Spatial and Temporal Patterns in Macroherbivore Grazing in a Multi-Species Tropical Seagrass Meadow of the Great Barrier Reef


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
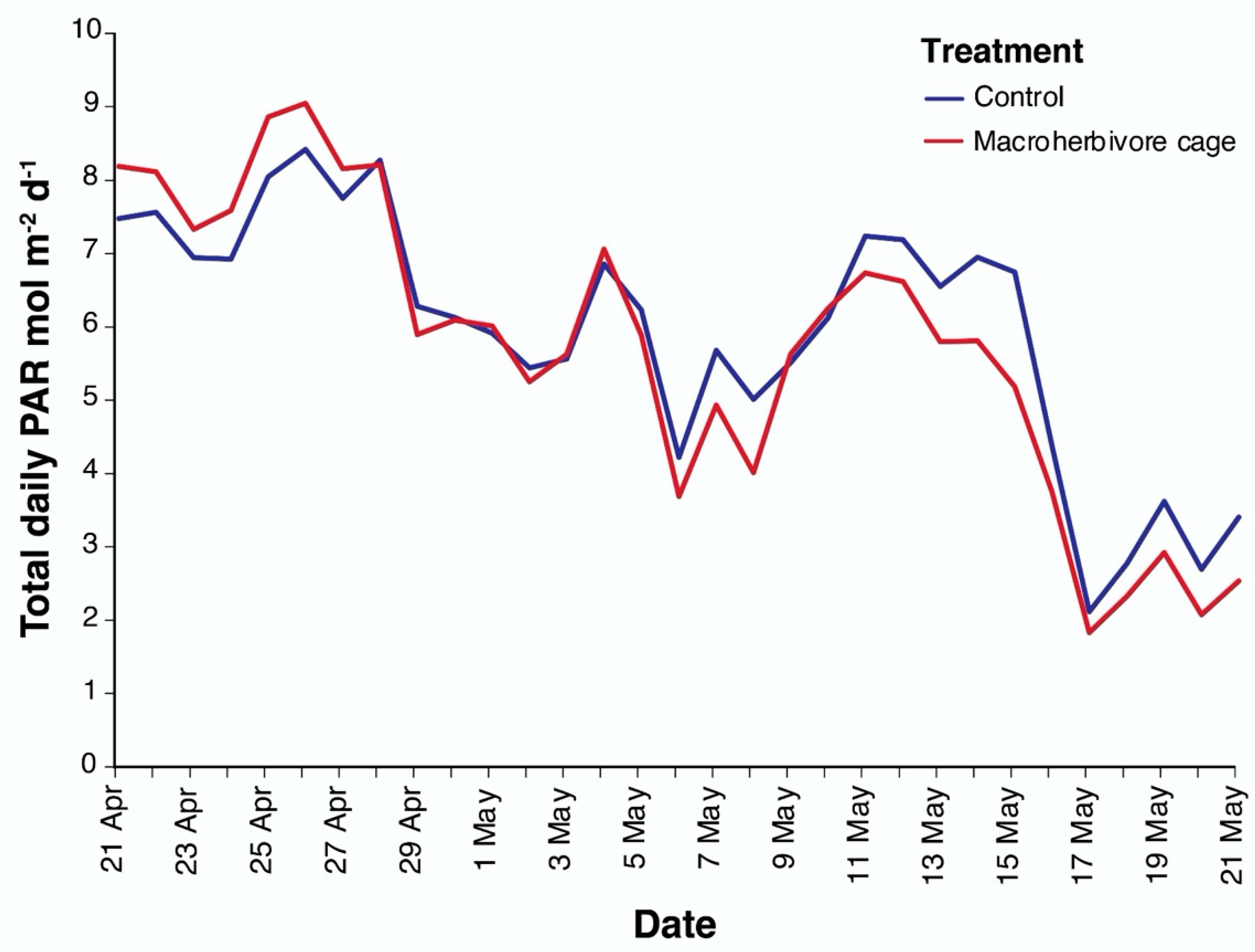
2.2. Exclusion cage Experiments
2.3. Tethering Experiments
2.4. Statistical Analysis
3. Results
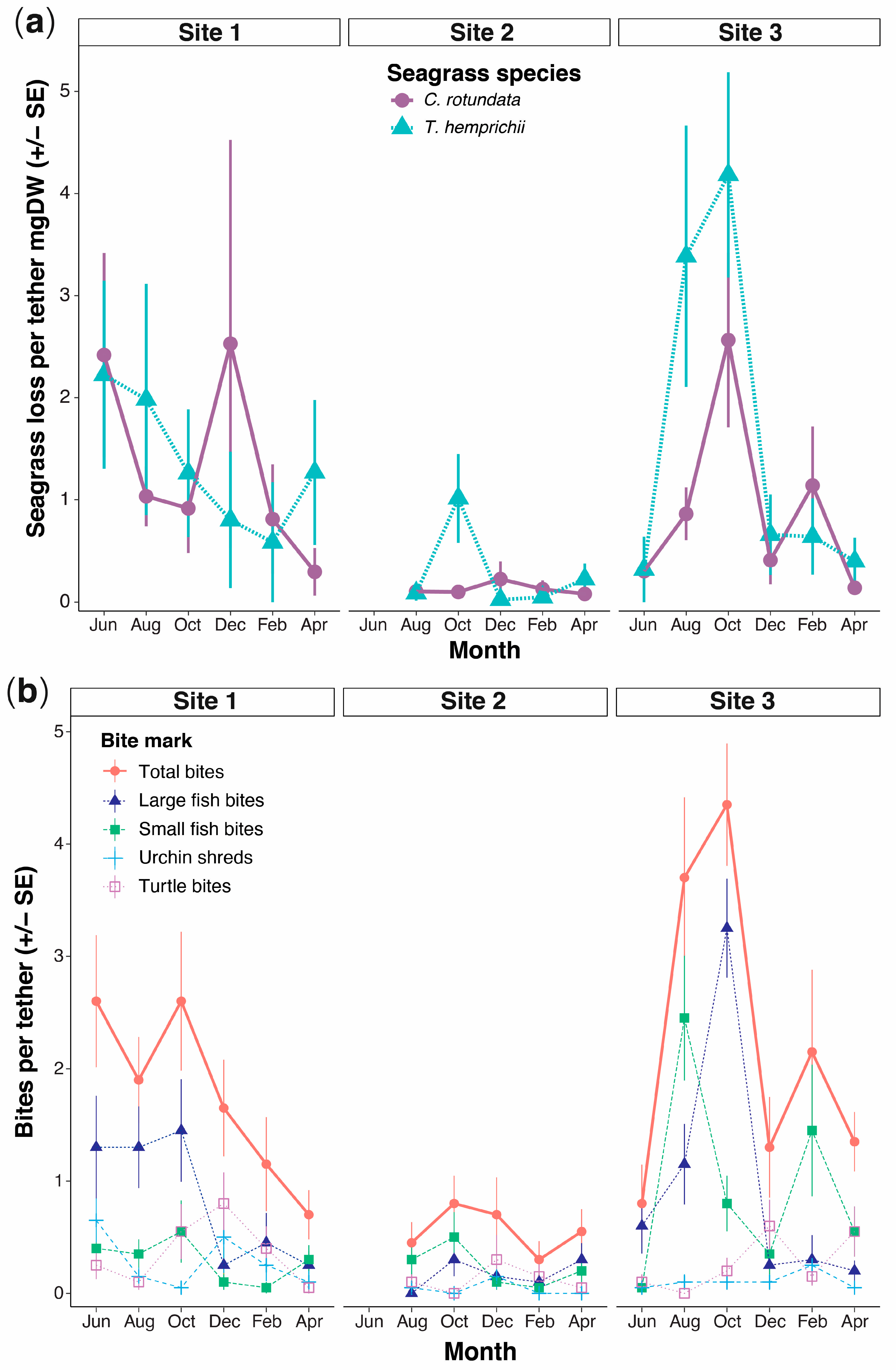
3.1. Rates of Herbivory
3.2. Spatial and Temporal Variability in Herbivory
3.3. Proportion of Productivity Consumed
3.4. Identity of Macroherbivores
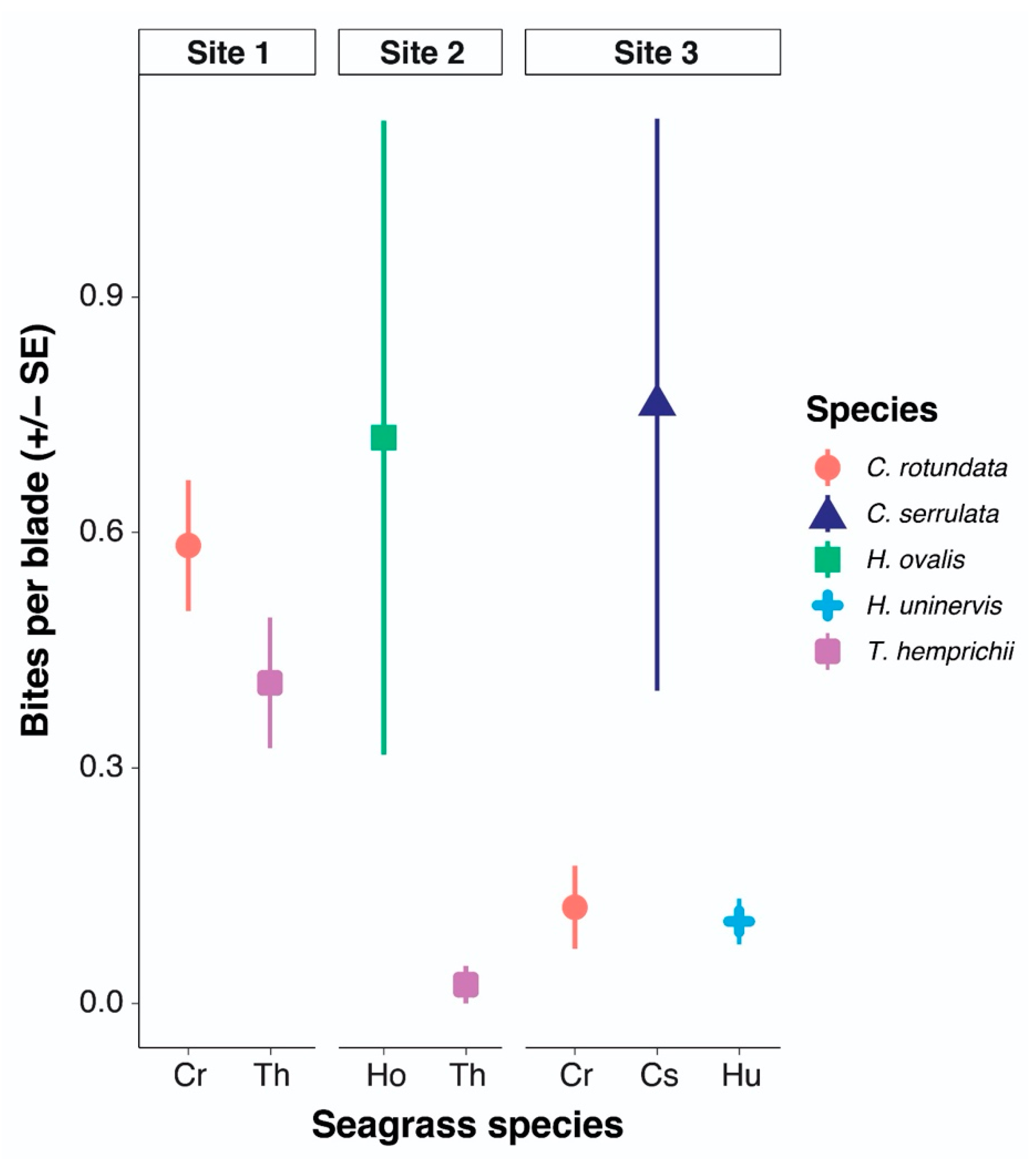
3.5. Species Preference
3.6. Impact of Macroherbivory on the Seagrass Meadow
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | June | August | October | December | February | April | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | |
| Ablennes hians | 11 (4) | 1 (1) | 58 (24) | 1 (1) | 1 (1) | 44 (13) | 1 (1) | 16 (9) | 23 (4) | ||||||||
| Arothron spp. | 1 (1) | 1 (1) | 1 (1) | ||||||||||||||
| Atherinidae | 18 (11) | 1 (1) | |||||||||||||||
| Blenniidae | 3 (1) | 3 (1) | 1 (1) | 2 (2) | |||||||||||||
| Caesionidae | 12 (8) | 3 (1) | 34 (7) | ||||||||||||||
| Carangoides orthogrammus | 1 (1) | ||||||||||||||||
| Caranx ignobilis | 25 (12) | 1 (1) | 1 (1) | ||||||||||||||
| Caranx melampygus | |||||||||||||||||
| Caranx papuensis | 1 (1) | 2 (1) | 1 (1) | 7 (2) | 4 (1) | 2 (1) | 6 (2) | 29 (21) | 1 (1) | 1 (1) | 19 (4) | ||||||
| Carharhinus limbatus | 1(1) | 1(1) | 1 (1) | 1 (1) | 3 (1) | 1 (1) | |||||||||||
| Cheilio inermis | 6 (2) | 15 (3) | 3 (2) | 4 (2) | 7 (3) | ||||||||||||
| Chrysiptera brownriggii | 2 (1) | ||||||||||||||||
| Echeneis naucrates | 1(1) | 1 (1) | 1 (1) | ||||||||||||||
| Gerres oyena | 52 (22) | ||||||||||||||||
| Gnathanodon speciosus | 12 (11) | ||||||||||||||||
| Gobiidae | 2 (2) | 2 (1) | |||||||||||||||
| Labroides spp. | 1 (1) | 1 (1) | |||||||||||||||
| Lethrinid | 5 (3) | 22 (3) | 2 (2) | 2 (2) | 1 (1) | 17 (2) | 12 (2) | 52 (4) | 62 (24) | 4 (1) | 91 (12) | 4 (2) | 1 (1) | 6 (3) | |||
| Lethrinus nebulosus | 2 (1) | 2 (1) | |||||||||||||||
| Lethrinus obsoletus | 2 (1) | ||||||||||||||||
| Myxus elongatus | 30 (13) | 35 (22) | 16 (8) | ||||||||||||||
| Parupeneus spp. | 1(1) | 3 (3) | 20 (5) | 1 (1) | 5 (2) | ||||||||||||
| Pastinachus sephen | 1 (1) | 1 (1) | |||||||||||||||
| Platax orbicularis | 1(1) | 1 (1) | 1 (1) | 1 (1) | |||||||||||||
| Rhinecanthus aculeatus | 1 (1) | ||||||||||||||||
| Siganus lineatus | 1 (1) | 3 (1) | 70 (8) | 30 (17) | 687 (30) | ||||||||||||
| Siganus doliatus | 44 (8) | ||||||||||||||||
