Out of Bound: A New Threatened Harlequin Toad (Bufonidae, Atelopus) from the Outer Borders of the Guiana Shield in Central Amazonia Described through Integrative Taxonomy


Abstract
1. Introduction
2. Materials and Methods
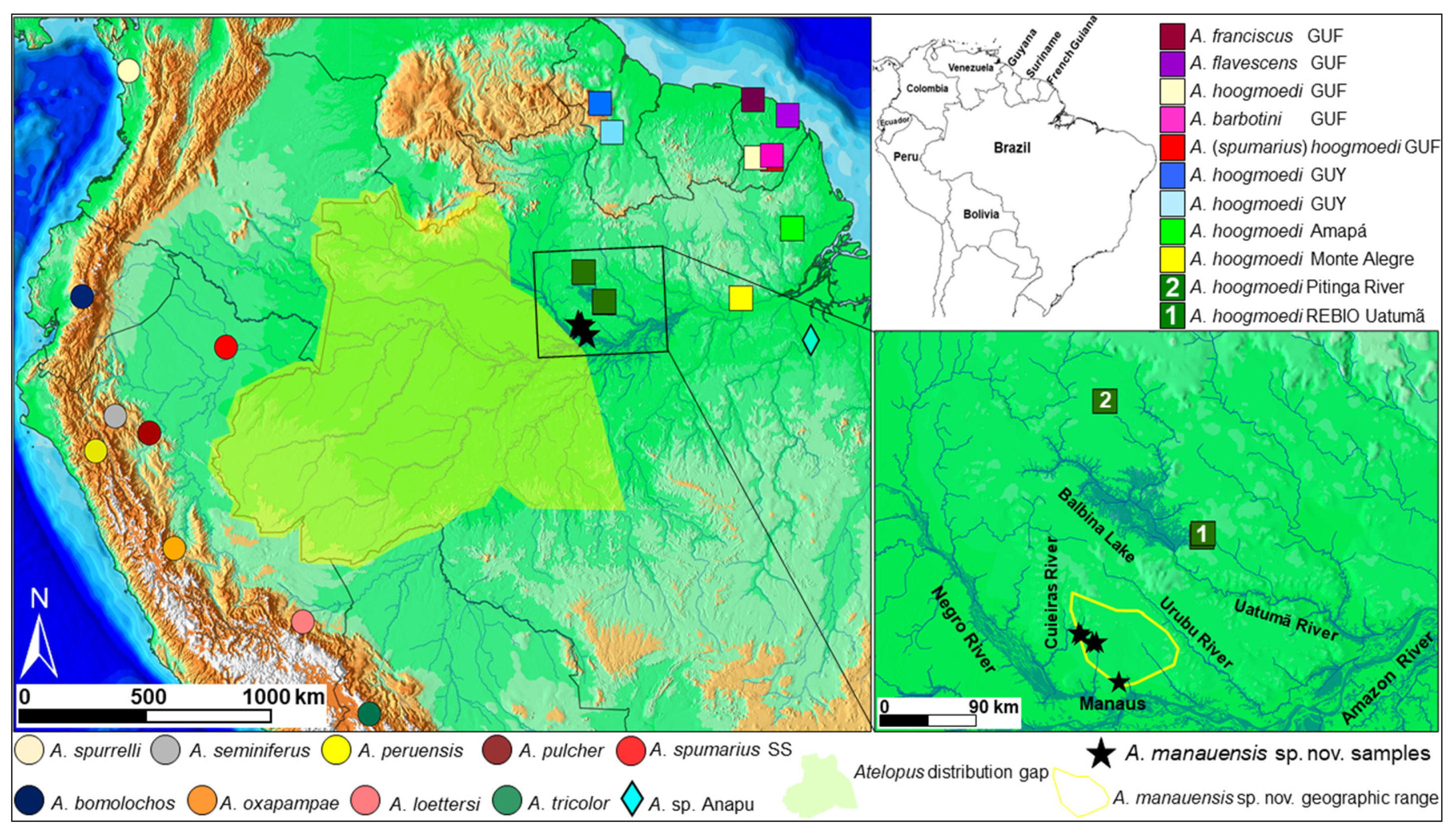
