
Sequencing of Organellar Genomes of Nowellia curvifolia (Cephaloziaceae Jungermanniales) Revealed the Smallest Plastome with Complete Gene Set and High Intraspecific Variation Suggesting Cryptic Speciation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussion
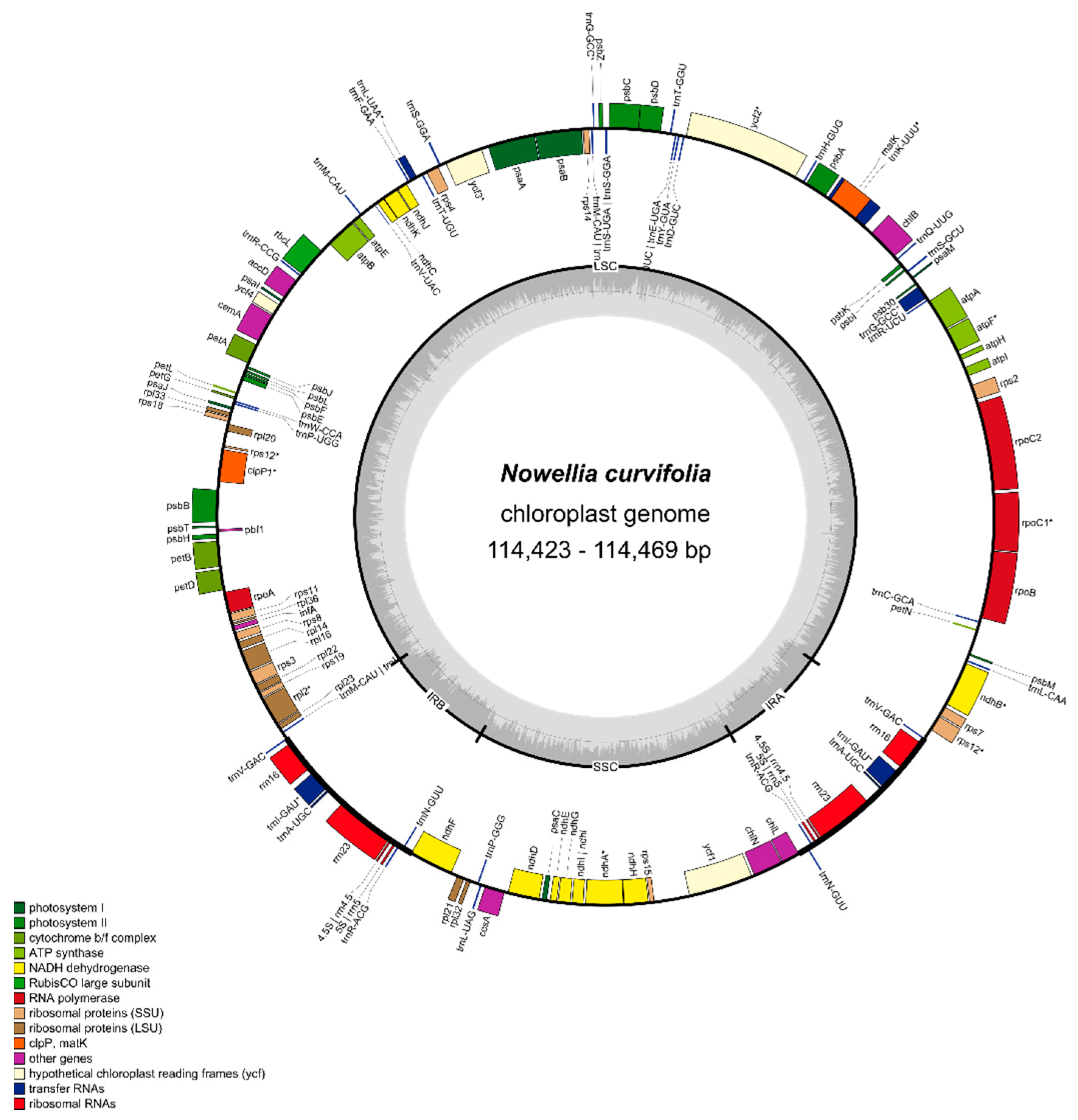
3.1. The Characteristics of Nowellia curvifolia Plastome
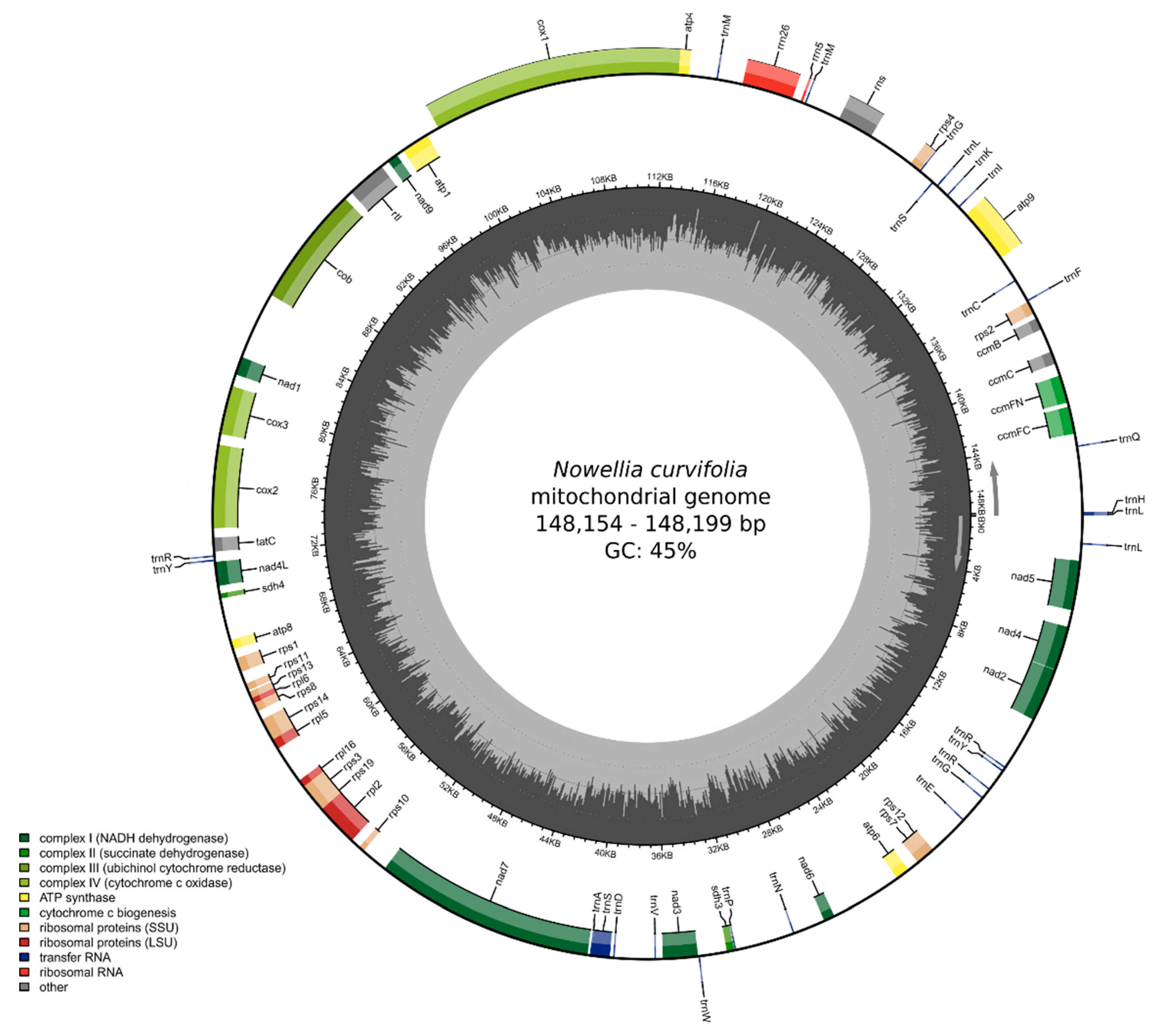
3.2. The Characteristics of Nowellia curvifolia Mitogenome
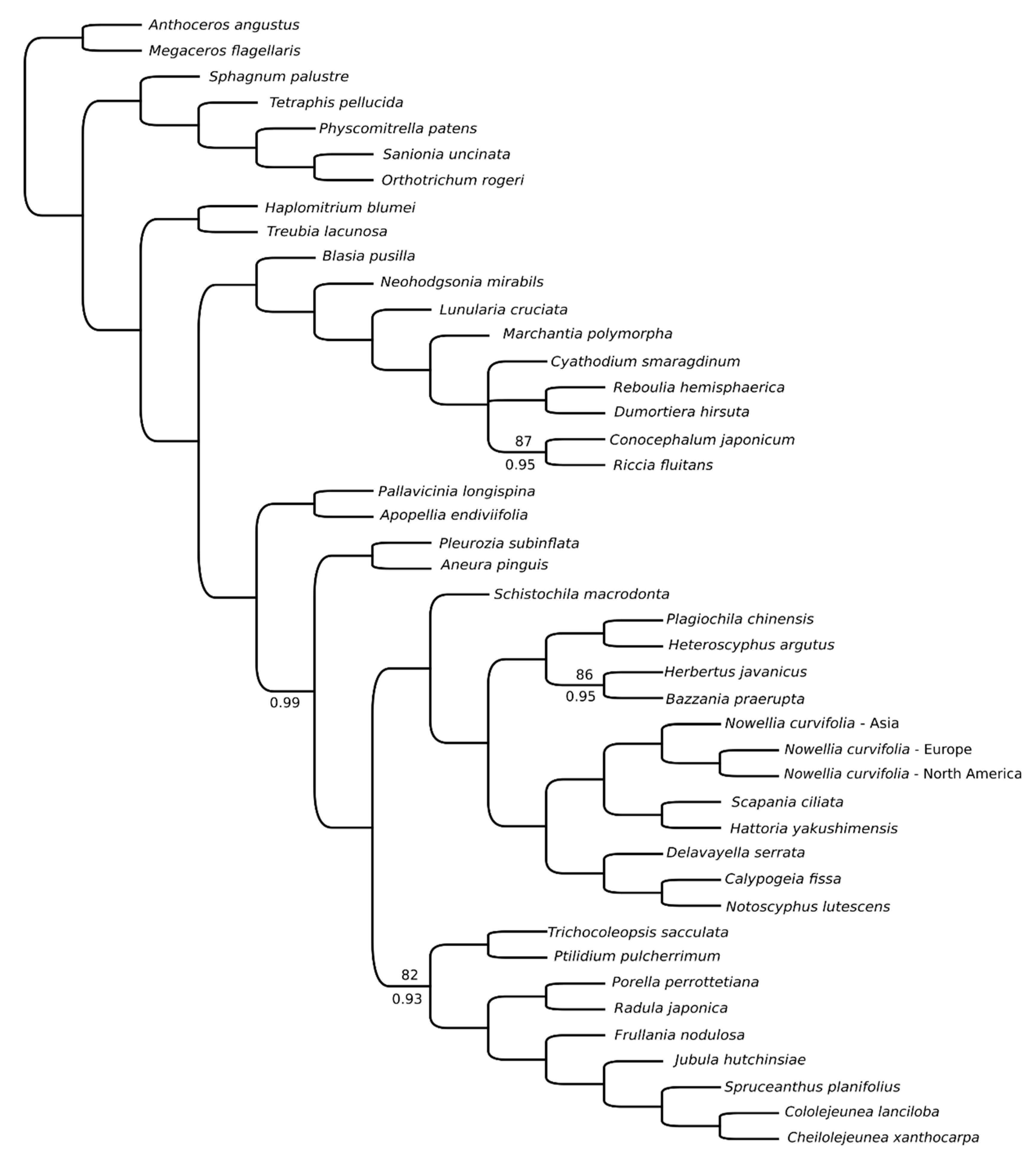
3.3. Intraspecific Variation of Nowellia curvifolia Organellar Genomes
3.4. RNA Editing in Organellar Genomes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Y.; Medina, R.; Goffinet, B. 350 My of Mitochondrial Genome Stasis in Mosses, an Early Land Plant Lineage. Mol. Biol. Evol. 2014, 31, 2586–2591. [Google Scholar] [CrossRef]
- Myszczyński, K.; Ślipiko, M.; Sawicki, J. Potential of Transcript Editing Across Mitogenomes of Early Land Plants Shows Novel and Familiar Trends. Int. J. Mol. Sci. 2019, 20, 2963. [Google Scholar] [CrossRef]
- Dong, S.; Zhao, C.; Zhang, S.; Zhang, L.; Wu, H.; Liu, H.; Zhu, R.; Jia, Y.; Goffinet, B.; Liu, Y. Mitochondrial genomes of the early land plant lineage liverworts (Marchantiophyta): Conserved genome structure, and ongoing low frequency recombination. BMC Genom. 2019, 20, 1–14. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, H.; Yang, J.; Ma, W.; Pressel, S.; Wu, Y.; Schneider, H. Exploring the plastid genome disparity of liverworts. J. Syst. Evol. 2019, 57, 382–394. [Google Scholar] [CrossRef]
- Ślipiko, M.; Myszczyński, K.; Buczkowska, K.; Bączkiewicz, A.; Szczecińska, M.; Sawicki, J. Molecular delimitation of European leafy liverworts of the genus Calypogeia based on plastid super-barcodes. BMC Plant Biol. 2020, 20, 243. [Google Scholar] [CrossRef]
- Ohyama, K.; Fukuzawa, H.; Kohchi, T.; Sano, T.; Sano, S.; Shirai, H.; Umesono, K.; Shiki, Y.; Takeuchi, M.; Aota, Z.C.S.-I.; et al. Structure and organization of Marchantia polymorpha chloroplast genome. J. Mol. Biol. 1988, 203, 281–298. [Google Scholar] [CrossRef]
