
A Forest Pool as a Habitat Island for Mites in a Limestone Forest in Southern Norway


 , ,
, ,
Abstract
1. Introduction
2. Material and Methods
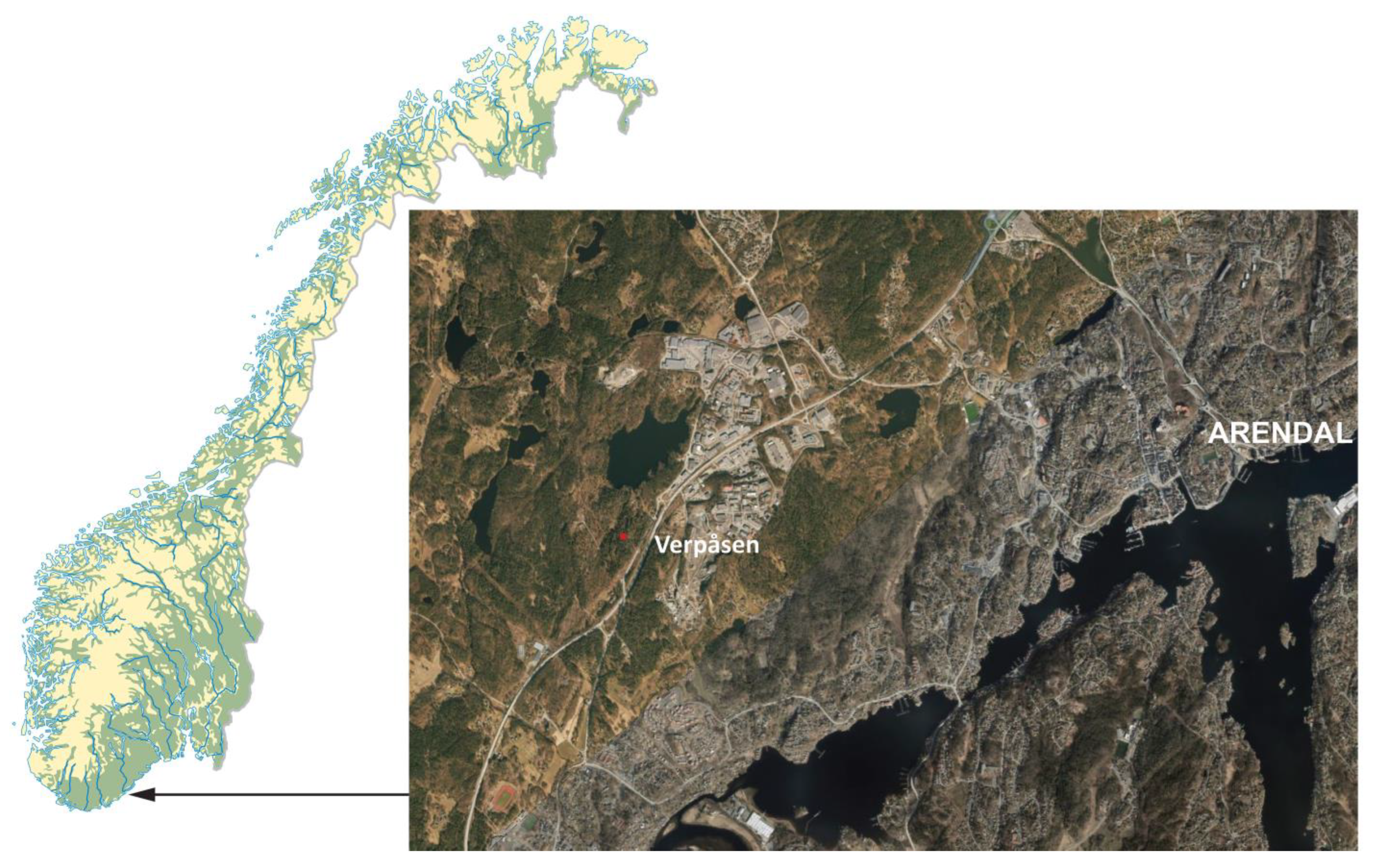
2.1. Study Site
2.2. Sampling and Identification
2.3. Statistical Analyses
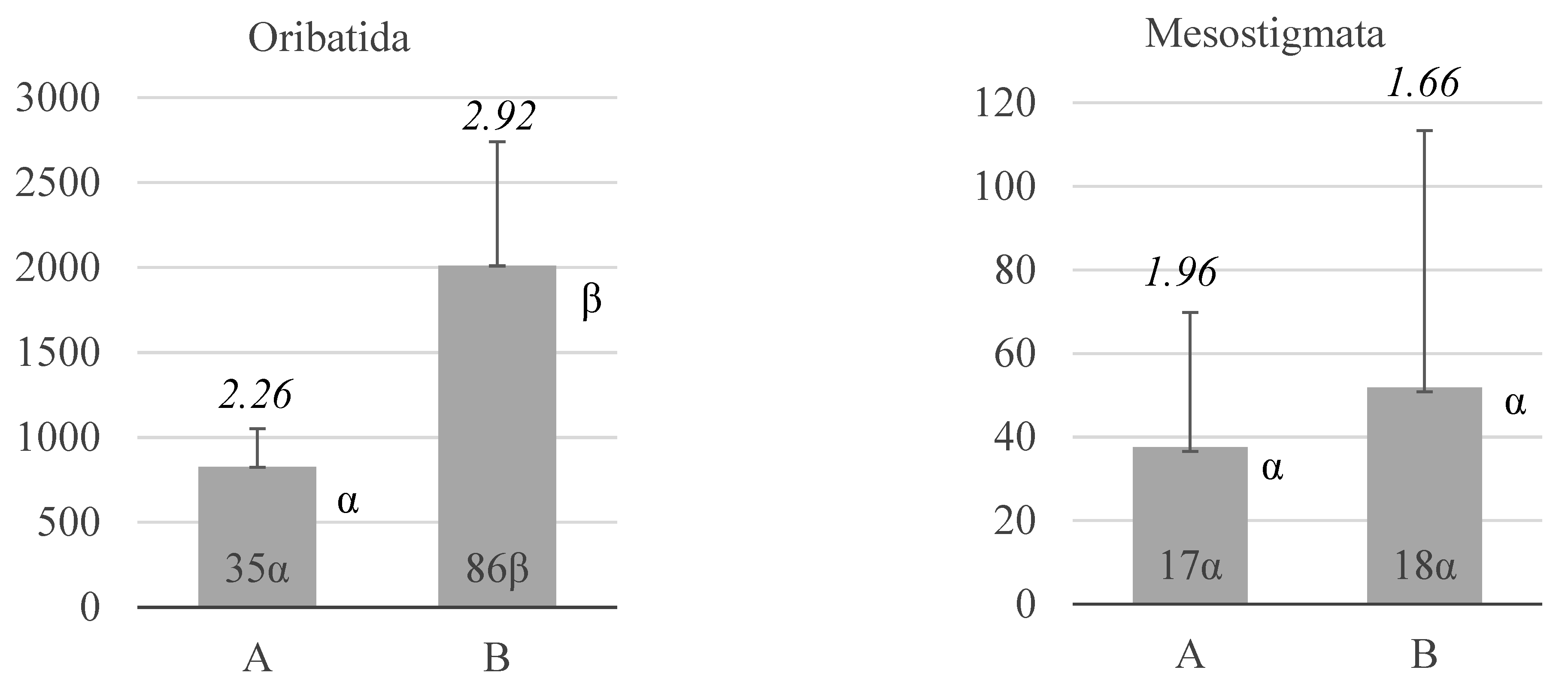
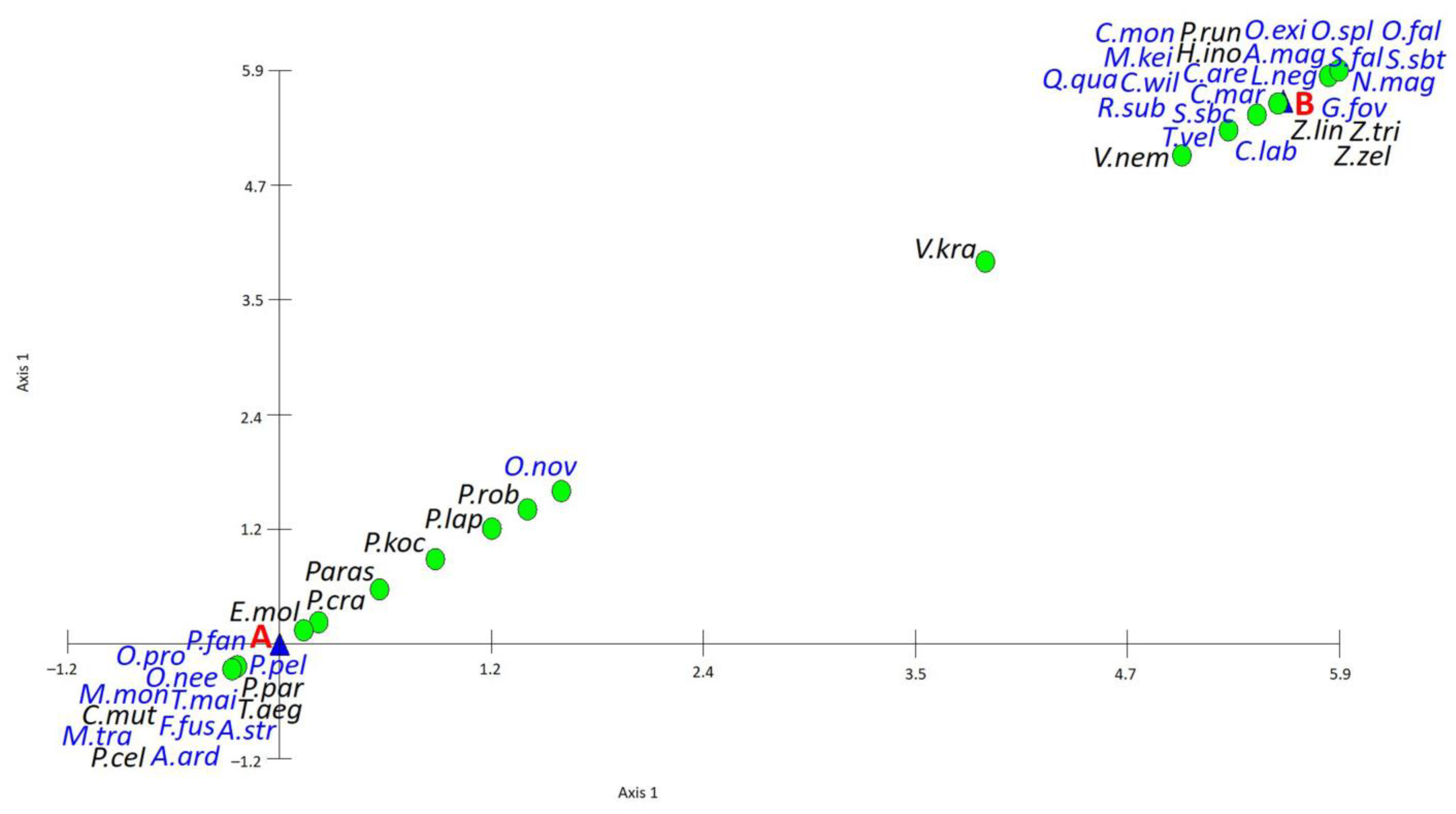
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waldon, B. Waloryzacja Przyrodnicza Śródleśnych Oczek Pojezierza Krajeńskiego; Banaszak, J., Tobolski, K., Eds.; Park Narodowy Bory Tucholskie: Charzykowy, Poland, 2002; pp. 289–299. [Google Scholar]
- Spyra, A. Acidic, neutral and alkaline forest ponds as a landscape element affecting the biodiversity of freshwater snails. Sci. Nat. 2017, 104, 73. [Google Scholar] [CrossRef]
- Namura-Ochalska, A. Śródleśne jeziora oligo-humotroficzne jako naturalne zbiorniki retencyjne. Stud. Mater. Cent. Eduk. Przyr. Leś. 2008, 2, 125–139. [Google Scholar]
- Seniczak, A. Mites (Acari) of the Shores of Forest Lakes and Ponds in Northern Poland, with Species Analysis of Oribatida; Wydawnictwa UTP: Bydgoszcz, Poland, 2011; pp. 1–231. ISSN 0209-0597. [Google Scholar]
- Hågvar, S.; Amundsen, T. Effects of liming and artificial acid rain on the mite (Acari) fauna in coniferous forest. Oikos 1981, 37, 7–20. [Google Scholar] [CrossRef]
- Hågvar, S. Ecological spotlights on mites (Acari) in Norwegian conifer forests: A review. Pests Control Acarol. 2018. [Google Scholar] [CrossRef]
