
Is Biodiversity of Uropodina Mites (Acari: Parasitiformes) Inhabiting Dead Wood Dependent on the Tree Species?


 ,
,
Abstract
:
1. Introduction
2. Material and Methods
3. Results
3.1. Species Composition and Community Structure of Uropodina Mite Communities in Dead Wood of Different Tree Species
3.2. Species Similarity of Uropodina Communities Inhabiting Dead Wood of Different Tree Species
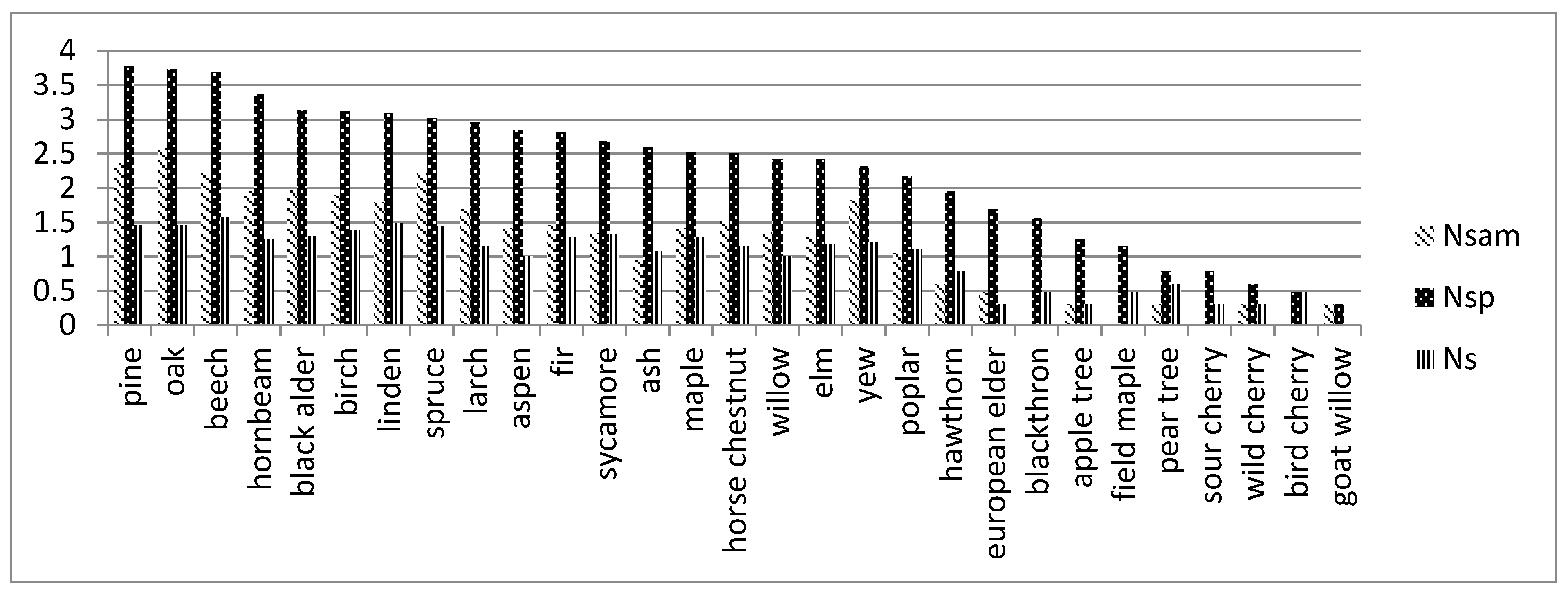
3.3. Impact of Dead Wood of Different Tree Species in Forest Ecosystems on Biodiversity and Abundance of Uropodina Mites
3.4. A Characteristic of Uropodina Communities in Dead Wood of Broadleaved and Coniferous Trees
3.5. A Thorough Analysis of Uropodina Mite Communities in Dead Wood Samples of Five Selected Tree Species over Decades
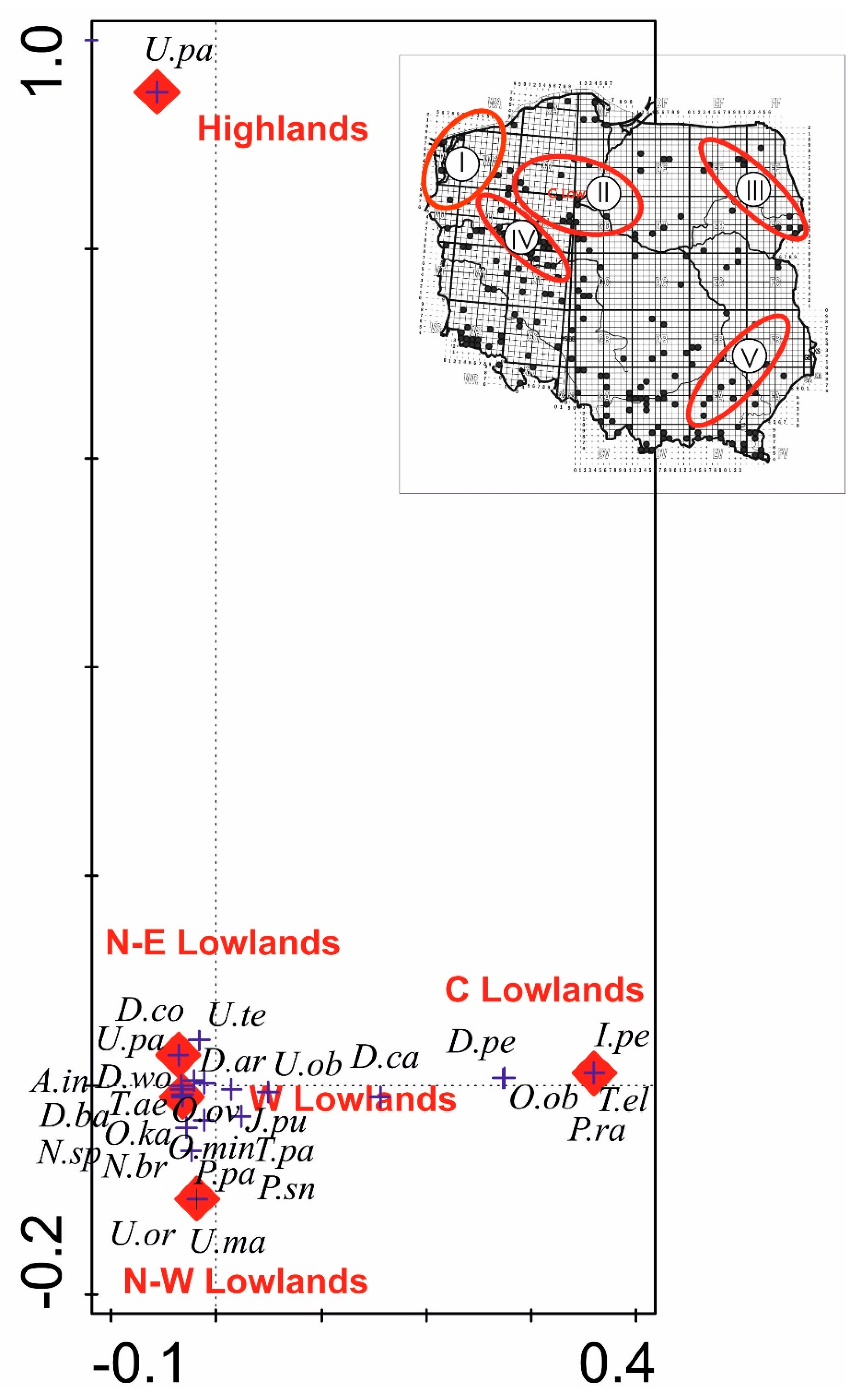
3.6. Multivariate Analysis for Uropodina Communities for Five Tree Species and Sampling Regions
3.7. Habitat Preferences of Uropodina Mite Species for Particular Tree Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Tree Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of samples | 164 | 62 | 384 | 233 | 161 | 79 | 22 | 92 | 26 | 29 | 90 | 65 | 19 | 33 | 49 | 12 | 12 | 9 | 26 | 23 | 4 | 2 | 1 | 1 | 1 | 3 | 2 | 1 | 2 | 1 | 2 |
| Number of specimens | 4857 | 1206 | 5377 | 5995 | 1090 | 1332 | 485 | 1383 | 334 | 645 | 2355 | 205 | 259 | 325 | 1530 | 149 | 407 | 395 | 691 | 275 | 90 | 6 | 3 | 14 | 36 | 49 | 4 | 6 | 18 | 1 | 2 |
| Number of species | 34 | 31 | 29 | 29 | 28 | 21 | 21 | 20 | 19 | 19 | 18 | 16 | 15 | 14 | 14 | 13 | 13 | 12 | 10 | 10 | 6 | 4 | 3 | 3 | 3 | 2 | 2 | 2 | 2 | 1 | 1 |
| Species | |||||||||||||||||||||||||||||||
| Oodinychus ovalis (Koch, 1839) | 967 | 311 | 3122 | 3370 | 447 | 800 | 294 | 731 | 144 | 81 | 1764 | 70 | 47 | 29 | 1129 | 65 | 328 | 319 | 199 | 226 | 60 | 3 | 30 | 48 | 1 | 3 | 6 | 1 | |||
