Morphological Disparity of the Humerus in Modern Birds

 , and
, and
Abstract
1. Introduction
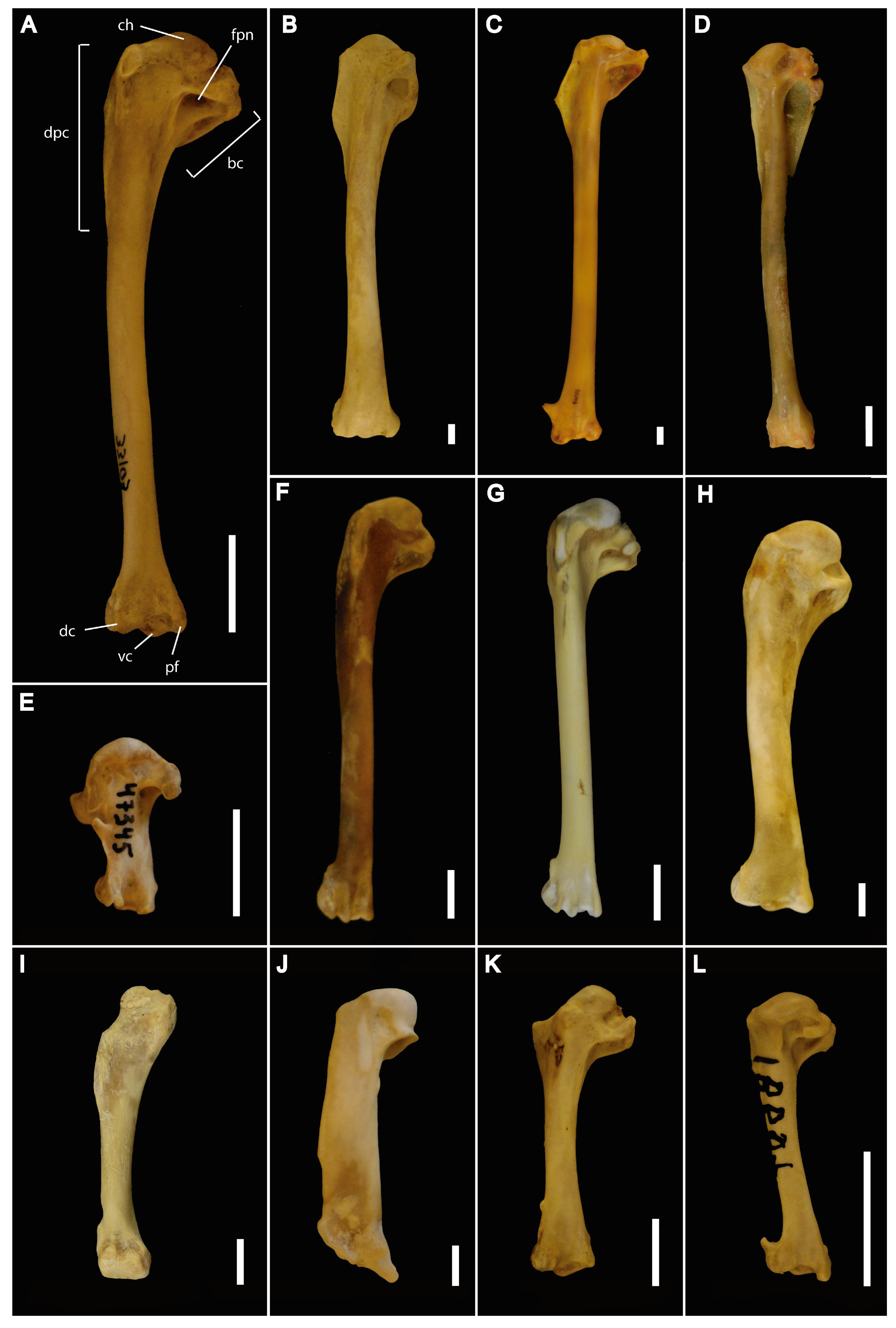
2. Material and Methods
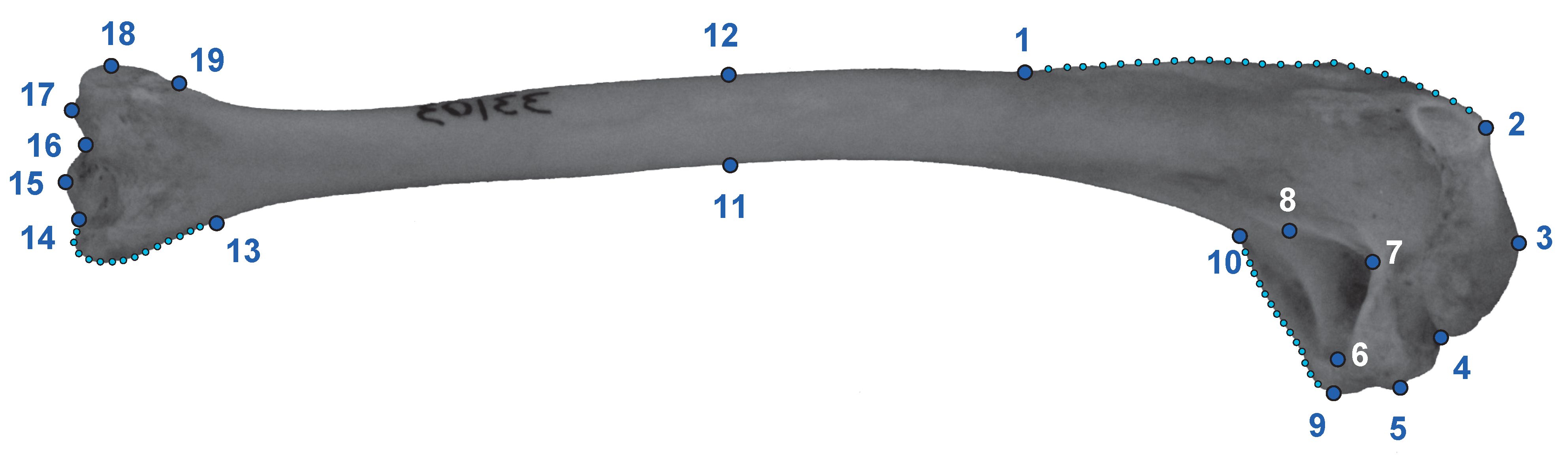
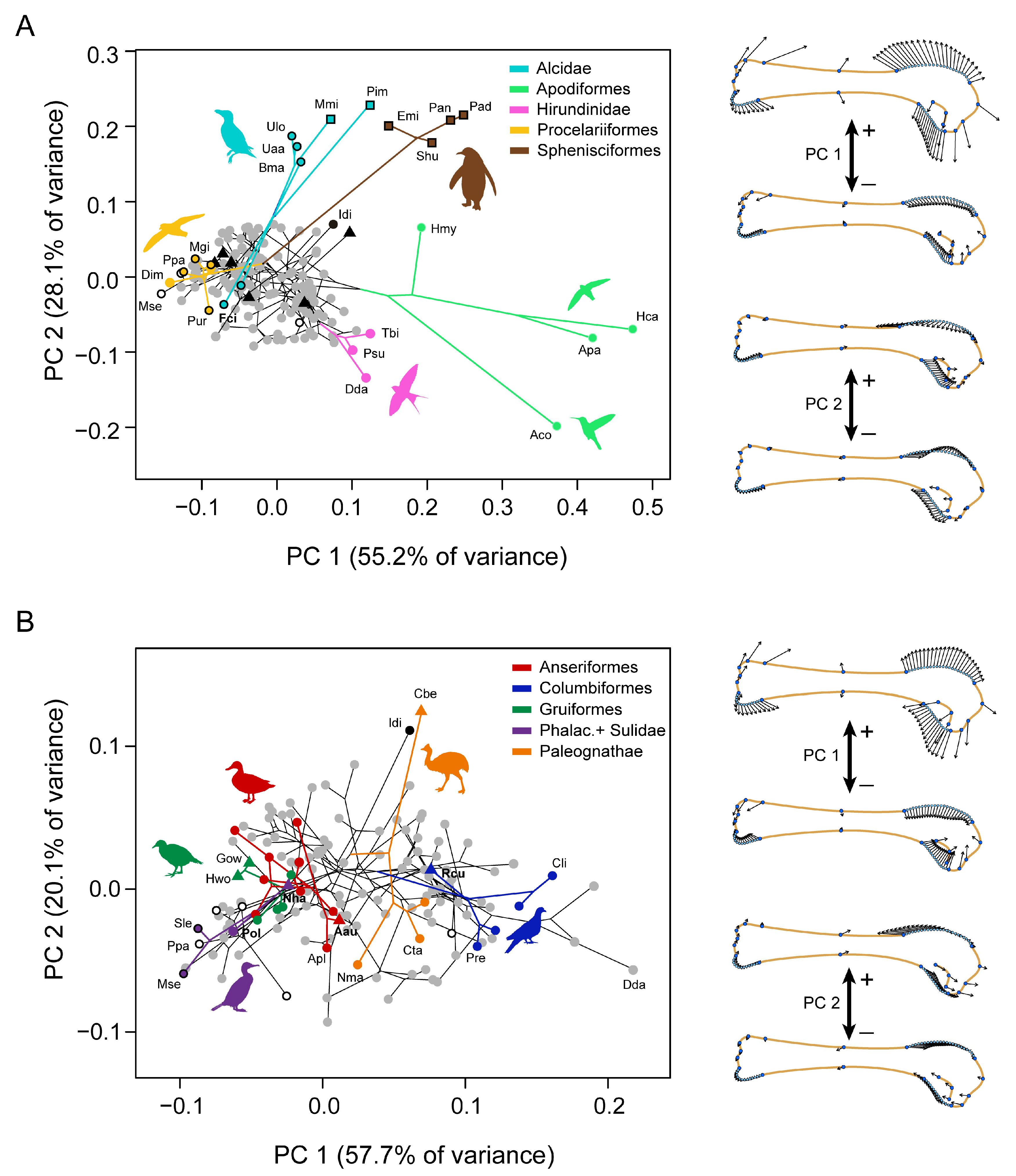
2.1. Data Acquisition
2.2. Analyses
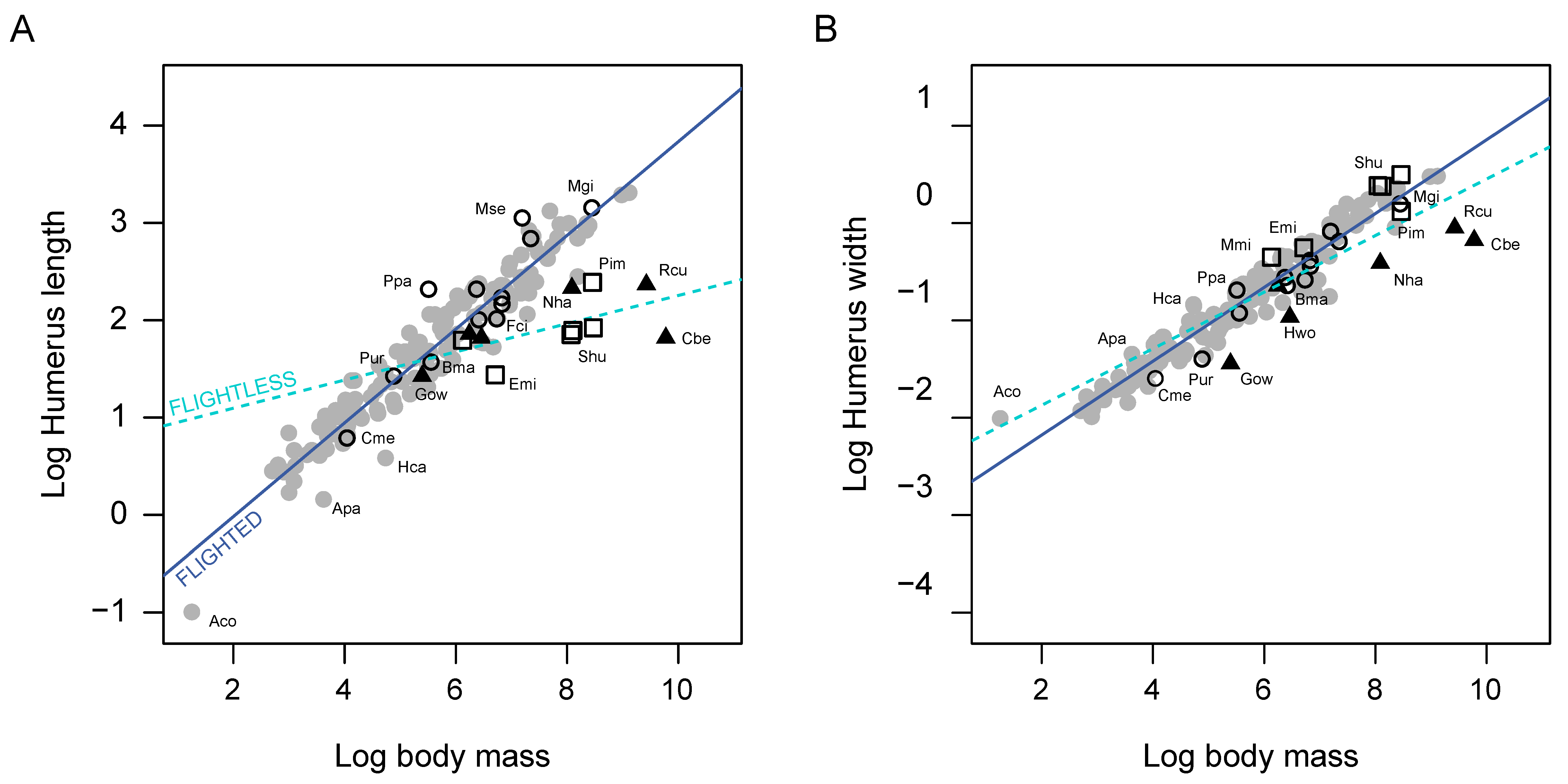
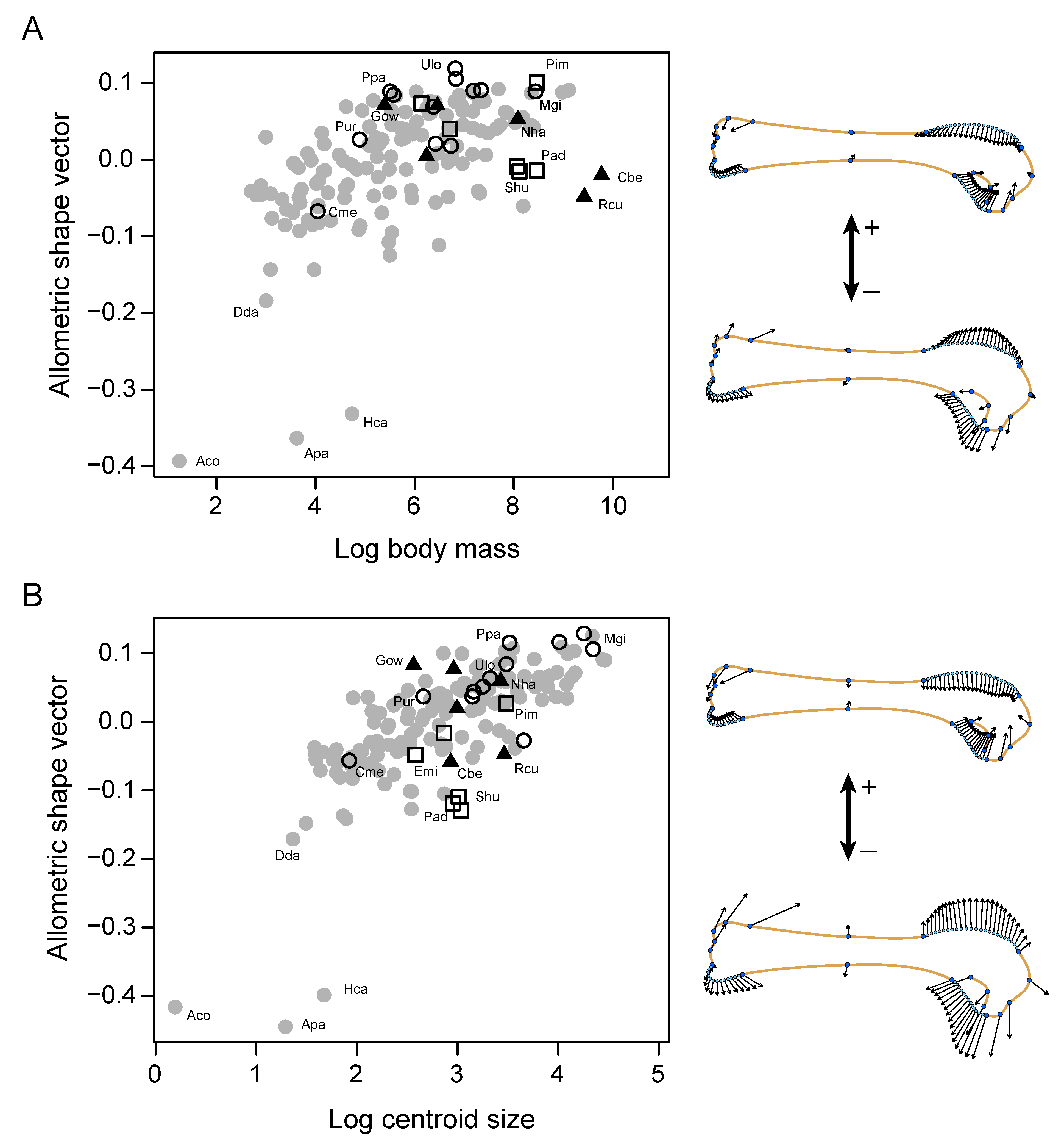
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barrowclough, G.F.; Cracraft, J.; Klicka, J.; Zink, R.M. How many kinds of birds are there and why does it matter? PLoS ONE 2016, 11, e0166307. [Google Scholar] [CrossRef] [PubMed]
- Norberg, U.L. Flight and scaling of flyers in nature. Flow Phenom. Nat. 2007, 1, 120–154. [Google Scholar]
- Butler, P.J. The physiological basis of bird flight. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150384. [Google Scholar] [CrossRef] [PubMed]
