Abiotic Community Constraints in Extreme Environments: Epikarst Copepods as a Model System




Abstract

1. Introduction
1.1. Background
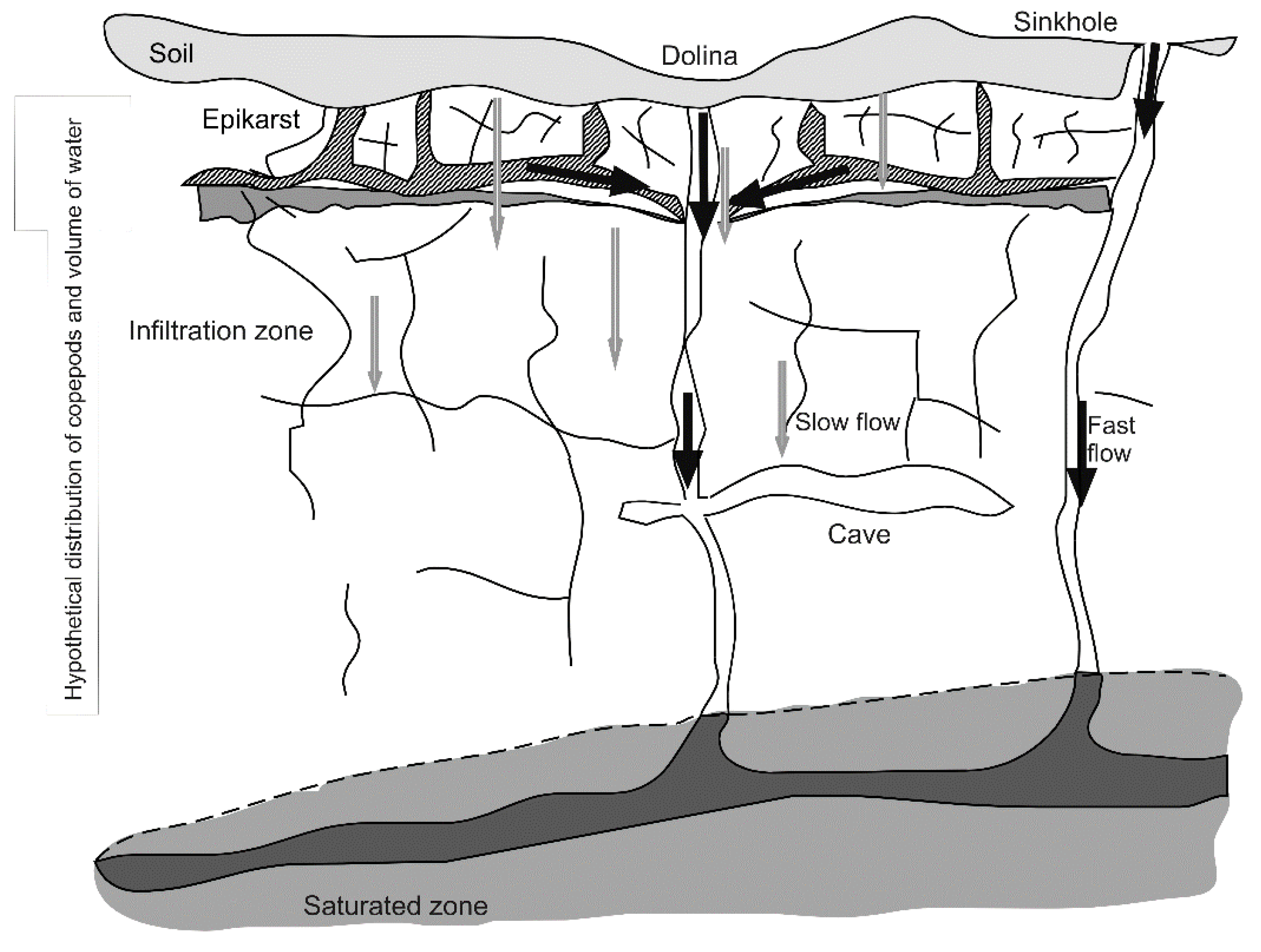
1.2. Epikarst as a Model Extreme Environment
1.3. Aims and Goals
2. Methods and Materials
2.1. Study Area and Sampling Procedures