| Pulchellaobovella pulchella (Berlese, 1904) | 804 | 9 | 224 | 1895 | 26 | 200 | 5 | 376 | 3 | 380 | 275 | 1 | 11 | 7 | 128 | 5 | 21 | 92 | 2 | 1 | 1 | 3 | 3 | 12 | 2 | ||||||
| Olodiscus minima (Kramer, 1882) | 19 | 18 | 117 | 303 | 4 | 22 | 6 | 12 | 14 | 51 | 4 | 1 | 3 | 33 | 2 | 4 | 15 | 1 | 8 | 1 | 1 | 1 | |||||||||
| Trachytes aegrota (C. L. Koch, 1841) | 189 | 51 | 156 | 154 | 295 | 54 | 12 | 12 | 9 | 21 | 60 | 12 | 2 | 18 | 113 | 1 | 5 | 12 | 1 | 13 | 5 | ||||||||||
| Dinychus carinatus Berlese, 1903 | 788 | 27 | 165 | 14 | 7 | 23 | 10 | 49 | 19 | 4 | 18 | 5 | 17 | 3 | 338 | 20 | |||||||||||||||
| Dinychus woelkiei (Hirschmann et Zirngiebl-Nicol, 1969) | 86 | 4 | 317 | 6 | 58 | 21 | 4 | 105 | 2 | 4 | 38 | 3 | 9 | 46 | 8 | 1 | |||||||||||||||
| Discourella (?) baloghi (Hirschmann et Z.-Nicol, 1969) | 517 | 21 | 425 | 1 | 16 | 106 | 83 | 5 | 5 | 74 | 2 | 77 | 14 | 2 | 3 | 2 | |||||||||||||||
| Uroobovella pyriformis (Berlese, 1920) | 866 | 336 | 439 | 1 | 3 | 2 | 61 | 36 | 10 | 221 | 1 | 3 | 5 | 32 | 3 | ||||||||||||||||
| Dinychus perforatus Kramer, 1882 | 81 | 3 | 9 | 6 | 2 | 5 | 10 | 1 | 3 | 2 | 26 | 45 | 2 | 4 | 11 | ||||||||||||||||
| Trematurella elegans (Kramer, 1882) | 4 | 15 | 28 | 31 | 10 | 2 | 2 | 3 | 5 | 1 | 79 | 38 | 42 | 1 | 1 | 3 | |||||||||||||||
| Urodiaspis tecta (Kramer, 1876) | 92 | 11 | 93 | 26 | 50 | 22 | 1 | 25 | 9 | 51 | 14 | 7 | 16 | 14 | 5 | 2 | 15 | ||||||||||||||
| Oodinychus karawaiewi (Berlese, 1903) | 1 | 9 | 31 | 13 | 33 | 7 | 3 | 1 | 5 | 1 | 4 | 6 | 3 | 5 | |||||||||||||||||
| Dinychus arcuatus (Trägårdh, 1922) | 107 | 38 | 6 | 81 | 34 | 2 | 5 | 6 | 16 | 7 | 2 | 5 | |||||||||||||||||||
| Pseudouropoda sp. | 5 | 7 | 9 | 18 | 2 | 1 | 1 | 1 | 1 | 16 | 1 | 3 | |||||||||||||||||||
| Iphiduropoda penicillata (Hirschmann et Z.-Nicol, 1961) | 2 | 4 | 8 | 6 | 1 | 2 | 2 | 23 | 1 | 1 | 1 | 2 | |||||||||||||||||||