| Stethojulis strigiventer | 12 (3) | 4 (2) | 2 (2) | 8 (3) | 87 (7) | 7 (4) | 52 (6) | 4 (3) | |||||||||
| Trachinotus blochii | 1(1) | 1 (1) | 3 (1) | 3 (2) | 2 (1) | 2 (1) | 10 (5) | ||||||||||
References
- Rasheed, M.A.; Dew, K.R.; McKenzie, L.J.; Coles, R.G.; Kerville, S.P.; Campbell, S.J. Productivity, carbon assimilation and intra-annual change in tropical reef platform seagrass communities of the Torres Strait, north-eastern Australia. Cont. Shelf Res. 2008, 28, 2292–2303. [Google Scholar] [CrossRef]
- Duarte, C.M.; Cebrián, J. The fate of marine autotrophic production. Limnol. Oceanogr. 1996, 41, 1758–1766. [Google Scholar] [CrossRef]
- Hyndes, G.A.; Nagelkerken, I.; Mcleod, R.J.; Connolly, R.M.; Lavery, S.; Vanderklift, M.A. Mechanisms and ecological role of carbon transfer within coastal seascapes. Biol. Rev. 2014, 89, 232–254. [Google Scholar] [CrossRef]
- Heck, K.L.; Samsonova, M.; Poore, A.G.B.B.; Hyndes, G.A. Global Patterns in Seagrass Herbivory: Why, Despite Existing Evidence, There Are Solid Arguments in Favor of Latitudinal Gradients in Seagrass Herbivory. Estuaries Coasts 2020. [Google Scholar] [CrossRef]
- Scott, A.L.; York, P.H.; Rasheed, M.A. Green turtle (Chelonia mydas) grazing plot formation creates structural changes in a multi-species Great Barrier Reef seagrass meadow. Mar. Environ. Res. 2020, 162, 105183. [Google Scholar] [CrossRef] [PubMed]
- Aragones, L.; Marsh, H. Impact of dugong grazing and turtle cropping on tropical seagrass communities. Pacific Conserv. Biol. 2000, 5, 286–288. [Google Scholar] [CrossRef]
- Scott, A.L.; York, P.H.; Rasheed, M.A. Herbivory Has a Major Influence on Structure and Condition of a Great Barrier Reef Subtropical Seagrass Meadow. Estuaries Coasts 2020. [Google Scholar] [CrossRef]
- Holzer, K.K.; Seekell, D.A.; McGlathery, K.J. Bucktooth parrotfish Sparisoma radians grazing on Thalassia in Bermuda varies seasonally and with background nitrogen content. J. Exp. Mar. Bio. Ecol. 2013, 443, 27–32. [Google Scholar] [CrossRef]
- Kirsch, K.D.; Valentine, J.F.; Heck, K.L. Parrotfish grazing on turtlegrass Thalassia testudinum: Evidence for the importance of seagrass consumption in food web dynamics of the Florida Keys National Marine Sanctuary. Mar. Ecol. Prog. Ser. 2002, 227, 71–85. [Google Scholar] [CrossRef][Green Version]
- Tomas, F.; Turon, X.; Romero, J. Seasonal and small-scale spatial variability of herbivory pressure on the temperate seagrass Posidonia oceanica. Mar. Ecol. Prog. Ser. 2005, 301, 95–107. [Google Scholar] [CrossRef]