- Seniczak, A.; Bolger, T.; Roth, S.; Seniczak, S.; Djursvoll, P.; Jordal, B.H. Diverse mite communities (Acari: Oribatida, Mesostigmata) from a broadleaf forest in western Norway. Ann. Zool. Fenn. 2019, 56, 121–136. [Google Scholar] [CrossRef]
- Seniczak, A.; Seniczak, S.; Starý, J.; Kaczmarek, S.; Jordal, B.H.; Kowalski, J.; Roth, S.; Djursvoll, P.; Bolger, T. High diversity of mites (Acari: Oribatida, Mesostigmata) supports the high conservation value of a broadleaf forest in Eastern Norway. Forests 2021, 12, 1098. [Google Scholar] [CrossRef]
- Walter, D.E.; Proctor, H.C. Mites: Ecology, Evolution and Behaviour; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2013. [Google Scholar] [CrossRef]
- Seniczak, A.; Seniczak, S.; Graczyk, R.; Waldon-Rudzionek, B.; Nowicka, A.; Pacek, S. Seasonal Dynamics of Oribatid Mites (Acari, Oribatida) in a Bog in Poland. Wetlands 2019, 39, 853–864. [Google Scholar] [CrossRef]
- Seniczak, A.; Niedbała, W.; Iturrondobeitia, J.C.; Seniczak, S.; Roth, S.; Jordal, B.H. Type of broadleaf forest matters most for ptyctimous mite communities (Acari, Oribatida) in Norway. Biodivers. Conserv. 2021, 30, 2929–2953. [Google Scholar] [CrossRef]
- Meteorological Institute. Available online: https://www.met.no (accessed on 8 June 2021).
- Moen, A.; Lillethun, A. National Atlas of Norway: Vegetation; Norwegian Mapping Authorities: Hønefoss, Norway, 1999. [Google Scholar]
- Miljødirektoratet. Available online: https://faktaark.naturbase.no (accessed on 18 July 2021).