2.2. Statistical Methods
3. Results
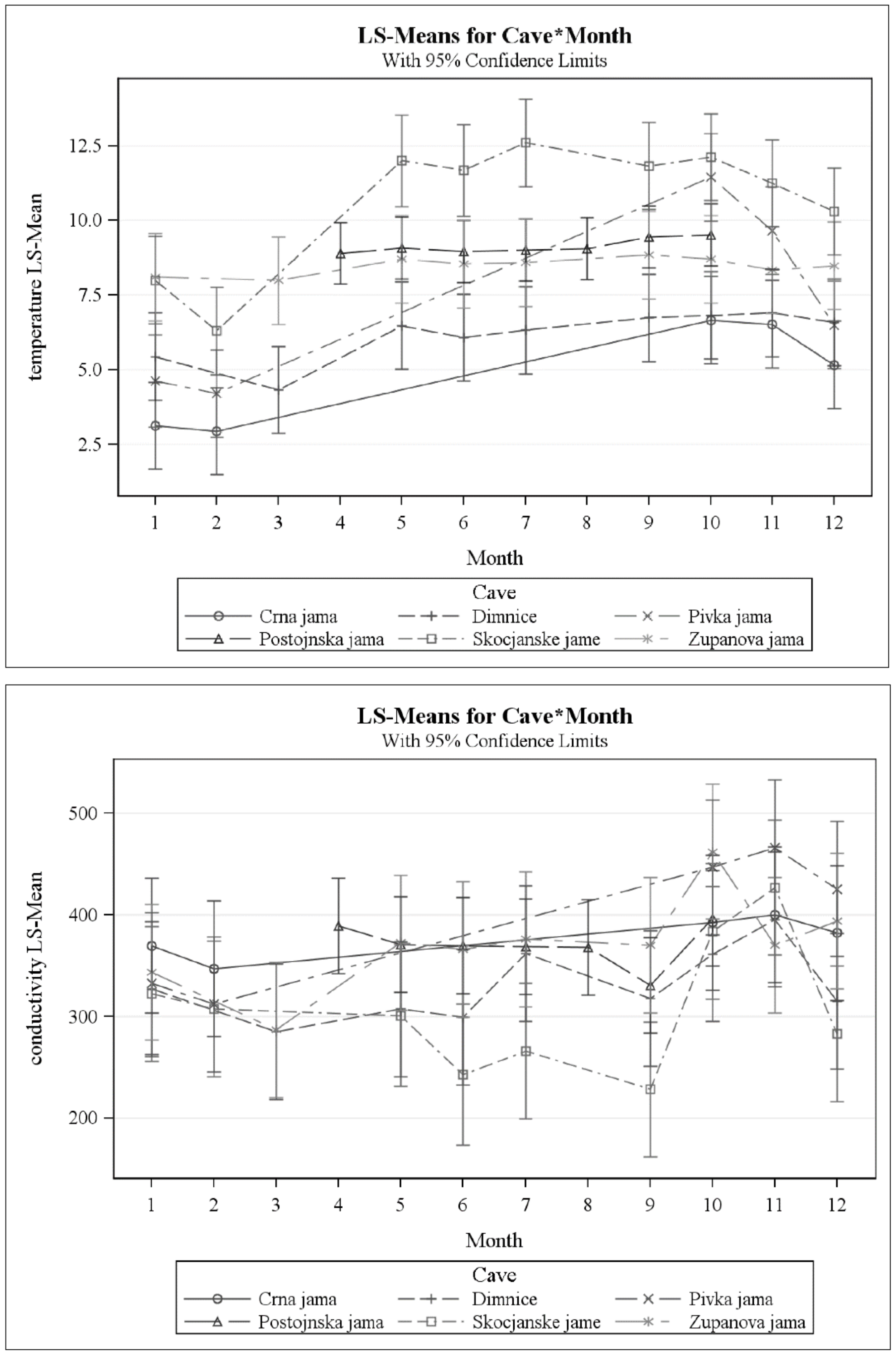
3.1. What Are the Physical and Chemical Conditions Present in Epikarst Water?
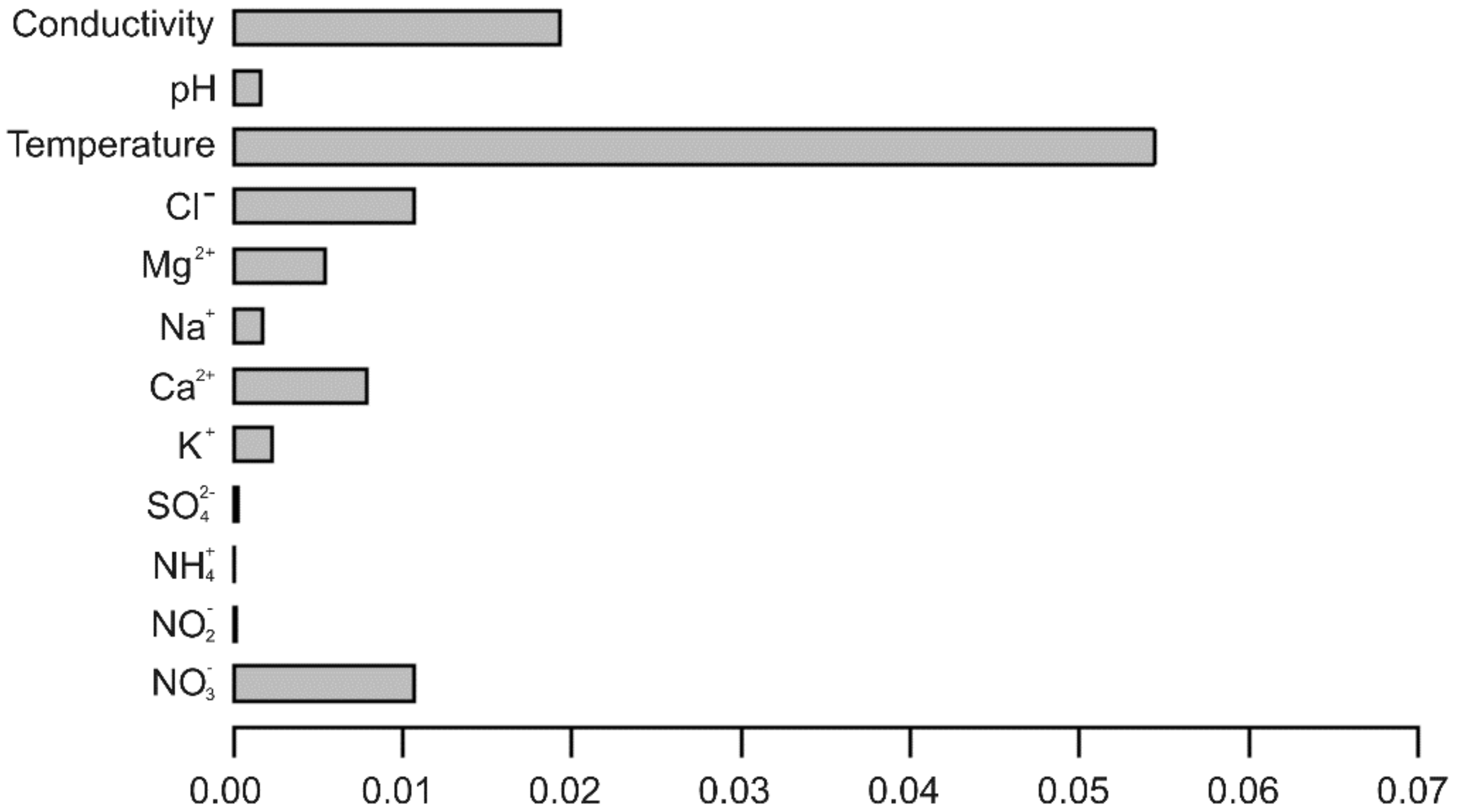
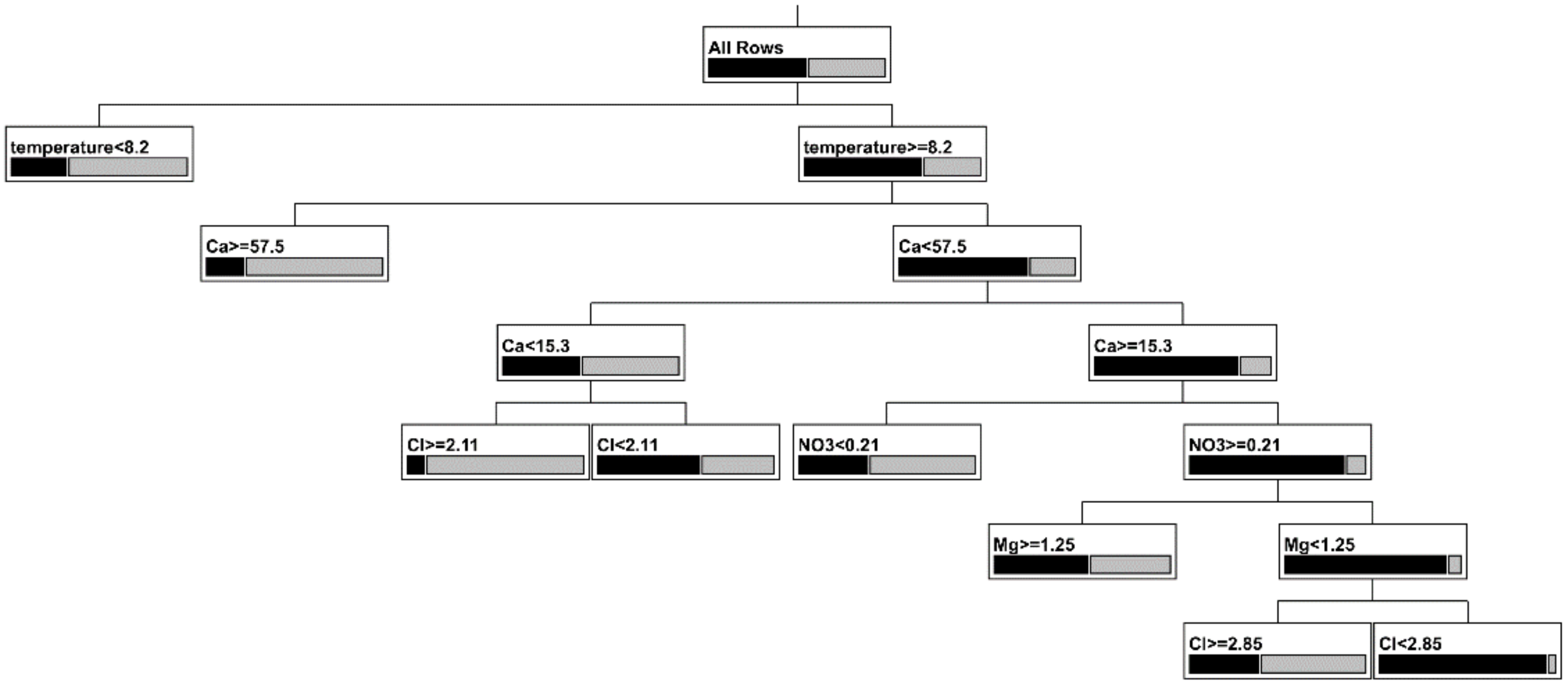
3.2. Which Variables Could Be Important in Limiting Copepod Distribution?
4. Discussion
4.1. Patterns of Variation of Physico-Chemical Factors
4.2. The Epikarst Copepod Physico-Chemical Niche
- Temperature,
- Ca2+,
- Mg2+,
- Cl−,
- NO3−.
- Temperature, which is likely a reflection of flow velocities rather than community structure.
- Calcium and perhaps magnesium ions, which are important, both as essential nutrients and in molting.
- Anthropogenically augmented ions—Cl− and perhaps NO3−—may indicate contamination from upgradient well water, or they may be surrogates for particular epikarst sites, where some unmeasured variable is important.