| Polyaspis patavinus (Berlese, 1881) | 2 | 230 | 8 | 5 | 3 | 7 | 1 | 5 | 19 | 21 | 1 | 8 | |||||||||||||||||||
| Trachytes pauperior (Berlese, 1914) | 15 | 2 | 38 | 8 | 17 | 2 | 4 | 4 | 1 | 3 | 20 | 1 | |||||||||||||||||||
| Uroobovella obovata (Canestrini et Berlese, 1884) | 11 | 5 | 13 | 14 | 2 | 8 | 15 | 1 | 1 | 3 | |||||||||||||||||||||
| Uroobovella sp. | 8 | 5 | 7 | 1 | 2 | 5 | 1 | 1 | 3 | ||||||||||||||||||||||
| Polyaspis sansonei (Berlese, 1916) | 2 | 1 | 93 | 3 | 7 | 1 | 44 | 1 | 1 | ||||||||||||||||||||||
| Apionoseius infirmus (Berlese, 1887) | 57 | 56 | 39 | 4 | 4 | 1 | 1 | 1 | |||||||||||||||||||||||
| Oodinychus obscurasimilis (Hirschmann et Z.-Nicol, 1961) | 19 | 3 | 2 | 1 | 3 | 1 | |||||||||||||||||||||||||
| Oodinychus spatulifera (Moniez, 1892) | 6 | 37 | 12 | 1 | 7 | 2 | |||||||||||||||||||||||||
| Oplitis sp. | 1 | 1 | 1 | 1 | 2 | 5 | |||||||||||||||||||||||||
| Dinychus inermis (C. L. Koch, 1841) | 1 | 3 | 7 | 2 | |||||||||||||||||||||||||||
| Leiodinychus orbicularis (C. L. Koch, 1839) | 23 | 15 | 52 | 1 | 39 | 6 | 1 | ||||||||||||||||||||||||
| Neodiscopoma splendida (Kramer, 1882) | 7 | 27 | 1 | 1 | 9 | 1 | |||||||||||||||||||||||||
| Cilliba cassideasimilis (Błoszyk, Stachowiak et Halliday, 2008) | 2 | 5 | 4 | 2 | |||||||||||||||||||||||||||
| Phaulodiaspis rackei (Oudemans, 1912) | 1 | 1 | 1 | 2 | |||||||||||||||||||||||||||
| Trachytes irenae (Pecina, 1970) | 2 | 28 | 1 | 3 | |||||||||||||||||||||||||||
| Urodiaspis pannonica (Willmann, 1952) | 8 | 1 | 1 | 7 | |||||||||||||||||||||||||||
| Discourella modesta (Leonardi, 1889) | 4 | 2 | 1 | 1 | |||||||||||||||||||||||||||
| Olodiscus misella (Berlese, 1916) | 9 | 1 | 2 | ||||||||||||||||||||||||||||
| Oplitis alophora (Berlese, 1903) | 8 | 1 | |||||||||||||||||||||||||||||
| Trachytes lamda Berlese, 1903 | 1 | 1 | 7 | ||||||||||||||||||||||||||||
| Trachytes minima (Trägårdh, 1910) | 3 | 1 | |||||||||||||||||||||||||||||