- Ghiljarov, M.S.; Krivolutskij, D.A. Opredelitel Obitajushchikh v Pochve Kleshchei, Sarcoptiformes; Nauka: Moskva, Russia, 1975; pp. 1–492. [Google Scholar]
- Pérez-Íñigo, C. Acari, Oribatei, Poronota. In Fauna Ibérica; Ramos, M.A., Ed.; Museo Nacional de Ciencias Naturales CSIC: Madrid, Spain, 1993; Volume 3, pp. 1–320. [Google Scholar]
- Pérez-Íñigo, C. Acari, Oribatei, Gymnonota, I. In Fauna Ibérica; Ramos, M.A., Ed.; Museo Nacional de Ciencias Naturales CSIC: Madrid, Spain, 1997; Volume 9, pp. 1–373. [Google Scholar]
- Weigmann, G. Hornmilben (Oribatida). In Die Tierwelt Deutschlands; Goecke and Evers: Keltern, Germany, 2006; Volume 76, pp. 1–520. [Google Scholar]
- Seniczak, S. The morphology of juvenile stages of moss mites of the family Pelopidae Ewing (Acarida: Oribatida), II. Ann. Zool. 1988, 41, 383–393. [Google Scholar]
- Seniczak, S. The morphology of juvenile stages of moss-mites of the subfamily Sphaerozetinae (Acarida: Oribatida), II. Ann. Zool. 1989, 42, 237–248. [Google Scholar]
- Seniczak, S. The morphology of juvenile stages of moss mites of the family Camisiidae (Acari: Oribatida), V. Zool. Anz. 1991, 227, 173–184. [Google Scholar]
- Seniczak, S. The morphology of juvenile stages of moss mites of the family Malaconothridae (Acari, Oribatida). I. Zool. Anz. 1993, 231, 59–72. [Google Scholar]
- Seniczak, S.; Klimek, A. The morphology of juvenile stages of moss mites of the family Camisiidae (Acari: Oribatida). Zool. Anz. 1990, 225, 71–86. [Google Scholar]
- Seniczak, S.; Żelazna, E. The Morphology of juvenile stages of moss mites of the family Nothridae (Acari, Oribatida). II Zool. Anz. 1992, 229, 149–162. [Google Scholar]
- Seniczak, S.; Seniczak, A. Morphology of juvenile stages of Parachipteria bella (Sellnick, 1928) and P. willmanni Hammen, 1952 (Acari: Oribatida: Achipteriidae). Ann. Zool. 2007, 57, 533–540. [Google Scholar] [CrossRef]
- Seniczak, S.; Seniczak, A. Morphology of three species of Crotonioidea Thorel, 1876 (Acari: Oribatida), and relations between some genera. Zool. Anz. 2009, 248, 195–211. [Google Scholar] [CrossRef]
- Seniczak, S.; Seniczak, A. Differentiation of external morphology of Damaeidae (Acari: Oribatida) in light of the ontogeny of three species. Zootaxa 2011, 2775, 1–36. [Google Scholar] [CrossRef]
- Seniczak, A.; Seniczak, S. Morphological ontogeny of Caleremaeus monilipes (Michael, 1882) (Acaria: Oribatida: Caleremaeidae) with comments on Caleremaeus Berlese. Syst. Appl. Acarol. 2019, 24, 1995–2009. [Google Scholar] [CrossRef]
- Ermilov, S.G. Ontogeny of oribatid mite Nanhermannia coronata (Acari, Oribatida, Nanhermanniidae). Zool. Zhurn. 2009, 88, 429–437. [Google Scholar] [CrossRef]
- Ermilov, S.G.; Łochyńska, M. Morphology of juvenile stages of Epidamaeus kamaensis (Sellnick, 1925) and Porobelba spinosa (Sellnick, 1920) (Acari: Oribatida: Damaeidae). Ann. Zool. 2009, 59, 527–544. [Google Scholar] [CrossRef]
- Seniczak, S.; Norton, R.A.; Seniczak, A. Morphology of Eniochthonius minutissimus (Berlese, 1904) and Hypochthonius rufulus C.L. Koch, 1835 (Acari: Oribatida: Hypochthonioidea). Ann. Zool. 2009, 59, 373–386. [Google Scholar] [CrossRef]
- Seniczak, S.; Seniczak, A.; Kaczmarek, S.; Słowikowska, M. Variability of external morphology of Eueremaeus Mihelčič, 1963 (Acari, Oribatida, Eremaeidae) in the light of ontogeny of three species. Int. J. Acarology 2014, 40, 81–108. [Google Scholar] [CrossRef]
- Seniczak, A.; Seniczak, S.; Sgardelis, S.; Graczyk, R. Morphological ontogeny, distribution and ecology of Edwardzetes edwardsii and Sphaerozetes orbicularis (Acari, Oribatida, Ceratozetidae). Syst. Appl. Acarol. 2016, 21, 713–744. [Google Scholar] [CrossRef]
- Seniczak, A.; Seniczak, S.; Graczyk, R.; Bukowski, G. Morphological ontogeny, ecology and some biological parameters of Achipteria magna (Acari: Oribatida: Achipteriidae). Syst. Appl. Acarol. 2017, 22, 980–992. [Google Scholar] [CrossRef]
- Seniczak, A.; Seniczak, S.; Kaczmarek, S.; Chachaj, B. Morphological ontogeny and ecology of Adoristes ovatus (Acari: Oribatida: Liacaridae), with comments on Adoristes Hull. Syst. Appl. Acarol. 2017, 22, 2038–2056. [Google Scholar] [CrossRef]
- Seniczak, S.; Seniczak, A.; Kaczmarek, S.; Marquardt, T. Morphological ontogeny, ecology and some biological parameters of Achipteria nitens (Acari: Oribatida: Achipteriidae). Zootaxa 2019, 4717, 030–046. [Google Scholar] [CrossRef]
- Pfingstl, T.; Krisper, G. No difference in the juveniles of two Tectocepheus species (Acari: Oribatida, Tectocepheidae). Acarologia 2011, 51, 199–218. [Google Scholar] [CrossRef][Green Version]
- Ermilov, S.G.; Kolesnikov, V.B. Morphology of juvenile instars of Furcoribula furcillata and Zygoribatula exilis (Acari, Oribatida). Acarina 2012, 20, 48–59. [Google Scholar]
- Schatz, H. Catalogue of oribatid mites (Acari: Oribatida) from Vorarlberg (Austria). Zootaxa 2020, 4783, 1–106. [Google Scholar] [CrossRef]
- Norton, R.A.; Ermilov, S.G. Catalogue and historical overview of juvenile instars of oribatid mites (Acari: Oribatida). Zootaxa 2014, 3833, 1–132. [Google Scholar] [CrossRef]
- Subías, L.S. Listado sistemático, sinonímico y biogeográfico de los Ácaros Oribátidos (Acariformes, Oribatida) del mundo (1758–2002). Graellsia 2021, 60, 3–305, unpublished electronic update, 15ª actualización, 527p. [Google Scholar] [CrossRef]
- Bhattacharyya, S.K. A revision of the British mites of the genus Pergamasus Berlese s lat. (Acari: Mesostigmata). Bull. Br. Mus. nat. Hist. Zool. 1963, 11, 133–242. [Google Scholar]
- Ghiljarov, M.S.; Bregetova, N.G. Opredelitel Obitajushchikh v Pochve Kleshchei; Mesostigmata; Nauka: Moskva, Russia, 1977; pp. 1–718. [Google Scholar]
- Lundqvist, L.; Johnston, D.E. Description of Zercon lindrothi sp. n. and a redescription of Zercon colligans Berlese, 1920 (Acari, Mesostigmata: Zerconidae). Insect Syst. Evol. 1985, 16, 345–350. [Google Scholar] [CrossRef]
- Micherdziński, W. Die Familie Parasitidae Oudemans 1901 (Acarina, Mesostigmata); PWN: Warszawa, Poland, 1969; pp. 1–690. [Google Scholar]
- Błaszak, C. Zerconidae (Acari, Mesostigmata) Polski. Monogr. Fauny Pol. 1974, 3, 1–315. [Google Scholar]
- Hyatt, K.H.; Emberson, R.M. A review of the Macrochelidae (Acari: Mesostigmata) of the British Isles. Bull. Br. Mus. Nat. Hist. Zool. 1988, 54, 63–125. [Google Scholar]
- Karg, W. Acari (Acarina), Milben, Unterordnung Parasitiformes (Anactinochaeta), Uropodina Kramer, Schildkrötenmilben; Gustav Fischer Verlag: Jena, Germany, 1989; pp. 1–203. [Google Scholar]
- Karg, W. Raubmilben: Acari (Acarina), Milben Parasitiformes (Allactinochaeta) Cohors Gamasina Leach; Gustav Fischer Verlag: Jena, Germany, 1993; pp. 1–524. [Google Scholar]
- Gwiazdowicz, D.J. Ascid Mites (Acari, Mesostigmata) from Selected Forest Ecosystems and Microhabitats in Poland; Wydawnictwo Akademii Rolniczej im. Augusta Cieszkowskiego: Poznań, Poland, 2007; pp. 1–248. [Google Scholar]