2.1. Sampling
2.2. Phylogenetic Analyses
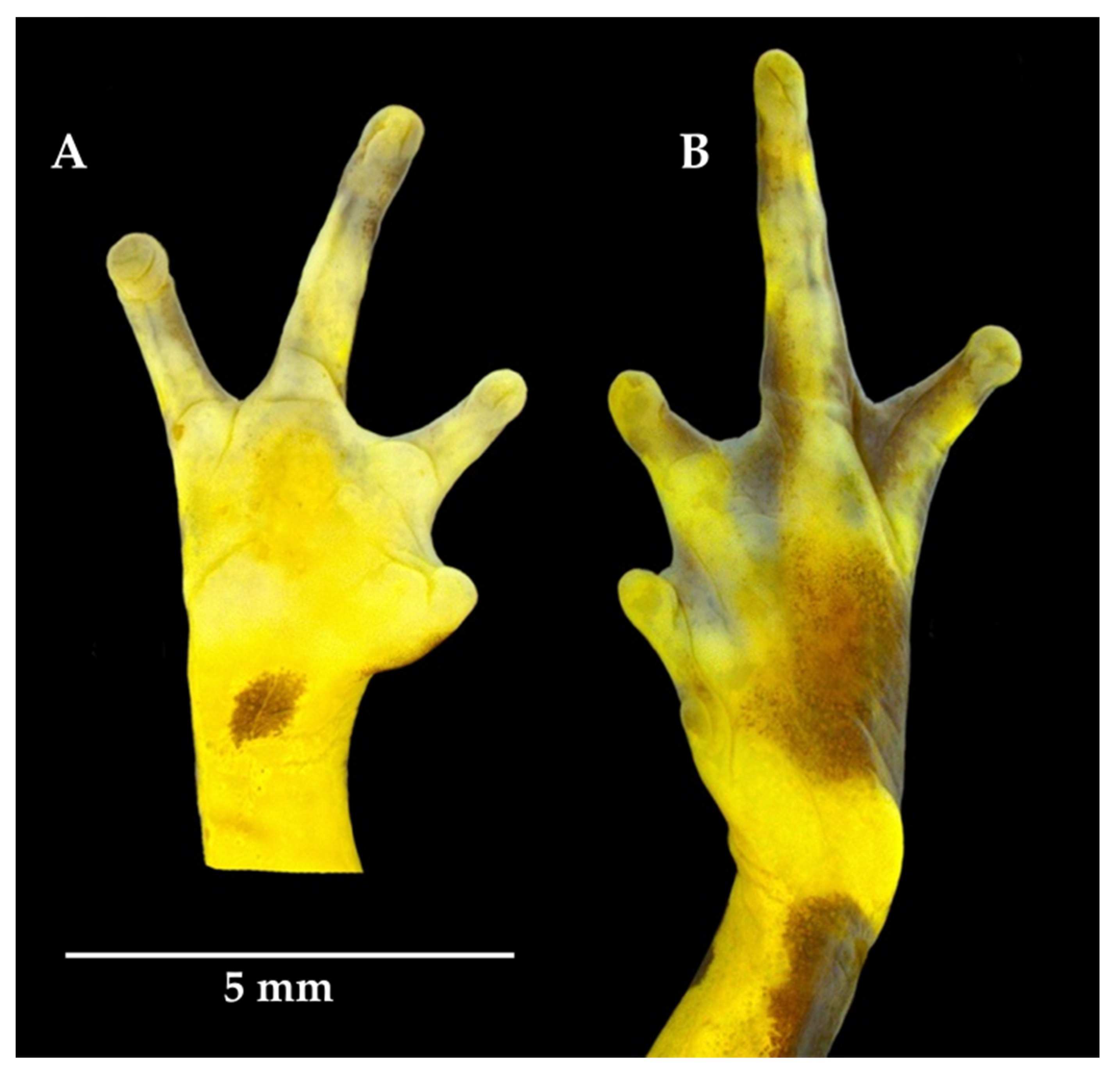
2.3. Morphological Analyses
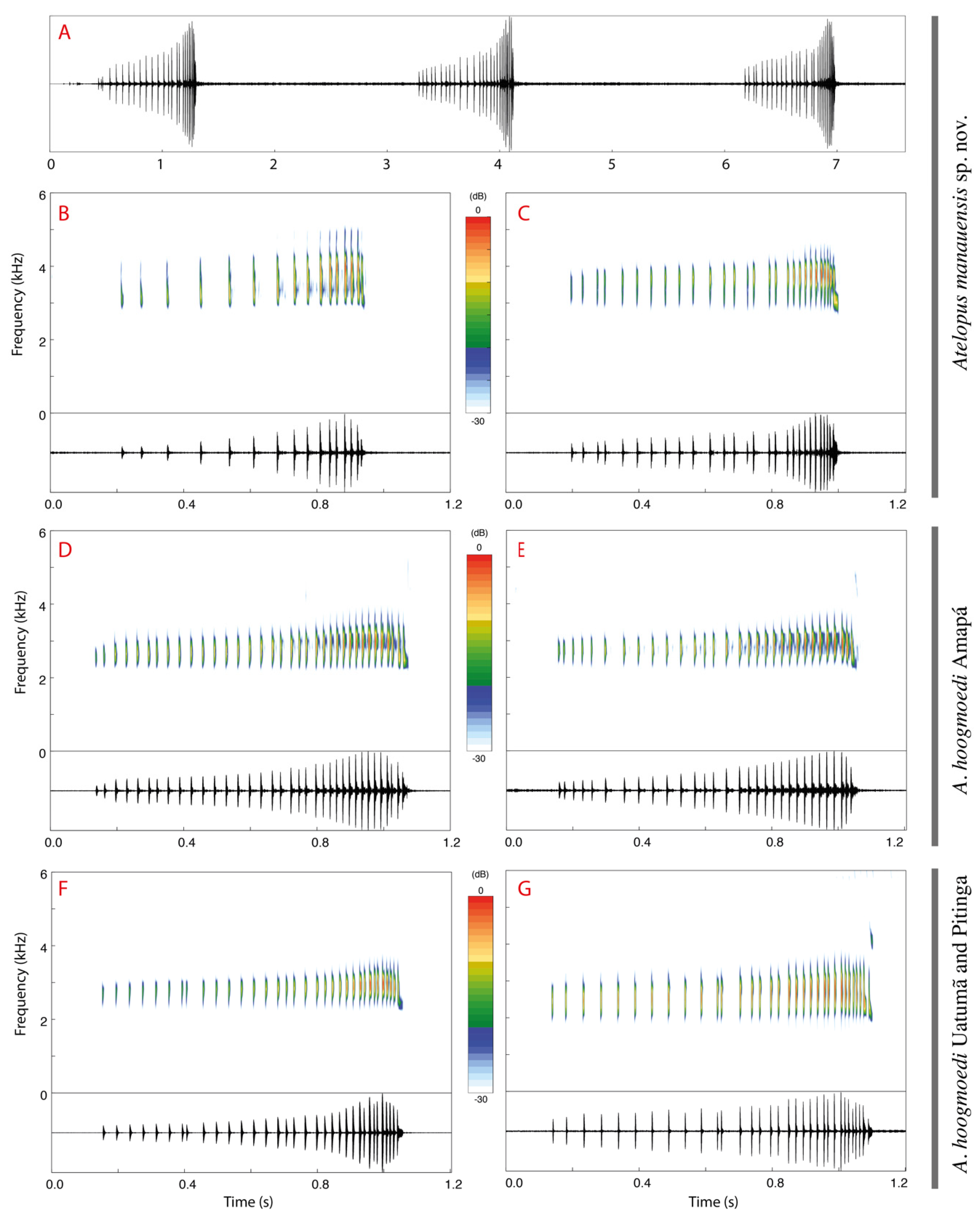
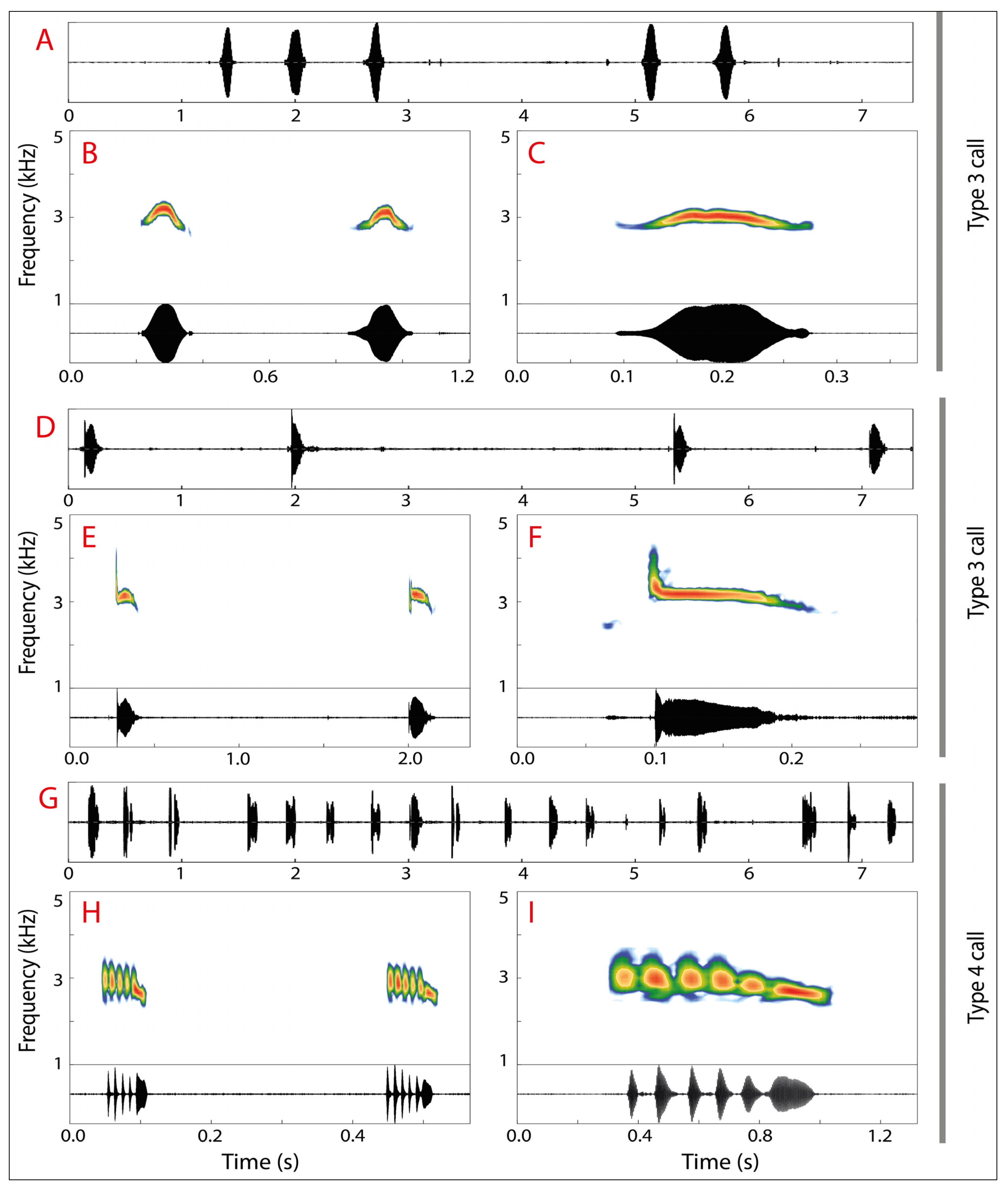
2.4. Bioacoustics
2.5. Natural History
3. Results
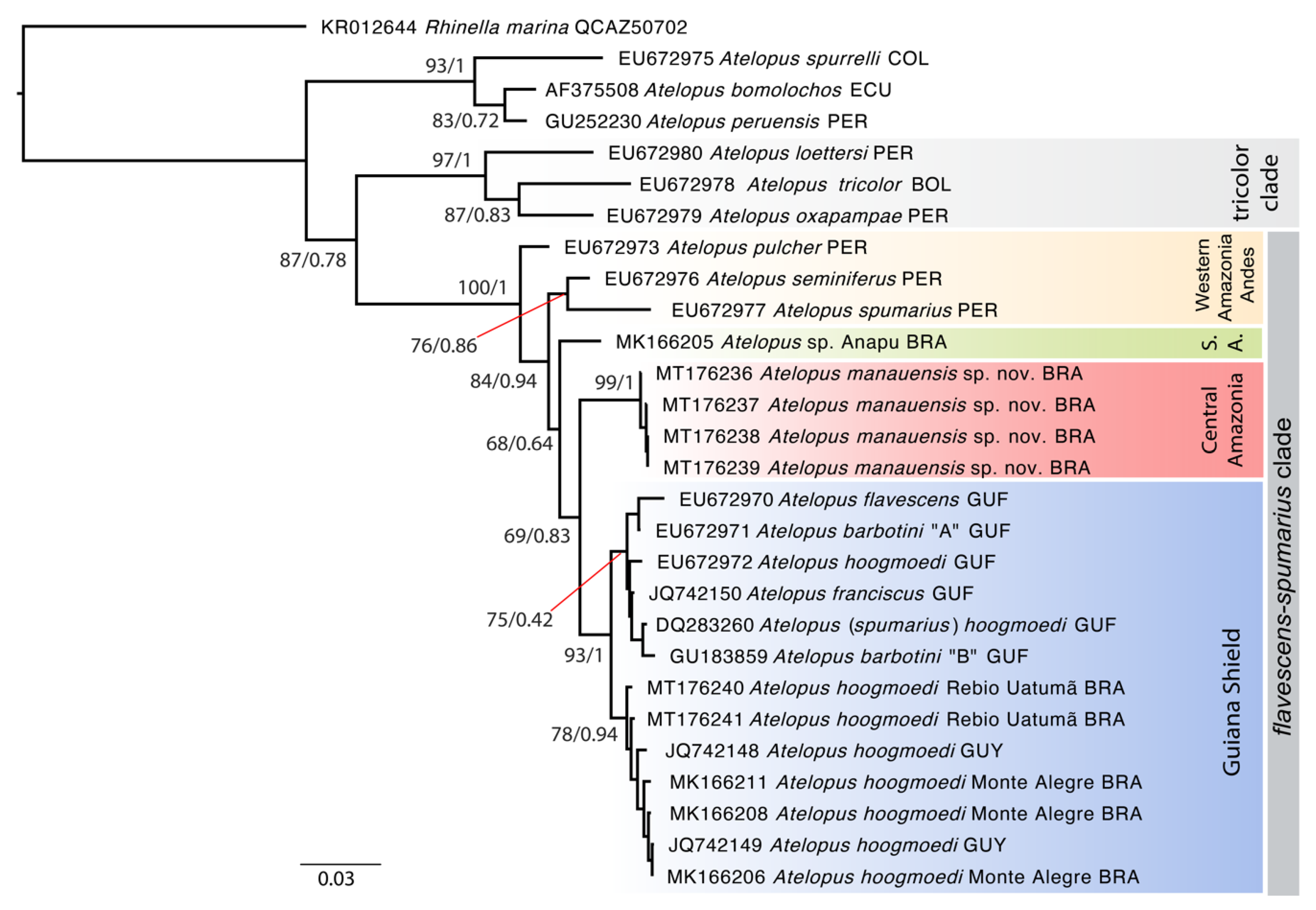
