
How Many Sipunculan Species Are Hiding in Our Oceans?

Abstract
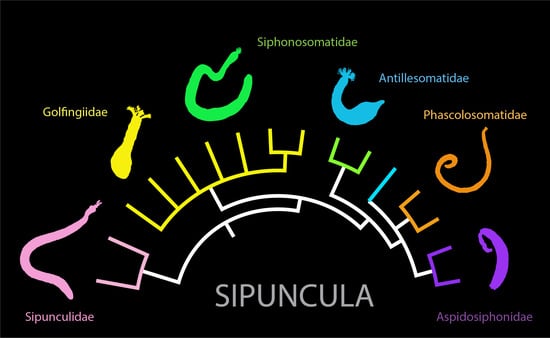
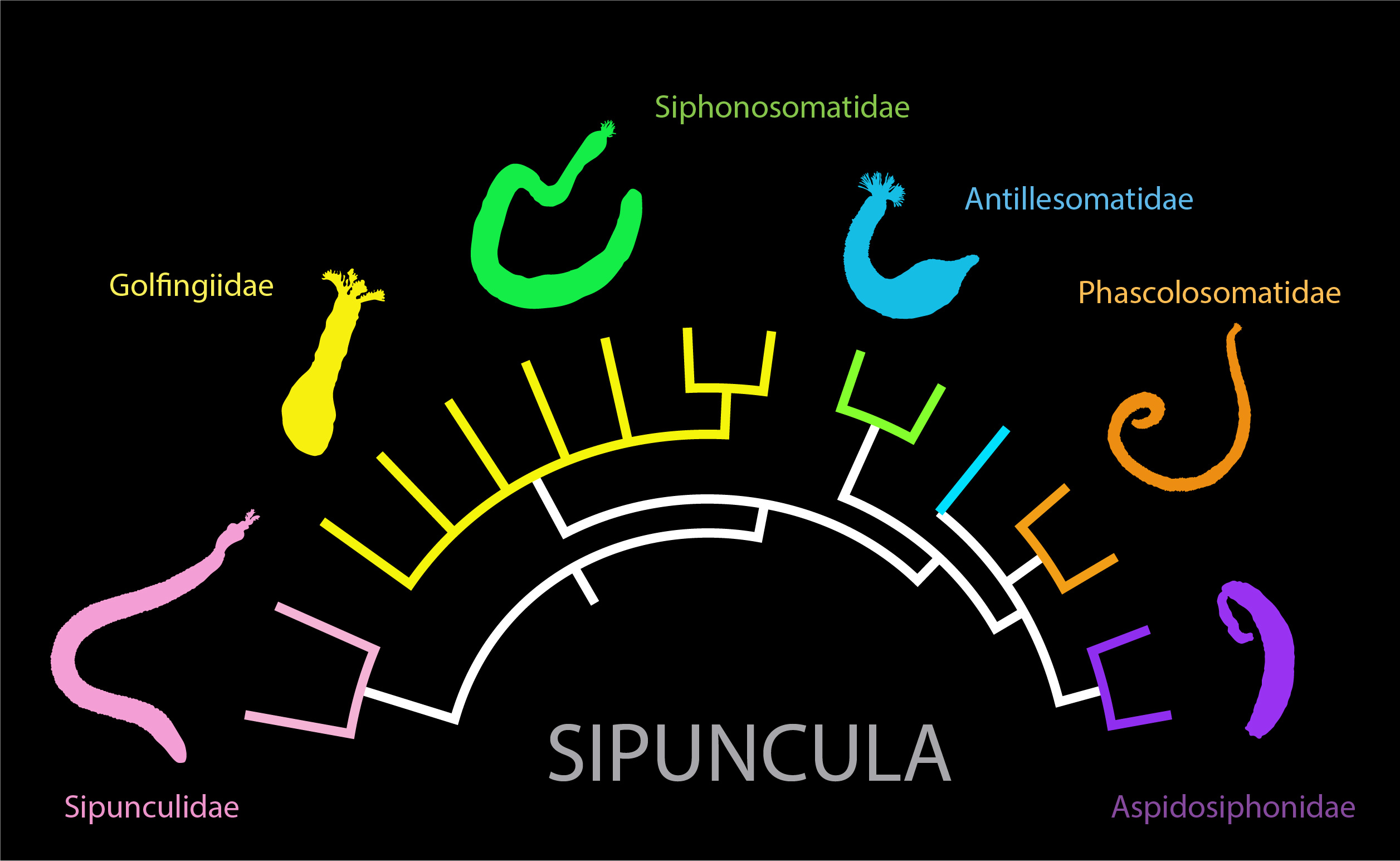{kind=link}
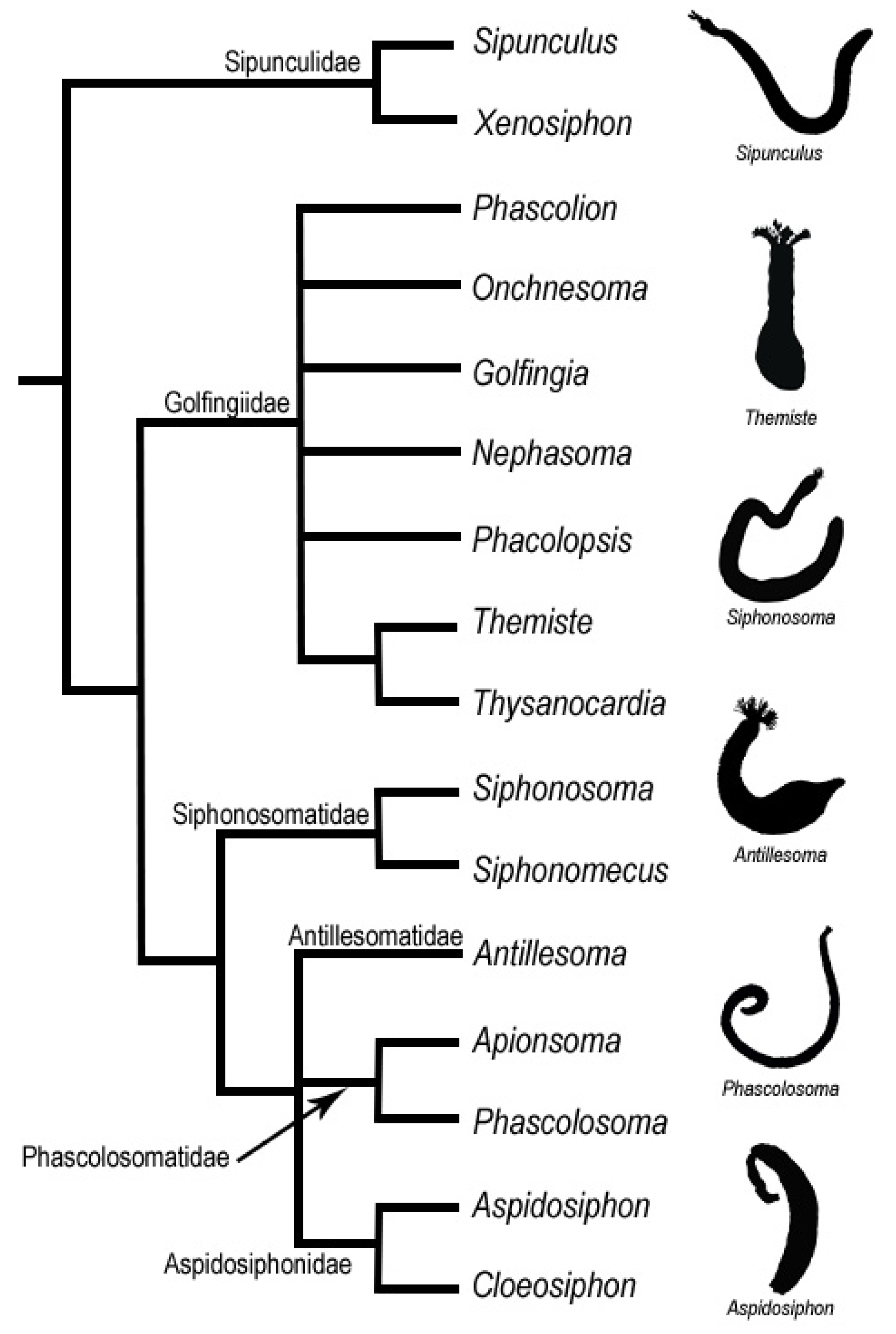{kind=link}
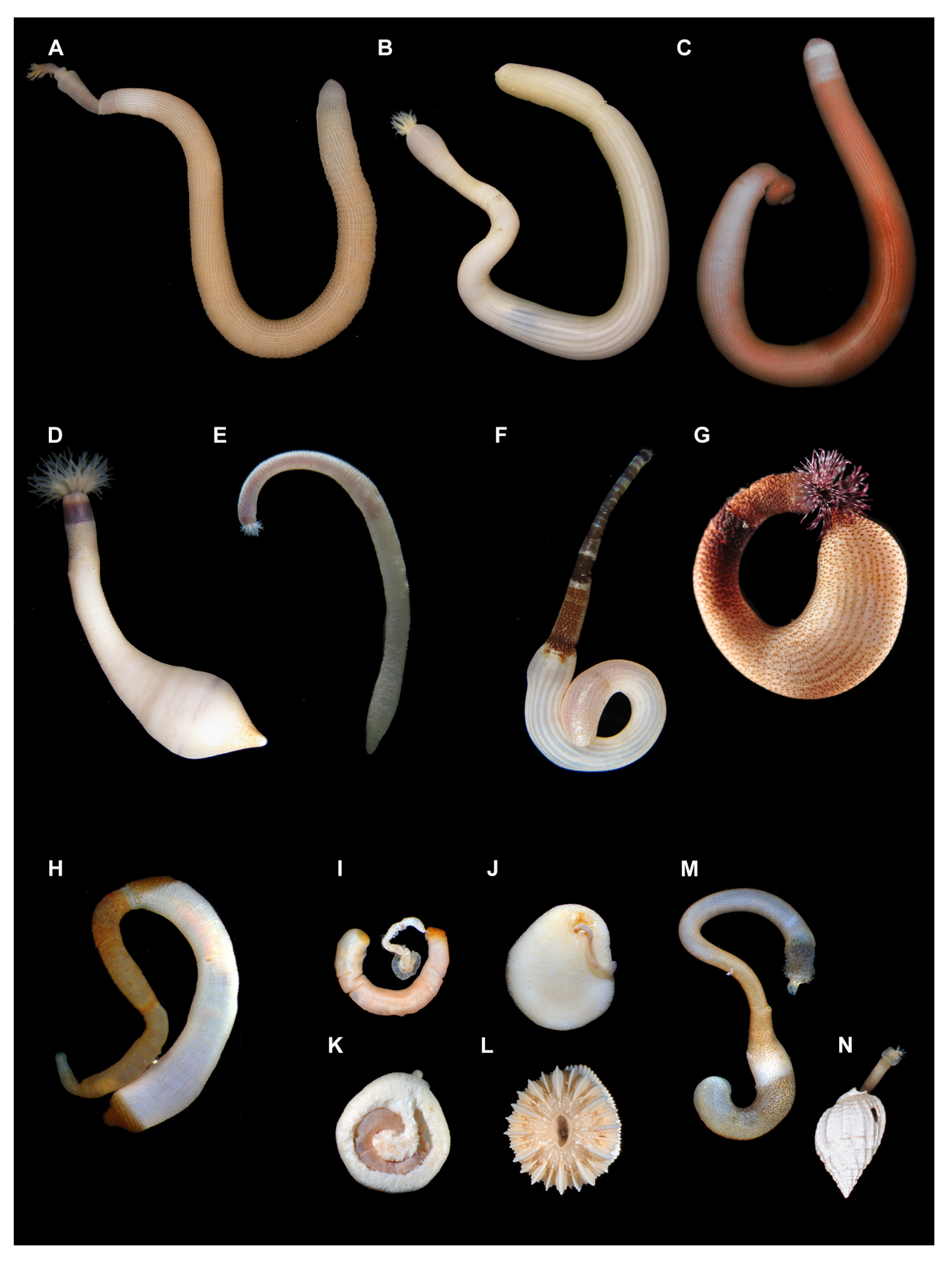{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
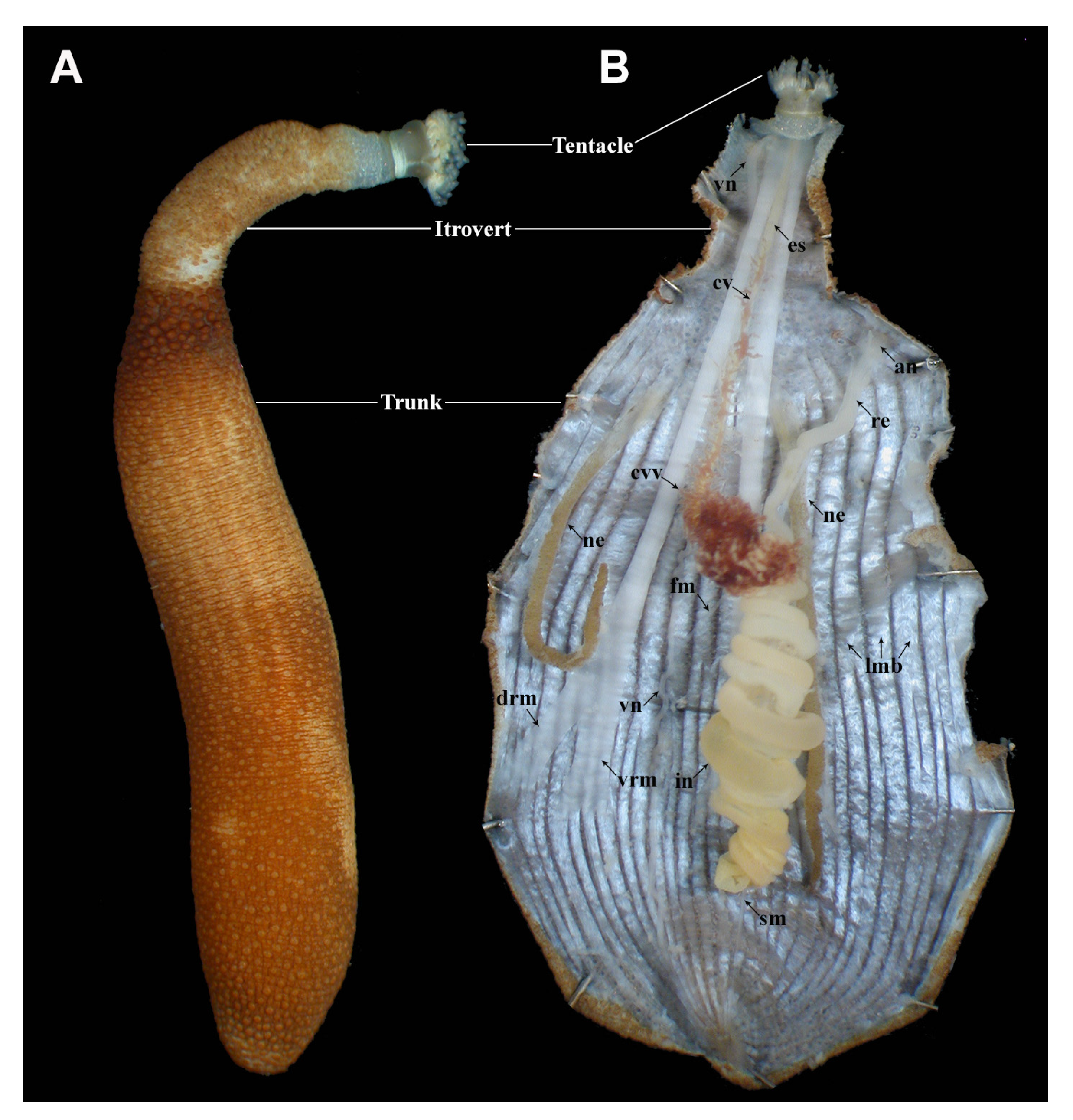
2. External Morphology
3. Anatomy
4. Ecology
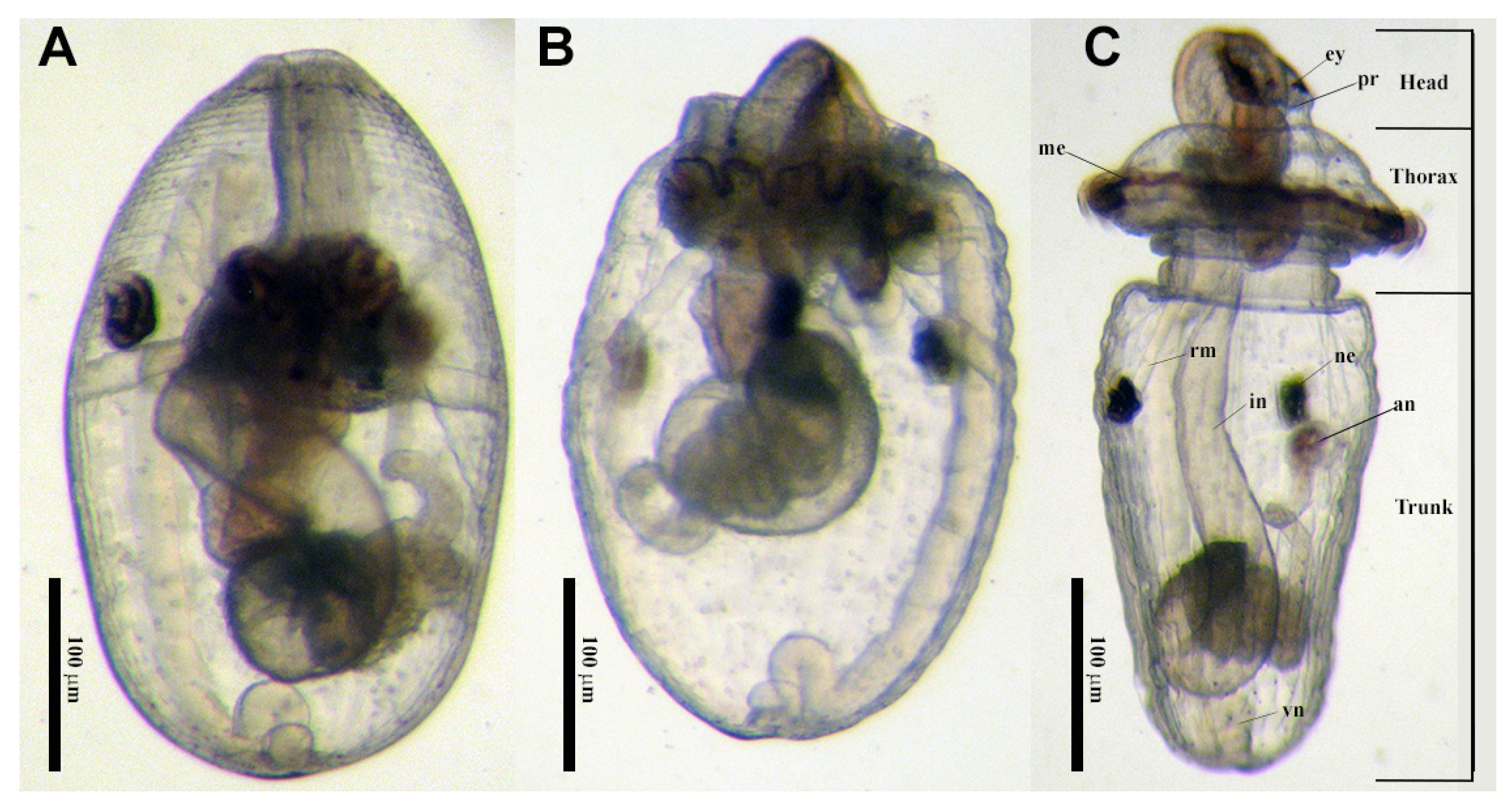
5. Reproduction and Development
6. Species Complexes
6.1. Sipunculus nudus Complex
6.2. Themiste pyroides/hexadactyla Complex
6.3. Thysanocarida nigra/Golfingia pugettensis Complex
6.4. Phascolosoma agassizii Complex
6.5. Phascolosoma nigrescens/varians Complex
6.6. Phascolosoma perlucens Complex
6.7. Apionsoma misakianum Complex
6.8. Antillesoma antillarum Complex
6.9. Aspidosiphon Species Complexes
7. Geographic Coverage
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boore, J.L.; Staton, J.L. The mitochondrial genome of the sipunculid Phascolopsis gouldii supports its association with Annelida rather than Mollusca. Mol. Biol. Evol. 2002, 19, 127–137. [Google Scholar] [CrossRef]
- Telford, M.J.; Wise, M.J.; Gowri-Shankar, V. Consideration of RNA secondary structure siginificantly improves likelihood-based estimates of phylogeny: Examples from the Bilateria. Mol. Biol. Evol. 2005, 22, 1129–1136. [Google Scholar] [CrossRef]
- Dunn, C.W.; Hejnol, A.; Matus, D.Q.; Pang, K.; Browne, W.E.; Smith, S.A.; Seaver, E.; Rouse, G.W.; Obst, M.; Edgecombe, G.D.; et al. Broad phylogenomic sampling improves resolution of the animal tree of life. Nature 2008, 452. [Google Scholar] [CrossRef]
- Mwinyi, A.; Meyer, A.; Bleidorn, C.; Lieb, B.; Bartolomaeus, T.; Podsiadlowski, L. Mitochondrial genome sequence and gene order of Sipunculus nudus give additional support for an inclusion of Sipuncula into Annelida. BMC Genom. 2009, 10, 27. [Google Scholar] [CrossRef]
- Struck, T.H.; Paul, C.; Hill, N.; Hartmann, S.; Hosel, C.; Kube, M.; Lieb, B.; Meyer, A.; Tiedemann, R.; Purschke, G.; et al. Phylogenomic analyses unravel annelid evolution. Nature 2011, 471, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Weigert, A.; Golombek, A.; Gerth, M.; Schwarz, F.; Struck, T.H.; Bleidorn, C. Evolution of mitochondrial gene order in Annelida. Mol. Phylogenet. Evol. 2016, 94, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Weigert, A.; Helm, C.; Meyer, M.; Nickel, B.; Arendt, D.; Hausdorf, B.; Santos, S.R.; Halanych, K.M.; Purschke, G.; Bleidorn, C.; et al. Illuminating the base of the annelid tree using transcriptomics. Mol. Biol. Evol. 2014, 31, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Baltodano, A.M.; Boyle, M.J.; Rice, M.E.; Meyer, N.P. Developmental architecture of the nervous system in Themiste lageniformis (Sipuncula): New evidence from confocal laser scanning microscopy and gene expression. J. Morphol. 2019, 280, 1628–1650. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Chen, J.; Vannier, J.; Saiz Salinas, J.I. Early Cambrian sipunculan worms from southwest China. Proc. R. Soc. Lond. Ser. B 2004, 271, 1671–1676. [Google Scholar] [CrossRef]