- Lee, C.L.; Huang, Y.H.; Chen, C.H.; Lin, H.J. Remote underwater video reveals grazing preferences and drift export in multispecies seagrass beds. J. Exp. Mar. Bio. Ecol. 2016, 476, 1–7. [Google Scholar] [CrossRef]
- Lee, C.L.; Huang, Y.H.; Chung, C.Y.; Hsiao, S.C.; Lin, H.J. Herbivory in multi-species, tropical seagrass beds. Mar. Ecol. Prog. Ser. 2015, 525, 65–80. [Google Scholar] [CrossRef]
- Tertschnig, W.P. Diel Activity Patterns and Foraging Dynamics of the Sea Urchin Tripneustes ventricosus in a Tropical Seagrass Community and a Reef Environment (Virgin Islands). Mar. Ecol. 1989, 10, 3–21. [Google Scholar] [CrossRef]
- Vonk, J.A.; Pijnappels, M.H.J.; Stapel, J. In situ quantification of Tripneustes gratilla grazing and its effects on three co-occurring tropical seagrass species. Mar. Ecol. Prog. Ser. 2008, 360, 107–114. [Google Scholar] [CrossRef][Green Version]
- Unsworth, R.K.F.; Taylor, J.D.; Powell, A.; Bell, J.J.; Smith, D.J. The contribution of scarid herbivory to seagrass ecosystem dynamics in the Indo-Pacific. Estuar. Coast. Shelf Sci. 2007, 74, 53–62. [Google Scholar] [CrossRef]
- Burnell, O.W.; Connell, S.D.; Irving, A.D.; Russell, B.D. Asymmetric patterns of recovery in two habitat forming seagrass species following simulated overgrazing by urchins. J. Exp. Mar. Bio. Ecol. 2013, 448, 114–120. [Google Scholar] [CrossRef]
- Carnell, P.E.; Ierodiaconou, D.; Atwood, T.B.; Macreadie, P.I. Overgrazing of Seagrass by Sea Urchins Diminishes Blue Carbon Stocks. Ecosystems 2020, 23, 1437–1448. [Google Scholar] [CrossRef]
- Alcoverro, T.; Mariani, S. Effects of sea urchin grazing on seagrass (Thalassodendron ciliatum) beds of a Kenyan lagoon. Mar. Ecol. Prog. Ser. 2002, 226, 255–263. [Google Scholar] [CrossRef]
- Hay, M.E. Patterns of Fish and Urchin Grazing on Caribbean Coral Reefs: Are Previous Results Typical? Ecol. Soc. Am. 1984, 65, 446–454. [Google Scholar] [CrossRef]
- Wressnig, A.; Booth, D.J. Patterns of seagrass biomass removal by two temperate Australian fishes (Monacanthidae). Mar. Freshw. Res. 2008, 59, 408–417. [Google Scholar] [CrossRef]
- Bessey, C.; Heithaus, M.R.; Fourqurean, J.W.; Gastrich, K.R.; Burkholder, D.A. Importance of teleost macrograzers to seagrass composition in a subtropical ecosystem with abundant populations of megagrazers and predators. Mar. Ecol. Prog. Ser. 2016, 553, 81–92. [Google Scholar] [CrossRef]
- Valentine, J.F.; Heck, K.L.; Kirsch, K.D.; Webb, D. Role of sea urchin Lytechinus variegatus grazing in regulating subtropical turtlegrass Thalassia testudinum meadows in the Florida Keys (USA). Mar. Ecol. Prog. Ser. 2000, 200, 213–228. [Google Scholar] [CrossRef][Green Version]