| Uroplitella paradoxa (Canestrini et Berlese, 1884) | 1 | 1 | 1 | ||||||||||||||||||||||||||||
| Uropoda sp. | 1 | 16 | 8 | ||||||||||||||||||||||||||||
| Cilliba rafalskii sp.n | 3 | 3 | |||||||||||||||||||||||||||||
| Dinychura cordieri (Berlese, 1916) | 1 | 7 | |||||||||||||||||||||||||||||
| Nenteria stylifera (Berlese, 1904) | 2 | 15 | 3 | ||||||||||||||||||||||||||||
| Nenteria breviunguiculata (Willmann, 1949) | 1 | 2 | |||||||||||||||||||||||||||||
| Pseudouropoda calcarata (Hirschmann et Z.-Nicol, 1961) | 1 | 2 | |||||||||||||||||||||||||||||
| Pseudouropoda tuberosa (Hirschmann et Z.-Nicol, 1961) | 1 | 5 | |||||||||||||||||||||||||||||
| Uroobovella marginata (C. L. Koch, 1829) | 22 | 1 | |||||||||||||||||||||||||||||
| Uropoda orbicularis ((Müller, 1776) | 1 | ||||||||||||||||||||||||||||||
| Allodinychus flagelliger (Berlese, 1910) | 1 | ||||||||||||||||||||||||||||||
| Cilliba erlangensis (Hirschmann et Z.-Nicol, 1969) | 1 | ||||||||||||||||||||||||||||||
| Metagynella carpatica (Balogh, 1943) | 168 | ||||||||||||||||||||||||||||||
| Olodiscus kargi (Hirschamann et Z.-Nicol, 1969) | 4 | ||||||||||||||||||||||||||||||
| Polyaspinus schweizeri (Huţu, 1976) | 1 | ||||||||||||||||||||||||||||||
| Trachytes montana (Willmann, 1953) | 1 | ||||||||||||||||||||||||||||||
| Trachyuropoda coccinea (Michael, 1891) | 6 | ||||||||||||||||||||||||||||||
| Urodiaspis stammeri (Hirschmann et Z.-Nicol, 1969) | 1 |




References
- Sullivan, G.T.; Ozman-Sullivan, S.K. Alarming evidence of widespread mite extinctions in the shadows of plant, insect and vertebrate extinctions. Austral Ecol. 2021, 46, 163–176. [Google Scholar] [CrossRef]
- Ozman-Sullivan, S.K.; Sullivan, G.T. The newly formed Mite Specialist Group of the IUCN’s Species Survival Commission and the conservation of global mite diversity. Acaraological Stud. 2021, 3, 51–55. [Google Scholar] [CrossRef]