- Tobalske, B.W. Avian Flight. In Handbook of Bird Biology, 3rd ed.; Lovette, I.J., Fitzpatrick, J.W., Eds.; Wiley-Blackwell: The Cornell Lab of Ornithology: Ithaca, NY, USA, 2016; pp. 149–169. [Google Scholar]
- Pennycuick, C.J. Modelling the Flying Bird, 1st ed.; AP Theoretical Ecology Series; Academic Press: Oxford, UK, 2008; Volume 5, p. 496. [Google Scholar]
- Habib, M. The structural mechanics and evolution of aquaflying birds. Biol. J. Linn. Soc. 2010, 99, 687–698. [Google Scholar] [CrossRef]
- Smith, N.A.; Clarke, J.A. Osteological histology of the Pan-Alcidae (Aves, Charadriiformes): Correlates of wing-propelled diving and flightlessness. Anat. Rec. 2014, 297, 188–199. [Google Scholar] [CrossRef]
- De Margerie, E.; Sanchez, S.; Cubo, J.; Castanet, J. Torsional resistance as a principal component of the structural design of long bones: Comparative multivariate evidence in birds. Anat. Record Part A 2005, 282, 49–66. [Google Scholar]
- Nudds, R.L.; Dyke, G.J.; Rayner, J.M.V. Avian brachial index and wing kinematics: Putting movement back into bones. J. Zool. 2007, 272, 218–226. [Google Scholar] [CrossRef]
- Habib, M.B.; Ruff, C. The effects of locomotion on the structural characteristics of avian limb bones. Zool. J. Linn. Soc. 2008, 153, 601–624. [Google Scholar] [CrossRef]
- Simons, E.L.; Hieronymus, T.L.; O’Connor, P.M. Cross sectional geometry of the forelimb skeleton and flight mode in pelecaniform birds. J. Morphol. 2011, 272, 958–971. [Google Scholar] [CrossRef]
- Raikow, R.J. Locomotor system. In Form and Function in Birds; King, A.S., McLellan, J., Eds.; Academic Press: Cambridge, MA, USA, 1985; Volume 3, pp. 57–147. [Google Scholar]
- Baumel, J.J.; King, A.S.; Breazile, J.; Evans, H.; Vanden Berge, J.C. Handbook of Avian Anatomy: Nomina Anatomica Avium, 2nd ed.; Harvard Univ Nuttall Ornithological Club: Cambridge, MA, USA, 1993; p. 779. [Google Scholar]