- Witaliński, W. Key to the world species of Holoparasitus Oudemans, 1936 (Acari: Parasitiformes: Parasitidae). Zootaxa 2017, 4277, 301–351. [Google Scholar] [CrossRef]
- Mehl, R. Checklist of Norwegian ticks and mites (Acari). Fauna Norv. B 1979, 26, 31–45. [Google Scholar]
- Seniczak, A.; Seniczak, S.; Iturrondobeitia, J.C.; Solhøy, T.; Flatberg, K.I. Diverse Sphagnum mosses support rich moss mite communities (Acari, Oribatida) in mires of western Norway. Wetlands 2020, 40, 1339–1351. [Google Scholar] [CrossRef]
- Seniczak, A.; Seniczak, S.; Schwarzfeld, M.D.; Coulson, S.J.; Gwiazdowicz, D.J. Diversity and Distribution of Mites (Acari: Ixodida, Mesostigmata, Trombidiformes, Sarcoptiformes) in the Svalbard Archipelago. Diversity 2020, 12, 323. [Google Scholar] [CrossRef]
- Seniczak, A.; Seniczak, S. Diversity of oribatid mites (Acari: Oribatida) in the Svalbard archipelago: A historical overview. Zootaxa 2020, 4834, 41–65. [Google Scholar] [CrossRef] [PubMed]
- Seniczak, A.; Seniczak, S. Morphological ontogeny of Fuscozetes coulsoni sp. nov. (Acari: Oribatida: Ceratozetidae) from Svalbard, Norway. Syst. Appl. Acarol. 2020, 25, 680–696. [Google Scholar] [CrossRef]
- Seniczak, A.; Seniczak, S. Morphological ontogeny of Limnozetes solhoyorum sp. nov. (Acari: Oribatida: Limnozetidae) from Norway, with comments on Limnozetes Hull. Syst. Appl. Acarol. 2020, 25, 327–348. [Google Scholar] [CrossRef]
- Seniczak, A.; Seniczak, S. Morphological ontogeny of Limnozetes schatzi sp. nov. (Acari: Oribatida: Limnozetidae) from Norway. Syst. Appl. Acarol. 2021, 26, 1974–1991. [Google Scholar] [CrossRef]
- Lebedeva, N.V.; Taskaeva, A.A.; Melekhina, E.N.; Zazovskaya, E.P. Invertebrates in the gradient of different age soils under a birdcliff at the Grønfjord Area (Svalbard, Arctic). Cauc. Ent. Bull. 2021, 16, 283–294. [Google Scholar] [CrossRef]
- Thunes, K.H.; Søli, G.E.E.; Thuróczy, C.; Fjellberg, A.; Olberg, S.; Roth, S.; Coulianos, C.-C.; Disney, R.H.L.; Starý, J.; Vierbergen, G.; et al. The Arthropod Fauna of Oak (Quercus spp., Fagaceae) Canopies in Norway. Diversity 2021, 13, 332. [Google Scholar] [CrossRef]
- Lundqvist, L. Bibliografi och checklist över Sveriges oribatider (Acari: Oribatei). Entomol. Tidskr. 1987, 108, 3–12. [Google Scholar]
- Niemi, R.; Karppinen, E.; Uusitalo, M. Catalogue of the Oribatida (Acari) of Finland. Acta Zool. Fenn. 1997, 207, 1–39. [Google Scholar]
- Koponen, S.; Rinne, V.; Clayhills, T. Arthropods on oak branches in SW Finland, collected by a new trap type. Entomol. Fenn. 1997, 8, 177–183. [Google Scholar] [CrossRef]
- Huhta, V.; Sulkava, P.; Viberg, K. Interactions between enchytraeid (Cognettia sphagnetorum), microarthropod and nematode populations in forest soil at different moistures. Appl. Soil Ecol. 1998, 9, 53–58. [Google Scholar] [CrossRef]
- Huhta, V.; Räty, M.; Ahlroth, P.; Hänninen, S.M.; Mattila, J.; Penttinen, R.; Rintala, T. Soil fauna of deciduous forests in central Finland. Memo. Soc. Fauna Flora Fenn. 2005, 81, 52–70. [Google Scholar]
- Huhta, V.; Siira-Pietikäinen, A.; Penttinen, R.; Räty, M. Soil fauna of Finland: Acarina, Collembola and Enchytraeidae. Memo. Soc. Fauna Flora Fenn. 2010, 86, 59–82. [Google Scholar]
- Huhta, V.; Siira-Pietikäinen, A.; Penttinen, R. Importance of dead wood for soil mite (Acarina) communities in boreal old-growth forests. Soil Org. 2012, 84, 499–512. [Google Scholar]
- Huhta, V.; Penttinen, R.; Pitkänen, E. Cultural factors in the distribution of soil mites in Finland. Memo. Soc. Fauna Flora Fenn. 2012, 88, 52–70. [Google Scholar]
- Froberg, L.; Solhøy, T.; Baur, A.; Baur, B. Lichen specificity of Oribatid mites (Acari; Oribatida) on limestone walls in the Great Alvar of Gland, Sweden. Entomol. Tidskr. 2003, 124, 177–182. [Google Scholar]
- Huhta, V.; Niemi, R. Communities of soil mites (Acarina) in planted birch stands as compared with natural forests in central Finland. Can. J. For. Res. 2003, 33, 171–180. [Google Scholar] [CrossRef]
- Penttinen, R.; Siira-Pietikäinen, A.; Huhta, V. Oribatid mites in eleven different habitats in Finland. In Integrative Acarology: Proceedings of the 6th European Congress of the EURAAC; Bertrand, M., Kreiter, S., McCoy, K.D., Migeon, A., Navajas, M., Tixier, M.S., Vial, L., Eds.; European Association of Acarologists: Bari, Italy, 2008; pp. 237–244. [Google Scholar]
- Siira-Pietikäinen, A.; Penttinen, R.; Huhta, V. Oribatid mites (Acari: Oribatida) in boreal forest floor and decaying wood. Pedobiologia 2008, 52, 111–118. [Google Scholar] [CrossRef]
- Penttinen, R.; Huhta, V. Ptyctima (Acari, Oribatida) in various habitats in Finland. In Trends in Acarology: Proceedings of the 12th International Congress on Acarology; Sabelis, M.W., Bruin, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 167–170. [Google Scholar]
- Elo, R.A.; Penttinen, R.; Sorvari, J. Distribution of oribatid mites is moisture-related within red wood ant Formica polyctena nest mounds. Appl. Soil Ecol. 2018, 124, 203–210. [Google Scholar] [CrossRef]
- Elo, R.A.; Penttinen, R.; Sorvari, J. A comparative study of oribatid mite communities in red wood ant Formica polyctena nests and surrounding soil in a Finnish oak forest. Insect Conserv. Divers. 2016, 9, 210–223. [Google Scholar] [CrossRef]
- Niedbała, W.; Liu, D. Catalogue of ptyctimous mites (Acari, Oribatida) of the world. Zootaxa 2018, 4393, 1–238. [Google Scholar] [CrossRef] [PubMed]
- Gwiazdowicz, D.J.; Gulvik, M.E. Checklist of Norwegian mesostigmatid mites (Acari, Mesostigmata). Nor. J. Entomol. 2005, 52, 117–125. [Google Scholar]
- Gwiazdowicz, D.J.; Gulvik, M.E. The first records of five mite species (Acari, Mesostigmata) in Norway. Nor. J. Entomol. 2007, 54, 125–127. [Google Scholar]
- Słomian, S.; Gulvik, M.E.; Madej, G.; Austad, I. Gamasina and Microgyniina (Acari, Gamasida) from soil and tree hollows at two traditional farms in Sogn og Fjordane, Norway. Nor. J. Entomol. 2005, 52, 39–48. [Google Scholar]
- Gwiazdowicz, D.J.; Solhøy, T.; Kaasa, K. Five mesostigmatid mites (Acari, Mesostigmata) new to the Norwegian fauna. Nor. J. Entomol. 2013, 60, 8–10. [Google Scholar]
- Bolger, T.; Devlin, M.; Seniczak, A. First records of ten species of Mesostigmata (Acari, Mesostigmata) added to the published Norwegian species list. Nor. J. Entomol. 2018, 65, 94–100. [Google Scholar]
- Kaczmarek, S.; Marquardt, T.; Seniczak, A. A new species of Zercon (Parasitiformes: Mesostigmata) from Norway, with notes on sexual dimorphism in Zerconidae. Syst. Appl. Acarol. 2021, 26, 1676–1702. [Google Scholar] [CrossRef]
- Huhta, V. Catalogue of the Mesostigmata mites in Finland. Memo. Soc. Fauna Flora Fenn. 2016, 92, 129–148. [Google Scholar]
- SLU Artdatabanken. Available online: https://artfakta.se (accessed on 20 September 2021).