- Cebrián, J.; Duarte, C.M.; Marbà, N. Herbivory on the seagrass Cymodocea nodosa (Ucria) ascherson in contrasting Spanish Mediterranean habitats. J. Exp. Mar. Bio. Ecol. 1996, 204, 103–111. [Google Scholar] [CrossRef]
- Swindells, K.L.; Murdoch, R.J.; Bazen, W.D.; Harman, N.W.; Unsworth, R.K.F. Habitat configuration alters herbivory across the tropical seascape. Front. Mar. Sci. 2017, 4, 48. [Google Scholar] [CrossRef]
- Chiquillo, K.L.; Mims, B.M.; Chong, A.U.; Johnson, C.; Habtes, S.Y.; Fong, C.R.; Barber, P.H.; Fong, P. Herbivory as a limiting factor for seagrass proximity to fringing reefs in Moorea, French Polynesia. Aquat. Bot. 2021, 168, 103294. [Google Scholar] [CrossRef]
- Valentine, J.F.; Jr, K.L.H.; Blackmon, D.; Goecker, M.E.; Christian, J.; Kroutil, R.M.; Peterson, B.J.; Vanderklift, M.A.; Kirsch, K.D.; Beck, M. Exploited species impacts on trophic linkages along reef-seagrass interfaces in the Florida Keys. Ecol. Appl. 2008, 18, 1501–1515. [Google Scholar] [CrossRef] [PubMed]
- Planes, S.; Raventos, N.; Ferrari, B.; Alcoverro, T. Fish herbivory leads to shifts in seagrass Posidonia oceanica investments in sexual reproduction. Mar. Ecol. Prog. Ser. 2011, 431, 205–213. [Google Scholar] [CrossRef]
- Peterson, B.J.; Rose, C.D.; Rutten, L.M.; Fourqurean, J.W. Disturbance and recovery following catastrophic grazing: Studies of a successional chronosequence in a seagrass bed. Oikos 2002, 97, 361–370. [Google Scholar] [CrossRef]
- Eklöf, J.S.; de la Torre-Castro, M.; Gullström, M.; Uku, J.; Muthiga, N.; Lyimo, T.; Bandeira, S.O. Sea urchin overgrazing of seagrasses: A review of current knowledge on causes, consequences, and management. Estuar. Coast. Shelf Sci. 2008, 79, 569–580. [Google Scholar] [CrossRef]
- Gera, A.; Pagès, J.F.; Romero, J.; Alcoverro, T. Combined effects of fragmentation and herbivory on Posidonia oceanica seagrass ecosystems. J. Ecol. 2013, 101, 1053–1061. [Google Scholar] [CrossRef]
- Scott, A.L.; York, P.H.; Duncan, C.; Macreadie, P.I.; Connolly, R.M.; Ellis, M.T.; Jarvis, J.C.; Jinks, K.I.; Marsh, H.; Rasheed, M.A. The Role of Herbivory in Structuring Tropical Seagrass Ecosystem Service Delivery. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Rose, C.D.; Sharp, W.C.; Kenworthy, W.J.; Hunt, J.H.; Lyons, W.G.; Valentine, J.F.; Hall, M.O.; Whitfield, P.E.; Fourqurean, J.W. Overgrazing of a large seagrass bed by the sea urchin Lytechinus variegatus in Outer Florida Bay. Mar. Ecol. Prog. Ser. 1999, 190, 211–222. [Google Scholar] [CrossRef]