3.1. Phylogenetic Analyses
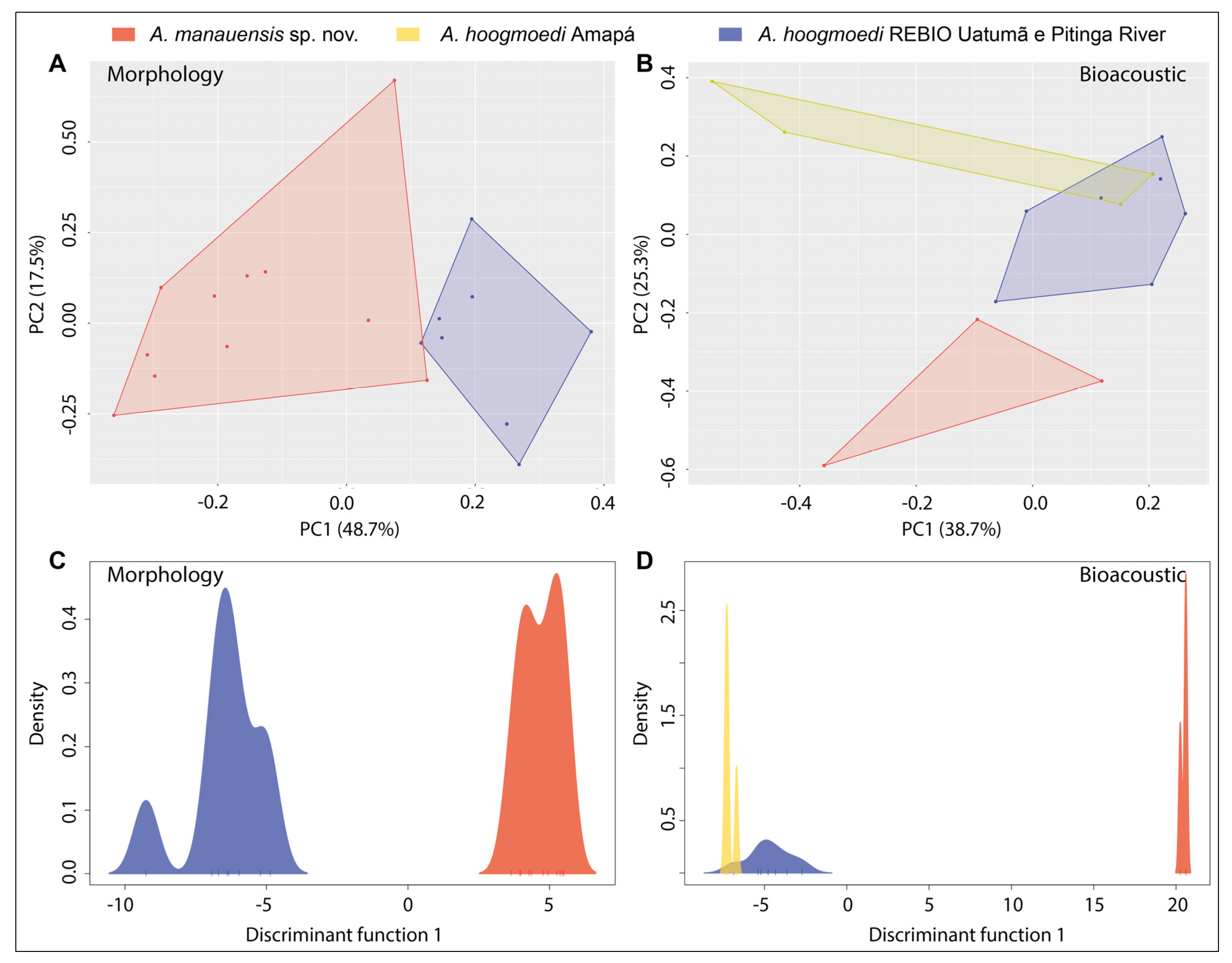
3.2. Morphological and Bioacoustic Analyses
3.3. Taxonomic Account
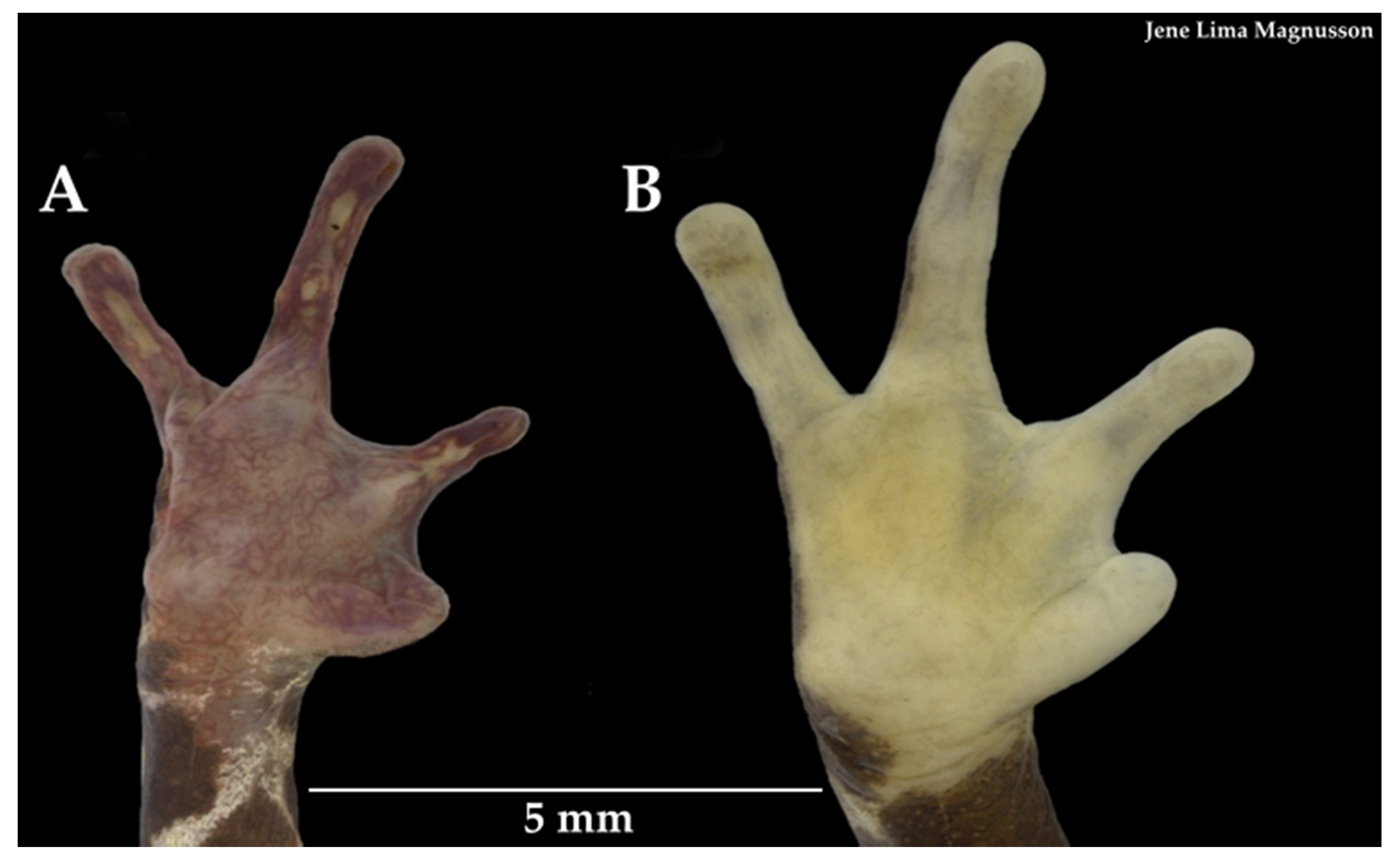
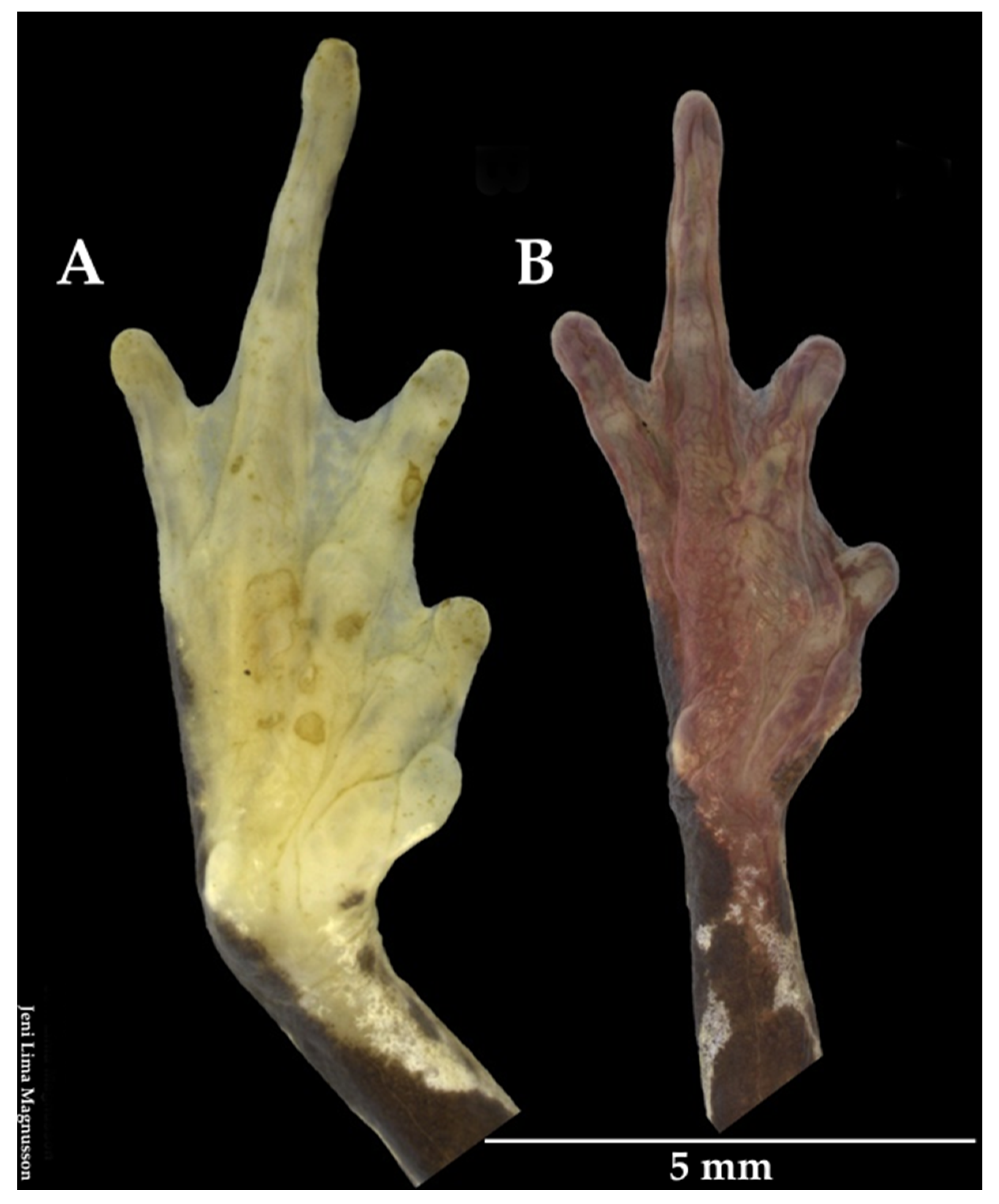
3.3.1. Taxonomic Comparisons
3.3.2. Variability
3.4. Distribution and Natural History
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Frost, D.R. Amphibian Species of the World: An Online Reference. Version 6.1. Available online: https://amphibiansoftheworld.amnh.org/index.php (accessed on 20 November 2019).