4.3. The Relationship between Community Physico-Chemical Niche and Individual Physico-Chemical Niches
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Engel, A.S. Chemolithoautotrophy. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: London, UK, 2019; pp. 267–276. [Google Scholar]
- Culver, D.C.; Pipan, T. Shallow Subterranean Habitats. Ecology, Evolution, and Conservation; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Christman, M.C. Sequential sampling for rare or geographically clustered populations. In Sampling Rare or Elusive Species; Thompson, W.L., Ed.; Island Press: Washington, DC, USA, 2004; pp. 134–148. [Google Scholar]
- Ter Braak, C.J.F.; Verdonschot, P.J.M. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 1015–1621. [Google Scholar] [CrossRef]
- Doledec, S.; Chessel, D.; Gimaret-Carpentier, C. Niche separation in community analysis: A new method. Ecology 2000, 81, 2914–2927. [Google Scholar] [CrossRef]
- Ward, D. The Biology of Deserts; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Van Dover, C. The Ecology of Deep-Sea Hydrothermal Vents; Princeton University Press: Princeton, NJ, USA, 2000. [Google Scholar]
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats, 2nd ed.; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Christiansen, K.A. Convergence and parallelism in Cave Entomobryinae. Evolution 1961, 15, 288–301. [Google Scholar] [CrossRef]
- Wiens, J.J.; Chippendale, P.T.; Hillis, D.M. When are phylogenetic analyses misled by convergence? A case study in Texas Cave Salamanders. Syst. Biol. 2003, 52, 501–514. [Google Scholar] [CrossRef]
- Pipan, T.; Culver, D.C. Convergence and divergence in the subterranean realm: A reassessment. Biol. J. Linn. Soc. 2012, 107, 1–14. [Google Scholar] [CrossRef]
- Protas, M.E.; Jeffery, W.R. Evolution and development in cave animals: From fish to crustaceans. Wiley Interdiscip. Rev. Dev. Biol. 2013, 1, 823–845. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. Shifting paradigms of the evolution of cave life. Acta Carsologica 2015, 44, 415–425. [Google Scholar] [CrossRef]
- Venarsky, M.P.; Huntsman, B.M.; Huryn, A.D.; Benstead, J.P.; Kuhajda, B.R. Quantitative food web analysis supports the energy limitation hypothesis in cave stream ecosystems. Oecologia 2014, 176, 859–869. [Google Scholar] [CrossRef]
- Venarsky, M.P.; Huntsman, B.M. Food webs in caves. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer: Gland, Switzerland, 2018; pp. 309–330. [Google Scholar]
- Bakalowicz, M. Epikarst. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: London, UK, 2019; pp. 394–398. [Google Scholar]
- Pipan, T.; Culver, D.C. Copepod distribution as an indicator of epikarst system connectivity. Hydrogeol. J. 2007, 15, 817–822. [Google Scholar] [CrossRef]
- Simon, K.S.; Pipan, T.; Culver, D.C. A conceptual model of the flow and distribution of organic carbon in caves. J. Caves Karst Stud. 2007, 69, 279–284. [Google Scholar]
- Pipan, T.; Culver, D.C. The unity and diversity of the subterranean realm with respect to invertebrate body size. J. Caves Karst Stud. 2017, 79, 1–9. [Google Scholar] [CrossRef]
- Pipan, T. Epikarst—A Promising Habitat; Založba ZRC: Ljubljana, Slovenia, 2005. [Google Scholar]
- Pipan, T.; Holt, N.; Culver, D.C. How to protect a diverse, poorly known, inaccessible fauna: Identification of source and sink habitats in the epikarst. Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 748–755. [Google Scholar] [CrossRef]