- Oda, K.; Yamato, K.; Ohta, E.; Nakamura, Y.; Takemura, M.; Nozato, N.; Akashi, K.; Kanegae, T.; Ogura, Y.; Kohchi, T.; et al. Gene organization deduced from the complete sequence of liverwort Marchantia polymorpha mitochondrial DNA. J. Mol. Biol. 1992, 223, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wickett, N.J.; Zhang, Y.; Hansen, S.K.; Roper, J.M.; Kuehl, J.V.; Plock, S.A.; Wolf, P.G.; Depamphilis, C.W.; Boore, J.L.; Goffinet, B. Functional Gene Losses Occur with Minimal Size Reduction in the Plastid Genome of the Parasitic Liverwort Aneura mirabilis. Mol. Biol. Evol. 2008, 25, 393–401. [Google Scholar] [CrossRef]
- Sawicki, J.; Bączkiewicz, A.; Buczkowska, K.; Górski, P.; Krawczyk, K.; Mizia, P.; Myszczyński, K.; Ślipiko, M.; Szczecińska, M. The Increase of Simple Sequence Repeats during Diversification of Marchantiidae, An Early Land Plant Lineage, Leads to the First Known Expansion of Inverted Repeats in the Evolutionarily-Stable Structure of Liverwort Plastomes. Genes 2020, 11, 299. [Google Scholar] [CrossRef] [PubMed]
- Myszczyński, K.; Górski, P.; Ślipiko, M.; Sawicki, J. Sequencing of organellar genomes of Gymnomitrion concinnatum (Jungermanniales) revealed the first exception in the structure and gene order of evolutionary stable liverworts mitogenomes. BMC Plant Biol. 2018, 18, 321. [Google Scholar] [CrossRef] [PubMed]
- Ślipiko, M.; Myszczyński, K.; Buczkowska-Chmielewska, K.; Bączkiewicz, A.; Szczecińska, M.; Sawicki, J. Comparative Analysis of Four Calypogeia Species Revealed Unexpected Change in Evolutionarily-Stable Liverwort Mitogenomes. Genes 2017, 8, 395. [Google Scholar] [CrossRef]
- Liu, Y.; Xue, J.-Y.; Wang, B.; Li, L.; Qiu, Y.-L. The Mitochondrial Genomes of the Early Land Plants Treubia lacunosa and Anomodon rugelii: Dynamic and Conservative Evolution. PLoS ONE 2011, 6, e25836. [Google Scholar] [CrossRef]
