R. Local variation in herbivore feeding activity on an inshore reef of the Great Barrier Reef. Coral Reefs 2008, 28, 127–133. [Google Scholar] [CrossRef]
- Plass-Johnson, J.G.; Ferse, S.C.A.; Jompa, J.; Wild, C.; Teichberg, M. Fish herbivory as key ecological function in a heavily degraded coral reef system. Limnol. Oceanogr. 2015, 60, 1382–1391. [Google Scholar] [CrossRef]
- Paul, V.; Hay, M. Seaweed susceptibility to herbivory: Chemical and morphological correlates. Mar. Ecol. Prog. Ser. 1986, 33, 255–264. [Google Scholar] [CrossRef]
- Choat, J.H.; Robbins, W.D.; Clements, K.D. The trophic status of herbivorous fishes on coral reefs. Mar. Biol. 2004, 145, 445–454. [Google Scholar] [CrossRef]
- Clements, K.D.; Raubenheimer, D.; Choat, J.H. Nutritional ecology of marine herbivorous fishes: Ten years on. Funct. Ecol. 2009, 23, 79–92. [Google Scholar] [CrossRef]
- Fox, R.J.; Bellwood, D.R. Remote video bioassays reveal the potential feeding impact of the rabbitfish Siganus canaliculatus (f: Siganidae) on an inner-shelf reef of the Great Barrier Reef. Coral Reefs 2008, 27, 605–615. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Limited functional redundancy in a high diversity system: Single species dominates key ecological process on coral reefs. Ecosystems 2009, 12, 1316–1328. [Google Scholar] [CrossRef]
- Ford, A.K.; Bejarano, S.; Marshell, A.; Mumby, P.J. Linking the biology and ecology of key herbivorous unicornfish to fisheries management in the Pacific. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 790–805. [Google Scholar] [CrossRef]
- Hoey, A.S.; Brandl, S.J.; Bellwood, D.R. Diet and cross-shelf distribution of rabbitfishes (f. Siganidae) on the northern Great Barrier Reef: Implications for ecosystem function. Coral Reefs 2013, 32, 973–984. [Google Scholar] [CrossRef]
- Loffler, Z.; Bellwood, D.R.; Hoey, A.S. Among-habitat algal selectivity by browsing herbivores on an inshore coral reef. Coral Reefs 2015, 34, 597–605. [Google Scholar] [CrossRef]
- Streit, R.P.; Hoey, A.S.; Bellwood, D.R. Feeding characteristics reveal functional distinctions among browsing herbivorous fishes on coral reefs. Coral Reefs 2015, 34, 1037–1047. [Google Scholar] [CrossRef]
- Kelly, E.L.A.; Eynaud, Y.; Clements, S.M.; Gleason, M.; Sparks, R.T.; Williams, I.D.; Smith, J.E. Investigating functional redundancy versus complementarity in Hawaiian herbivorous coral reef fishes. Oecologia 2016, 182, 1151–1163. [Google Scholar] [CrossRef]
- Brandl, S.J.; Bellwood, D.R. Individual-based analyses reveal limited functional overlap in a coral reef fish community. J. Anim. Ecol. 2014, 83, 661–670. [Google Scholar] [CrossRef]
- Cheevaporn, V.; Menasveta, P. Water pollution and habitat degradation in the Gulf of Thailand. Mar. Pollut. Bull. 2003, 47, 43–51. [Google Scholar] [CrossRef]
- Adger, W.N.; Hughes, T.P.; Folke, C.; Carpenter, S.R.; Rockström, J. Social-ecological resilience to coastal disasters. Science 2005, 309, 1036–1039. [Google Scholar] [CrossRef] [PubMed]
- Stuhldreier, I.; Bastian, P.; Schönig, E.; Wild, C.; Schoenig, E. Effects of simulated eutrophication and overfishing on algae and invertebrate settlement in a coral reef of Koh Phangan, Gulf of Thailand. Mar. Pollut. Bull. 2015, 92, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Buranapratheprat, A.; Bunpapong, M. A two dimensional hydrodynamic model for the Gulf of Thailand. In Proceedings of the IOC/WESTPAC Fourth International Scientific Symposium, Okinawa, Japan, 2 February 1998; Volume 469, p. 478. [Google Scholar]