- Kristof, A.; Wollesen, T.; Wanninger, A. Segmental mode of neural patterning in Sipuncula. Curr. Biol. 2008, 18, 1129–1132. [Google Scholar] [CrossRef]
- Kristof, A.; Wollesen, T.; Maiorova, A.S.; Wanninger, A. Cellular and muscular growth patterns during sipunculan development. J. Exp. Zool. Part. B Mol. Dev. Evol. 2011, 316B, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Saiz Salinas, J.I. Almost five centuries of systematic study of the enigmatic sipunculan worms. In Proceedings of the Second International Symposium on the Biology of the Sipuncula; Boyle, M.J., Kawauchi, G.Y., Eds.; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2018; pp. 219–235. [Google Scholar]
- De Quatrefages, A. Etudes sur les types inférieurs de l’embranchement des annelés. Mémoire sur l’echiure de Gaertner (Echiurus Gaertnerii NOB). Ann. Sci. Nat. Zool. (Paris) 1847, 7, 307–343. [Google Scholar]
- Hyman, L.H. Phylum Sipunculida. In Smaller Coelomate Groups; McGraw-Hill: New York, NY, USA, 1959; Volume 5, pp. 610–690. [Google Scholar]
- Selenka, E.; de Man, J.G.; Bülow, C. Die Sipunculiden, eine systematische Monographie. Semper Reis. Archipel Phillippinen 2 1883, 4, 1–131. [Google Scholar]
- Fisher, W.K. The sipunculid worms of California and Baja California. Proc. U. S. Natl. Mus. 1952, 102, 371–450. [Google Scholar] [CrossRef]
- Stephen, A.C. Echiura and Sipuncula from the Isreal South Red Sea Expedition. Sea Fish. Res. Stn. Haifa Bull. 1965, 40, 79–83. [Google Scholar]
- Stephen, A.C.; Edmonds, S.J. The Phyla Sipuncula and Echiura; Trustees of the British Museum (Natural History): London, UK, 1972. [Google Scholar]
- Cutler, E.B. The Sipuncula. Their Systematics, Biology and Evolution; Cornell University Press: Ithaca, NY, USA, 1994; p. 453. [Google Scholar]
- Scheltema, R.S.; Hall, J.R. The dispersal of pelagosphera larvae by ocean currents and the geographical distribution of sipunculans. In Proceedings of the International Symposium on the Biology of the Sipuncula and Echiura; Rice, M.E., Todorovic, M., Eds.; Nauçno Delo: Belgrade, Serbia, 1975; pp. 103–116. [Google Scholar]
- Silva-Morales, I. Reinstatement of Phascolosoma (Phascolosoma) varians Keferstein, 1865 (Sipuncula: Phascolosomatidae) based on morphological and molecular data. PeerJ 2020, 8, e10238. [Google Scholar] [CrossRef]
- Kawauchi, G.Y.; Rice, M.E. Two New Species of Nephasoma (Sipuncula: Golfingiidae) from the Western Atlantic Ocean. Proc. Biol. Soc. Wash. 2009, 122, 1–13. [Google Scholar] [CrossRef]
- Saiz Salinas, J.I.; Bustamante, M.; Tajadura, J.; Vijapure, T.; Sukumaran, S. A new subspecies of Phascolion Théel, 1875 (Sipuncula: Golfingiidae) from Indian waters. Zootaxa 2015, 3931, 433–437. [Google Scholar] [CrossRef][Green Version]
