Facilitation or Competition? Effects of Lions on Brown Hyaenas and Leopards

Abstract
1. Introduction
2. Materials and Methods
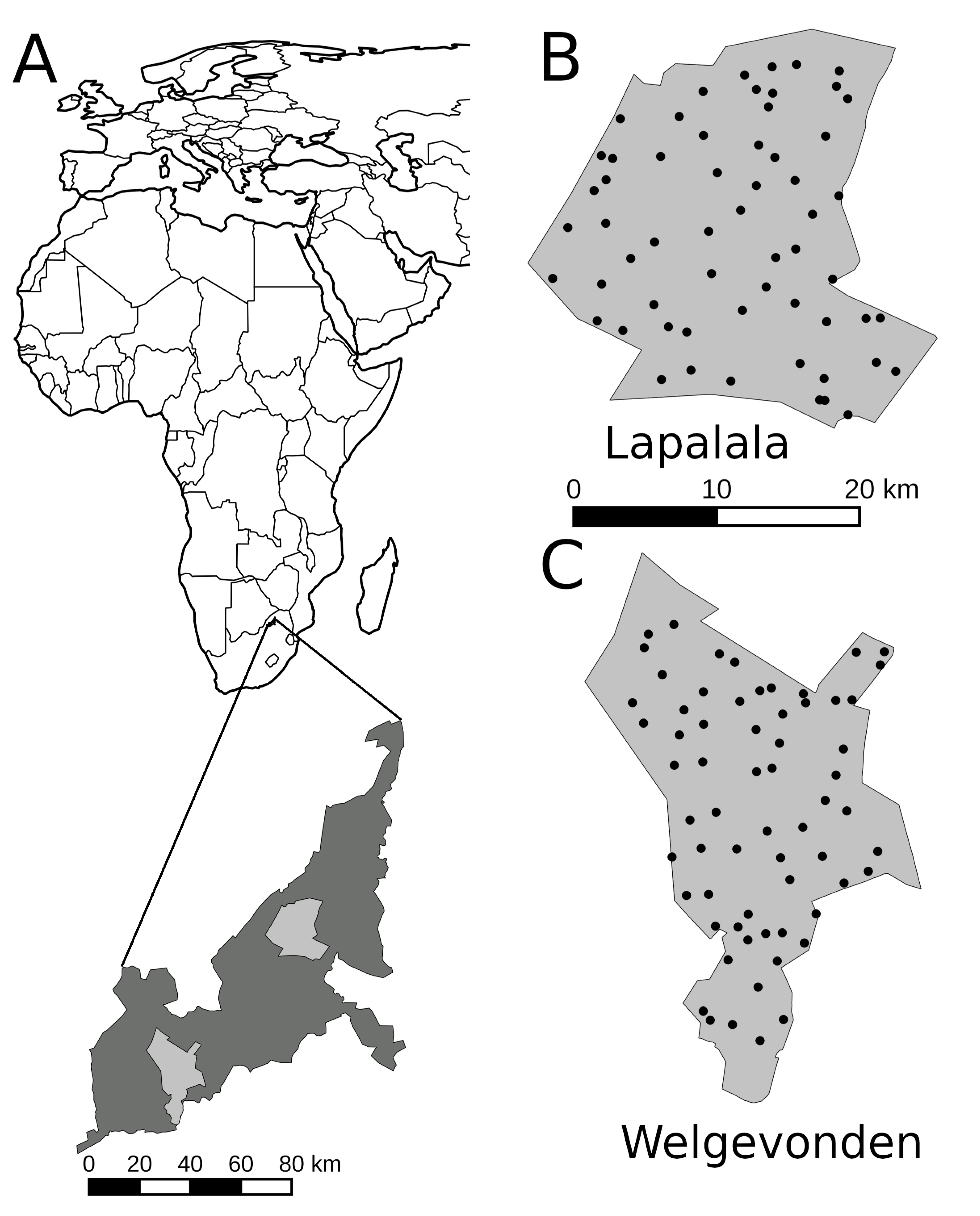
2.1. Study Area
2.2. Camera Surveys
2.3. Estimation of Prey Abundance
2.4. Estimation of Brown Hyaena and Leopard Diet
2.5. Data Analyses
3. Results
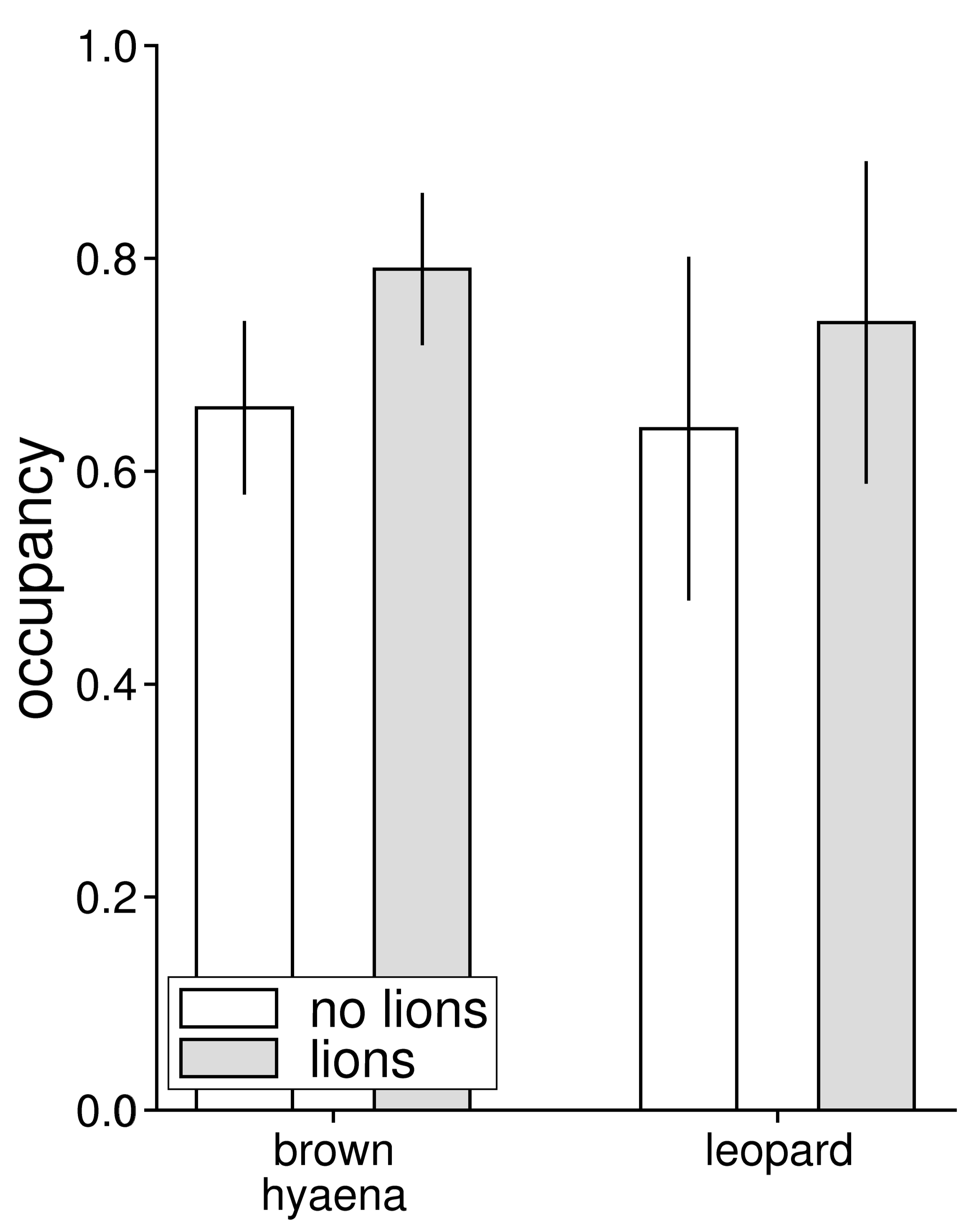
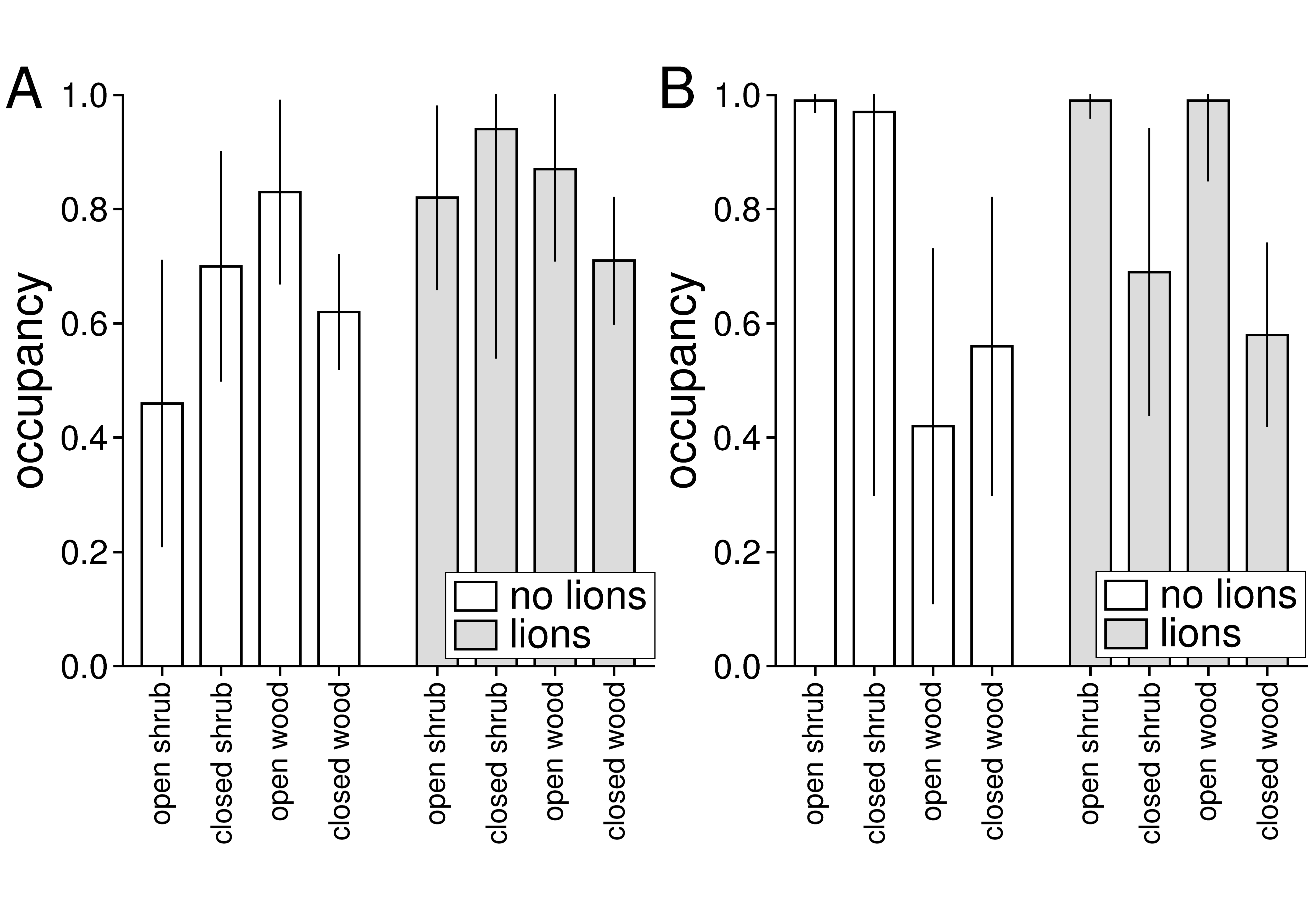
3.1. Relative Abundance and Habitat Use
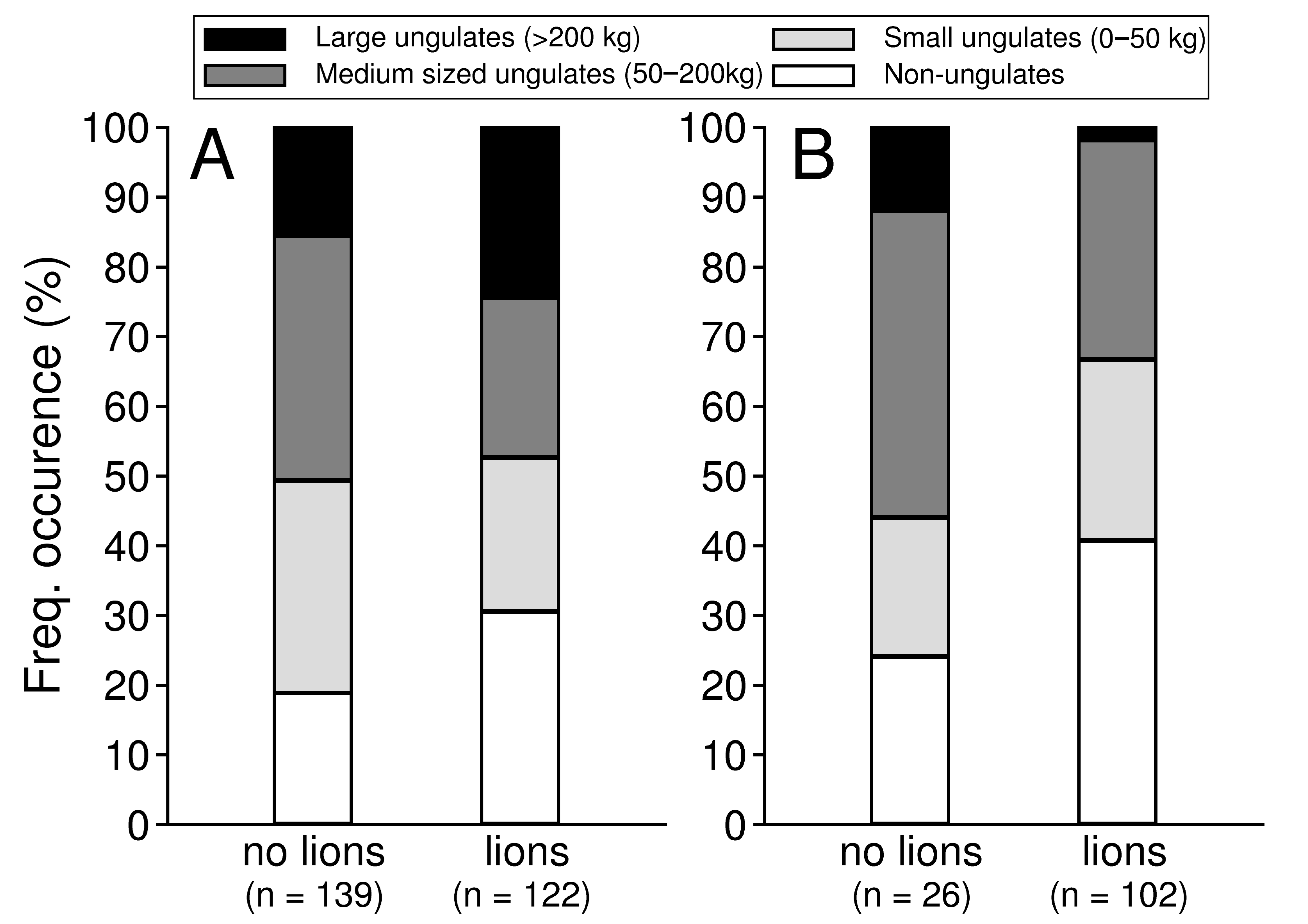
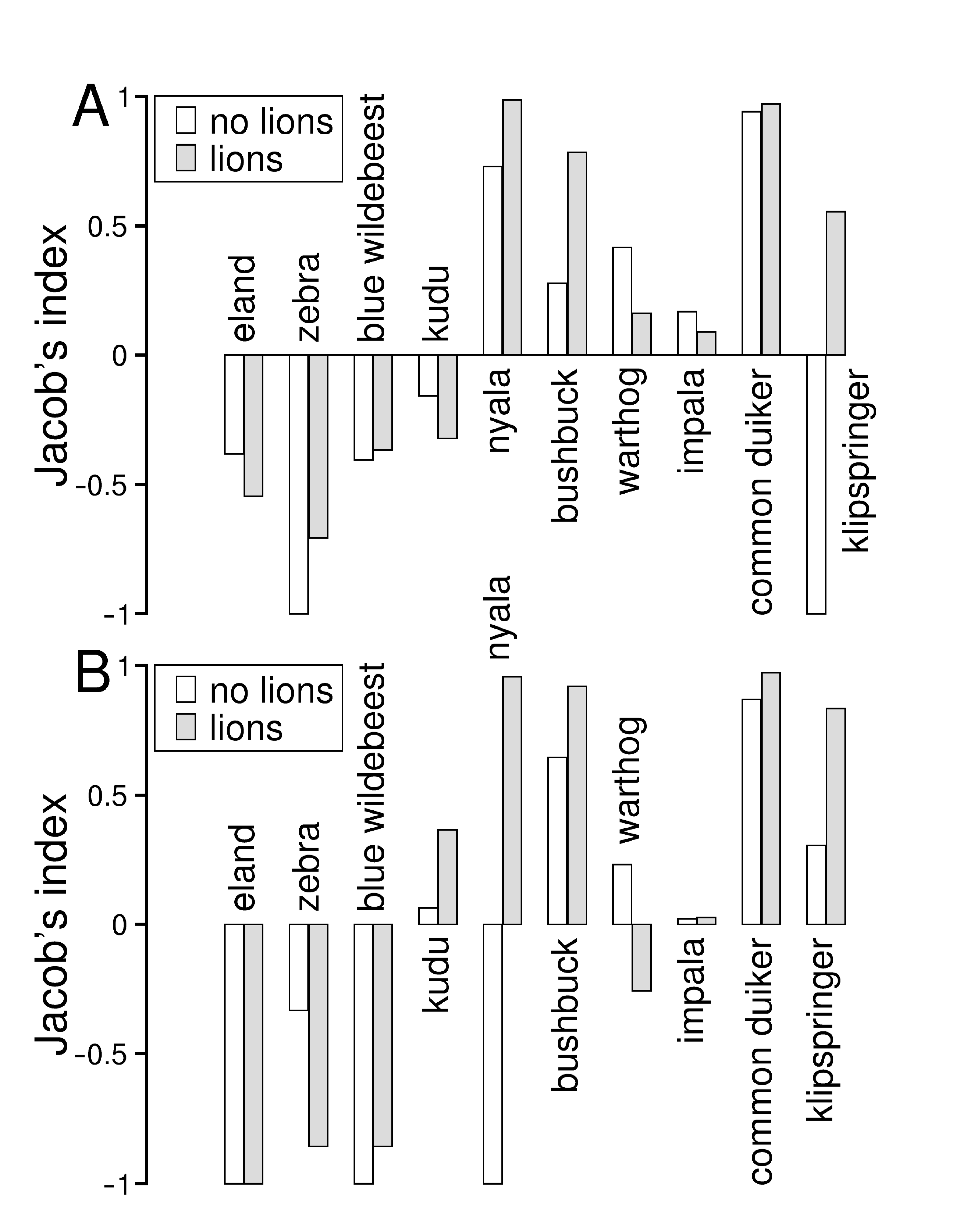
3.2. Diet and Prey Selection
4. Discussions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Latin Name | Lapalala | Welgevonden | |
|---|---|---|---|---|
| 2008 | 2008 | 2009 | ||
| Impala | Aepyceros melampus | 1090 | 695 | 701 |
| Burchell’s zebra | Equus burchellii | 1156 | 472 | 573 |
| Blue wildebeest | Connochaetes taurinus | 409 | 469 | 557 |
| Greater kudu | Tragelaphus strepsiceros | 606 | 183 | 166 |
| Common warthog | Phacochoerus africanus | 293 | 220 | 144 |
| Eland | Taurotragus oryx | 200 | 158 | 156 |
| Waterbuck | Kobus ellipsiprymnus | 206 | 140 | 123 |
| Klipspringer | Oreotragus oreotragus | 123 | 44 | 34 |
| Giraffe | Giraffa camelopardalis | 67 | 30 | 24 |
| Bushbuck | Tragelaphus scriptus | 106 | 6 | 4 |
| Mountain reedbuck | Redunca fulvorufula | 44 | 28 | 20 |
| Common duiker | Sylvicapra grimmia | 35 | 6 | 2 |
| Nyala | Tragelaphus angasii | 30 | 1 | 4 |
| Common Name | Latin Name | Size Class |
|---|---|---|
| Bushbuck | Tragelaphus scriptus | Small (<50kg) |
| Common duiker | Sylvicapra grimmia | Small (<50kg) |
| Klipspringer | Oreotragus oreotragus | Small (<50kg) |
| Greater kudu | Tragelaphus strepsiceros | Medium (50–200kg) |
| Impala | Aepyceros melampus | Medium (50–200kg) |
| Nyala | Tragelaphus angasii | Medium (50–200kg) |
| Common warthog | Phacochoerus africanus | Medium (50–200kg) |
| Blue wildebeest | Connochaetes taurinus | Large (>200kg) |