- Kulbicki, M.; Guillemot, N.; Amand, M. A general approach to length-weight relationships for New Caledonian lagoon fishes. Cybium 2005, 29, 235–252. [Google Scholar]
- Hill, J.; Wilkinson, C.R. Methods for ecological monitoring of coral reefs. In A Resource for Managers, Version 1; Australian Institute of Marine Science: Townsville, Australia, 2004; ISBN 9780642322371. [Google Scholar]
- Low, J.K.Y.; Fong, J.; Todd, P.A.; Chou, L.M.; Bauman, A.G. Seasonal variation of Sargassum ilicifolium (phaeophyceae) growth on equatorial coral reefs. J. Phycol. 2018, 55, 289–296. [Google Scholar] [CrossRef]
- Vieira, C.; D’Hondt, S.; De Clerck, O.; Payri, C.E. Toward an inordinate fondness for stars, beetles and Lobophora? Species diversity of the genus Lobophora (Dictyotales, Phaeophyceae) in New Caledonia. J. Phycol. 2014, 50, 1101–1119. [Google Scholar] [CrossRef] [PubMed]
- Mantyka, C.S.; Bellwood, D.R. Direct evaluation of macroalgal removal by herbivorous coral reef fishes. Coral Reefs 2007, 26, 435–442. [Google Scholar] [CrossRef]
- Davis, A.; Manly, B.; McDonald, L.; Thomas, D. Resource selection by animals: Statistical design and analysis for field studies. J. Anim. Ecol. 1994, 63, 745. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org (accessed on 26 July 2020).
- Strauss, R.E. Reliability estimates for Ivlev’s electivity index, the forage ratio, and a proposed linear index of food selection. Trans. Am. Fisheries Soc. 1979, 108, 344–352. [Google Scholar] [CrossRef]
- Lechowicz, M.J. The sampling characteristics of electivity indices. Oecologia 1982, 52, 22–30. [Google Scholar] [CrossRef]
- Roa, R. Design and analysis of multiple-choice feeding-preference experiments. Oecologia 1992, 89, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, J.R., III. On the statistical analysis of multiple-choice feeding preference experiments. Oecologia 1998, 116, 475–481. [Google Scholar] [CrossRef]
- Hollander, M.; Wolfe, D.A.; Chicken, E. Nonparametric Statistical Methods, 3rd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; ISBN 0470387378. [Google Scholar]
- Roff, G.; Mumby, P.J. Global disparity in the resilience of coral reefs. Trends Ecol. Evol. 2012, 27, 404–413. [Google Scholar] [CrossRef]
- Edwards, C.B.; Friedlander, A.M.; Green, A.G.; Hardt, M.J.; Sala, E.; Sweatman, H.P.; Williams, I.D.; Zgliczynski, B.; Sandin, S.A.; Smith, J.E. Global assessment of the status of coral reef herbivorous fishes: Evidence for fishing effects. Proc. R. Soc. B Boil. Sci. 2014, 281, 20131835. [Google Scholar] [CrossRef]
- Chong-Seng, K.M.; Nash, K.L.; Bellwood, D.R.; Graham, N.A.J. Macroalgal herbivory on recovering versus degrading coral reefs. Coral Reefs 2014, 33, 409–419. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Suppression of herbivory by macroalgal density: A critical feedback on coral reefs? Ecol. Lett. 2011, 14, 267–273. [Google Scholar] [CrossRef]
- Madin, E.M.P.; Gaines, S.D.; Warner, R.R. Field evidence for pervasive indirect effects of fishing on prey foraging behavior. Ecology 2010, 91, 3563–3571. [Google Scholar] [CrossRef]
- Fox, R.J.; Bellwood, D.R. Niche partitioning of feeding microhabitats produces a unique function for herbivorous rabbitfishes (Perciformes, Siganidae) on coral reefs. Coral Reefs 2012, 32, 13–23. [Google Scholar] [CrossRef]