- Pagès, J.F.; Farina, S.; Gera, A.; Arthur, R.; Romero, J.; Alcoverro, T. Indirect interactions in seagrasses: Fish herbivores increase predation risk to sea urchins by modifying plant traits. Funct. Ecol. 2012, 26, 1015–1023. [Google Scholar] [CrossRef]
- Heck, K.L.; Valentine, J.F. Sea urchin herbivory: Evidence for long-lasting effects in subtropical seagrass meadows. J. Exp. Mar. Bio. Ecol. 1995, 189, 205–217. [Google Scholar] [CrossRef]
- Bell, S.Y.; Fraser, M.W.; Statton, J.; Kendrick, G.A. Salinity stress drives herbivory rates and selective grazing in subtidal seagrass communities. PLoS ONE 2019, 14, e0214308. [Google Scholar] [CrossRef]
- Mariani, S.; Alcoverro, T. A mulitple-choice feeding-preference experiment utilising seagrasses with a natural population of herbivorous fish. Mar. Ecol. Prog. Ser. 1999, 189, 295–299. [Google Scholar] [CrossRef]
- McGlathery, K.J. Nutrient and grazing influences on a subtropical seagrass community. Mar. Ecol. Prog. Ser. 1995, 122, 239–252. [Google Scholar] [CrossRef]
- Goecker, M.E.; Heck, K.L.; Valentine, J.F. Effects of nitrogen concentrations in turtlegrass Thalassia testudinum on consumption by the bucktooth parrotfish Sparisoma radians. Mar. Ecol. Prog. Ser. 2005, 286, 239–248. [Google Scholar] [CrossRef]
- Prado, P.; Heck, K.L. Seagrass selection by omnivorous and herbivorous consumers: Determining factors. Mar. Ecol. Prog. Ser. 2011, 429, 45–55. [Google Scholar] [CrossRef]
- Ogilvie, P. Marine protected areas in Queensland: Past and present. In Big, Bold and Blue: Lessons from Australia’s Marine Protected Areas; Wescott, G., Fitzsimmons, J., Eds.; CSIRO Publishing: Melbourne, VIC, Australia, 2016; p. 195. [Google Scholar]
- Alcoverro, T.; Mariani, S. Patterns of fish and sea urchin grazing on tropical Indo-Pacific seagrass beds. Ecography (Cop.). 2004, 3, 361–365. [Google Scholar] [CrossRef]
- Finke, D.L.; Denno, R.F. Predator diversity and the functioning of ecosystems: The role of intraguild predation in dampening trophic cascades. Ecol. Lett. 2005, 8, 1299–1306. [Google Scholar] [CrossRef]
- Prado, P.; Farina, S.; Tomas, F.; Romero, J.; Alcoverro, T. Marine protection and meadow size alter fish herbivory in seagrass ecosystems. Mar. Ecol. Prog. Ser. 2008, 371, 11–21. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hoey, A.S.; Choat, J.H. Limited functional redundancy in high diversity systems: Resilience and ecosystem function on coral reefs. Ecol. Lett. 2003, 6, 281–285. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, D.M.; Jones, G.P.; McCook, L.J. Interactions between herbivorous fish guilds and their influence on algal succession on a coastal coral reef. J. Exp. Mar. Bio. Ecol. 2011, 399, 60–67. [Google Scholar] [CrossRef]