- Lötters, S. The neotropical toad genus Atelopus. Checklist. In Biology and Distribution; M. Vences, F. Glaw Verlags GbR: Köln, Germany, 1996; p. 143. [Google Scholar]
- Lötters, S.; Van Der Meijden, A.; Coloma, L.A.; Boistel, R.; Cloetens, P.; Ernst, R.; Lehr, E.; Veith, M. Assessing the molecular phylogeny of a near extinct group of vertebrates: The Neotropical harlequin frogs (Bufonidae: Atelopus). Syst. Biodivers. 2011, 9, 45–57. [Google Scholar] [CrossRef]
- Lötters, S.; Haas, W.; Schick, S.; Böhme, W. On the systematics of the harlequin frogs (Amphibia: Bufonidae: Atelopus) from Amazonia. II: Redescription of Atelopus pulcher (Boulenger, 1882) from the eastern Andean versant in Peru. Salamandra 2002, 38, 165–184. [Google Scholar]
- Kok, P.J.R. A survey of the anuran fauna of Montagne Belvédère, county of Saül, French Guiana: Field list with comments on taxonomy and ecology. Br. Herpetol. Soc. Bull. 2000, 71, 6–26. [Google Scholar]
- Cope, E.D. Ninth contribution to the herpetology of Tropical America. Proc. Acad. Nat. Sci. Phila. 1871, 23, 200–222. [Google Scholar]
- Lescure, J. Contribution a l’etude des Amphibiens de Guyane francaise. VIII. Validation d’Atelopus spumarius Cope, 1871, et designation d’un neotype. Description d’Atelopus spumarius barbotini nov. ssp. Donnees etho-ecologiques et biogeographiques sur les d’Atelopus du groupe flavescens (Anoures, Bufonides). Bull. Mus. Natl. Hist. Nat. B 1981, 4, 893–910. [Google Scholar]
- Rivero, J.A. More on the Atelopus (Amphibia, Salientia) from western South America. Caribb. J. Sci. 1968, 8, 19–29. [Google Scholar]
- Lescure, J. Contribution a l’étude des amphibiens de Guyane Française i. notes sur Atelopus flavescens Duméril et Bibron et description d’une nouvelle espèce. Vie Milieu. 1972–1973, 23, 125–141. [Google Scholar]
- Lötters, S.; de La Riva, I. Redescription of Atelopus tricolor Boulenger from Southeastern Peru and adjacent Bolivia, with comments on related forms. J. Herpetol. 1998, 32, 481–488. [Google Scholar] [CrossRef]
- Noonan, B.P.; Gaucher, P. Phylogeography and demography of Guianan harlequin toads (Atelopus): Diversification within a refuge. Mol. Ecol. 2005, 14, 3017–3031. [Google Scholar] [CrossRef]
- Zimmerman, B.L. A comparison of strucutural features of calls of open and forest habitat frog species in the Central Amazon. Herpetologica 1983, 39, 235–246. [Google Scholar]
- Zimmerman, B.L.; Bierregaard, R.O. Relevance of the equilibrium theory of island biogeography and species-area relations to conservation with a case from Amazonia. J. Biogeogr. 1986, 13, 133–143. [Google Scholar] [CrossRef]
- Zimmerman, B.L.; Simberloff, D. An historical interpretation of habitat used by frogs in a Central Amazonian Forest. J. Biogeogr. 1996, 23, 27–43. [Google Scholar] [CrossRef]
- Gascon, C. The tadpole of Atelopus pulcher Boulenger (Annura: Bufonidae) from Manaus, Amazonas. Zoologia 1989, 6, 235–239. [Google Scholar] [CrossRef]
- Boulenger, G.A. Catalogue of the Batrachia Salientia s. Ecaudata in the Collection of the British Museum, 2nd ed.; London Printed by order of the Trustees: London, UK, 1882; p. 528. [Google Scholar]
- Peters, J.A. The Frog Genus Atelopus in Ecuador (Anura: Bufonidae). Smithson. Contrib. Zool. 1973, 145, 1–49. [Google Scholar] [CrossRef]
- Lima, A.P.; Magnusson, W.E.; Menin, M.; Erdtmann, L.K.; Rodrigues, D.J.; Keller, C.; Hödl, W. Guide to the Frogs of Reserva Ducke–Central Amazonia, 2nd ed.; Áttema Design Editorial: Manaus, Brazil, 2006; p. 188. [Google Scholar]
- Rojas-Ahumada, D.P.; Menin, M. Composition and abundance of anurans in riparian and non-riparian areas in a forest in central Amazonia, Brazil. S. Am. J. Herpetol. 2010, 5, 157–167. [Google Scholar] [CrossRef]
- Menin, M.; Waldez, F.; Lima, A.P. Effects of environmental and spatial factors on the distribution of anuran species with aquatic reproduction in central Amazonia. Herpetol. J. 2011, 21, 255–261. [Google Scholar]
- Siqueira, S.; Aguiar Jr, O.; Lima, A.P.; Recco-Pimentel, S.M. Cytogenetics and sperm ultrastructure of Atelopus spumarius (Anura: Bufonidae) from the Brazilian Amazon. Genet. Mol. Biol. (Impresso) 2013, 36, 528–532. [Google Scholar] [CrossRef]
- Jorge, R.F.; Simões, P.I.; Magnusson, W.E.; Lima, A.P. Fine-scale habitat heterogeneity explains the local distribution of two Amazonian frog species of concern for conservation. Biotropica 2016, 48, 694–703. [Google Scholar] [CrossRef]
- Rößler, D.C.; Lötters, S.; Mappes, J.; Valkonen, J.K.; Menin, M.; Lima, A.P.; Pröhl, H. Sole coloration as an unusual aposematic signal in a Neotropical toad. Sci. Rep. 2018, 9, 1128. [Google Scholar] [CrossRef]
- Duellman, W.E.; Lynch, J.D. Descriptions of Atelopus tadpoles and their relevance to Atelopodid classification. Herpetology 1969, 25, 231–240. [Google Scholar]
- Lötters, S.; Mebs, D.; Köhler, G.; Vargas, J.; La Marca, E. The voice from the hereafter: Vocalizations in three species of Atelopus from the Venezuelan Andes, likely to be extinct. Herpetozoa 2019, 32, 267–275. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2 new methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Jorge, R.F.; Magnusson, W.E.; Silva, D.A.; Polo, E.M.; Lima, A.P. Urban growth threatens the lowland Amazonian Manaus harlequin frog which represents an evolutionarily significant unit within the genus Atelopus (Amphibia: Anura: Bufonidae). J. Zool. Syst. Evol. Res. 2020, 1–11. [Google Scholar] [CrossRef]