- Brandl, S.J.; Bellwood, D.R. Microtopographic refuges shape consumer-producer dynamics by mediating consumer functional diversity. Oecologia 2016, 182, 203–217. [Google Scholar] [CrossRef]
- Fox, R.J.; Bellwood, D.R. Quantifying herbivory across a coral reef depth gradient. Mar. Ecol. Prog. Ser. 2007, 339, 49–59. [Google Scholar] [CrossRef]
- Russ, G. Distribution and abundance of herbivorous grazing fishes in the central Great Barrier Reef. I. Levels of variability across the entire continental shelf. Mar. Ecol. Prog. Ser. 1984, 20, 23–34. [Google Scholar] [CrossRef]
- Wismer, S.; Hoey, A.; Bellwood, D.R. Cross-shelf benthic community structure on the Great Barrier Reef: Relationships between macroalgal cover and herbivore biomass. Mar. Ecol. Prog. Ser. 2009, 376, 45–54. [Google Scholar] [CrossRef]
- Choat, J.H.; Bellwood, D.R. Interactions amongst herbivorous fishes on a coral reef: Influence of spatial variation. Mar. Biol. 1985, 89, 221–234. [Google Scholar] [CrossRef]
- Klumpp, D.; McKinnon, A. Temporal and spatial patterns in primary production of a coral-reef epilithic algal community. J. Exp. Mar. Biol. Ecol. 1989, 131, 1–22. [Google Scholar] [CrossRef]
- Russ, G.R. Grazer biomass correlates more strongly with production than with biomass of algal turfs on a coral reef. Coral Reefs 2003, 22, 63–67. [Google Scholar] [CrossRef]
- Mumby, P.J.; Harborne, A.R.; Williams, J.; Kappel, C.V.; Brumbaugh, D.R.; Micheli, F.; Holmes, K.E.; Dahlgren, C.P.; Paris, C.B.; Blackwell, P.G. Trophic cascade facilitates coral recruitment in a marine reserve. Proc. Natl. Acad. Sci. USA 2007, 104, 8362–8367. [Google Scholar] [CrossRef] [PubMed]
- Bauman, A.G.; Seah, J.C.L.; Januchowski-Hartley, F.A.; Hoey, A.S.; Fong, J.; Todd, P.A. Fear effects associated with predator presence and habitat structure interact to alter herbivory on coral reefs. Biol. Lett. 2019, 15, 20190409. [Google Scholar] [CrossRef] [PubMed]
- Fong, C.R.; Sura, S.A.; Ford, A.T.; Howard, H.B.; Molina, N.E.; Smith, N.N.; Fong, P. Testing the conceptual and operational underpinnings of field herbivory assays: Does variation in predictability of resources, assay design, and deployment method affect outcomes? J. Exp. Mar. Biol. Ecol. 2020, 533, 151469. [Google Scholar] [CrossRef]
- Steinberg, P.; Paul, V. Fish feeding and chemical defenses of tropical brown algae in Western Australia. Mar. Ecol. Prog. Ser. 1989, 58, 253–259. [Google Scholar] [CrossRef]
- Stiger, V.; Deslandes, E.; Payri, C.E. Phenolic contents of two brown algae, Turbinaria ornata and Sargassum mangarevense on Tahiti (French Polynesia): Interspecific, ontogenic and spatio-temporal variations. Bot. Mar. 2004, 47. [Google Scholar] [CrossRef]
- Bittick, S.J.; Bilotti, N.D.; Peterson, H.A.; Stewart, H.L. Turbinaria ornata as an herbivory refuge for associate algae. Mar. Biol. 2009, 157, 317–323. [Google Scholar] [CrossRef]
- Prathep, A.; Wichachucherd, B.; Thongroy, P. Spatial and temporal variation in density and thallus morphology of Turbinaria ornata in Thailand. Aquat. Bot. 2007, 86, 132–138. [Google Scholar] [CrossRef]
- Seah, J.C.L.; Bauman, A.G.; Todd, P.A. Temporal variation in macroalgal removal: Insights from an impacted equatorial coral reef system. Mar. Biol. 2021, 168, 1–12. [Google Scholar] [CrossRef]