| Burchell’s zebra | Equus burchellii | Large (>200kg) |
| Eland | Taurotragus oryx | Large (>200kg) |
| Species | Data | Occupancy Covariates | Detection Covariates | Par. | Δ AIC |
|---|---|---|---|---|---|
| Brown hyaena | Both | Reserve x Habitat | Reserve + Habitat | 13 | 0 |
| Brown hyaena | Both | Reserve x Habitat | Reserve x Habitat | 16 | 1.92 |
| Brown hyaena | Both | Reserve x Habitat | None | 9 | 25.35 |
| Brown hyaena | Both | Reserve x Habitat | Reserve | 10 | 27.35 |
| Brown hyaena | Lapalala | Habitat | None | 5 | 0.92 |
| Brown hyaena | Lapalala | Habitat | Habitat | 8 | 0 |
| Brown hyaena | Welgevonden | Habitat | Habitat | 8 | 0 |
| Brown hyaena | Welgevonden | Habitat | None | 5 | 6.40 |
| Leopard | Both | Reserve x Habitat | None | 9 | 2.00 |
| Leopard | Both | Reserve x Habitat | Reserve | 10 | 1.26 |
| Leopard | Both | Reserve x Habitat | Reserve + Habitat | 13 | 0 |
| Leopard | Both | Reserve x Habitat | Reserve x Habitat | 16 | 3.57 |
| Leopard | Lapalala | Habitat | None | 5 | 0 |
| Leopard | Lapalala | Habitat | Habitat | 8 | 5.07 |
| Leopard | Welgevonden | Habitat | Habitat | 8 | 0 |
| Leopard | Welgevonden | Habitat | None | 5 | 7.91 |
References
- Werner, E.E.; Peacor, S.D. A review of trait-mediated indirect interactions in ecological communities. Ecology 2003, 84, 1083–1100. [Google Scholar] [CrossRef]
- Root, R.B. The niche exploitation pattern of the blue-gray gnatcatcher. Ecol. Monogr. 1967, 37, 317–350. [Google Scholar] [CrossRef]
- Simberloff, D.; Dayan, T. The guild concept and the structure of ecological communities. Ann. Rev. Ecol. Syst. 1991, 22, 115–143. [Google Scholar] [CrossRef]
- Keddy, P.A. Competition, 2nd ed.; Springer: New York, NY, USA, 2001. [Google Scholar]
- Stachowicz, J.J. Mutualism, facilitation, and the structure of ecological communities. Bioscience 2001, 51, 235–246. [Google Scholar] [CrossRef]
