
Spatial Niche Expansion at Multiple Habitat Scales of a Tropical Freshwater Turtle in the Absence of a Potential Competitor

 ,
,  ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- (1)
- Are the study species co-occurring in a same micro-habitat or do they exhibit apparent differences in habitat utilization?
- (2)
- If the two species differ in their habitat utilization, are these differences found at multiple spatial scales?
- (3)
- Are the eventual habitat differences due, at least in part, to interspecific competition?
- (4)
- If so, do habitat preferences of either of these two species change when allopatric from a potential competitor?
- (5)
- Is the assumption that P. cupulatta habitat use is similar to its sister species (Pelusios niger) confirmed by our field evidence?
2. Materials and Methods
2.1. Study Species
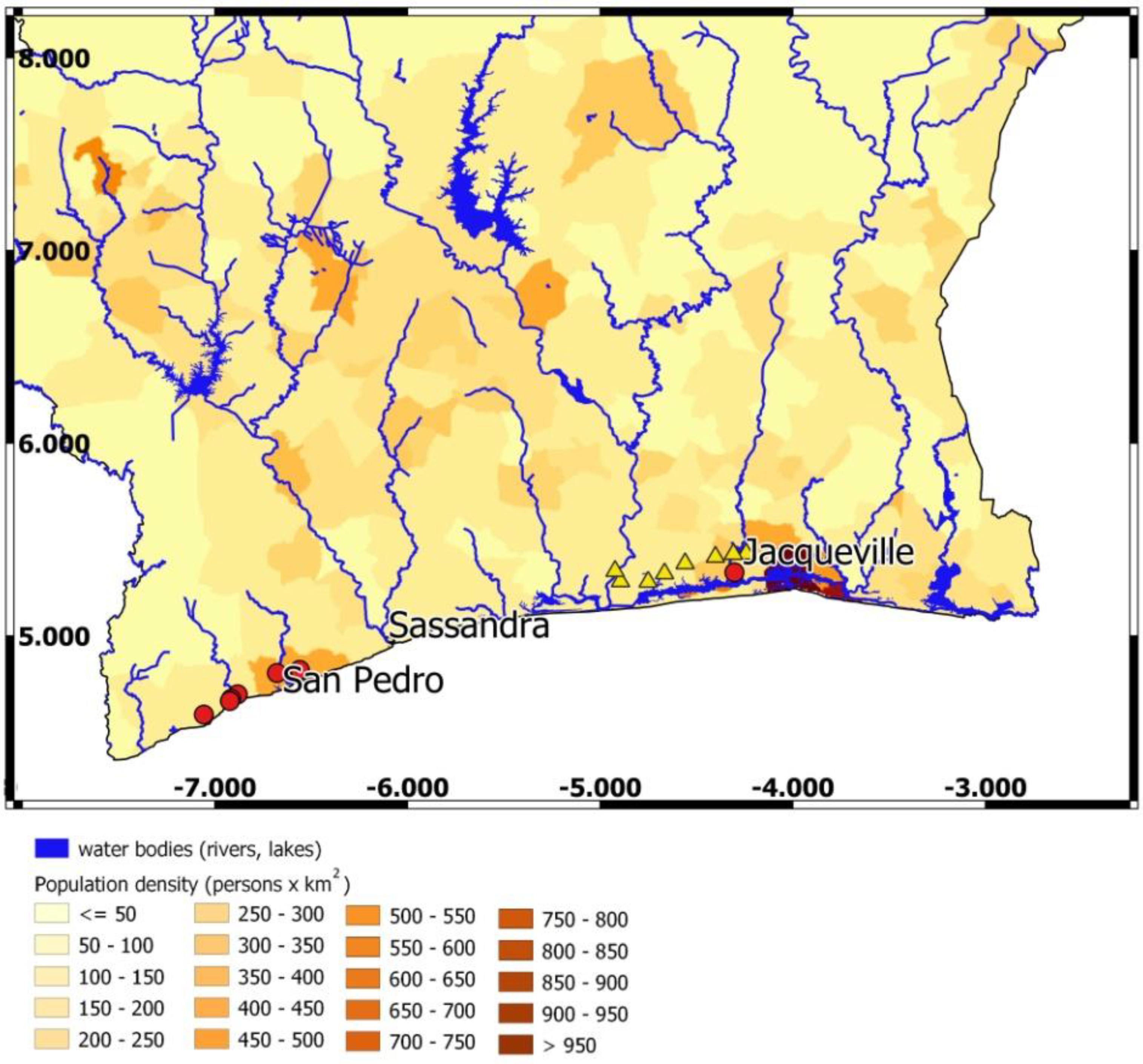
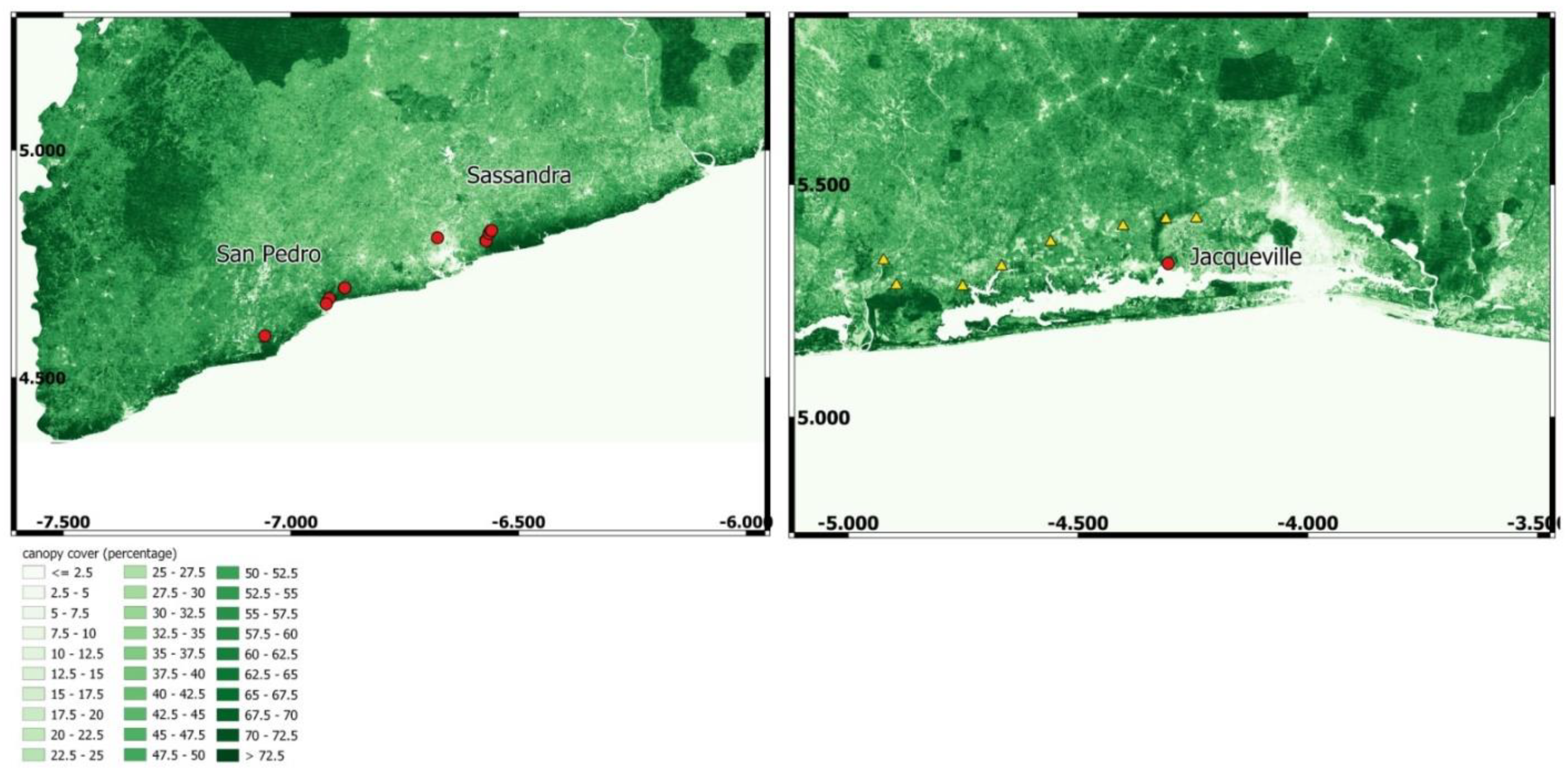
2.2. Study Sites
2.3. Protocol
- (a)
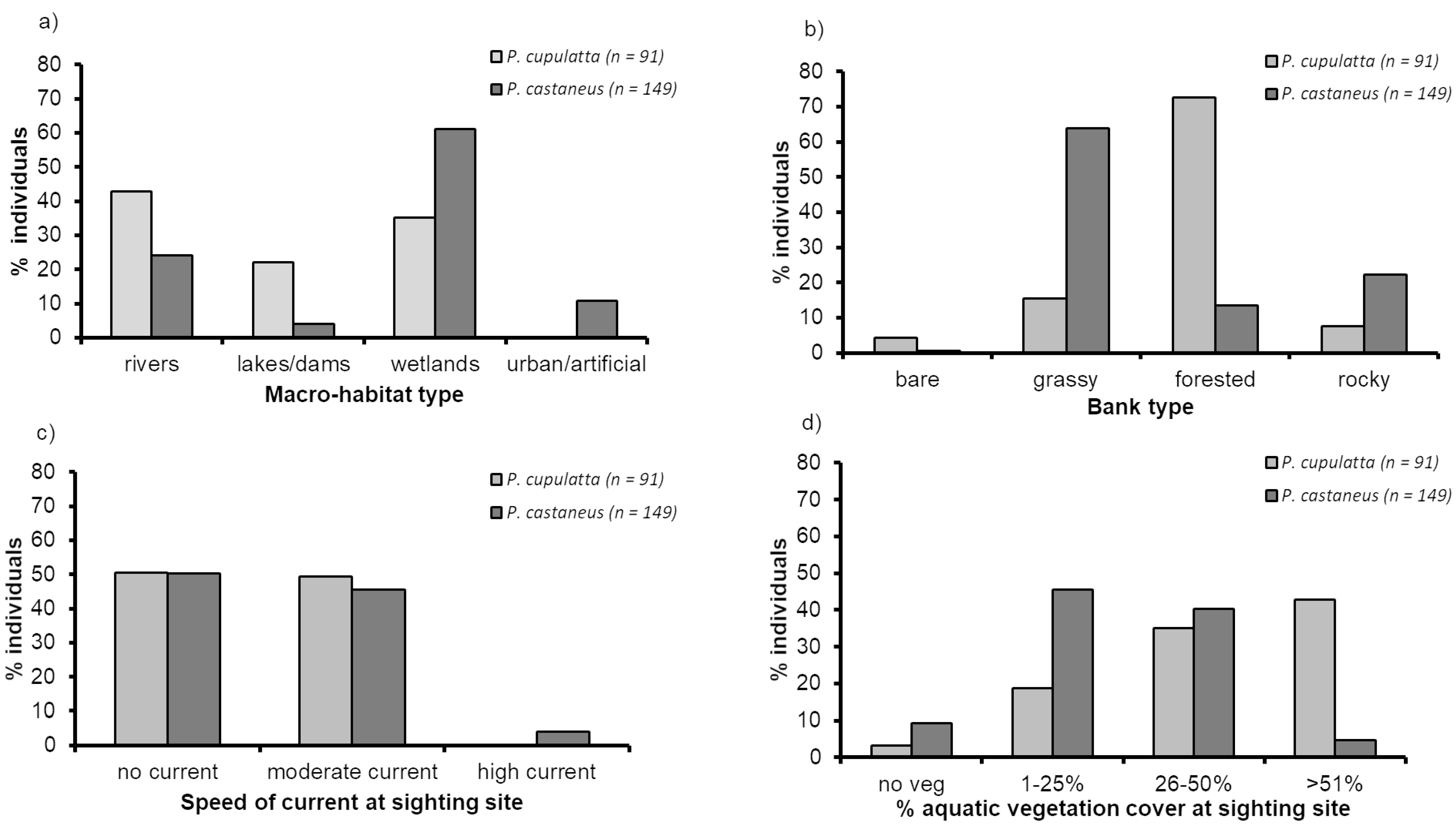
- Water current speed—within a 10 m radius around the sighting point of each individual turtle we evaluated the water current speed by eye in one of three categories: 0 = no current; 1 = moderate current; 2 = high current.