- da Silva, G.W.B.; Cornélio, G.S.; de Oliveira, E.A.; Trindade, N.G.P.; França, I.; Hernández Ruz, E.J. A candidate species currently classified as Atelopus hoogmoedi (Anura: Bufonidae) in the eastern Amazon, Pará, Brazil. Genet. Mol. Res. 2020, 19, gmr18392. [Google Scholar] [CrossRef]
- Kok, P.J.; MacCulloch, R.D.; Means, D.B.; Roelants, K.; Van Bocxlaer, I.; Bossuyt, F. Low genetic diversity in tepui summit vertebrate. Curr. Biol. 2012, 22, 589–590. [Google Scholar] [CrossRef]
- Lötters, S.; van der Meijden, A.; Rödder, D.; Köster, T.E.; Kraus, T.; La Marca, E.; Haddad, C.F.B.; Veith, M. Reinforcing and expanding the predictions of the disturbance vicariance hypothesis in Amazonian harlequin frogs: A molecular phylogenetic and climate envelope modelling approach. Biodivers. Conserv. 2010, 19, 2125–2146. [Google Scholar] [CrossRef]
- Van Bocxlaer, I.; Loader, S.P.; Roelants, K.; Biju, S.D.; Menegon, M.; Bossuyt, F. Gradual adaptation toward a range-expansion phenotype initiated the global radiation of toads. Science 2010, 327, 679–682. [Google Scholar] [CrossRef]
- Frost, D.R.; Grant, T.; Faivovich, J.; Bain, R.; Haas, A.; Haddad, C.F.B.; de Sa, R.O.; Channing, A.; Wilkinson, M.; Donnellan, S.C.; et al. The Amphibian tree of life. Bull. Am. Mus. Nat. Hist. 2006, 297, 1–291. [Google Scholar] [CrossRef]
- dos Santos, S.P.; Ibanez, R.; Ron, S.R. Systematics of the Rhinella margaritifera complex (Anura, Bufonidae) from western Ecuador and Panama with insights in the biogeography of Rhinella alata. Zookeys 2015, 501, 109–145. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2011, 61, 539–542. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA 6: Molecular evolutionary genetics analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Gray, P.; Cannatella, D.C. A new species of Atelopus (Anura: Bufonidae) from the Andes of Northern Peru. Copeia 1985, 4, 910–917. [Google Scholar] [CrossRef]
- Coloma, L.A.; Lötters, S.; Salas, A.W. Taxonomy of the Atelopus ignescens complex (Anura:Bufonidae): Designation of a Neotype of Atelopus ignescens and recognition of Atelopus exiguus. Herpetologica 2000, 56, 303–324. [Google Scholar]
- Savage, J.M.; Heyer, W.R. Variation and distribution in the tree-frog genus Phyllomedusa in Costa Rica, Central America. Beiträge zur Neotropischen 1967, 5, 111–131. [Google Scholar] [CrossRef]
- Savage, J.M.; Heyer, W.R. Digital webbing formulae for anurans: A refinement. Herpetol. Rev. 1997, 28, 131. [Google Scholar]
- Myers, C.W.; Duellman, W.E. A new species of Hyla from Cerro Colorado, and other tree frog records and geographical notes from Western Panama. Am. Mus. Novit. 1982, 2752, 1–32. [Google Scholar]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computin; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Liaw, A.; Wiener, M. Classification and Regression by RandomForest. R News 2002, 2, 18–22. [Google Scholar]
- Center for Conservation Bioacoustics. Raven Pro: Interactive Sound Analysis Software Version 1.6.1. Computer Software; The Cornell Lab of Ornithology: Ithaca, NY, USA, 2019. [Google Scholar]
- Costa-Campos, C.E.; Carvalho, T.R. The advertisement call of the Hoogmoed’s harlequin toad Atelopus hoogmoedi Lescure, 1974 from northern Brazil (Anura: Bufonidae). Zootaxa 2018, 4521, 141–144. [Google Scholar] [CrossRef]
- Köhler, J.; Jansen, M.; Rodríguez, A.; Kok, F.J.R.; Toledo, L.F.; Emmrich, M.; Glaw, F.; Haddad, C.F.B.; Rӧdel, M.-O.; Vences, M. The use of bioacoustics in anuran taxonomy: Theory, terminology, methods and recommendations for best practice. Zootaxa 2017, 4251, 1–124. [Google Scholar] [CrossRef]
- Lescure, J. Présence d’une sous-espèce Atelopus pulcher (Amphibien, Anoure) dans les Guyanes: Atelopus pulcher hoogmoedi. Bull. Mus. Natl. Hist. Nat. 1973, 108, 997–1005. [Google Scholar]
- Lötters, S.; Boistel, R.; Blanc, M.; Haddad, C.F.B.; van der Meijden, A. Atelopus hoogmoedi Lescure, 1974. In Ranas Arlequines; Almonacid, J.V.R., Mahecha, J.V.R., la Marca, E., Lötters, S., Kahn, T., Angulo, A., Eds.; Conservácion International: Bogotá, Colombia, 2005; p. 132. [Google Scholar]
- Kok, P.J.R.; Kalamandeen, M. Introduction of the taxonomy of the amphibians of Kaieteur National Park, Guyana. ABC Taxa 2008, 5, 1–278. [Google Scholar]
- de La Riva, I.; Castroviejo-Fisher, S.; Chaparro, J.C.; Boistel, R.; Padial, J.M. A new species of Atelopus (Anura: Bufonidae) from the Amazonian slopes of the Andes in south-eastern Peru. Salamandra 2011, 47, 161–168. [Google Scholar]
- Lötters, S.; Shulte, R. Atelopus seminiferus Cope, 1874. In Ranas Arlequinas; Almonacid, J.V.R., Mahecha, J.V.R., la Marca, E., Lötters, S., Kahn, T., Angulo, A., Eds.; Conservácion International: Bogotá, Colombia, 2005; p. 132. [Google Scholar]
- Cope, E.D. On some Batrachia and Nematognathi brought from the Upper Amazon by Prof. Orton. Proc. Acad. Nat. Sci. Phila. 1874, 26, 120–137. [Google Scholar]
- La Marca, E.; Lips, K.L.; Lötters, S.; Puschendorf, R.; Ibáñez, R.; Rueda-Almonacid, J.V.; Schulte, R.; Marty, C.; Castro, F.; Manzanilla-Puppo, J.; et al. Catastrophic population declines and extinctions in Neotropical harlequin frogs (Bufonidae: Atelopus). Biotropica 2005, 37, 190–201. [Google Scholar] [CrossRef]
- Lambertini, C.; Missassi, A.F.R.; Jorge, R.F.; Leite, D.A.; Lima, A.P.; Toledo, L.F. Climatic relationships between the killing-chytrid fungus and wild amphibians in the Brazilian Amazonia. Dis. Aquat. Org. Submitted manuscript.