- Meleg, I.N.; Fiers, F.; Moldovan, O.T. Assessing copepod (Crustacea: Copepoda) species richness at different spatial scales in Northwestern Romanian caves. Subterr. Biol. 2011, 9, 103–112. [Google Scholar] [CrossRef]
- Meleg, I.N.; Moldovan, O.T.; Iepure, S.; Fiers, F.; Brad, T. Diversity patterns of fauna in dripping water of caves from Transylvania. Ann. Limnol. Int. J. Lim. 2011, 47, 185–197. [Google Scholar] [CrossRef]
- Papi, F.; Pipan, T. Ecological studies of an epikarst community in Snežna jama na planini Arto—An ice cave in North Central Slovenia. Acta Carsol. 2011, 40, 505–513. [Google Scholar] [CrossRef]
- Fong, D.W.; Culver, D.C.; Hobbs, H.H., III; Pipan, T. The Invertebrate Cave Fauna of West Virginia, 2nd ed.; West Virginia Speleological Survey: Barrackville, WV, USA, 2007. [Google Scholar]
- Pipan, T. Ekologija Ceponožnih Rakov (Crustacea: Copepoda) v Prenikajoči Vodi Izbranih Kraških jam. Ph.D. Thesis, University of Ljubljana, Ljubljana, Slovenia, May 2003. [Google Scholar]
- Pipan, T.; Blejec, A.; Brancelj, A. Multivariate analysis of copepod assemblages in epikarstic waters of some Slovenian caves. Hydrobiologia 2006, 559, 213–223. [Google Scholar] [CrossRef]
- Strobl, C.; Boulestein, A.L.; Kneib, T.; Augustin, T.; Zeileis, A. Conditional variable importance for random forests. BMC Bioinform. 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Strobl, C.; Malley, J.; Tutz, G. An introduction to recursive partitioning: Rationale, application, and characteristics of classification and regression trees, bagging, and random forests. Psychol. Methods 2009, 14, 323–348. [Google Scholar] [CrossRef] [PubMed]
- Nicodemus, K.K.; Malley, J.D.; Strobl, C.; Ziegler, A. The behavior of random forest permutation-based variable importance under predictor correlation. BMC Bioinfom. 2010, 11. [Google Scholar] [CrossRef]
- Herman, J.S. Water chemistry in caves. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: London, UK, 2019; pp. 1136–1143. [Google Scholar]
- Kogovšek, J. Characteristics of Percolation Through the Karst Vadose Zone; ZRC Publishing: Ljubljana, Slovenia, 2010. [Google Scholar]
- Kluge, T.; Reichelman, D.F.C.; Wieser, M.; Spötl, C.; Sültenfuß, J.; Schröder Ritzrau, A.; Niggemann, S.; Aeschbach-Hertig, W. Dating cave drip water by tritium. J. Hydrol. 2010, 394, 396–406. [Google Scholar] [CrossRef]
- Gospodarič, R. Speleogeological data about Taborska jama and its vicinity. Acta Carsol. 1987, 16, 19–34. [Google Scholar]
- Kogovšek, J. Natural purification of sanitary sewage during the vertical percolation in Pivka jama. Acta Carsol. 1987, 16, 123–139. [Google Scholar]
- Mercado, A. Nitrate and chloride pollution of aquifers; a regional study with the aid of a single cell model. Water Resour. Res. 1976, 12, 731–747. [Google Scholar] [CrossRef]
- Williams, P.W. The role of the epikarst in karst and cave hydrogeology: A review. Int. J. Speleol. 2008, 37, 1–10. [Google Scholar] [CrossRef]
- Berthelin, R.; Hartmann, A. The shallow subsurface of karst systems: Review and directions. In Eurokarst 2018, Besançon: Advances in the Hydrogeology and Karst and Carbonate Reservoirs; Bertrand, S., Denimal, M., Steinmann, R.P., Eds.; Springer Nature: Gland, Switzerland, 2020; pp. 61–68. [Google Scholar]
- Badino, G. Underground meteorology—“What’s the weather underground? ” Acta Carsol. 2010, 39, 427–448. [Google Scholar] [CrossRef]