- Colwell, R.K.; Futuyma, D.J. On the measurement of niche breadth and overlap. Ecology 1971, 52, 567–576. [Google Scholar] [CrossRef]
- Abrams, P. Some comments on measuring niche overlap. Ecology 1980, 61, 44–49. [Google Scholar] [CrossRef]
- Wilmers, C.C.; Crabtree, R.L.; Smith, D.W.; Murphy, K.M.; Getzsource, W.M. Trophic facilitation by introduced top predators: Grey wolf subsidies to scavengers in Yellowstone National Park. J. Anim. Ecol. 2003, 72, 909–916. [Google Scholar] [CrossRef]
- van Valkenburgh, B. Locomotor diversity within past and present guilds of large predatory mammals. Paleobiology 1985, 11, 406–428. [Google Scholar] [CrossRef]
- Grossnickle, D.M. Feeding ecology has a stronger evolutionary influence on functional morphology than on body mass in mammals. Evolution 2020, 74, 610–628. [Google Scholar] [CrossRef]
- Polis, G.A.; Myers, C.A.; Holt, R.D. The ecology and evolution of intraguild predation—Potential competitors that eat each other. Ann. Rev. Ecol. Syst. 1989, 20, 297–330. [Google Scholar] [CrossRef]
- Palomares, F.; Caro, T.M. Interspecific killing among mammalian carnivores. Am. Nat. 1999, 153, 492–508. [Google Scholar] [CrossRef] [PubMed]
- Donadio, E.; Buskirk, S.W. Diet, morphology, and interspecific killing in Carnivora. Am. Nat. 2006, 167, 524–536. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.C. Carnivore biogeography and conservation in the African rainforest: A community perspective. In African Rainforest Ecology and Conservation; Weber, W., White, L.J.T., Vedder, A., Naughton-Treves, L., Eds.; Yale University Press: New Haven, CT, USA, 2001; pp. 214–232. [Google Scholar]
- Woodroffe, R.; Ginsberg, J. King of the beast: Evidence for guild redundancy among large mammalian carnivores. In Large Carnivores and the Conservation of Biodiversity; Ray, J.C., Redford, K.H., Berger, J., Eds.; Island Press: Washington, DC, USA, 2005; pp. 154–175. [Google Scholar]