- (b)
- Bank type—we described the type of bank linked to the water body where each individual turtle was sighted as: 0 = no bank vegetation (only organic substratum, bare); 1 = rocky banks (rocky); 2 = bank with mainly herbaceous vegetation (grassy); 3 = forested banks, i.e., when the bank vegetation consisted of a strip of gallery forest.
- (c)
- Aquatic vegetation—classified as: 0 = no aquatic vegetation; 1 = moderate aquatic vegetation (1–25% covered by aquatic plants); 2 = high aquatic vegetation (26–50% cover); 3 = very high aquatic vegetation (>51% cover). The percentage of aquatic vegetation cover was evaluated by eye within a radius of 10 m around the site of sighting of each individual turtle. Since we grouped vegetation in four large categories based on the estimated percentage of vegetation cover, this design would have been problematic in cases where the percentage of vegetative cover was at the limit between two categories (such as 24% in one site and 26% in another site). These sites would have been categorized such that they were as different as two sites that had e.g., 2% and 49% cover. Although these were clearly not equivalent comparisons, we defined the three percentages of vegetative cover categories after having examined the distribution of sample sizes to verify that there were no such “threshold cases” in our samples.
2.4. Statistical Analyses
3. Results
3.1. General Data
3.2. Spatial Niche Characteristics in Sympatric Populations
3.3. Spatial Niche Characteristics of Pelusios castaneus in Allopatry
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bodie, J.R.; Semlitsch, R.D. Spatial and temporal use of floodplain habitats by lentic and lotic species of aquatic turtles. Oecologia 2000, 122, 138–146. [Google Scholar] [CrossRef]
- Bodie, J.R.; Semlitsch, R.D.; Renken, R.B. Diversity and structure of turtle assemblages: Associations with wetland characters across a floodplain landscape. Ecography 2000, 23, 444–456. [Google Scholar] [CrossRef]
- Conner, C.A.; Douthitt, B.A.; Ryan, T.J. Descriptive ecology of a turtle assemblage in an urban landscape. Am. Midl. Nat. 2005, 153, 428–435. [Google Scholar] [CrossRef][Green Version]
- Luiselli, L.; Akani, G.C.; Ajong, S.N.; George, A.; Di Vittorio, M.; Eniang, E.A.; Dendi, D.; Hema, E.M.; Petrozzi, F.; Fa, J.E. Predicting the structure of turtle assemblages along a megatransect in West Africa. Biol. J. Linn. Soc. 2020, 130, 296–309. [Google Scholar] [CrossRef]
- Luiselli, L. Resource partitioning in freshwater turtle communities: A null model meta-analysis of available data. Acta Oecol. 2008, 34, 80–88. [Google Scholar] [CrossRef]
- Lindeman, P.V. Aggressive interactions during basking among four species of emydid turtles. J. Herpetol. 1999, 33, 214–219. [Google Scholar] [CrossRef]
- Cadi, A.; Joly, P. Competition for basking sites between the endangered European pond turtle (Emys orbicularis galloitalica) and the introduced red-eared slider (Trachemys scripta elegans). Can. J. Zool. 2003, 81, 1392–1398. [Google Scholar] [CrossRef]
- Webb, R.G. North American recent soft-shelled turtles (Family Trionychidae). Univ. Kansas Publ. Mus. Nat. Hist. 1962, 13, 426–611. [Google Scholar]
- Auth, D.L. Behavioral ecology of basking in the yellowbellied turtle, Chrysemys scripta scripta (Schoepff). Bull. Florida State Mus. Biol. Sci. 1975, 20, 1–45. [Google Scholar]
- Bjorndal, K.A.; Bolten, A.B. Digestive processing in an herbivorous freshwater turtle: Consequences of small intestine fermentation. Physiol. Zool. 1990, 63, 1232–1247. [Google Scholar]