- Silva-Morales, I.; López-Aquino, M.J.; Islas-Villanueva, V.; Ruiz-Escobar, F.J.; Bastida-Zavala, R. Morphological and molecular differences between the Amphiamerican populations of Antillesoma (Sipuncula: Antillesomatidae), with the description of a new species. Rev. Biol. Trop. 2019, 67, S101–S109. [Google Scholar] [CrossRef]
- Staton, J.L. Phylogenetic analysis of the mitochondrial cytochrome c oxidase subunit 1 gene from 13 sipunculan genera: Intra-and interphylum relationships. Invertebr. Biol. 2003, 122, 252–264. [Google Scholar] [CrossRef]
- Maxmen, A.B.; King, B.F.; Cutler, E.B.; Giribet, G. Evolutionary relationships within the protostome phylum Sipuncula; a molecular analysis of ribosomal genes and histone H3 sequence data. Mol. Phylogenet. Evol. 2003, 27, 489–503. [Google Scholar] [CrossRef]
- Schulze, A.; Cutler, E.B.; Giribet, G. Reconstructing the phylogeny of the Sipuncula. Hydrobiologia 2005, 535/536, 277–296. [Google Scholar] [CrossRef]
- Schulze, A.; Cutler, E.B.; Giribet, G. Phylogeny of sipunculan worms: A combined analysis of four gene regions and morphology. Mol. Phylogenet. Evol. 2007, 42, 171–192. [Google Scholar] [CrossRef] [PubMed]
- Kawauchi, G.Y.; Sharma, P.P.; Giribet, G. Sipunculan phylogeny based on six genes, with a new classification and the descriptions of two new families. Zool. Scr. 2012, 41, 186–210. [Google Scholar] [CrossRef]
- Lemer, S.; Kawauchi, G.Y.; Andrade, S.C.S.; González, V.L.; Boyle, M.J.; Giribet, G. Re-evaluating the phylogeny of Sipuncula through transcriptomics. Mol. Phylogenetics Evol. 2015, 83, 174–183. [Google Scholar] [CrossRef]
- Schulze, A.; Rice, M.E. Musculature in sipunculan worms: Ontogeny and ancestral states. Evol. Dev. 2009, 11, 97–108. [Google Scholar] [CrossRef]
- Schulze, A.; Kawauchi, G.Y.; Migotto, A. 13 Sipuncula. In Guide to the Identification of Marine Meiofauna; Schmidt-Rhaesa, A., Ed.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2020; pp. 277–284. [Google Scholar]
- Cordes, E.E.; Carney, S.L.; Hourdez, S.; Carney, R.S.; Brooks, J.M.; Fisher, C.R. Cold seeps of the deep Gulf of Mexico: Community structure and biogeographic comparisons to Atlantic equatorial belt seep communities. Deep Sea Res. Part. I Oceanogr. Res. Pap. 2007, 54, 637–653. [Google Scholar] [CrossRef]
- MacAvoy, S.E.; Fisher, C.R.; Carney, R.S.; Macko, S.A. Nutritional associations among fauna at hydrocarbon seep communities in the Gulf of Mexico. Mar. Ecol. Prog. Ser. 2005, 292, 51–60. [Google Scholar] [CrossRef]
- Cordes, E.E.; Hourdez, S.; Predmore, B.L.; Redding, M.L.; Fisher, C.R. Succession of hydrocarbon seep communities associated with the long-lived foundation species Lamellibrachia luymesi. Mar. Ecol. Prog. Ser. 2005, 305, 17–29. [Google Scholar] [CrossRef]