- Cheal, A.J.; Emslie, M.J.; Macneil, M.A.; Miller, I.; Sweatman, H.P.A. Spatial variation in the functional characteristics of herbivorous fish communities and the resilience of coral reefs. Ecol. Appl. 2016, 23, 174–188. [Google Scholar] [CrossRef]
- Baxter, I.N. Green Island Information Review. A report to the Great Barrier Reef Marine Park Authority; Great Barrier Reef Marine Park Authority: Townsville, QLD, Australia, 1990. [Google Scholar]
- AIMS AIMS Long-term Monitoring Program: Visual Census Fish Data (Great Barrier Reef). Available online: https://apps.aims.gov.au/metadata/view/5be0b340-4ade-11dc-8f56-00008a07204e (accessed on 27 October 2020).
- Fuentes, M.M.P.B.; Lawler, I.R.; Gyuris, E. Dietary preferences of juvenile green turtles (Chelonia mydas) on a tropical reef flat. Wildl. Res. 2006, 33, 671–678. [Google Scholar] [CrossRef]
- Rasheed, M.A. Recovery and succession in a multi-species tropical seagrass meadow following experimental disturbance: The role of sexual and asexual reproduction. J. Exp. Mar. Biol. Ecol. 2004, 310, 13–45. [Google Scholar] [CrossRef]
- Pitt, J.M. The Feeding Ecology of Rabbitfish (Siganidae) at Green Island Reef: Ontogenetic and Interspecific Differences in Diet, Morphology and Habitat Utilisation. Ph.D. Thesis, James Cook University, Townsville, Australia, 1997. [Google Scholar]
- Kirk, J.T.O. Light and Photosynthesis in Aquatic Ecosystems; Cambridge University Press: Cambridge, UK, 1994; ISBN 9781139168212. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Harvey, E.S.; Cappo, M.; Butler, J.J.; Hall, N.; Kendrick, G.A. Bait attraction affects the performance of remote underwater video stations in assessment of demersal fish community structure. Mar. Ecol. Prog. Ser. 2007, 350, 245–254. [Google Scholar] [CrossRef]
- Zieman, J.C. Methods for the Study of the Growth and Production of Turtle Grass, Thalassia testudinum Konig. Aquaculture 1974, 4, 139–143. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 18 December 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. Available online: https://ggplot2.tidyverse.org/ (accessed on 18 December 2020).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: Cham, Switzerland, 2002. [Google Scholar]
- emmeans: Estimated Marginal Means, aka Least-Squares Means. Available online: https://rdrr.io/cran/emmeans/ (accessed on 18 December 2020).