- Feldberg, K.; Váňa, J.; Krusche, J.; Kretschmann, J.; Patzak, S.D.F.; Pérez-Escobar, O.A.; Rudolf, N.R.; Seefelder, N.; Schäfer-Verwimp, A.; Long, D.G.; et al. A phylogeny of Cephaloziaceae (Jungermanniopsida) based on nuclear and chloroplast DNA markers. Org. Divers. Evol. 2016, 16, 727–742. [Google Scholar] [CrossRef]
- Lincoln, M. Liverworts of New England; Memoirs of the New York Botanical Garden; Botanical Garden Press: New York, NY, USA, 2008; Volume 99. [Google Scholar]
- Szweykowski, J.; Buczkowska, K.; Odrzykoski, I.J. Conocephalum salebrosum (Marchantiopsida, Conocephalaceae)—A new Holarctic liverwort species. Plant Syst. Evol. 2005, 253, 133–158. [Google Scholar] [CrossRef]
- Buczkowska, K.; Bakalin, V.; Bączkiewicz, A.; Aguero, B.; Gonera, P.; Ślipiko, M.; Szczecińska, M.; Sawicki, J. Does Calypogeia azurea (Calypogeiaceae, Marchantiophyta) occur outside Europe? Molecular and morphological evidence. PLoS ONE 2018, 13, e0204561. [Google Scholar] [CrossRef]
- Heinrichs, J.; Hentschel, J.; Bombosch, A.; Fiebig, A.; Reise, J.; Edelmann, M.; Kreier, H.-P.; Schäfer-Verwimp, A.; Caspari, S.; Schmidt, A.R.; et al. One species or at least eight? Delimitation and distribution of Frullania tamarisci (L.) Dumort. s. l. (Jungermanniopsida, Porellales) inferred from nuclear and chloroplast DNA markers. Mol. Phylogenetics Evol. 2010, 56, 1105–1114. [Google Scholar] [CrossRef]
- Myszczyński, K.; Bączkiewicz, A.; Buczkowska, K.; Ślipiko, M.; Szczecińska, M.; Sawicki, J. The extraordinary variation of the organellar genomes of the Aneura pinguis revealed advanced cryptic speciation of the early land plants. Sci. Rep. 2017, 7, 9804. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty:de novoassembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Vand accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [PubMed]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [PubMed]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Yang, J.; Ma, W.; Pressel, S.; Liu, H.; Wu, Y.; Schneider, H. Chloroplast phylogenomics of liverworts: A reappraisal of the backbone phylogeny of liverworts with emphasis on Ptilidiales. Cladistics 2020, 36, 184–193. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Lenz, H.; Knoop, V. PREPACT 2.0: Predicting C-to-U and U-to-C RNA Editing in Organelle Genome Sequences with Multiple References and Curated RNA Editing Annotation. Bioinform. Biol. Insights 2013, 7. [Google Scholar] [CrossRef]
- Forrest, L.L.; Wickett, N.J.; Cox, C.J.; Goffinet, B. Deep sequencing of Ptilidium (Ptilidiaceae) suggests evolutionary stasis in liverwort plastid genome structure. Plant Ecol. Evol. 2011, 144, 29–43. [Google Scholar] [CrossRef]
- Grosche, C.; Funk, H.T.; Maier, U.G.; Zauner, S. The Chloroplast Genome of Pellia endiviifolia: Gene Content, RNA-Editing Pattern, and the Origin of Chloroplast Editing. Genome Biol. Evol. 2012, 4, 1349–1357. [Google Scholar] [CrossRef]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly Variable Chloroplast Markers for Evaluating Plant Phylogeny at Low Taxonomic Levels and for DNA Barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar] [CrossRef]
- Dong, W.; Xu, C.; Li, C.; Sun, J.; Zuo, Y.; Shi, S.; Cheng, T.; Guo, J.; Zhou, S. ycf1, the most promising plastid DNA barcode of land plants. Sci. Rep. 2015, 5, 8348. [Google Scholar] [CrossRef]
- Szczecińska, M.; Sawicki, J. Genomic Resources of Three Pulsatilla Species Reveal Evolutionary Hotspots, Species-Specific Sites and Variable Plastid Structure in the Family Ranunculaceae. Int. J. Mol. Sci. 2015, 16, 22258–22279. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, Y.; Yang, G.; Peng, J.; Chen, S.; Xu, Z. Determination of the evolutionary pressure on Camellia oleiferaon Hainan Island using the complete chloroplast genome sequence. PeerJ 2019, 7, e7210. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, A.J.C.; Crandall-Stotler, B.J.; Hart, M.L.; Long, D.G.; Forrest, L.L. Divergence times and the evolution of morphological complexity in an early land plant lineage (Marchantiopsida) with a slow molecular rate. New Phytol. 2015, 209, 1734–1746. [Google Scholar] [CrossRef] [PubMed]