- Yonge, C.M. A note on mutualism between sipunculans and scleractinian corals. In Proceedings of the International Symposium on the Biology of Sipuncula and Echiura; Rice, M.E., Todorovic, M., Eds.; Nauçno Delo: Belgrade, Serbia, 1975. [Google Scholar]
- Rice, M.E.; Piraino, J.; Reichardt, H.F. Observations on the ecology and reproduction of the sipunculan Phascolion cryptus in the Indian River Lagoon. Fla. Sci. 1983, 46, 382–396. [Google Scholar]
- Edmonds, S.J. Some notes on the abundance, environment, and nutrition of Sipunculus nudus L. (Sipunculoidea) at Morgat, Brittany. Cah. Biol. Mar. III 1962, 3, 183–190. [Google Scholar]
- Pilger, J.F. Ultrastructure of the tentacles of Themiste lageniformis (Sipuncula). Zoomorphology 1982, 100, 143–156. [Google Scholar] [CrossRef]
- Adrianov, A.V.; Malakhov, V.V.; Maiorova, A.S. Development of the tentacular apparatus in sipunculans (Sipuncula): I. Thysanocardia nigra (Ikeda, 1904) and Themiste pyroides (Chamberlin, 1920). J. Morphol. 2006, 267, 569–583. [Google Scholar] [CrossRef] [PubMed]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of worms emended: An update of polychaete feeding guilds. Annu. Rev. Mar. Sci. 2015, 7, 497–520. [Google Scholar] [CrossRef]
- Maiorova, A.S.; Adrianov, A.V. Distribution of peanut worms (Sipuncula) in the West Pacific. In Proceedings of the China-Russia Bilateral Symposium on “Comparison on Marine Biodiversity in the Northwest Pacific Ocean”, Qingdao, China, 10–11 October 2010. [Google Scholar]
- Paul, G. Über Petalostoma minutum Keferstein und verwandte Arten. Jahrb. Abtheilung Anat. 1909, 29, 1–50. [Google Scholar]
- Gibbs, P.E. Gametogenesis and spawning in a hermaphroditic population of Golfingia minuta (Sipuncula). J. Mar. Biol. Assoc. U. K. 1975, 55, 69–82. [Google Scholar] [CrossRef]
- Pilger, J.F. Reproductive biology and development of Themiste lageniformis, a parthenogenic sipunculan. Bull. Mar. Sci. 1987, 41, 59–67. [Google Scholar]
- Rice, M.E. Asexual reproduction in a sipunculan worm. Science 1970, 167, 1618–1620. [Google Scholar] [CrossRef]
- Rajulu, G.S.; Krishnan, N. Occurrence of asexual reproduction by budding in Sipunculida. Nature 1969, 223, 186–207. [Google Scholar] [CrossRef]
- Rice, M.E. A comparative study of the development of Phascolosoma agassizii, Golfingia pugettensis, and Themiste pyroides with a discussion of developmental patterns in the Sipuncula. Ophelia 1967, 4, 143–171. [Google Scholar] [CrossRef]
- Rice, M.E. Observations on the development of six species of Caribbean Sipuncula with a review of development in the phylum. In Proceedings of the International Symposium of Sipuncula and Echiura; Rice, M.E., Todorovic, M., Eds.; Naucno Delo Press: Belgrade, Serbia, 1975; pp. 141–160. [Google Scholar]
- Selenka, E. Eifurchung und Larvenbildung von Phascolosoma elongatum. Z. Wiss. Zool. 1875, 25, 442–450. [Google Scholar]
- Gerould, J.H. The development of Phascolosoma. Zool. Jahrb. Abtheilung Anat. 1906, 23, 77–162. [Google Scholar]