- Odum, E.P. Podstawy Ekologii; PWRiL: Warszawa, Poland, 1982. [Google Scholar]
- Stanisz, A. Easy Course of Statistic Using Statistica PL and Medicine Examples, 1. Basic Statistic; StatSoft Polska: Kraków, Poland, 2006. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier Science BV: Amsterdam, The Netherlands, 1998; p. 853. [Google Scholar]
- Leps, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003; p. 269. [Google Scholar]
- Dell. STATISTICA 13.3; Dell: Round Rock, TX, USA, 2020. [Google Scholar]
- Kovach Computing Services. MVSP: A Multivariate Statistical Package for Windows, Ver. 3.0; Kovach Computing Services: Wales, UK, 2011. [Google Scholar]
- Microsoft. MS Excel 365 Software; Microsoft: Redmond, WA, USA, 2020. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Seniczak, S.; Graczyk, R.; Seniczak, A.; Faleńczyk-Koziróg, K.; Kaczmarek, S.; Marquardt, T. Microhabitat preferences of Oribatida and Mesostigmata (Acari) inhabiting lowland beech forest in Poland and the trophic interactions between these mites. Eur. J. Soil Biol. 2018, 87, 25–32. [Google Scholar] [CrossRef]
- Harris, J.R.W.; Usher, M.B. Laboratory studies of predation by the grassland mite Pergamasus longicornis, Berlese and their possible implications for the dynamics of populations of Collembola. Sci. Proc. R. Dublin Soc. A 1976, 6, 143–153. [Google Scholar]
- Rajski, A. Faunistic-ecological investigations on moss mites (Acari, Oribatei) in several plant associations. I. Ecol. Bull. Soc. Amis. Sc. Lettr. Poznań Ser. B 1961, 25, 123–283. [Google Scholar]
- Niedbała, W. Mechowce—Roztocze Ekosystemów Lądowych; PWN: Warszawa, Poland, 1980; pp. 1–255. [Google Scholar]
- Schatz, H.; Behan-Pelletier, V.M. Global diversity of oribatids (Oribatida; Acari—Arachnida). Hydrobiologia 2008, 595, 323–328. [Google Scholar] [CrossRef]
- Mumladze, L.; Murvanidze, M.; Behan-Pelletier, V. Compositional patterns in Holarctic peat bog inhabiting oribatid mite (Acari: Oribatida) communities. Pedobiologia 2013, 56, 41–48. [Google Scholar] [CrossRef]
- Niedbała, W. Phthiracaroidea (Acari, Oribatida): Systematic Studies; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1992. [Google Scholar]
- Hågvar, S. Mites (Acari) developing inside decomposing spruce needles: Biology and effect on decomposition rate. Pedobiologia 1998, 42, 358–377. [Google Scholar]
- Weigmann, G. Oribatid communities in transects from bogs to forests in Berlin indicating the biotope qualities. In Modern Acarology; Dusbabek, F., Bukva, V., Eds.; Academia: Prague, Czech Republic, 1991; Volume 1, pp. 359–364. [Google Scholar]
- Niedbała, W.; Błoszyk, J.; Gutowski, J.M.; Konwerski, S.; Napierała, A. A characteristic of a community of ptyctimous mites (Acari: Oribatida) in the Białowieża Primeval Forest, Central Europe. In Mites (Acari) of the Białowieża Primeval Forest; Błoszyk, J., Napierała, A., Eds.; Wydawnictwo Kontekst: Poznan, Poland, 2020; pp. 61–87. [Google Scholar]
- Lehmitz, R.; Maraun, M. Small-scale spatial heterogeneity of stable isotopes signatures (δ15N, δ13C) in Sphagnum sp. transfers to all trophic levels in oribatid mites. Soil Biol. Biochem. 2016, 100, 242–251. [Google Scholar] [CrossRef]
- Behan-Pelletier, V.M.; Bisset, B. Oribatida of Canadian peatlands. Mem. Entomol. Soc. Can. 1994, 126, 73–88. [Google Scholar] [CrossRef]
- Salmane, I.; Brumelis, G. Species list and habitat preference of Mesostigmata mites (Acari, Parasitiformes) in Latvia. Acarologia 2010, 50, 373–394. [Google Scholar] [CrossRef]
- Bolger, T.; Arroyo, J.; Piotrowska, K. A catalogue of the species of Mesostigmata (Arachnida, Acari, Parasitiformes) recorded from Ireland including information on their geographical distribution and habitats. Zootaxa 2018, 4519, 1–220. [Google Scholar] [CrossRef] [PubMed]
- Wisdom, R.; Arroyo, J.; Bolger, T. A survey of the Oribatida and Mesostigmata (Acarina) of Irish peatlands. Bull. Irish Biogeogr. Soc. 2011, 35, 130–149. [Google Scholar]
- Heinrichs, S.; Winterhoff, W.; Schmidt, W. Vegetation dynamics of beech forests on limestone in central Germany over half a century—Effects of climate change, forest management, eutrophication or game browsing? Biodivers. Ecol. 2012, 4, 49–61. [Google Scholar] [CrossRef]
- Weigmann, G.; Horak, F.; Franke, K.; Christian, A. Verbreitung und Ökologie der Hornmilben (Oribatida) in Deutschland; Senckenberg Museum Für Naturkunde Görlitz: Görlitz, Germany, 2015; Volume 10, pp. 1–171. [Google Scholar]
- Dalenius, P. The oribatid fauna of South Sweden with remarks concerning its ecology and zoogeography. Kgl. Fysiogr. Sällsk. Lund. Förh. 1950, 20, 30–48. [Google Scholar]
- Moritz, M. Neue Oribatiden (Acari) aus Deutschland III: Suctobelba carcharodon n. sp. und Suctobelba prominens n. sp. Zool. Anz. 1966, 177, 276–282. [Google Scholar]
- Moritz, M.; Broen, B.V.; Messner, B.; Mohrig, W. Beiträge zur Arthropodenfauna aus Großhöhlen des Harzes und des Kyffhäusers. V. Oribatei (Acarina). Deutsche Entomol. Z.N.F. 1971, 18, 133–144. [Google Scholar] [CrossRef]
- Miko, L. Faunistic records of Slovak oribatids II. Oribatid mites (Acarina) of Busov Mountains, North-East Slovakia. Folia Faun. Slov. 2016, 21, 217–220. [Google Scholar]