- Dalerum, F.; Cameron, E.Z.; Kunkel, K.E.; Somers, M.J. Continental patterns of carnivore guild depletions: Implications for prioritizing global carnivore conservation. Biol. Lett. 2009, 5, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Mills, M.G.L. Hyaena brunnea. Mamm. Spec. 1982, 194, 1–5. [Google Scholar] [CrossRef]
- Mills, M.G.L. Kalahari Hyaenas: Comparative Behavioural Ecology of Two Species; Springer: Berlin, Germany, 1990. [Google Scholar]
- Stein, A.B.; Hayssen, V. Panthera pardus. Mamm. Spec. 2013, 45, 30–48. [Google Scholar] [CrossRef]
- Haas, S.K.; Hayssen, V.; Krausman, P.R. Panthera leo. Mamm. Spec. 2005, 762, 1–11. [Google Scholar] [CrossRef]
- Balme, G.A.; Pitman, R.T.; Robinson, H.S.; Miller, J.R.B.; Funston, P.J.; Hunter, L.T.B. Leopard distribution and abundance is unaffected by interference competition with lions. Behav. Ecol. 2017, 28, 1348–1358. [Google Scholar] [CrossRef]
- Vogel, J.T.; Somers, M.J.; Venter, J.A. Niche overlap and dietary resource partitioning in an African large carnivore guild. J. Zool. 2019, 309, 212–223. [Google Scholar] [CrossRef]
- Rafiq, K.; Hayward, M.W.; Wilson, A.M.; Meloro, C.; Jordan, N.R.; Wich, S.A.; McNutt, J.W.; Golabek, K.A. Spatial and temporal overlaps between leopards (Panthera pardus) and their competitors in the African large predator guild. J. Zool. 2020, 311. [Google Scholar] [CrossRef]
- Chaudhary, R.; Zehra, N.; Musavi, A.; Khan, J.A. Spatio-temporal partitioning and coexistence between leopard (Panthera pardus fusca) and Asiatic lion (Panthera leo persica) in Gir protected area, Gujarat, India. PLoS ONE 2020, 15, e0229045. [Google Scholar] [CrossRef]
- Yarnell, R.W.; Phipps, W.L.; Burgess, L.P.; Ellis, J.A.; Harrison, S.R.W.; Dell, S.; MacTavish, D.; MacTavish, L.M.; Scott, D.M. The influence of large predators on the feeding ecology of two African mesocarnivores: The black-backed jackal and the brown hyaena. Afr. J. Wildl. Res. 2013, 43, 155–166. [Google Scholar] [CrossRef]