- Åkesson, B. A study of the nervous system of the sipunculoideae, with some remarks on the development of the two species Phascolion strombi Montagu and Golfingia minuta Keferstein. Unders. Över Öresund 1958, 38, 1–249. [Google Scholar]
- Åkesson, B. The development of Golfingia elongata Keferstein (Sipunculidea) with some remarks on the development of neurosecretory cells in sipunculids. Ark. Zool. 1961, 13, 511–531. [Google Scholar]
- Schulze, A.; Boyle, M.J.; Kawauchi, G.Y. Chapter 6.1: Sipuncula. In Annelida; Volume 1: Annelida Basal Groups and Pleistoannelida; Sedentaria, I., Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany, 2019; Volume 1, pp. 177–201. [Google Scholar]
- Boyle, M.J.; Seaver, E.C. Expression of FoxA and GATA transcription factors correlates with regionalized gut development in two lophotrochozoan marine worms: Chaetopterus (Annelida) and Themiste lageniformis (Sipuncula). EvoDevo 2010, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Rice, M.E. Larvae adrift: Patterns and problems in life histories of sipunculans. Am. Zool. 1981, 21, 605–619. [Google Scholar] [CrossRef][Green Version]
- Boyle, M.J.; Rice, M.E. Comparative development and the evolution of life history diversity in Sipuncula. In Proceedings of the Second International Symposium on the Biology of the Sipuncula; Boyle, M.J., Kawauchi, G.Y., Eds.; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2018. [Google Scholar]
- Rice, M.E.; Piraino, J.; Reichardt, H.F.; Boyle, M.J. Observations on oceanic pelagosphera larvae (Sipuncula): Morphology, behavior, and metamorphosis. In Proceedings of the Second International Symposium on the Biology of the Sipuncula; Boyle, M.J., Kawauchi, G.Y., Eds.; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2018; pp. 41–81. [Google Scholar]
- Schulze, A.; Hipes, J.; Borda, E.; Rice, M.E. Who’s who in the Sipuncula: Matching larvae and adults using DNA. In Proceedings of the Second International Symposium on the Biology of the Sipuncula; Boyle, M.J., Kawauchi, G.Y., Eds.; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2018; pp. 83–93. [Google Scholar]
- Knowlton, N. Sibling species in the sea. Annu. Rev. Ecol. Syst. 1993, 24, 189–216. [Google Scholar] [CrossRef]
- Sáez, A.G.; Lozano, E. Body doubles. Nature 2005, 433, 111. [Google Scholar] [CrossRef]
- Staton, J.; Rice, M.E. Genetic Differentiation despite teleplanic larval dispersal: Allozyme variation in sipunculans of the Apionsoma misakianum species complex. Bull. Mar. Sci. 1999, 65, 467–480. [Google Scholar]
- Johnson, N.D.; Schulze, A. Genetic structure in two Phascolosoma species in the Pacific Ocean. Mar. Biol. Res. 2016, 12, 739–747. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; DeWaard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Biol. Sci. 2003, 270, S96–S99. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.F.E.; Gil, J.; Passos, A.M.; Pereira, P.; Melo, P.; Batista, F.; Fonseca, L.C.D. The market features of imported nonindigenous polychaetes in Portugal and consequent ecological concerns. Sci. Mar. 2006, 70, 287–292. [Google Scholar] [CrossRef]