- Jackson, D.R. Meat on the move: Diet of a predatory turtle, Deirochelys reticularia (Testudines: Emydidae). Chel. Cons. Biol. 1996, 2, 105–107. [Google Scholar]
- Aresco, M.J. Ecological Relationships of Turtles in Northern Florida Lakes: A study of Omnivory and the Structure of a Lake Food Web. Ph.D. Thesis, The Florida State University, Tallahassee, FL, USA, 2005, unpublished. [Google Scholar]
- Luiselli, L.; Akani, G.C. An indirect assessment of the effects of oil pollution on the diversity and functioning of turtle communities in the Niger Delta, Nigeria. Anim. Biodiv. Cons. 2003, 26, 57–65. [Google Scholar]
- Luiselli, L.; Akani, G.C.; Politano, E.; Odegbune, E.; Bello, O. Dietary shifts of sympatric freshwater turtles in pristine and oil-polluted habitats of the Niger Delta, southern Nigeria. Herpetol. J. 2004, 14, 57–64. [Google Scholar]
- Luiselli, L.; Akani, G.C.; Politano, E. Effects of habitat alteration cased by petrochemical activities and oil spills on the habitat use and interspecific relationships among four species of Afrotropical freshwater turtles. Biodiv. Cons. 2006, 15, 3751–3767. [Google Scholar] [CrossRef]
- Akani, G.C.; Eniang, E.A.; Amadi, N.; Dendi, D.; Hema, E.M.; Diagne, T.; Ségniagbeto, G.H.; Di Vittorio, M.; Gbewaa, S.B.; Pawels, O.S.G.; et al. Macrohabitat and microhabitat usage by two softshell turtles (Trionyx triunguis and Cyclanorbis senegalensis) in West and Central Africa. Herp. Cons. Biol. 2018, 13, 642–651. [Google Scholar]
- Stanford, C.B.; Iverson, J.B.; Rhodin, A.G.J.; Van Dijk, P.P.; Mittermeier, R.A.; Kuchling, G.; Berry, K.H.; Bertolero, A.; Blanck, T.E.G.; Bjorndal, K.A.; et al. Turtles and tortoises are in trouble. Curr. Biol. 2020, 30, R721–R735. [Google Scholar] [CrossRef]
- Branch, B. Tortoises, Terrapins and Turtles of Africa; New Holland Publishing: Cape Town, South Africa, 2008. [Google Scholar]
- Bour, R.; Luiselli, L.; Petrozzi, F.; Segniagbeto, G.H.; Chirio, L. Pelusios castaneus (Schweigger 1812)—West African Mud Turtle, Swamp Terrapin. In Conservation Biology of Freshwater Turtles and Tortoises: A Compilation Project of the IUCN/SSC Tortoise and Freshwater Turtle Specialist Group; Rhodin, A.G.J., Pritchard, P.C.H., van Dijk, P.P., Saumure, R.A., Buhlmann, K.A., Iverson, J.B., Mittermeier, R.A., Eds.; Chelonian Research Foundation and Turtle Conservancy: Arlington, VT, USA, 2016; Volume 5, pp. 095.1–095.11. Available online: https://iucn-tftsg.org/wp-content/uploads/file/Accounts/crm_5_095_castaneus_v1_2016.pdf (accessed on 21 December 2020). [CrossRef]
- Ficetola, G.F.; Padoa-Schioppa, E.; Monti, A.; Massa, R.; De Bernardi, F.; Bottoni, L. The importance of aquatic and terrestrial habitat for the European pond turtle (Emys orbicularis): Implications for conservation planning and management. Can. J. Zool. 2004, 82, 1704–1712. [Google Scholar]
- Wyneken, J.; Godfrey, M.H.; Bels, V. Biology of Turtles: From Structures to Strategies of Life; CRC Press: New York, NY, USA, 2008. [Google Scholar]
- Vignoli, L.; Bologna, M.A.; Manzini, S.; Rugiero, L.; Luiselli, L. Attributes of basking sites of the European pond turtle (Emys orbicularis) in central Italy. Amphib. Reptil. 2015, 36, 125–131. [Google Scholar] [CrossRef]
- Case, T.J.; Faaborg, J.; Sidell, R. The role of body size in the assembly of West Indian bird communities. Evolution 1983, 37, 1062–1074. [Google Scholar] [CrossRef]
- Munday, P.L.; Jones, G.P.; Caley, M.J. Interspecific competition and coexistence in a guild of coral-dwelling fishes. Ecology 2001, 82, 2177–2189. [Google Scholar] [CrossRef]