- Vences, M.; Thomas, M.; van Der Meijden, A.; Chiari, Y.; Vieites, D.R. Comparative performance of the 16S rRNA gene in DNA barcoding of amphibians. Front. Zool. 2005, 2, 1–12. [Google Scholar] [CrossRef]
- Fouquet, A.; Gilles, A.; Vences, M.; Marty, C.; Blanc, M.; Gemmell, N.J. Underestimation of species richness in Neotropical frogs revealed by mtDNA analyses. PLoS ONE 2007, 2, e1109. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Species | GenBank 16S/COI | Voucher | Country | Source |
|---|---|---|---|---|---|
| 1 | Atelopus manauensis sp. nov. | MT176236/MT184269 | INPA-H 041289 | Brazil | [29] |
| 2 | Atelopus manauensis sp. nov. | MT176237/MT184270 | INPA-H 041290 | Brazil | [29] |
| 3 | Atelopus manauensis sp. nov. | MT176238/MT184271 | INPA-H 041291 | Brazil | [29] |
| 4 | Atelopus manauensis sp. nov. | MT176239/MT184272 | INPA-H 041292 | Brazil | [29] |
| 5 | Atelopus hoogmoedi REBIO Uatumã | MT176240/MT184273 | INPA-H 041293 | Brazil | [29] |
| 6 | Atelopus hoogmoedi REBIO Uatumã | MT176241/MT184274 | INPA-H 041294 | Brazil | [29] |
| 7 | Atelopus sp. Anapu | MK166205 | KA14 | Brazil | [30] |
| 8 | Atelopus hoogmoedi Monte Alegre | MK166206 | LZA971 | Brazil | [30] |
| 9 | Atelopus hoogmoedi Monte Alegre | MK166208 | LZA990 | Brazil | [30] |
| 10 | Atelopus hoogmoedi Monte Alegre | MK166211 | LZA1046 | Brazil | [30] |
| 11 | Atelopus hoogmoedi | JQ742148 | IRSNB15781 | Guyana | [31] |
| 12 | Atelopus hoogmoedi | JQ742149 | IRSNB14477 | Guyana | [31] |
| 13 | Atelopus barbotini “A” | EU672971 | - | French Guiana | [32] |
| 14 | Atelopus barbotini “B” | GU183859 | BPN1697 | French Guiana | [33] |
| 15 | Atelopus franciscus | JQ742150 | PK3306 | French Guiana | [31] |
| 16 | Atelopus (spumarius) hoogmoedi | DQ283260 | BPN754UTA | French Guiana | [34] |
| 17 | Atelopus hoogmoedi | EU672972 | - | French Guiana | [32] |
| 18 | Atelopus flavescens | EU672970 | - | French Guiana | [32] |
| 19 | Atelopus seminiferus | EU672976 | - | Peru | [32] |
| 20 | Atelopus spumarius | EU672977 | - | Peru | [32] |
| 21 | Atelopus pulcher | EU672973 | KU211678 | Peru | [32] |
| 22 | Atelopus bomolochos | GU252227 | KU217468 | Ecuador | [33] |
| 23 | Atelopus peruensis | GU252229 | KU211631 | Peru | [33] |
| 24 | Atelopus spurrelli | EU672975/DQ502895 | MHNUC273 | Colombia | [32,34] |
| 25 | Atelopus loettersi | EU672980 | - | Peru | [32] |
| 26 | Atelopus oxapampae | EU672979 | MTD1276 | Peru | [32] |
| 27 | Atelopus tricolor | EU672978 | MNCN5885 | Bolivia | [32] |
| 28 | Rhinella marina | KR012644/ KR012546 | QCAZ50702 | Ecuador | [35] |
| ID | Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | A. spurrelli COL | 13.4 | 10.7 | 5.0 | 11.7 | 4.8 | 12.4 | 11.9 | 13.6 | 12.2 | 13.1 | 12.4 | 12.0 | 13.2 | 12.6 | 14.2 | 12.4 | 14.1 | 12.7 | |
| 2 | A. tricolor BOL | 12.1 | 6.7 | 11.8 | 5.6 | 12.2 | 12.0 | 11.8 | 12.3 | 12.4 | 12.8 | 12.6 | 12.7 | 12.5 | 13.1 | 13.2 | 12.2 | 13.3 | 12.5 | |
| 3 | A. loettersi PER | 9.9 | 6.3 | 10.7 | 6.1 | 11.1 | 13.7 | 10.7 | 12.3 | 11.6 | 13.1 | 12.0 | 11.2 | 12.0 | 12.3 | 12.7 | 11.7 | 13.1 | 11.9 | |
| 4 | A. bomolochos ECU | 4.8 | 10.8 | 10.0 | 10.7 | 2.2 | 11.3 | 10.4 | 12.1 | 10.7 | 10.9 | 11.4 | 11.2 | 11.6 | 11.6 | 12.6 | 11.2 | 12.4 | 10.9 | |
| 5 | A. oxapampae PER | 10.8 | 5.3 | 5.8 | 9.9 | 10.7 | 12.2 | 10.2 | 11.8 | 11.1 | 13.1 | 11.5 | 11.2 | 11.3 | 11.4 | 12.0 | 11.2 | 12.1 | 10.8 | |
| 6 | A. peruensis PER | 4.6 | 11.1 | 10.3 | 2.2 | 9.9 | 11.1 | 10.3 | 11.6 | 10.4 | 10.8 | 10.9 | 10.7 | 11.3 | 11.1 | 12.1 | 10.7 | 12.3 | 10.5 | |
| 7 | A. spumarius SS PER | 11.4 | 11.1 | 12.5 | 10.5 | 11.3 | 10.3 | 4.5 | 4.7 | 2.8 | 3.4 | 3.7 | 4.3 | 2.8 | 4.1 | 3.8 | 3.9 | 3.2 | 4.9 | |
| 8 | A. pulcher PER | 11.0 | 10.9 | 9.9 | 9.6 | 9.5 | 9.6 | 4.3 | 3.4 | 2.6 | 3.3 | 2.7 | 3.4 | 2.7 | 3.1 | 2.8 | 2.9 | 3.1 | 4.0 | |
| 9 | A. flavescens GUF | 12.4 | 11.3 | 11.3 | 11.2 | 10.9 | 10.7 | 4.6 | 3.3 | 3.2 | 2.6 | 1.1 | 1.7 | 0.4 | 1.5 | 1.1 | 1.3 | 0.8 | 3.0 | |
| 10 | A. seminiferus PER | 11.2 | 11.4 | 10.7 | 9.9 | 10.3 | 9.7 | 2.7 | 2.6 | 3.1 | 2.4 | 2.2 | 2.8 | 2.2 | 2.5 | 2.7 | 2.4 | 2.6 | 3.2 | |
| 11 | A. sp. Anapu BRA | 11.9 | 11.7 | 12.0 | 10.1 | 11.9 | 10.0 | 3.3 | 3.2 | 2.5 | 2.3 | 2.3 | 3.0 | 2.6 | 2.7 | 2.8 | 2.6 | 2.8 | 3.0 | |
| 12 | A. franciscus GUF | 11.4 | 11.6 | 11.0 | 10.5 | 10.6 | 10.1 | 3.6 | 2.7 | 1.1 | 2.1 | 2.3 | 0.5 | 0.2 | 0.4 | 0.5 | 0.4 | 0.4 | 2.7 | |
| 13 | A. barbotini “B” GUF | 11.0 | 11.7 | 10.4 | 10.4 | 10.3 | 9.9 | 4.2 | 3.3 | 1.7 | 2.7 | 2.9 | 0.5 | 0.8 | 0.7 | 1.0 | 1.3 | 1.1 | 3.0 | |
| 14 | A. barbotini “A” GUF | 12.0 | 11.5 | 11.0 | 10.7 | 10.4 | 10.4 | 2.7 | 2.6 | 0.4 | 2.2 | 2.5 | 0.2 | 0.8 | 0.5 | 0.4 | 0.4 | 0.6 | 2.3 | |