- Bączkiewicz, A.; Sawicki, J.; Buczkowska, K.; Polok, K.; Zieliński, R. Application of Different DNA Markers in Studies on Cryptic Species of Aneura Pinguis (Jungermanniopsida, Metzgeriales). Cryptogam. Bryol. 2008, 29, 3–21. [Google Scholar]
- Bączkiewicz, A.; Szczecinska, M.; Sawicki, J.; Stebel, A.; Buczkowska, K. DNA barcoding, ecology and geography of the cryptic species of Aneura pinguis and their relationships with Aneura maxima and Aneura mirabilis (Metzgeriales, Marchantiophyta). PLoS ONE 2017, 12, e0188837. [Google Scholar] [CrossRef] [PubMed]
- Bakalin, V.; Vilnet, A.; Klimova, K.; Nguyen, V.S. Calypogeia vietnamica sp. nov. (Calypogeiaceae, Hepaticae) from North Vietnam and Diversification in Calypogeia Taxa with Blue Oil Bodies. Herzogia 2019, 32, 219–229. [Google Scholar] [CrossRef]
- Feldberg, K.; Groth, H.; Wilson, R.; Heinrichs, J. Cryptic speciation in Herbertus (Herbertaceae, Jungermanniopsida): Range and morphology of Herbertus sendtneri inferred from nrITS sequences. Plant Syst. Evol. 2004, 249, 247–261. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Fedosov, V.E.; Fedorova, A.V.; Nguyen, V.S. Integrative Taxonomic Revision of Marsupella (Gymnomitriaceae, Hepaticae) Reveals Neglected Diversity in Pacific Asia. Cryptogam. Bryol. 2019, 40, 59. [Google Scholar] [CrossRef]
- Fiedorow, P.; Odrzykoski, I.; Szweykowski, J.; Szweykowska-Kulińska, Z. Phylogeny of the European species of the genus Pellia (Hepaticae; Metzgeriales) based on the molecular data from nuclear tRNA Leu CAA intergenic sequences. Gene 2001, 262, 309–315. [Google Scholar] [CrossRef]
- Diatta, J.; Bączkiewicz, A.; Drapikowska, M.; Rodkiewicz, P.; Sawicki, J.; Szczecinńska, M.; Buczkowska, K. Geochemical Al-kalinity and Acidity as Preferential Site-Specific for Three Liverwort Lineages of Aneura Pinguis Cryptic Species A. Sci. Rep. 2021, in press. [Google Scholar]
- Knoop, V. When you can’t trust the DNA: RNA editing changes transcript sequences. Cell. Mol. Life Sci. 2010, 68, 567–586. [Google Scholar] [CrossRef] [PubMed]