- Błoszyk, J.; Rutkowski, T.; Napierała, A.; Konwerski, S.; Zacharyasiewicz, M. Dead Wood as an Element Enriching Biodiversity of Forest Ecosystems: A Case Study Based on Mites from the Suborder Uropodina (Acari: Parasitiformes). Diversity 2021, 13, 476. [Google Scholar] [CrossRef]
- Gutowski, J.M.; Bobiec, A.; Pawlaczyk, P.; Zub, K. Drugie Życie Drzewa; WWF: Warszawa, Poland, 2004; 245p. [Google Scholar]
- Huhta, V.; Siira-Pietikäinen, A.; Penttinen, R. Importance of dead wood for soil mite (Acarina) communities in boreal old-growth forests. Soil Org. 2012, 84, 499–512. [Google Scholar]
- Siitonen, J. Microhabitats. In Biodiversity in Dead Wood, 1st ed.; Stokland, J.N., Siitonen, J., Jonsson, B.G., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 150–182. [Google Scholar]
- Stokland, J.N.; Siitonen, J.; Bengt, G.J. Dead wood in natural forest. In Biodiversity in Dead Wood, 1st ed.; Cambridge University Press: Cambridge, UK, 2012; 301p. [Google Scholar]
- Skubała, P.; Duras, M. Do decaying logs represent habitat islands? Oribatid mite communities in dead wood. Ann. Zool. 2008, 58, 453–466. [Google Scholar] [CrossRef]
- Błoszyk, J. Fauna Uropodina (Acari: Mesostigmata) spróchniałych pni drzew i dziupli w Polsce. Zesz. Probl. Post. Nauk Roln. 1990, 373, 217–235. [Google Scholar]
- Błoszyk, J. Geograficzne i Ekologiczne Zróżnicowanie Zgrupowań Roztoczy z Kohoryty Uropodina (Acari: Mesostigmata) w Polsce: Uropodina Lasów Grądowych (Carpinion betuli); Kontekst: Poznań, Poland, 1999; 245p. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004; 256p. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartig, F. DHARMa: Residual Diagnostics for Hierachical (Multi-Level/Mixed) Regression Models. R Package Version 0.3.3.0. 2020. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 22 November 2021).