- Covington, M.D.; Luhmann, A.J.; Gabrovšek, F.; Wicks, C.M. Mechanisms of heat exchange between water and rock in karst conduits. Water Resour. Res. 2011, 47. [Google Scholar] [CrossRef]
- Covington, M.D.; Perne, M. Consider a cylindrical cave: A physicists’ view of cave and karst science. Acta Carsol. 2015, 44, 363–380. [Google Scholar] [CrossRef]
- Rouch, R. Recherches sur lex eaux souterraines—12—Le système karstique du Baget. 1. La phénomène d’hémorragie’ au niveau de l’exutoire principal. Ann. Spéléol. 1970, 25, 665–709. [Google Scholar]
- Glazier, D.S.; Horne, M.T.; Lehman, M.E. Abundance, body composition and reproductive output of Gammarus minus (Crustacea: Amphipoda) in ten cold springs differing in pH and ionic content. Freshw. Biol. 1992, 28, 149–163. [Google Scholar] [CrossRef]
- Katz, B.G. Nitrate contamination of karst groundwater. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: London, UK, 2019; pp. 756–760. [Google Scholar]
- Simon, K.S.; Benfield, E.F. Ammomium retention and whole-stream metabolism in cave streams. Hydrobiologia 2002, 482, 31–39. [Google Scholar] [CrossRef]
- Pasternak, K.; Kocot, J.; Horecka, A. Biochemistry of magnesium. J. Elem. 2010, 15, 601–616. [Google Scholar] [CrossRef]
- Barton, H.A. Starving artists: Bacterial oligotrophic heterotrophy in caves. In Microbial Life of Cave Systems; Engel, A.S., Ed.; De Gruyter: Berlin, Germany, 2015; pp. 79–104. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Postojnska Jama | Pivka Jama | Črna Jama | Škocjanske Jame | Dimnice | Županova Jama |
|---|---|---|---|---|---|---|
| Jan | X | X | X | X | X | |
| Feb | X | X | X | |||
| Mar | X | X | ||||
| Apr | X | |||||
| May | X | X | X | X | ||
| Jun | X | X | X | X | ||
| Jul | X | X | X | X | ||
| Aug | X | |||||
| Sep | X | X | X | X | ||
| Oct | X | X | X | X | X | X |
| Nov | X | X | X | X | X | |
| Dec | X | X | X | X | X |
| Statistic | Temperature | Conductivity | pH | Ca2+ | K+ | Mg2+ | Na+ | NH4+ | Cl− | NO2− | NO3− | SO42− |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | 8.16 | 356.64 | 7.79 | 37.71 | 0.47 | 0.93 | 1.36 | 0.096 | 2.34 | 0.0030 | 2.84 | 6.18 |
| S.E. of Mean | 0.106 | 4.641 | 0.012 | 0.747 | 0.013 | 0.025 | 0.086 | 0.005 | 0.219 | 0.0004 | 0.360 | 0.135 |
| Median | 8.91 | 365.00 | 7.73 | 40.60 | 0.41 | 0.91 | 0.95 | 0.084 | 1.44 | 0.000 | 0.78 | 5.95 |
| S.E. of Median | 0.043 | 4.91 | 0.012 | 1.47 | 0.013 | 0.027 | 0.043 | 0.002 | 0.05 | 0.001 | 0.083 | 0.15 |
| NO3− | NO2− | NH4+ | SO42− | K+ | Ca2+ | Na+ | Mg+ | Cl− | °C | pH | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NO3− | 1 | ||||||||||
| NO2− | 0.202 * | 1 | |||||||||
| NH4+ | 0.140 * | 0.065 | 1 | ||||||||
| SO42− | 0.075 | 0.025 | 0.061 | 1 | |||||||
| K+ | 0.500 * | 0.316 * | 0.327 * | 0.261 * | 1 | ||||||
| Ca2+ | 0.194 * | −0.034 | 0.034 | 0.492 * | 0.282 * | 1 | |||||
| Na+ | 0.622 * | 0.198 * | 0.264 * | 0.183 * | 0.470 * | 0.210 * | 1 | ||||
| Mg2+ | 0.247 * | 0.023 | 0.261 * | 0.588 * | 0.420 * | 0.461 * | 0.406 * | 1 | |||
| Cl− | 0.480 * | 0.030 | 0.162 * | 0.023 | 0.088 | 0.157 * | 0.687 * | 0.216 * | 1 | ||
| Temperature | −0.113 | 0.094 | −0.029 | 0.072 | −0.062 | −0.271 * | −0.208 * | 0.022 | −0.258 * | 1 | |
| pH | 0.121 | −0.009 | −0.035 | −0.074 | 0.156 * | −0.023 | 0.058 | −0.105 | 0.021 | −0.039 | 1 |
| Conductivity | 0.294 * | 0.055 | −0.023 | −0.228 * | −0.039 | 0.121 | 0.005 | −0.053 | 0.085 | −0.047 | −0.123 |