- Gullström, M.; Berkström, C.; Öhman, M.C.; Bodin, M.; Dahlberg, M. Scale-dependent patterns of variability of a grazing parrotfish (Leptoscarus vaigiensis) in a tropical seagrass-dominated seascape. Mar. Biol. 2011, 158, 1483–1495. [Google Scholar] [CrossRef]
- Cebrián, J.; Duarte, C.M. Patterns in leaf herbivory on seagrasses. Aquat. Bot. 1998, 60, 67–82. [Google Scholar] [CrossRef]
- Loffler, Z.; Bellwood, D.R.; Hoey, A.S. Among-habitat algal selectivity by browsing herbivores on an inshore coral reef. Coral Reefs 2015, 34, 597–605. [Google Scholar] [CrossRef]
- Fox, R.J.; Bellwood, D.R. Remote video bioassays reveal the potential feeding impact of the rabbitfish Siganus canaliculatus (f: Siganidae) on an inner-shelf reef of the Great Barrier Reef. Coral Reefs 2008, 27, 605–615. [Google Scholar] [CrossRef]
- Ferrari, B.; Raventos, N.; Planes, S. Assessing effects of fishing prohibition on Posidonia oceanica seagrass meadows in the Marine Natural Reserve of Cerbère-Banyuls. Aquat. Bot. 2008, 88, 295–302. [Google Scholar] [CrossRef]
- Carroll, J.M.; Stubler, A.D.; Finelli, C.M.; Peterson, B.J. The potential use of seagrass herbivory patterns as an indicator of herbivore community change after tropical marine protected area establishment. Gulf Caribb. Res. 2019, 30, SC1–SC6. [Google Scholar] [CrossRef]
- Duffy, J.E.; Cardinale, B.J.; France, K.E.; McIntyre, P.B.; Thébault, E.; Loreau, M. The functional role of biodiversity in ecosystems: Incorporating trophic complexity. Ecol. Lett. 2007, 10, 522–538. [Google Scholar] [CrossRef]
- Burkholder, D.A.; Heithaus, M.R.; Fourqurean, J.W.; Wirsing, A.; Dill, L.M. Patterns of top-down control in a seagrass ecosystem: Could a roving apex predator induce a behaviour-mediated trophic cascade? J. Anim. Ecol. 2013, 82, 1192–1202. [Google Scholar] [CrossRef]
- Wallner-Hahn, S.; de la Torre-Castro, M.; Eklöf, J.S.; Gullström, M.; Muthiga, N.A.; Uku, J. Cascade effects and sea-urchin overgrazing: An analysis of drivers behind the exploitation of sea urchin predators for management improvement. Ocean Coast. Manag. 2015, 107, 16–27. [Google Scholar] [CrossRef]
- Heithaus, M.R.; Frid, A.; Wirsing, A.J.; Dill, L.M.; Fourqurean, J.W.; Burkholder, D.; Thomson, J.; Bejder, L. State-dependent risk-taking by green sea turtles mediates top-down effects of tiger shark intimidation in a marine ecosystem. J. Anim. Ecol. 2007, 76, 837–844. [Google Scholar] [CrossRef]
- Wirsing, A.J.; Heithaus, M.R.; Dill, L.M. Fear factor: Do dugongs (Dugong dugon) trade food for safety from tiger sharks (Galeocerdo cuvier)? Oecologia 2007, 153, 1031–1040. [Google Scholar] [CrossRef]
- Ruíz, J.M.; Pérez, M.; Romero, J.; Tomas, F. The importance of herbivory in the decline of a seagrass (Posidonia oceanica) meadow near a fish farm: An experimental approach. Bot. Mar. 2009, 52, 449–458. [Google Scholar] [CrossRef]
- Hoffmann, L.; Edwards, W.; York, P.H.; Rasheed, M.A. Richness of primary producers and consumer abundance mediate epiphyte loads in a tropical seagrass system. Diversity 2020, 12, 384. [Google Scholar] [CrossRef]
- Young, M.A.L.; Bellwood, D.R. Diel patterns in sea urchin activity and predation on sea urchins on the Great Barrier Reef. Coral Reefs 2011, 30, 729–736. [Google Scholar] [CrossRef]
- Fox, R.J.; Bellwood, D.R. Unconstrained by the clock? Plasticity of diel activity rhythm in a tropical reef fish, Siganus lineatus. Funct. Ecol. 2011, 25, 1096–1105. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Cross-shelf variation in the role of parrotfishes on the Great Barrier Reef. Coral Reefs 2008, 27, 37–47. [Google Scholar] [CrossRef]