- RStudio Team. RStudio: Integrated Development Environment for R. RStudio, PBC, Boston, MA, USA. 2021. Available online: http://www.rstudio.com/ (accessed on 22 November 2021).
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar] [CrossRef] [Green Version]
- Napierała, A.; Błoszyk, J. Unstable microhabitats (merocenoses) as specific habitats of Uropodina mites (Acari: Mesostigmata). Exp. Appl. Acarol. 2013, 60, 163–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Błoszyk, J.; Napierała, A. Zróżnicowane granice zasięgów roztoczy z podrzędu Uropodina (Acari: Mesostigmata) w Polsce. In Proceedings of the Ogólnopolska Konferencja Zoologiczna, Zoologia dziś: Trendy, wyzwania, kierunki na przyszłość, Rzeszów, Poland, 7–8 September 2021. [Google Scholar]
- Napierała, A.; Konwerski, S.; Gutowski, J.M.; Błoszyk, J. Species diversity of Uropodina communities (Acari: Parasitiformes) in soil and selected microhabitats in the Białowieza Primeval Forest. In Mites (Acari) of the Białowieza Primeval Forest, 1st ed.; Błoszyk, J., Napierała, A., Eds.; Kontekst: Poznań, Poland, 2020; pp. 11–60. [Google Scholar]







| Type of Trees | Deciduous | Coniferous | ||||||
|---|---|---|---|---|---|---|---|---|
| Number of Samples | 1061 | 578 | ||||||
| H’ Index | H’ = 1.706 | H’ = 1.454 | ||||||
| Species | n | D% | F% | X ± SD | n | D% | F% | X ± SD |
| O. ovalis | 9338 | 47.45 | 46.84 | 8.842 ± 25.077 | 5097 | 53.86 | 45.50 | 9.005 ± 22.627 |
| U. pyriformis | 1993 | 10.13 | 5.18 | 1.878 ± 24.181 | 38 | 0.40 | 1.04 | 0.066 ± 1.076 |
| P. pulchella | 1926 | 9.79 | 13.95 | 1.815 ± 11.500 | 2430 | 25.68 | 19.38 | 4.204 ± 19.878 |
| D. carinatus | 1457 | 7.40 | 11.59 | 1.373 ± 9.548 | 42 | 0.44 | 3.11 | 0.073 ± 0.557 |
| D. baloghi | 1257 | 6.39 | 4.34 | 1.19 ± 15.712 | 91 | 0.96 | 1.04 | 0.161 ± 2.861 |
| T. aegrota | 596 | 3.03 | 14.70 | 0.564 ± 2.875 | 595 | 6.29 | 21.80 | 1.051 ± 6.747 |
| D. woelkiei | 573 | 2.91 | 5.75 | 0.540 ± 5.222 | 77 | 0.81 | 2.77 | 0.133 ± 2.027 |
| U. tecta | 345 | 1.75 | 8.95 | 0.327 ± 1.805 | 104 | 1.10 | 8.82 | 0.184 ± 0.889 |
| P. patavinus | 301 | 1.53 | 1.41 | 0.285 ± 4.619 | 6 | 0.06 | 0.52 | 0.011 ± 0.178 |
| O. minima | 290 | 1.47 | 9.05 | 0.275 ± 1.335 | 344 | 3.64 | 13.15 | 0.608 ± 2.664 |
| D. arcuatus | 199 | 1.01 | 3.39 | 0.188 ± 1.714 | 105 | 1.11 | 2.94 | 0.182 ± 1.382 |
| T. elegans | 185 | 0.94 | 3.20 | 0.175 ± 2.734 | 81 | 0.86 | 2.60 | 0.143 ± 1.777 |