- Subías, L.S. Los ácaros oribátidos (Acari, Oribatida) de la Sierra de Albarracín (noreste de España). Rev. Ibérica Aracnol. 2015, 26, 81–84. [Google Scholar]
- Starý, J. Půdní roztoči—pancířníci (Oribatida)—Soil mites—oribatid mites. In Grasslands of the White Carpatian Mountains; Jongepierova, I., Ed.; ZO CSOP Bílé Karpaty: Veselí nad Moravau, Czech Republic, 2008; pp. 210–212. [Google Scholar]
- Niemi, R.; Skubała, P. New species of Moritzoppia and Medioppia from the Beskidy Mountains, Poland (Acarina, Oribatida, Oppiidae). Entomol. Fenn. 1993, 4, 195–200. [Google Scholar] [CrossRef]
- Skubała, P.; Gurgul, B. Importance of tree hollows for biodiversity of mites (Acari) in the forest reserve “Śrubita” (Carpathian Mountains, south Poland). Biol. Lett. 2011, 48, 97–106. [Google Scholar] [CrossRef]
- Mahunka, S.; Mahunka-Papp, L. Faunistical and taxonomical studies on oribatids collected in Albania (Acari: Oribatida), I. Opusc. Zool. Budapest 2006, 37, 43–62. [Google Scholar]
- Håpnes, A. Background Note: Natural Forest Heritage in Norway. WWF Norway. 2003. Available online: http://wwf.panda.org/wwf_news/?6748/Background-paper-Natural-forest-heritage-in-Norway (accessed on 8 June 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order/Suborder and Family | Species | Abbreviations for DCA Analysis | Microhabitat A | Microhabitat B | ANOVA Rang Kruskal-Wallis | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Oribatida | A | C | D | A | C | D | H | p | ||
| Brachychthoniidae Thor, 1934 | Liochthonius brevis (Michael, 1888) | 0.0 | 1.3 | 17 | 0.07 | 0.83 | ns | |||
| L. neglectus Moritz, 1976 | L.neg | 0.0 | 39.5 | 33 | 1.97 | 1.83 | ns | |||
| L. tuxeni (Forsslund, 1957) | 0.6 | 20 | 0.07 | 0.2 | 17 | 0.01 | 0.07 | ns | ||
| Neobrachychthonius magnus Moritz, 1976 | N.mag | 0.0 | 46.0 | 67 | 2.29 | 4.47 | 0.035 | |||
| Sellnickochthonius jacoti (Evans, 1952) | 0.0 | 0.7 | 17 | 0.03 | 0.83 | ns | ||||
| Eniochthoniidae Grandjean, 1947 | Eniochthonius minutissimus (Berlese, 1904) | 0.2 | 20 | 0.02 | 2.2 | 17 | 0.11 | 0.00 | ns | |
| Hypochthoniidae Berlese, 1910 | Hypochthonius rufulus C.L. Koch, 1835 | 2.2 | 40 | 0.27 | 0.8 | 17 | 0.04 | 0.66 | ns | |
| Euphthiracaridae Jacot, 1930 | Acrotritia ardua (C.L. Koch, 1841) | A.ard | 8.6 | 100 | 1.04 | 0.0 | 8.97 | 0.003 | ||
| A. duplicata (Grandjean, 1953) | 0.0 | 0.2 | 17 | 0.01 | 0.83 | ns | ||||
| Euphthiracarus cribrarius (Berlese, 1904) | 0.2 | 20 | 0.02 | 0.7 | 33 | 0.03 | 0.49 | ns | ||
| Phthiracaridae Perty, 1841 | Phthiracarus anonymus Grandjean, 1933 | 2.8 | 40 | 0.34 | 0.2 | 17 | 0.01 | 1.09 | ns | |
| P. bryobius Jacot, 1930 | 2.0 | 80 | 0.24 | 1.3 | 67 | 0.07 | 0.31 | ns | ||
| P. clavatus Parry, 1979 | 0.0 | 1.5 | 17 | 0.07 | 0.83 | ns | ||||
| P. crinitus (C.L. Koch, 1841) | 0.2 | 20 | 0.02 | 0.0 | 1.20 | ns | ||||
| P. laevigatus (C.L. Koch, 1841) | 0.2 | 20 | 0.02 | 0.0 | 1.20 | ns | ||||
| P. longulus (C.L. Koch, 1841) | 0.4 | 40 | 0.05 | 3.5 | 83 | 0.17 | 3.31 | ns | ||
| Steganacaridae Niedbała, 1986 | Atropacarus striculus (C.L. Koch, 1835) | A.str | 115.8 | 100 | 14.02 | 0.0 | 8.92 | 0.003 | ||
| Steganacarus magnus (Nicolet, 1855) | 3.4 | 60 | 0.41 | 0.2 | 17 | 0.01 | 2.53 | ns | ||
| S. spinosus (Sellnick, 1920) | 0.6 | 40 | 0.07 | 8.8 | 83 | 0.44 | 3.51 | ns | ||
| Crotoniidae Thorell, 1876 | Camisia biurus (C.L. Koch, 1839) | 0.0 | 2.0 | 33 | 0.10 | 1.83 | ns | |||
| C. spinifer (C.L. Koch, 1836) | 0.0 | 0.2 | 17 | 0.01 | 0.83 | ns | ||||
| Platynothrus peltifer (C.L. Koch, 1839) | P.pel | 19.0 | 80 | 2.30 | 0.2 | 17 | 0.01 | 5.24 | 0.022 | |
| Malaconothridae Berlese, 1916 | Malaconothrus monodactylus (Michael, 1888) | M.mon | 71.2 | 60 | 8.62 | 0.0 | 4.37 | 0.037 | ||
| Tyrphonothrus maior (Berlese, 1910) | T.mai | 98.6 | 100 | 11.94 | 0.00 | 8.92 | 0.003 | |||
| Nanhermanniidae Sellnick, 1928 | Nanhermannia coronata Berlese, 1913 | 6.4 | 80 | 0.78 | 0.0 | 6.44 | 0.011 | |||
| Nothridae Berlese, 1896 | Nothrus silvestris Nicolet, 1855 | 0.0 | 2.7 | 67 | 0.13 | 4.47 | 0.035 | |||
| Damaeidae Berlese, 1896 | Damaeus clavipes (Hermann, 1804) | 0.0 | 0.2 | 17 | 0.01 | 0.83 | ns | |||
| D. gracilipes (Kulczynski, 1902) | 0.0 | 0.3 | 17 | 0.02 | 0.83 | ns | ||||
| Porobelba spinosa (Sellnick, 1920) | 0.0 | 8.7 | 83 | 0.43 | 6.19 | 0.013 | ||||
| Cepheusidae Berlese, 1896 | Cepheus cepheiformis (Nicolet, 1855) | 0.0 | 0.5 | 17 | 0.02 | 0.83 | ns | |||
| Caleremaeidae Grandjean, 1965 | Caleremaeus monilipes (Michael, 1882) | C.mon | 0.0 | 59.8 | 33 | 2.98 | 1.83 | ns | ||
| Eremaeidae Oudemans, 1900 | Eueremaeus silvestris (Forsslund, 1956) | 0.0 | 1.0 | 33 | 0.05 | 1.83 | ns | |||
| E. valkanovi (Kunst, 1957) | 0.0 | 18.8 | 50 | 0.94 | 3.03 | ns | ||||
| Astegistidae Balogh, 1961 | Cultroribula bicultrata (Berlese, 1905) | 0.0 | 0.3 | 33 | 0.02 | 1.85 | ns | |||
| Furcoribula furcillata (Nordenskiöld, 1901) | 1.3 | 33 | 0.07 | 1.83 | ns | |||||
| Liacaridae Sellnick, 1928 | Adoristes ovatus (C.L. Koch, 1839) | 0.8 | 60 | 0.10 | 4.2 | 67 | 0.21 | 0.89 | ns | |
| Liacarus coracinus (C.L. Koch, 1841) | 0.0 | 13.2 | 33 | 0.66 | 1.83 | ns | ||||