- Knie, N.; Grewe, F.; Fischer, S.; Knoop, V. Reverse U-to-C editing exceeds C-to-U RNA editing in some ferns—A monilophyte-wide comparison of chloroplast and mitochondrial RNA editing suggests independent evolution of the two processes in both organelles. BMC Evol. Biol. 2016, 16, 1–12. [Google Scholar] [CrossRef]
- Dong, S.; Zhao, C.; Zhang, S.; Wu, H.; Mu, W.; Wei, T.; Li, N.; Wan, T.; Liu, H.; Cui, J.; et al. The Amount of RNA Editing Sites in Liverwort Organellar Genes Is Correlated with GC Content and Nuclear PPR Protein Diversity. Genome Biol. Evol. 2019, 11, 3233–3239. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawicki, J.; Krawczyk, K.; Ślipiko, M.; Szczecińska, M. Sequencing of Organellar Genomes of Nowellia curvifolia (Cephaloziaceae Jungermanniales) Revealed the Smallest Plastome with Complete Gene Set and High Intraspecific Variation Suggesting Cryptic Speciation. Diversity 2021, 13, 81. https://doi.org/10.3390/d13020081
Sawicki J, Krawczyk K, Ślipiko M, Szczecińska M. Sequencing of Organellar Genomes of Nowellia curvifolia (Cephaloziaceae Jungermanniales) Revealed the Smallest Plastome with Complete Gene Set and High Intraspecific Variation Suggesting Cryptic Speciation. Diversity. 2021; 13(2):81. https://doi.org/10.3390/d13020081
Chicago/Turabian StyleSawicki, Jakub, Katarzyna Krawczyk, Monika Ślipiko, and Monika Szczecińska. 2021. "Sequencing of Organellar Genomes of Nowellia curvifolia (Cephaloziaceae Jungermanniales) Revealed the Smallest Plastome with Complete Gene Set and High Intraspecific Variation Suggesting Cryptic Speciation" Diversity 13, no. 2: 81. https://doi.org/10.3390/d13020081
APA StyleSawicki, J., Krawczyk, K., Ślipiko, M., & Szczecińska, M. (2021). Sequencing of Organellar Genomes of Nowellia curvifolia (Cephaloziaceae Jungermanniales) Revealed the Smallest Plastome with Complete Gene Set and High Intraspecific Variation Suggesting Cryptic Speciation. Diversity, 13(2), 81. https://doi.org/10.3390/d13020081