- Mzezewa, J.; Misi, T.; van Rensburg, L. Characterisation of rainfall at a semi-arid ecotope in the Limpopo Province (South Africa) and its implications for sustainable crop production. Water SA 2010, 36, 19–26. [Google Scholar] [CrossRef]
- Low, A.B.; Rebelo, T.G. Vegetation of South Africa, Lesotho and Swaziland; Department of Environmental Affairs and Tourism: Pretoria, South Africa, 1996. [Google Scholar]
- Isaacs, L.; Somers, M.J.; Dalerum, F. Effects of prescribed burning and mechanical bush clearing on ungulate space use in an African savannah. Restor. Ecol. 2013, 21, 260–266. [Google Scholar] [CrossRef]
- Mucina, L.; Rutherford, M.C. The Vegetation of South Africa, Lesotho and Swaziland; Streliza 19; South African National Biodiversity Institute: Pretoria, South Africa, 2006. [Google Scholar]
- Dalerum, F.; Belton, L. African ungulates recognize a locally extinct native predator. Behav. Ecol. 2015, 26, 215–222. [Google Scholar] [CrossRef]
- Periquet, S.; Richardson, P.; Cameron, E.Z.; Ganswindt, A.; Belton, L.; Loubster, E.; Dalerum, F. Effects of lions on behaviour and endocrine stress in plains zebras. Ethology 2017, 123, 667–674. [Google Scholar] [CrossRef]
- Chizzola, M.; Belton, L.; Ganswindt, A.; Greco, I.; Hall, G.; Swanepoel, L.; Dalerum, F. Landscape level effects of lion presence (Panthera leo) on two contrasting prey species. Front. Ecol. Evol. 2018, 6, 191. [Google Scholar] [CrossRef]
- Ramnanan, R.; Swanepoel, L.; Somers, M. The diet and presence of African wild dogs (Lycaon pictus) on private land in the Waterberg region, South Africa. Afr. J. Wildl. Res. 2013, 43, 68–73. [Google Scholar] [CrossRef]
- Skead, C.J. Historical Incidence of the Larger Land Mammals in the Broader Western and Northern Cape, 2nd ed.; Nelson Mandela Metropolitan University: Port Elizabeth, South Africa, 2011. [Google Scholar]