| Covariance Parameter Estimates (SE) for the Random Effects | ||||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Transformation | Sampling Site within Cave | Residual | Numerator df | Denominator df | F-Value | p | Caves with Monthly Differences |
| Presence of copepods | Binary | 2.050 (1.193) | na | 37 | 179 | 0.74 | 0.86 | None |
| Temperature | None | 1.603 (0.462) | 1.070 (0.113) | 43 | 178.1 | 11.64 | <0.0001 | All caves except Postojnska jama and Županova jama |
| Conductivity | None | 3595.6 (1026.4) | 1935.9 (204.8) | 43 | 177.8 | 5.22 | <0.0001 | All caves except Črna jama |
| pH | None | 0.00566 (0.00239) | 0.0263 (0.0028) | 43 | 179.4 | 3.31 | <0.0001 | Pivka jama, Postojnska jama, Županova jama |
| Ca2+ | None | 17.90 (7.37) | * | 43 | 96.5 | 16.58 | <0.0001 | All caves |
| K+ | None | 0.00923 (0.00408) | ** | 43 | 93.5 | 3.00 | <0.0001 | Pivka jama, Postojnska jama, Županova jama |
| Mg2+ | None | 0.0838 (0.0257) | 0.0772 (0.0082) | 43 | 176.2 | 5.26 | <0.0001 | All caves except Županova jama |
| Na+ | Log | 0.232 (0.075) | 0.320 (0.035) | 43 | 176.4 | 4.71 | <0.0001 | All caves except Županova jama |
| NH4+ | None | 0 | *** | 43 | 105.8 | 2.09 | 0.0012 | Postojnska jama |
| Cl− | Square root | 0.258 (0.071) | 0.0841 (0.0089) | 43 | 177.3 | 4.66 | <0.0001 | All caves except Postojnska jama and Županova jama |
| NO3− | Log | 0.888 (0.242) | 0.232 (0.025) | 43 | 177.1 | 7.77 | <0.0001 | All caves |
| SO42− | None | 1.449 (0.501) | 3.126 (0.301) | 43 | 177.2 | 9.25 | <0.0001 | All caves |
| Number of Variables | χ2 | Variables |
|---|---|---|
| 1 | 12.978 | Temperature |
| 2 | 18.265 | Temperature, Conductivity |
| 3 | 22.426 | Temperature, Conductivity, K+ |
| 4 | 24.350 | Temperature, Conductivity, K+, NO2− |
| 5 | 26.117 | Temperature, Conductivity, K+, NO2−, Na+ |
| 6 | 27.782 | Temperature, Conductivity, K+, NO2−, Na+, Cl− |
| 7 | 28.441 | Temperature, Conductivity, K+, NO2−, Na+, Cl−, Ca2+ |
| Temperature | Ca2+ | Cl− | NO3− | Mg2+ | Frequency with Copepods | Frequency without Copepods | n |
|---|---|---|---|---|---|---|---|
| <8.2 | 0.668 | 0.332 | 85 | ||||
| ≥8.2 | ≥57.5 | 0.758 | 0.242 | 22 | |||
| ≥8.2 | <15.3 | ≥2.11 | 0.843 | 0.157 | 9 | ||
| ≥8.2 | <15.3 | <2.11 | 0.410 | 0.590 | 22 | ||
| ≥8.2 | <57.5 and ≥15.3 | <0.21 | 0.587 | 0.413 | 15 | ||
| ≥8.2 | <57.5 and ≥15.3 | ≥0.21 | ≥1.25 | 0.447 | 0.553 | 11 | |
| ≥8.2 | <57.5 and ≥15.3 | ≥2.85 | ≥0.21 | <1.25 | 0.556 | 0.444 | 5 |
| ≥8.2 | <57.5 and ≥15.3 | <2.85 | ≥0.21 | <1.25 | 0.052 | 0.948 | 83 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pipan, T.; Christman, M.C.; Culver, D.C. Abiotic Community Constraints in Extreme Environments: Epikarst Copepods as a Model System. Diversity 2020, 12, 269. https://doi.org/10.3390/d12070269
Pipan T, Christman MC, Culver DC. Abiotic Community Constraints in Extreme Environments: Epikarst Copepods as a Model System. Diversity. 2020; 12(7):269. https://doi.org/10.3390/d12070269
Chicago/Turabian StylePipan, Tanja, Mary C. Christman, and David C. Culver. 2020. "Abiotic Community Constraints in Extreme Environments: Epikarst Copepods as a Model System" Diversity 12, no. 7: 269. https://doi.org/10.3390/d12070269
APA StylePipan, T., Christman, M. C., & Culver, D. C. (2020). Abiotic Community Constraints in Extreme Environments: Epikarst Copepods as a Model System. Diversity, 12(7), 269. https://doi.org/10.3390/d12070269