- Valentine, J.F.; Heck, K.L.; Blackmon, D.; Goecker, M.E.; Christian, J.; Kroutil, R.M.; Kirsch, K.D.; Peterson, B.J.; Beck, M.; Vanderklift, M.A. Food web interactions along seagrass-coral reef boundaries: Effects of piscivore reductions on cross-habitat energy exchange. Mar. Ecol. Prog. Ser. 2007, 333, 37–50. [Google Scholar] [CrossRef]
- Randall, J.E. Grazing effect on sea grasses by herbivorous reef fishes in the West Indies. Ecology 1965, 46, 255–260. [Google Scholar]
- Sluka, R.D.; Miller, M.W. Herbivorous fish assemblages and herbivory pressure on Laamu Atoll, Republic of Maldives. Coral Reefs 2001, 20, 255–262. [Google Scholar] [CrossRef]
- Tribble, G.W. Reef-basel herbivores and the distribution of two seagrasses (Syringodium filiforme and Thalassia testudinum) in the San Blas Islands (Western Caribbean). Mar. Biol. 1981, 65, 277–281. [Google Scholar] [CrossRef]
- Jinks, K.I.; Brown, C.J.; Rasheed, M.A.; Scott, A.L.; Sheaves, M.; York, P.H.; Connolly, R.M. Habitat complexity influences the structure of food webs in Great Barrier Reef seagrass meadows. Ecosphere 2019, 10. [Google Scholar] [CrossRef]





| Site | Species Composition | Depth | Mean Aboveground Biomass/gDW m−2 | Mean Shoot Density/m−2 | Habitat | Experiment |
|---|---|---|---|---|---|---|
| 1 | C. rotundata T. hemprichii | 0–2 m | 213.8 | 1605 | In meadow | Cages and tethering |
| 2 | H. ovalis T. hemprichii | 0–2.5 m | 13.1 | 1208 | In reef patch | Cages |
| 2a | C. rotundata T. hemprichii | 0–2.5 m | - | - | In meadow | Tethering |
| 3 | C. rotundata C. serrulata H. uninervis S. isoetifolium T. hemprichii | 1–3 m | 181.3 | 8649 | In meadow | Cages and tethering |
| Site | Season | Species | Productivity | Seagrass Consumption | Productivity Consumed by |
|---|---|---|---|---|---|
| (mg day−1 shoot−1) | (mg day−1 shoot−1) | Macroherbivores | |||
| 1 | Wet | C. rotundata | 1.68 ± 0.56 | 0.21 ± 0.41 | 8–25% |
| T. hemprichii | 1.57 ± 0.56 | 0.15 ± 0.29 | 6–13% | ||
| Dry | C. rotundata | 1.32 ± 0.48 | 0.19 ± 0.28 | 4–30% | |
| T. hemprichii | 1.63 ± 0.93 | 0.30 ± 0.45 | 13–23% | ||
| 2 | Wet | C. rotundata | 1.68 ± 0.56 | 0.02 ± 0.06 | 1–2% |
| T. hemprichii | 1.57 ± 0.56 | 0.06 ± 0.15 | 0.25–11% | ||
| Dry | C. rotundata | 1.32 ± 0.48 | 0.13 ± 0.44 | 1–30% | |
| T. hemprichii | 1.63 ± 0.93 | 0.09 ± 0.26 | 1–13% | ||
| 3 | Wet | C. rotundata | 1.68 ± 0.56 | 0.27 ± 0.36 | 4–25% |
| T. hemprichii | 1.57 ± 0.56 | 0.30 ± 0.39 | 7–44% | ||
| Dry | C. rotundata | 1.32 ± 0.48 | 0.07 ± 0.12 | 2–11% | |
| T. hemprichii | 1.63 ± 0.93 | 0.26 ± 0.46 | 3–33% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scott, A.L.; York, P.H.; Rasheed, M.A. Spatial and Temporal Patterns in Macroherbivore Grazing in a Multi-Species Tropical Seagrass Meadow of the Great Barrier Reef. Diversity 2021, 13, 12. https://doi.org/10.3390/d13010012
Scott AL, York PH, Rasheed MA. Spatial and Temporal Patterns in Macroherbivore Grazing in a Multi-Species Tropical Seagrass Meadow of the Great Barrier Reef. Diversity. 2021; 13(1):12. https://doi.org/10.3390/d13010012
Chicago/Turabian StyleScott, Abigail L., Paul H. York, and Michael A. Rasheed. 2021. "Spatial and Temporal Patterns in Macroherbivore Grazing in a Multi-Species Tropical Seagrass Meadow of the Great Barrier Reef" Diversity 13, no. 1: 12. https://doi.org/10.3390/d13010012
APA StyleScott, A. L., York, P. H., & Rasheed, M. A. (2021). Spatial and Temporal Patterns in Macroherbivore Grazing in a Multi-Species Tropical Seagrass Meadow of the Great Barrier Reef. Diversity, 13(1), 12. https://doi.org/10.3390/d13010012