| Carabodidae C.L. Koch, 1843 | Carabodes areolatus Berlese, 1916 | C.are | 0.2 | 20 | 0.02 | 40.5 | 83 | 2.01 | 4.53 | 0.033 |
| C. coriaceus C.L. Koch, 1835 | 0.0 | 0.7 | 17 | 0.03 | 0.83 | ns | ||||
| C. femoralis (Nicolet, 1855) | 0.0 | 9.5 | 33 | 0.47 | 1.83 | ns | ||||
| C. labyrinthicus (Michael, 1879) | C.lab | 1.0 | 60 | 0.12 | 24.8 | 100 | 1.24 | 7.57 | 0.006 | |
| C. marginatus (Michael, 1884) | C.mar | 0.0 | 24.7 | 17 | 1.23 | 0.83 | ns | |||
| C. ornatus Storkán, 1925 | 0.2 | 20 | 0.02 | 5.8 | 83 | 0.29 | 4.07 | 0.044 | ||
| C. rugosior Berlese, 1916 | 0.2 | 20 | 0.02 | 1.20 | ns | |||||
| C. tenuis Forsslund, 1953 | 0.0 | 3.3 | 17 | 0.17 | 0.83 | ns | ||||
| C. willmanni Bernini, 1975 | C.wil | 0.0 | 37.2 | 33 | 1.85 | 1.83 | ns | |||
| Odontocepheus elongatus (Michael, 1879) | 0.0 | 5.7 | 83 | 0.28 | 6.23 | 0.013 | ||||
| Autognetidae Grandjean, 1960 | Autogneta longilamellata (Michael, 1885) | 0.0 | 0.2 | 17 | 0.01 | 0.83 | ns | |||
| Conchogneta dalecarlica (Forsslund, 1947) | 0.0 | 2.2 | 67 | 0.11 | 4.50 | 0.034 | ||||
| Oppiidae Sellnick, 1937 | Dissorhina ornata (Oudemans, 1900) | 0.0 | 14.3 | 100 | 0.71 | 8.25 | 0.004 | |||
| Graptoppia foveolata (Paoli, 1908) | G.fov | 0.0 | 35.8 | 50 | 1.78 | 3.03 | ns | |||
| Lauroppiabeskidyensis(Niemi et Skubala, 1993) | 0.0 | 0.2 | 17 | 0.01 | 0.83 | ns | ||||
| Microppia minus (Paoli, 1908) | 0.0 | 0.5 | 50 | 0.02 | 3.13 | ns | ||||
| Moritzoppia keilbachi (Moritz, 1969) | M.kei | 0.0 | 26.8 | 83 | 1.33 | 6.19 | 0.013 | |||
| M.translamellata (Willmann, 1923) | M.tra | 15.6 | 20 | 1.89 | 0.0 | 1.20 | ns | |||
| Oppiella falcata (Paoli, 1908) | O.fal | 0.0 | 465.5 | 100 | 23.16 | 8.25 | 0.004 | |||
| O. neerlandica (Oudemans, 1900) | O.nee | 60.0 | 100 | 7.27 | 0.0 | 8.92 | 0.003 | |||
| O. nova (Oudemans, 1902) | O.nov | 47.6 | 100 | 5.76 | 67.7 | 100 | 3.37 | 0.21 | ns | |
| O. propinqua Mahunka et Mahunka-Papp, 2000 | O.pro | 38.8 | 100 | 4.70 | 0.0 | 8.92 | 0.003 | |||
| O. splendens (C.L. Koch, 1841) | O.spl | 0.0 | 215.2 | 83 | 10.70 | 6.19 | 0.013 | |||
| O.uliginosa (Willmann, 1919) | 0.7 | 33 | 0.03 | 1.83 | ns | |||||
| Rhinoppia subpectinata (Oudemans, 1900) | R.sub | 0.0 | 24.0 | 50 | 1.19 | 3.03 | ns | |||
| Quadroppiidae Balogh, 1983 | Quadroppia monstruosa (Hammer, 1979) | 0.0 | 3.3 | 50 | 0.17 | 3.03 | ns | |||
| Q. quadricarinata (Michael, 1885) | Q.qua | 0.0 | 48.8 | 100 | 2.43 | 8.25 | 0.004 | |||
| Thyrisomidae Grandjean, 1953 | Banksinoma lanceolata (Michael, 1885) | 0.6 | 40 | 0.07 | 0.2 | 17 | 0.01 | 0.87 | ns | |
| Suctobelbidae Jacot, 1938 | Suctobelba regia Moritz, 1970 | 0.0 | 13.2 | 83 | 0.66 | 6.19 | 0.013 | |||
| S. trigona (Michael, 1888) | 0.0 | 2.0 | 17 | 0.10 | 0.83 | ns | ||||
| Suctobelbella lobata (Strenzke, 1950) | 0.0 | 1.0 | 17 | 0.05 | 0.83 | ns | ||||
| S. carcharodon (Moritz, 1966) | 0.0 | 0.8 | 17 | 0.04 | 0.83 | ns | ||||
| S. falcata (Forsslund, 1941) | S.fal | 0.0 | 42.7 | 33 | 2.12 | 1.83 | ns | |||
| S. sarekensis (Forsslund, 1941) | 0.0 | 18.7 | 50 | 0.93 | 3.03 | ns | ||||
| S. similis (Forsslund, 1941) | 0.0 | 0.3 | 17 | 0.02 | 0.83 | ns | ||||
| Suctobelbella sp. 1 | 0.0 | 2.7 | 17 | 0.13 | 0.83 | ns | ||||
| S. subcornigera (Forsslund, 1941) | S.sbc | 2.6 | 60 | 0.31 | 87.8 | 100 | 4.37 | 6.13 | 0.013 | |
| S. subtrigona (Oudemans, 1900) | S.sbt | 0.0 | 75.8 | 67 | 3.77 | 4.47 | 0.035 | |||
| Tectocepheidae Grandjean, 1954 | Tectocepheus velatus (Michael, 1880) | T.vel | 16.6 | 100 | 2.01 | 302.0 | 100 | 15.02 | 7.50 | 0.006 |
| Licneremaeidae Grandjean, 1954 | Licneremaeus licnophorus (Michael, 1882) | 0.0 | 0.7 | 17 | 0.03 | 0.83 | ns | |||
| Phenopelopidae Petrunkevich, 1955 | Eupelops plicatus (C.L. Koch, 1835) | 0.2 | 20 | 0.02 | 0.7 | 50 | 0.03 | 1.15 | ns | |
| E. torulosus (C.L. Koch, 1839) | 1.0 | 60 | 0.12 | 10.5 | 100 | 0.52 | 7.11 | 0.008 | ||
| Achipteriidae Thor, 1929 | Achipteria magna (Sellnick, 1928) | A.mag | 0.0 | 63.2 | 67 | 3.14 | 4.47 | 0.035 | ||
| A. nitens (Nicolet, 1855) | 0.0 | 5.3 | 33 | 0.27 | 1.83 | ns | ||||
| Parachipteria fanzagoi (Jacot, 1929) | P.fan | 278.8 | 100 | 33.76 | 1.5 | 33 | 0.07 | 7.86 | 0.005 | |
| Oribatellidae Jacot, 1925 | Oribatella quadricornuta Michael, 1880 | 0.0 | 0.2 | 17 | 0.01 | 0.83 | ns | |||
| Ophidiotrichus tectus (Michael, 1884) | 0.0 | 0.2 | 17 | 0.01 | 0.83 | ns | ||||
| Haplozetidae Grandjean, 1936 | Lagenobates lagenulus (Berlese, 1904) | 0.0 | 0.2 | 17 | 0.01 | 0.83 | ns | |||
| Oribatulidae Thor, 1929 | Oribatula exilis (Nicolet, 1855) | O.exi | 0.0 | 85.2 | 100 | 4.24 | 8.25 | 0.004 | ||
| O. tibialis (Nicolet, 1855) | 0.0 | 2.2 | 50 | 0.11 | 3.03 | ns | ||||
| Parakalummidae Grandjean, 1936 | Neoribates aurantiacus (Oudemans, 1914) | 0.0 | 0.3 | 17 | 0.02 | 0.83 | ns | |||
| Scheloribatidae Grandjean, 1933 | Scheloribates initialis (Berlese, 1908) | 3.4 | 60 | 0.41 | 4.0 | 67 | 0.20 | 0.08 | ns | |
| S. pallidulus (C.L. Koch, 1841) | 0.0 | 0.5 | 17 | 0.02 | 0.83 | ns | ||||
| Liebstadia longior (Berlese, 1908) | 0.0 | 0.2 | 17 | 0.01 | 0.83 | ns | ||||
| L. similis (Michael, 1888) | 0.0 | 1.0 | 33 | 0.05 | 1.83 | ns | ||||