- Clements, K.D.; German, D.P.; Piché, J.; Tribollet, A.; Choat, J.H. Integrating ecological roles and trophic diversification on coral reefs: Multiple lines of evidence identify parrotfishes as microphages. Biol. J. Linn. Soc. 2016. [Google Scholar] [CrossRef]
- Perpetua, M.D.; Gorospe, J.G.; Torres, M.A.J.; Demayo, C.G. Diet composition based on stomach content of the Streaked spinefoot (Siganus javus) from three coastal bays in Mindanao, Philippines. Adv. Environ. Sci. 2013, 5, 49–61. [Google Scholar]
- Miyake, S.; Ngugi, D.K.; Stingl, U. Diet strongly influences the gut microbiota of surgeonfishes. Mol. Ecol. 2015, 24, 656–672. [Google Scholar] [CrossRef]
- Ebrahim, A.; Martin, T.S.H.; Mumby, P.J.; Olds, A.D.; Tibbetts, I.R. Differences in diet and foraging behaviour of commercially important rabbitfish species on coral reefs in the Indian Ocean. Coral Reefs 2020, 39, 977–988. [Google Scholar] [CrossRef]
- Nanami, A. Spatial distributions, feeding ecologies, and behavioral interactions of four rabbitfish species (Siganus unimaculatus, S. virgatus, S. corallinus, and S. puellus). PeerJ 2018, 6, e6145. [Google Scholar] [CrossRef]
- Bauman, A.G.; Hoey, A.S.; Dunshea, G.; Feary, D.A.; Low, J.; Todd, P.A. Macroalgal browsing on a heavily degraded, urbanized equatorial reef system. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kuriiwa, K.; Hanzawa, N.; Yoshino, T.; Kimura, S.; Nishida, M. Phylogenetic relationships and natural hybridization in rabbitfishes (Teleostei: Siganidae) inferred from mitochondrial and nuclear DNA analyses. Mol. Phylogenet. Evol. 2007, 45, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Fox, R.J.; Sunderland, T.L.; Hoey, A.; Bellwood, D.R. Estimating ecosystem function: Contrasting roles of closely related herbivorous rabbitfishes (Siganidae) on coral reefs. Mar. Ecol. Prog. Ser. 2009, 385, 261–269. [Google Scholar] [CrossRef]
- Cheal, A.; Emslie, M.; Miller, I.; Sweatman, H. The distribution of herbivorous fishes on the Great Barrier Reef. Mar. Biol. 2012, 159, 1143–1154. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Choat, J.H. A functional analysis of grazing in parrotfishes (family Scaridae): The ecological implications. Environ. Boil. Fishes 1990, 28, 189–214. [Google Scholar] [CrossRef]
- Brandl, S.J.; Rasher, D.B.; Côté, I.M.; Casey, J.M.; Darling, E.S.; Lefcheck, J.S.; Duffy, J.E. Coral reef ecosystem functioning: Eight core processes and the role of biodiversity. Front. Ecol. Environ. 2019, 17, 445–454. [Google Scholar] [CrossRef]
- Fong, C.R.; Chancellor, K.S.; Renzi, J.J.; Robinson, D.R.; Barber, P.H.; Habtes, S.Y.; Fong, P. Epibionts on Turbinaria ornata, a secondary foundational macroalga on coral reefs, provide diverse trophic support to fishes. Mar. Environ. Res. 2018, 141, 39–43. [Google Scholar] [CrossRef]
- Clements, K.D.; Choat, J.H. Comparison of herbivory in the closely-related marine fish genera Girella and Kyphosus. Mar. Biol. 1997, 127, 579–586. [Google Scholar] [CrossRef]
- Puk, L.; Cernohorsky, N.; Marshell, A.; Dwyer, J.; Wolfe, K.; Mumby, P. Species-specific effects of herbivorous fishes on the establishment of the macroalga Lobophora on coral reefs. Mar. Ecol. Prog. Ser. 2020, 637, 1–14. [Google Scholar] [CrossRef]
- Fulton, C.J.; Abesamis, R.A.; Berkström, C.; Depczynski, M.; Graham, N.A.; Holmes, T.H.; Tinkler, P. Form and function of tropical macroalgal reefs in the Anthropocene. Funct. Ecol. 2019, 33, 989–999. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).