- Swanepoel, L.; Somers, M.; Dalerum, F. Density of leopards Panthera pardus on protected and non-protected land in the Waterberg Biosphere, South Africa. Wildl. Biol. 2015, 21, 263–268. [Google Scholar] [CrossRef]
- Karanth, K.U.; Chundawat, R.S.; Nichols, J.D.; Kumar, N.S. Estimation of tiger densities in the tropical dry forests of Panna, central India, using photographic capture–recapture sampling. Anim. Conserv. 2004, 7, 285–290. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Edwards, D. A broad-scale structural classification of vegetation for practical purposes. Bothalia 1983, 14, 705–712. [Google Scholar] [CrossRef]
- Gerber, B.; Karpanty, S.; Kelly, M.J. Evaluating the potential biases in carnivore capture–recapture studies associated with the use of lure and varying density estimation techniques using photographic-sampling data of the Malagasy civet. Popul. Ecol. 2012, 54, 43–54. [Google Scholar] [CrossRef]
- Caughley, G. Bias in aerial survey. J. Wildl. Manag. 1974, 38, 921–933. [Google Scholar] [CrossRef]
- Jachmann, H. Comparisons of aerial counts with ground counts for large African herbivores. J. Appl. Ecol. 2002, 39, 841–852. [Google Scholar] [CrossRef]
- Foguekem, D.; Tchamba, M.N.; Omondi, P. Aerial survey of elephants (Loxodonta africana africana), other large mammals and human activities in Waza National Park, Cameroon. Afr. J. Environ. Sci. Technol. 2010, 4, 401–411. [Google Scholar]
- Craig, G.C. Aerial Survey Standards for the MIKE Programme, version 2.0. CITES MIKE Programme, Nairobi, Kenya. 2012. Available online: https://cites.org/sites/default/files/common/prog/mike/survey/MIKE_aerial_standards.v2.pdf (accessed on 25 August 2020).
- Hulsman, A.; Dalerum, F.; Swanepoel, L.; Ganswindt, A.; Sutherland, C.; Paris, M. Patterns of scat deposition by brown hyaenas (Hyaena brunnea) in a mountain savanna region of South Africa. Wildl. Biol. 2010, 16, 445–451. [Google Scholar] [CrossRef]