- Kohn, A.J. Predation on Sipuncula. In Proceedings of the International Symposium on the Biology of Sipuncula and Echiura; Rice, M.E., Todorovic, M., Eds.; Nauçno Delo: Belgrade, Serbia, 1975; pp. 113–133. [Google Scholar]
- Kawauchi, G.Y.; Giribet, G. Sipunculus nudus Linnaeus, 1766 (Sipuncula): Cosmopolitan or a group of pseudo-cryptic species? An integrated molecular and morphological approach. Mar. Ecol. 2014, 35, 478–491. [Google Scholar] [CrossRef]
- Adrianov, A.V.; Maiorova, A.S.; Malakhov, V.V. Embryonic and larval development of the peanut worm Themiste pyroides (Sipuncula: Sipunculoidea) from the Sea of Japan. Invertebr. Reprod. Dev. 2008, 52, 143–151. [Google Scholar] [CrossRef]
- Sato, H. Report on the biological survey of Mutsu Bay. 15. Sipunculoidea. Sci. Rep. Ser. 4 Biol. 1930, 5, 1–40. [Google Scholar]
- Cutler, E.B.; Cutler, N.J. A reconsideration of Sipuncula named by I. Ikeda and H. Sato. Publ. Seto Mar. Biol. Lab. 1981, 26, 51–93. [Google Scholar]
- Cutler, E.B.; Cutler, N.J. A revision of the genus Themiste (Sipuncula). Proc. Biol. Soc. Wash. 1988, 101, 741–766. [Google Scholar]
- Schulze, A.; Maiorova, A.; Timm, L.E.; Rice, M.E. Sipunculan larvae and “cosmopolitan” species. Integr. Comp. Biol. 2012, 52, 497–510. [Google Scholar] [CrossRef]
- Adrianov, A.; Maiorova, A. Reproduction and development of common species of peanut worms (Sipuncula) from the Sea of Japan. Russ. J. Mar. Biol. 2010, 36, 1–15. [Google Scholar] [CrossRef]
- Maiorova, A.S.; Adrianov, A.V. Peanut worms of the phylum Sipuncula from the Sea of Japan with a key to species. Deep Sea Res. Part. II Top. Stud. Oceanogr. 2013, 86–87, 140–147. [Google Scholar] [CrossRef]
- Nishikawa, T. Some comments on the taxonomy of the peanut worms (Annelida: Sipuncula) in Japanese waters toward a future revision. In Species Diversity of Animals in Japan; Motokawa, M., Kajihara, H., Eds.; Springer Japan: Tokyo, Japan, 2017; pp. 469–476. [Google Scholar] [CrossRef]
- Rice, M.E. Gametogenesis in three species of Sipuncula: Phascolosoma agassizii, Golfingia pugettensis, and Themiste pyroides. La Cell. 1974, 70, 295–313. [Google Scholar]
- Gibbs, P.E.; Cutler, E.B.; Cutler, N.J. A review of the genus Thysanocardia Fisher (Sipuncula). Zool. Scr. 1983, 12, 295–304. [Google Scholar]
- Haldar, B.P. Sipunculans of the Indian coast. Mem. Zool. Surv. India 1991, 17, 1–169. [Google Scholar]
- Murina, V.V. New records of Echiura and Sipuncula in the North Atlantic Ocean, with the description of a new species of Jacobia. Mar. Biol. Res. 2008, 4, 152–156. [Google Scholar] [CrossRef]
- Açik, Ş. Checklist of Sipuncula from the coasts of Turkey. Turk. J. Zool. 2014, 38, 723–733. [Google Scholar]