- Pritchard, P.C. Observations on body size, sympatry, and niche divergence in softshell turtles (Trionychidae). Chel. Cons. Biol. 2001, 4, 5–27. [Google Scholar]
- Luiselli, L. Food habits of the pelomedusid turtle Pelusios castaneus castaneus in southeastern Nigeria. Chel. Cons. Biol. 1998, 3, 106–107. [Google Scholar]
- Ude, L.; Luiselli, L.; Angelici, F.M.; Akani, G.C.; Wariboko, S. Seasonal variation in habitat use in sympatric Afrotropical semi-aquatic snakes, Grayia smythii and Afronatrix anoscopus (Colubridae). Amphib. Reptil. 2005, 26, 372–376. [Google Scholar] [CrossRef]
- Connell, J.H. On the prevalence and relative importance of interspecific competition: Evidence from field experiments. Am. Nat. 1983, 122, 661–696. [Google Scholar] [CrossRef]
- Tilman, D. The importance of the mechanisms of interspecific competition. Am. Nat. 1987, 129, 769–774. [Google Scholar] [CrossRef]
- Luiselli, L.; Bour, R.; Petrozzi, F.; Akani, G.C.; Segniagbeto, G.H. Pelusios niger (Duméril and Bibron 1835)—West African Black Mud Turtle. In Conservation Biology of Freshwater Turtles and Tortoises: A Compilation Project of the IUCN/SSC Tortoise and Freshwater Turtle Specialist Group; Rhodin, A.G.J., Iverson, J.B., van Dijk, P.P., Stanford, C.B., Goode, E.V., Buhlmann, K.A., Pritchard, P.C.H., Mittermeier, R.A., Eds.; Chelonian Research Foundation and Turtle Conservancy: Arlington, VT, USA, 2018; Volume 5, pp. 105.1–105.8. Available online: https://iucn-tftsg.org/wp-content/uploads/file/Accounts/crm_5_105_niger_v1_2018.pdf (accessed on 21 December 2020). [CrossRef]
- Tesche, M.R.; Hodges, K.E. Unreliable population inferences from common trapping practices for freshwater turtles. Glob. Ecol. Cons. 2015, 3, 802–813. [Google Scholar] [CrossRef]
- Luiselli, L.; Angelici, F.M.; Politano, E. Ecological correlates of the distribution of terrestrial and freshwater chelonians in the Niger Delta, Nigeria: A biodiversity assessment with conservation implications. Rev. Ecol. (Terre et Vie) 2000, 55, 3–23. [Google Scholar]
- McCoy, C.J.; Flores-Villela, O.A.; Vogt, R.C.; Pappas, M.; McCoy, J.K. Ecology of riverine turtle communities in the Southern United States: Food resource use and trophic niche dimensions. Chel. Cons. Biol. 2020, 19, 197–208. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrozzi, F.; Ajong, S.N.; Pacini, N.; Dendi, D.; Bi, S.G.; Fa, J.E.; Luiselli, L. Spatial Niche Expansion at Multiple Habitat Scales of a Tropical Freshwater Turtle in the Absence of a Potential Competitor. Diversity 2021, 13, 55. https://doi.org/10.3390/d13020055
Petrozzi F, Ajong SN, Pacini N, Dendi D, Bi SG, Fa JE, Luiselli L. Spatial Niche Expansion at Multiple Habitat Scales of a Tropical Freshwater Turtle in the Absence of a Potential Competitor. Diversity. 2021; 13(2):55. https://doi.org/10.3390/d13020055
Chicago/Turabian StylePetrozzi, Fabio, Stephanie N. Ajong, Nic Pacini, Daniele Dendi, Sery Gonedele Bi, Julia E. Fa, and Luca Luiselli. 2021. "Spatial Niche Expansion at Multiple Habitat Scales of a Tropical Freshwater Turtle in the Absence of a Potential Competitor" Diversity 13, no. 2: 55. https://doi.org/10.3390/d13020055
APA StylePetrozzi, F., Ajong, S. N., Pacini, N., Dendi, D., Bi, S. G., Fa, J. E., & Luiselli, L. (2021). Spatial Niche Expansion at Multiple Habitat Scales of a Tropical Freshwater Turtle in the Absence of a Potential Competitor. Diversity, 13(2), 55. https://doi.org/10.3390/d13020055