- Orr, A.J.; Laake, J.L.; Dhruv, M.I.; Banks, A.S.; de Long, R.L.; Huber, H.R. Comparison of processing pinniped scat samples using a washing machine and nested sieves. Wildl. Soc. Bull. 2003, 30, 253–257. [Google Scholar]
- Perrin, M.R.; Campbell, B.S. Key to the mammals of the Andries Vosloo Kudu Reserve (Eastern Cape), based on their hair morphology, for use in predator scat analysis. Afr. J. Wildl. Res. 1979, 10, 1–14. [Google Scholar]
- Keogh, H.J. A photographic reference system of the microstructure of the hair of southern African bovids. Afr. J. Wildl. Res. 1983, 13, 89–132. [Google Scholar]
- Buys, D.; Keogh, H.J. Notes on the microstructure of hair of the Orycteropodidae, Elephantidae, Equidae, Suidae and Giraffidae. Afr. J. Wildl. Res. 1984, 14, 111–119. [Google Scholar]
- Douglas, R.M. A new method of cross-sectioning hair of larger mammals. Afr. J. Wildl. Res. 1989, 19, 73–76. [Google Scholar]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modelling: Inferring Patterns and Dynamics of Species Occurrence; Academic Press: San Diego, CA, USA, 2006. [Google Scholar]
- MacKenzie, D.I.; Nichols, J.D. Occupancy as a surrogate for abundance estimation. Anim. Biodivers. Conserv. 2004, 27, 461–467. [Google Scholar]
- Neyman, J.; Pearson, E.S. On the problem of the most efficient tests of statistical hypotheses. Philosophical Transactions of the Royal Society of London Series A 1933, 231, 289–337. [Google Scholar]
- Akaike, H. A New Look at the Statistical Model Definition; IEEE: Piscataway, NJ, USA, 1974; Volume 19, pp. 716–723. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Jacobs, J. Quantitative measurement of food selection: A modification of the forage ratio and Ivlev’s Electivity Index. Oecologia 1974, 14, 413–417. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, 2020. Available online: https://www.r-project.org (accessed on 26 August 2020).