| 15 | A. hoogmoedi GUF | 11.6 | 11.9 | 11.2 | 10.7 | 10.5 | 10.2 | 4.0 | 3.0 | 1.4 | 2.5 | 2.7 | 0.4 | 0.7 | 0.5 | 0.4 | 0.6 | 0.3 | 2.9 | |
| 16 | A. hoogmoedi GUY | 12.9 | 12.1 | 11.6 | 11.5 | 11.1 | 11.1 | 3.7 | 2.8 | 1.1 | 2.7 | 2.8 | 0.5 | 1.0 | 0.4 | 0.4 | 0.4 | 0.4 | 2.7 | |
| 17 | A. hoogmoedi REUA BRA | 11.4 | 11.3 | 10.7 | 10.4 | 10.3 | 9.9 | 3.8 | 2.9 | 1.3 | 2.3 | 2.5 | 0.4 | 1.2 | 0.4 | 0.6 | 0.4 | 0.2 | 2.7 | |
| 18 | A. hoogmoedi MA BRA | 12.8 | 12.1 | 12.0 | 11.3 | 11.1 | 11.3 | 3.1 | 3.0 | 0.8 | 2.5 | 2.7 | 0.4 | 1.0 | 0.6 | 0.3 | 0.4 | 0.2 | 2.4 | |
| 19 | A. manauensis sp. nov. BRA | 11.6 | 11.5 | 11.0 | 10.1 | 10.0 | 9.7 | 4.7 | 3.9 | 3.0 | 3.1 | 2.9 | 2.7 | 2.9 | 2.3 | 2.8 | 2.7 | 2.6 | 2.3 |
| Morphology | PC1 | PC2 | Call | PC1 | PC2 |
|---|---|---|---|---|---|
| SVL | 0.3731 | −0.0989 | Call_dur | −0.2697 | 0.3854 |
| HW | −0.3729 | −0.0944 | N_pulses | 0.3058 | 0.3564 |
| HL | −0.3726 | −0.0171 | Puls_dur_1 | −0.3007 | −0.0782 |
| EYDM | −0.3512 | −0.0744 | Interpulse_1 | −0.4138 | −0.0042 |
| ITNA | −0.3482 | 0.0172 | Puls_dur_2 | −0.2513 | 0.0300 |
| IOD | −0.2834 | −0.0745 | Interpulse_2 | −0.3626 | −0.3038 |
| EYNO | −0.0531 | −0.0017 | Puls_dur_3 | 0.0295 | 0.1371 |
| SW | −0.3230 | −0.1655 | Interpulse_3 | −0.2898 | 0.3132 |
| FOOT | 0.0910 | −0.5572 | Dur_3_first_puls | −0.4367 | −0.0368 |
| THBL | 0.2361 | −0.2680 | Dur_3_last_puls | −0.2934 | 0.3254 |
| HAND | 0.0977 | −0.5271 | Domi_frequency | −0.1183 | −0.4472 |
| TL | −0.2663 | −0.0910 | bandwidth | 0.0440 | −0.4483 |
| RDUL | −0.0798 | −0.5233 | – | – | – |
| Explanation (%) | 48.7 | 17.5 | Explanation (%) | 38.7 | 25.3 |
| Morphology | Holotype | Males (n = 11) | Females (n = 2) |
|---|---|---|---|
| SVL | 21.2 | 21.8 ± 2.4 (19.1–26.4) | 27.9–28.8 |
| SW | 6.5 | 7.1 ± 1.1 (5.7–8.6) | 8.4–8.5 |
| HW | 7.3 | 7.4 ± 0.5 (6.5–8.3) | 8.3–8.4 |
| HL | 7.5 | 7.8 ± 0.6 (6.8–8.7) | 8.9–9.2 |
| EYDM | 2.4 | 2.4 ± 0.2 (2.1–2.7) | 2.7–3.0 |
| EYNO | 2.4 | 2.5 ± 0.3 (2.1–3.0) | 2.8–3.0 |
| IOD | 3.0 | 2.9 ± 0.3 (2.5–3.6) | 3.1–3.1 |
| ITNA | 2.6 | 2.7 ± 0.2 (2.4–3.1) | 3.1–3.1 |
| RDUL | 6.3 | 6.7 ± 0.8 (6.0–8.2) | 8.7–10.0 |
| HAND | 5.0 | 5.2 ± 0.6 (4.9–6.6) | 6.4–7.5 |
| THBL | 2.2 | 2.2 ± 0.2 (1.9–2.5) | 2.5–3.0 |
| FOOT | 7.3 | 7.7 ± 1.1 (6.4–10.4) | 10.4–10.9 |
| TL | 9.8 | 10.3 ± 0.8 (9.8–11.9) | 12.1–12.9 |
| Acoustic Traits | Atelopus manauensis sp. nov. (n = 3) | Atelopus hoogmoedi Amapá (n = 4) | Atelopus hoogmoedi REBIO Uatumã and Pitinga River (n = 7) |
|---|---|---|---|
| Call_dur (ms) | 744 ± 84 (689–840) | 1164 ± 387 (873–1710) | 858 ± 89 (733–1071) |
| N_pulses | 19 ± 6 (15–26) | 29 ± 5 (23–35) | 29 ± 5 (21–37) |
| Puls_dur_1 (ms) | 7 ± 1 (6–8) | 7 ± 1 (6–9) | 6 ± 2 (3–11) |
| Puls_dur_2 (ms) | 6 ± 1 (5–7) | 8 ± 1 (7–9) | 7 ± 2 (4–12) |
| Puls_dur_3 (ms) | 14 ± 6 (8–19) | 14 ± 5 (10–22) | 15 ± 4 (7–27) |
| Interpulse_1 (ms) | 54 ± 16 (36–67) | 48 ± 33 (15–78) | 35 ± 9 (19–50) |
| Interpulse_2 (ms) | 50 ± 13 (40–65) | 37 ± 11 (23–47) | 32 ± 9 (15–54) |
| Interpulse_3 (ms) | 7 ± 3 (4–9) | 10 ± 3 (7–13) | 7 ± 2 (4–13) |
| Dur_3_first_puls (ms) | 132 ± 40 (92–172) | 120 ± 66 (61–178) | 88 ± 16 (56–116) |
| Dur_3_last_puls (ms) | 43 ± 6 (38–49) | 54 ± 5 (49–59) | 42 ± 7 (30–59) |
| Domi_frequency (Hz) | 3334 ± 263 (3088–3610) | 2825 ± 13 (2812–2838) | 2741 ± 161 (2498–3058) |
| Bandwidth (Hz) | 743 ± 151 (633–915) | 442 ± 127 (280–568) | 555 ± 94 (388–754) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jorge, R.F.; Ferrão, M.; Lima, A.P. Out of Bound: A New Threatened Harlequin Toad (Bufonidae, Atelopus) from the Outer Borders of the Guiana Shield in Central Amazonia Described through Integrative Taxonomy. Diversity 2020, 12, 310. https://doi.org/10.3390/d12080310
Jorge RF, Ferrão M, Lima AP. Out of Bound: A New Threatened Harlequin Toad (Bufonidae, Atelopus) from the Outer Borders of the Guiana Shield in Central Amazonia Described through Integrative Taxonomy. Diversity. 2020; 12(8):310. https://doi.org/10.3390/d12080310
Chicago/Turabian StyleJorge, Rafael F., Miquéias Ferrão, and Albertina P. Lima. 2020. "Out of Bound: A New Threatened Harlequin Toad (Bufonidae, Atelopus) from the Outer Borders of the Guiana Shield in Central Amazonia Described through Integrative Taxonomy" Diversity 12, no. 8: 310. https://doi.org/10.3390/d12080310
APA StyleJorge, R. F., Ferrão, M., & Lima, A. P. (2020). Out of Bound: A New Threatened Harlequin Toad (Bufonidae, Atelopus) from the Outer Borders of the Guiana Shield in Central Amazonia Described through Integrative Taxonomy. Diversity, 12(8), 310. https://doi.org/10.3390/d12080310