| D. perforatus | 168 | 0.85 | 2.54 | 0.159 ± 1.937 | 41 | 0.43 | 1.73 | 0.072 ± 1.079 |
| M. carpatica | 168 | 0.85 | 0.28 | 0.159 ± 5.017 | ||||
| A. infirmus | 155 | 0.79 | 1.70 | 0.147 ± 1.932 | 8 | 0.08 | 0.35 | 0.014 ± 0.238 |
| L. orbicularis | 131 | 0.67 | 1.89 | 0.124 ± 1.661 | 6 | 0.06 | 0.35 | 0.011 ± 0.214 |
| O. karawaiewi | 96 | 0.49 | 2.07 | 0.091 ± 1.15 | 23 | 0.24 | 1.04 | 0.041 ± 0.469 |
| T. pauperior | 69 | 0.35 | 3.02 | 0.065 ± 0.596 | 45 | 0.48 | 3.98 | 0.08 ± 0.471 |
| U. obovata | 57 | 0.29 | 1.89 | 0.054 ± 0.54 | 16 | 0.17 | 0.69 | 0.028 ± 0.551 |
| O. spatulifera | 53 | 0.27 | 0.66 | 0.050 ± 0.863 | 12 | 0.13 | 0.35 | 0.021 ± 0.399 |
| I. penicillata | 45 | 0.23 | 1.32 | 0.042 ± 0.74 | 8 | 0.08 | 0.69 | 0.014 ± 0.195 |
| N. splendida | 35 | 0.18 | 0.66 | 0.033 ± 0.837 | 11 | 0.12 | 0.52 | 0.019 ± 0.383 |
| Pseudouropoda sp. | 35 | 0.18 | 1.32 | 0.033 ± 0.524 | 30 | 0.32 | 1.90 | 0.053 ± 0.491 |
| Uroobovella sp. | 30 | 0.15 | 1.41 | 0.028 ± 0.291 | 3 | 0.03 | 0.52 | 0.005 ± 0.073 |
| U. marginata | 22 | 0.11 | 0.28 | 0.021 ± 0.563 | ||||
| N. stylifera | 20 | 0.10 | 0.28 | 0.019 ± 0.475 | ||||
| Uropoda sp. | 17 | 0.09 | 0.19 | 0.016 ± 0.493 | 8 | 0.08 | 0.17 | 0.014 ± 0.336 |
| P. sansonei | 13 | 0.07 | 0.57 | 0.012 ± 0.232 | 140 | 1.48 | 1.90 | 0.247 ± 2.797 |
| D. inermis | 12 | 0.06 | 0.38 | 0.011 ± 0.199 | 1 | 0.01 | 0.17 | 0.002 ± 0.042 |
| C. cassideasimilis | 11 | 0.06 | 0.38 | 0.010 ± 0.202 | 2 | 0.02 | 0.17 | 0.004 ± 0.084 |
| O. misella | 11 | 0.06 | 0.38 | 0.010 ± 0.206 | 1 | 0.01 | 0.17 | 0.002 ± 0.042 |
| Oplitis sp. | 9 | 0.05 | 0.38 | 0.009 ± 0.171 | 2 | 0.02 | 0.35 | 0.004 ± 0.059 |
| U. pannonica | 9 | 0.05 | 0.57 | 0.009 ± 0.141 | 8 | 0.08 | 0.69 | 0.014 ± 0.222 |
| D. modesta | 8 | 0.04 | 0.47 | 0.008 ± 0.115 | ||||
| O. obscurasimilis | 7 | 0.04 | 0.57 | 0.007 ± 0.092 | 22 | 0.23 | 1.04 | 0.039 ± 0.645 |
| P. tuberosa | 6 | 0.03 | 0.19 | 0.006 ± 0.157 | ||||
| T. coccinea | 6 | 0.03 | 0.19 | 0.006 ± 0.138 | ||||
| O. kargi | 4 | 0.02 | 0.19 | 0.004 ± 0.097 | ||||
| P. rackei | 4 | 0.02 | 0.28 | 0.004 ± 0.075 | 1 | 0.01 | 0.17 | 0.002 ± 0.042 |
| C. rafalskii | 3 | 0.02 | 0.19 | 0.003 ± 0.069 | 3 | 0.03 | 0.17 | 0.005 ± 0.126 |
| T. irenae | 3 | 0.02 | 0.19 | 0.003 ± 0.069 | 31 | 0.33 | 0.87 | 0.055 ± 0.793 |
| T. minima | 3 | 0.02 | 0.09 | 0.003 ± 0.092 | 1 | 0.01 | 0.17 | 0.002 ± 0.042 |
| N. breviunguiculata | 2 | 0.01 | 0.19 | 0.002 ± 0.043 | 2 | 0.02 | 0.35 | 0.004 ± 0.059 |