- Fiske, I.; Chandler, R. Unmarked: An R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef]
- Mugerwa, B.; du Preez, B.; Tallents, L.A.; Loveridge, A.J.; Macdonald, D.W. Increased foraging success or competitor avoidance? Diel activity of sympatric large carnivores. J. Mammal. 2017, 98, 1443–1452. [Google Scholar] [CrossRef]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Timothy, S.E.; Holt, R.D.; Jackson, J.B.C.; et al. Trophic downgrading of planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef]
- Kohl, M.; Stahler, D.R.; Metz, M.C.; Forester, J.D.; Kauffman, M.J.; Varley, N.; White, P.J.; Smith, D.W.; MacNulty, D.R. Diel predator activity drives a dynamic landscape of fear. Ecol. Monogr. 2018, 88, 638–652. [Google Scholar] [CrossRef]
- Hargreaves, A.; Germain, R.M.; Bontrager, M.; Persi, J.; Angert, A.L. Local adaptation to biotic interactions: A meta-analysis across latitudes. Am. Nat. 2020, 195, 395–411. [Google Scholar] [CrossRef]
- Middleton, A.D.; Kauffman, M.J.; McWhirther, E.J.; Jimenez, M.J.; Cook, R.C.; Cook, J.G.; Albeke, S.E.; Sawyer, H.; White, P.J. Linking anti-predator behavior to prey demography reveals limited risk effects of an actively hunting large carnivore. Ecol. Lett. 2013, 16, 1023–1030. [Google Scholar] [CrossRef]
- Greco, I.; Chizzola, M.; Meloro, C.; Swanepoel, L.; Tamagnini, D.; Dalerum, F. Similarities in size, morphology and diel activity between lions and sympatric carnivores. J. Zool. under review.
- Haswell, P.M.; Kusak, J.; Hayward, M.W. Large carnivore impacts are context-dependent. Food Webs 2017, 12, 3–13. [Google Scholar] [CrossRef]
- Miller, J.R.B.; Pitman, R.T.; Mann, G.K.H.; Fuller, A.K.; Balme, G.A. Lions and leopards coexist without spatial, temporal or demographic effects of interspecific competition. J. Anim. Ecol. 2018, 87, 1709–1726. [Google Scholar] [CrossRef]
- Hunter, M.D.; Price, P.W. Playing chutes and ladders: Heterogeneity and the relative roles of bottom-up and top-down forces in natural communities. Ecology 1992, 73, 724–732. [Google Scholar]
- Sivy, K.; Pozzanghera, C.B.; Colson, K.E.; Mumma, M.A.; Prugh, L.R. Apex predators and the facilitation of resource partitioning among mesopredators. Oikos 2018, 127, 607–621. [Google Scholar] [CrossRef]
- Prugh, L.R.; Sivy, K. Enemies with benefits: Integrating positive and negative interactions among terrestrial carnivores. Ecol. Lett. 2020, 23, 902–918. [Google Scholar] [CrossRef] [PubMed]
- van der Merwe, I.; Tambling, C.J.; Thorn, M.; Scott, D.M.; Yarnell, R.W.; Green, M.; Cameron, E.Z.; Bateman, P.W. An assessment of diet overlap of two mesocarnivores in the North West Province, South Africa. Afr. J. Zool. 2009, 44, 288–291. [Google Scholar] [CrossRef]
- Hurlbert, S.H. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef]
- Ríos-Saldaña, C.; Delibes-Mateos, M.; Ferrerira, C.C. Are field studies being relegated to second place in conservation science? Glob. Ecol. Cons. 2018, 14, e00389. [Google Scholar]
- Marucco, F.; Pletcher, D.H.; Boitani, L. Accuracy of scat sampling for carnivore diet analysis: Wolves in the Alps as a case study. J. Mammal. 2008, 89, 665–673. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bashant, J.; Somers, M.; Swanepoel, L.; Dalerum, F. Facilitation or Competition? Effects of Lions on Brown Hyaenas and Leopards. Diversity 2020, 12, 325. https://doi.org/10.3390/d12090325
Bashant J, Somers M, Swanepoel L, Dalerum F. Facilitation or Competition? Effects of Lions on Brown Hyaenas and Leopards. Diversity. 2020; 12(9):325. https://doi.org/10.3390/d12090325
Chicago/Turabian StyleBashant, Janelle, Michael Somers, Lourens Swanepoel, and Fredrik Dalerum. 2020. "Facilitation or Competition? Effects of Lions on Brown Hyaenas and Leopards" Diversity 12, no. 9: 325. https://doi.org/10.3390/d12090325
APA StyleBashant, J., Somers, M., Swanepoel, L., & Dalerum, F. (2020). Facilitation or Competition? Effects of Lions on Brown Hyaenas and Leopards. Diversity, 12(9), 325. https://doi.org/10.3390/d12090325