| Ceratozetidae Jacot, 1925 | Fuscozetes fuscipes (C.L. Koch, 1844) | F.fus | 36.6 | 100 | 4.43 | 0.0 | 8.92 | 0.003 | ||
| Sphaerozetes orbicularis (C.L. Koch, 1835) | 0.0 | 0.25 | 50 | 0.12 | 3.03 | ns | ||||
| Chamobatidae Thor, 1937 | Chamobates borealis Trägårdh, 1902 | 0.22 | 80 | 0.27 | 0.22 | 50 | 0.11 | 0.08 | ns | |
| C. pusillus (Berlese, 1895) | 0.0 | 0.43 | 33 | 0.22 | 1.83 | ns | ||||
| C. rastratus (Hull, 1914) | 0.0 | 0.02 | 17 | 0.01 | 0.83 | ns | ||||
| Galumnidae Jacot, 1925 | Pergalumna nervosa (Berlese, 1914) | 0.0 | 0.03 | 17 | 0.02 | 0.83 | ns | |||
| Mesostigmata | ||||||||||
| Epicriidae Berlese, 1885 | Epicrius mollis (Kramer, 1876) | E.mol | 1.8 | 40 | 4.79 | 0.3 | 17 | 0.64 | 1.09 | ns |
| Zerconidae Berlese, 1892 | Prozercon kochi Sellnick, 1943 | P.koc | 2.2 | 40 | 5.85 | 1.5 | 50 | 2.89 | 0.04 | ns |
| Zercon lindrothi Lundqvist et Johonston, 1986 | Z.lin | 0.0 | 1.5 | 50 | 2.89 | 3.06 | ns | |||
| Z. triangularis C.L. Koch, 1836 | Z.tri | 0.0 | 3.5 | 50 | 6.75 | 3.06 | ns | |||
| Z. zelawaiensis Sellnick, 1944 | Z.zel | 0.0 | 30.2 | 67 | 58.20 | 4.47 | 0.035 | |||
| Macrochelidae Vitzthum, 1930 | Macrocheles opacus (C.L. Koch, 1839) | 0.2 | 20 | 0.53 | 0.0 | 1.20 | ns | |||
| Parasitidae Oudemans, 1901 | Holoparasitus inornatus (Berlese, 1906) | H.ino | 0.0 | 1.2 | 50 | 2.25 | 3.03 | ns | ||
| Paragamasus celticus (Bhattacharyya, 1963) | P.cel | 1.6 | 60 | 4.26 | 0.0 | 4.40 | 0.036 | |||
| P. robustus (Oudemans, 1902) | P.rob | 0.4 | 20 | 1.06 | 0.5 | 50 | 0.96 | 0.41 | ns | |
| P. runcatellus (Berlese, 1903) | P.run | 0.0 | 1.7 | 50 | 3.22 | 3.06 | ns | |||
| P. parrunciger (Bhattacharyya, 1963) | P.par | 16.6 | 40 | 44.15 | 0.0 | 2.64 | ns | |||
| P. lapponicus (Trägårdh, 1910) | P.lap | 5.2 | 60 | 13.83 | 5.5 | 67 | 10.61 | 0.32 | ns | |
| Pergamasus crassipes (Linne, 1758) | P.cra | 2.4 | 60 | 6.38 | 0.5 | 33 | 0.96 | 1.68 | ns | |
| P. septentrionalis (Oudemans, 1902) | 0.2 | 20 | 0.53 | 0.2 | 17 | 0.32 | 0.02 | ns | ||
| Parasitus lunulatus (J. Muller, 1859) | 0.2 | 20 | 0.53 | 0.0 | 1.20 | ns | ||||
| Vulgarogamasus kraepelini (Berlese, 1905) | V.kra | 0.2 | 20 | 0.53 | 1.0 | 17 | 1.93 | 0.00 | ns | |
| Other Parasitidae, juveniles | Paras | 2.4 | 80 | 6.38 | 1.0 | 50 | 1.93 | 0.57 | ns | |
| Veigaiidae Oudemnas, 1939 | Veigaia cerva (Kramer, 1876) | 0.0 | 0.2 | 17 | 0.32 | 0.83 | ns | |||
| V. kochi (Trägårdh, 190 1) | 0.2 | 20 | 0.53 | 0.2 | 17 | 0.32 | 0.02 | ns | ||
| V. nemorensis (C.L. Koch, 1839) | V.nem | 0.2 | 20 | 0.53 | 2.5 | 50 | 4.82 | 1.61 | ns | |
| Ascidae Voigts et Oudemans, 1905 | Asca aphidioides (Linne, 1758) | 0.0 | 0.3 | 17 | 0.64 | 0.83 | ns | |||
| Cheiroseius mutilus (Berlese, 1916) | C.mut | 1.6 | 60 | 4.26 | 0.0 | 4.37 | 0.037 | |||
| Laelapidae Berlese, 1892 | Pachylaelaps dubius Hirschmann et Krauss, 1965 | 0.0 | 0.2 | 17 | 0.32 | 0.83 | ns | |||
| Trachytidae Trägårdh, 1938 | Trachytes aegrota (C.L. Koch, 1841) | T.aeg | 2.0 | 40 | 5.32 | 0.0 | 2.64 | ns | ||
| Uropodidae Kramer, 1881 | Uropoda misella (Berlese, 1916) | 0.2 | 20 | 0.53 | 0.0 | 1.20 | ns | |||
| Species | Specificity | Fidelity | p-Value |
|---|---|---|---|
| A | |||
| A. ardua | 1.00 | 1.00 | 0.01 |
| A. striculus | 1.00 | 1.00 | 0.01 |
| F. fuscipes | 1.00 | 1.00 | 0.01 |
| O. neerlandica | 1.00 | 1.00 | 0.01 |
| O. propinqua | 1.00 | 1.00 | 0.01 |
| P. fanzagoi | 0.99 | 1.00 | 0.01 |
| N. coronata | 1.00 | 0.80 | 0.04 |
| T. maior | 1.00 | 0.80 | 0.05 |
| P. peltifer | 0.98 | 0.80 | 0.04 |
| B | |||
| D. ornata | 1.00 | 1.00 | 0.01 |
| O. falcata | 1.00 | 1.00 | 0.01 |
| Q. quadricartinata | 1.00 | 1.00 | 0.01 |
| O. exilis | 1.00 | 1.00 | 0.01 |
| S. subcornigera | 0.97 | 1.00 | 0.02 |
| C. labyrinthicus | 0.96 | 1.00 | 0.01 |
| E. torulosus | 0.93 | 1.00 | 0.04 |
| M. keilbachi | 1.00 | 0.83 | 0.05 |
| O. elongatus | 1.00 | 0.83 | 0.02 |
| O. splendens | 1.00 | 0.83 | 0.03 |
| P. spinosa | 1.00 | 0.83 | 0.05 |
| S. regia | 1.00 | 0.83 | 0.02 |
| C. areolatus | 0.99 | 0.83 | 0.05 |
| C. ornatus | 0.97 | 0.83 | 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seniczak, A.; Seniczak, S.; Graczyk, R.; Kaczmarek, S.; Jordal, B.H.; Kowalski, J.; Djursvoll, P.; Roth, S.; Bolger, T. A Forest Pool as a Habitat Island for Mites in a Limestone Forest in Southern Norway. Diversity 2021, 13, 578. https://doi.org/10.3390/d13110578
Seniczak A, Seniczak S, Graczyk R, Kaczmarek S, Jordal BH, Kowalski J, Djursvoll P, Roth S, Bolger T. A Forest Pool as a Habitat Island for Mites in a Limestone Forest in Southern Norway. Diversity. 2021; 13(11):578. https://doi.org/10.3390/d13110578
Chicago/Turabian StyleSeniczak, Anna, Stanisław Seniczak, Radomir Graczyk, Sławomir Kaczmarek, Bjarte H. Jordal, Jarosław Kowalski, Per Djursvoll, Steffen Roth, and Thomas Bolger. 2021. "A Forest Pool as a Habitat Island for Mites in a Limestone Forest in Southern Norway" Diversity 13, no. 11: 578. https://doi.org/10.3390/d13110578
APA StyleSeniczak, A., Seniczak, S., Graczyk, R., Kaczmarek, S., Jordal, B. H., Kowalski, J., Djursvoll, P., Roth, S., & Bolger, T. (2021). A Forest Pool as a Habitat Island for Mites in a Limestone Forest in Southern Norway. Diversity, 13(11), 578. https://doi.org/10.3390/d13110578