- Johnson, N.D.; Sanders, C.; Maiorova, A.; Schulze, A. Cryptic species in Pacific sipunculans (Sipuncula: Phascolosomatidae): East-west divergence between non-sister taxa. Zool. Scr. 2016, 45, 455–463. [Google Scholar] [CrossRef]
- Adrianov, A.V.; Maiorova, A.S.; Malakhov, V.V. Embryonic and larval development of the peanut worm Phascolosoma agassizii (Keferstein 1867) from the Sea of Japan (Sipuncula: Phascolosomatidea). Invertebr. Reprod. Dev. 2011, 55, 22–29. [Google Scholar] [CrossRef]
- Rice, M.E. Morphology, behavior and histogenesis of the pelagosphera larva of Phascolosoma agassizii (Sipuncula). Smithson. Contrib. Zool. 1973, 132, 1–51. [Google Scholar]
- Keferstein, W. Beitrage zur anatomischen und systematischen Kenntniss der Sipunculiden. Z. Fur Wiss. Zool. 1865, 15, 404–445. [Google Scholar]
- Rice, M.E. Survey of the Sipuncula of the coral and beach rock communities of the Caribbean Sea. In Proceedings of the International Symposium of Sipuncula and Echiura; Rice, M.E., Todorovic, M., Eds.; Naucno Delo Press: Belgrade, Serbia, 1975; pp. 35–49. [Google Scholar]
- tenBroeke, A. Westindische Sipunculiden und Echuiriden. Bijdr. Tot De Dierkd. 1925, 24, 81–96. [Google Scholar]
- Baird, W.B. Monograph on the species of worms belonging to the subclass Gephyreae. Proc. Zool. Soc. Lond. 1868, 1868, 77–114. [Google Scholar]
- Murina, V.V. On the sipunculid fauna of the littoral of Cuba. Zool. Zhurnal 1967, 46, 35–47. [Google Scholar]
- Shipley, A.E. Report on the Gephyrean worms collected by Mr. J. Stanley Gardiner at Rotuma and Funafuti. J. Zool. 1898, 66, 468–473. [Google Scholar] [CrossRef]
- Edmonds, S.J. A revision of the systematics of Australian sipunculans (Sipuncula). Rec. South. Aust. Mus. 1980, 18, 1–74. [Google Scholar]
- Cutler, E.B.; Cutler, N.J.; Nishikawa, T. The Sipuncula of Japan: Their systematics and distribution. Publ. Seto Mar. Biol. Lab. 1984, 29, 249–322. [Google Scholar] [CrossRef]
- Fischer, W. Westindische Gephyreen. Zool. Anz. 1922, 55, 10–18. [Google Scholar]
- Cutler, E.B.; Kirsteuer, E. Additional notes on some Sipuncula from Madagascar. Results of the Australian Indo-West Pacific Expedition, 1959–1960. Part 12. Zool. Anz. 1968, 180, 352–356. [Google Scholar]
- Cutler, E.B.; Cutler, N.J. Madagascar and Indian Ocean Sipuncula. Bull. Mus. Natl. D’hist. Nat. Paris 1979, 4, 941–990. [Google Scholar]
- Stephen, A.C. Echiuroidea and Sipunculoidea from Senegal, West Africa. Bull. L’inst. Fr. D’afrique Noire 1960, 22A, 512–520. [Google Scholar]
- Cutler, N.J.; Cutler, E.B. A revision of the subgenus Phascolosoma (Sipuncula, Phascolosoma). Proc. Biol. Soc. Wash. 1990, 3, 691–730. [Google Scholar]
- Kawauchi, G.; Giribet, G. Are there true cosmopolitan sipunculan worms? A genetic variation study within Phascolosoma perlucens (Sipuncula, Phascolosomatidae). Mar. Biol. 2010, 157, 1417–1431. [Google Scholar] [CrossRef]
- Rice, M.E. Factors influencing larval metamorphosis in Golfingia misakiana (Sipuncula). Bull. Mar. Sci. 1986, 39, 362–375. [Google Scholar]