| T. lamda | 2 | 0.01 | 0.19 | 0.002 ± 0.043 | 7 | 0.07 | 0.17 | 0.012 ± 0.294 |
| A. flagelliger | 1 | 0.01 | 0.09 | 0.001 ± 0.031 | ||||
| C. erlangensis | 1 | 0.01 | 0.09 | 0.001 ± 0.031 | ||||
| D. cordieri | 1 | 0.01 | 0.09 | 0.001 ± 0.031 | 7 | 0.07 | 0.35 | 0.012 ± 0.256 |
| O. alophora | 1 | 0.01 | 0.09 | 0.001 ± 0.031 | 8 | 0.08 | 0.52 | 0.014 ± 0.23 |
| P. calcarata | 1 | 0.01 | 0.09 | 0.001 ± 0.031 | 2 | 0.02 | 0.17 | 0.004 ± 0.084 |
| U. stammeri | 1 | 0.01 | 0.09 | 0.001 ± 0.031 | ||||
| U. paradoxa | 1 | 0.01 | 0.09 | 0.001 ± 0.031 | 2 | 0.02 | 0.35 | 0.004 ± 0.059 |
| P. schweizeri | 1 | 0.01 | 0.17 | 0.002 ± 0.042 | ||||
| T. montana | 1 | 0.01 | 0.17 | 0.002 ± 0.042 | ||||
| Total | 19,681 | 100.00 | 64.18 | 18.549 ± 52.262 | 9463 | 100.00 | 64.71 | 16.372 ± 36.067 |
| Species | Beech | Birch | Oak | Hornbeam | Pine | Spruce | Pearson Χ2 Statistic | p-Value |
|---|---|---|---|---|---|---|---|---|
| P. pulchella | −0.401 | 1.118 | −3.418 | 0.358 | 5.259 | −2.845 | 58.9086 | <0.001 |
| O. minima | −1.995 | −0.327 | −1.112 | 1.754 | 4.114 | −3.262 | 97.1116 | <0.001 |
| O. ovalis | −0.268 | −0.552 | 0.806 | 0.747 | 0.929 | −2.685 | 13.3092 | <0.025 |
| T. aegrota | 0.113 | −0.079 | −1.286 | −0.321 | 0.289 | 1.996 | 4.9874 | >0.05 |
| U. tecta | −0.113 | 0.737 | −0.175 | −0.149 | −1.602 | 2.363 | 7.5201 | >0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zacharyasiewicz, M.; Napierała, A.; Kurek, P.; Grossmann, K.; Błoszyk, J. Is Biodiversity of Uropodina Mites (Acari: Parasitiformes) Inhabiting Dead Wood Dependent on the Tree Species? Diversity 2021, 13, 609. https://doi.org/10.3390/d13120609
Zacharyasiewicz M, Napierała A, Kurek P, Grossmann K, Błoszyk J. Is Biodiversity of Uropodina Mites (Acari: Parasitiformes) Inhabiting Dead Wood Dependent on the Tree Species? Diversity. 2021; 13(12):609. https://doi.org/10.3390/d13120609
Chicago/Turabian StyleZacharyasiewicz, Michał, Agnieszka Napierała, Przemysław Kurek, Kamila Grossmann, and Jerzy Błoszyk. 2021. "Is Biodiversity of Uropodina Mites (Acari: Parasitiformes) Inhabiting Dead Wood Dependent on the Tree Species?" Diversity 13, no. 12: 609. https://doi.org/10.3390/d13120609
APA StyleZacharyasiewicz, M., Napierała, A., Kurek, P., Grossmann, K., & Błoszyk, J. (2021). Is Biodiversity of Uropodina Mites (Acari: Parasitiformes) Inhabiting Dead Wood Dependent on the Tree Species? Diversity, 13(12), 609. https://doi.org/10.3390/d13120609