- Hall, J.R.; Scheltema, R.S. Comparative morphology of open-ocean pelagosphera. In Proceedings of the International Symposium on the Biology of the Sipuncula and Echiura; Rice, M.E., Todorovic, M., Eds.; Nauçno Delo Press: Belgrade, Serbia, 1975; pp. 183–197. [Google Scholar]
- Ikeda, I. The Gephyrea of Japan. J. Coll. Sci. Imp. Univ. Tokyo 1904, 20, 1–87. [Google Scholar]
- Gunton, L.M.; Kupriyanova, E.; Alvestad, T.; Avery, L.; Blake, J.A.; Biriukova, O.; Böggemann, M.; Borisova, P.; Budaeva, N.; Burghardt, I.; et al. Annelids of the eastern Australian abyss collected by the 2017 RV Investigator voyage. ZooKeys, in press.
- Murina, V.V. The geographic distribution of marine worms of the phylum Sipuncula of the world ocean. In Proceedings of the International Symposium on the Biology of the Sipuncula and Echiura; Rice, M.E., Todorovic, M., Eds.; Nauçno Delo: Bêlgrade, Serbia, 1975; Volume 1, pp. 3–18. [Google Scholar]
- Cutler, E.B.; Cutler, N.J. Sipuncula from Southern Brazil. Bol. Inst. Oceanogr. Sao Paulo 1980, 29, 1–8. [Google Scholar] [CrossRef]
- Migotto, A.E.; Ditadi, A.S.F. Aspidosiphonidae (Sipuncula) from the northern and northeastern coast of Brazil. Rev. Bras. Biol. 1988, 48, 245–263. [Google Scholar]
- Kawauchi, G.Y.; Migotto, A. Filo Sipuncula. In Biodiversidade Bentônica da Região Sudeste-Sul do Brasil; Amaral, A.C.Z., Rossi-Wongtschowski, C.L.D.B., Eds.; USP Instituto Oceanográfico: São Paulo, Brasil, 2004; pp. 74–76. [Google Scholar]
- Pires Vanin, A.M.S.; Corbisier, T.N.; Arasaki, E.; Möellmann, A.M. Composição e distribuição espaço-temporal da fauna bêntica no Canal de São Sebastião. Realt. Técnico Inst. Oceanogr. 1997, 41, 29–46. [Google Scholar]
- Migotto, A.E.; Tiago, C.G. Síntese. In Biodiversidade do Estado de São Paulo, Brasil: Síntese do Conhecimento ao Final do Século XX, Volume 3: Invertebrados Marinhos; Migotto, A.E., Tiago, C.G., Eds.; Fundação de Amparo à Pesquisa do Estado de São Paulo, FAPESP: São Paulo, Brasil, 1999; pp. 301–310. [Google Scholar]
- Lana, P.C.; Camargo, M.G.; Brogin, R.A.; Isaac, V.J. O bentos da Costa Brasileira: Avaliação Crítica e Levantamento Bibliográfico (1858–1996); FEMAR: Rio de Janeiro, Brasil, 1996; p. 432. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schulze, A.; Kawauchi, G.Y. How Many Sipunculan Species Are Hiding in Our Oceans? Diversity 2021, 13, 43. https://doi.org/10.3390/d13020043
Schulze A, Kawauchi GY. How Many Sipunculan Species Are Hiding in Our Oceans? Diversity. 2021; 13(2):43. https://doi.org/10.3390/d13020043
Chicago/Turabian StyleSchulze, Anja, and Gisele Y. Kawauchi. 2021. "How Many Sipunculan Species Are Hiding in Our Oceans?" Diversity 13, no. 2: 43. https://doi.org/10.3390/d13020043
APA StyleSchulze, A., & Kawauchi, G. Y. (2021). How Many Sipunculan Species Are Hiding in Our Oceans? Diversity, 13(2), 43. https://doi.org/10.3390/d13020043