- Bright, J.A.; Marugán-Lobón, J.; Cobb, S.N.; Rayfield, E.J. The shapes of bird beaks are highly controlled by nondietary factors. Proc. Natl. Acad. Sci. USA 2016, 113, 5352–5357. [Google Scholar] [CrossRef]
- Marugán-Lobón, J.; Watanabe, A.; Kawabe, S. Studying avian encephalization with geometric morphometrics. J. Anat. 2016, 229, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Cooney, C.R.; Bright, J.A.; Capp, E.J.; Chira, A.M.; Hughes, E.C.; Moody, C.J.; Nouri, L.O.; Varley, Z.K.; Thomas, G.H. Mega-evolutionary dynamics of the adaptive radiation of birds. Nature 2017, 542, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Felice, R.N.; Goswami, A. Developmental origins of mosaic evolution in the avian cranium. Proc. Natl. Acad. Sci. USA 2018, 115, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Navalón, G.; Bright, J.A.; Marugán-Lobón, J.; Rayfield, E.J. The evolutionary relationship among beak shape, mechanical advantage, and feeding ecology in modern birds. Evolution 2019, 73, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Navalón, G.; Marugán-Lobón, J.; Bright, J.A.; Cooney, C.R.; Rayfield, E.J. The consequences of craniofacial integration for the adaptive radiations of Darwin’s finches and Hawaiian honeycreepers. Nat. Ecol. Evol. 2020, 4, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Shatkovska, O.V.; Ghazali, M. Integration of skeletal traits in some passerines: Impact (or the lack thereof) of body mass, phylogeny, diet and habitat. J. Anat. 2020, 236, 274–287. [Google Scholar] [CrossRef]
- Close, R.A.; Rayfield, E.J. Functional morphometric analysis of the furcula in Mesozoic birds. PLoS ONE 2012, 7, e36664. [Google Scholar] [CrossRef]
- Wang, X.; Clarke, J.A. The evolution of avian wing shape and previously unrecognized trends in covert feathering. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151935. [Google Scholar] [CrossRef]
- Watanabe, J. Clade-specific evolutionary diversification along ontogenetic major axes in avian limb skeleton. Evolution 2018, 72, 2632–2652. [Google Scholar] [CrossRef]
- Nebreda, S.M.; Navalón, G.; Menéndez, I.; Sigurdsen, T.; Chiappe, L.M.; Marugán-Lobón, J. Disparity and macroevolutionary transformation of the maniraptoran manus. Bull. Am. Mus. Nat. Hist. in press.
- Navalón, G.; Meng, Q.; Marugán-Lobón, J.; Zhang, Y.; Wang, B.; Xing, H.; Liu, D.; Chiappe, L. Diversity and evolution of the Confuciusornithidae: Evidence from a new 131-million-year-old specimen from the Huajiying Formation in NE China. J. Asian Earth Sci. 2018, 152, 12–22. [Google Scholar] [CrossRef]
- Mayr, G. Avian Evolution: The Fossil Record of Birds and its Paleobiological Significance; John Wiley and Sons: Chichester, UK, 2017; p. 306. [Google Scholar]
- Olmos, M.; Casinos, A.; Cubo, J. Limb allometry in birds. Annales des Sciences Naturelles-Zoologie et Biologie Animale 1996, 17, 39–49. [Google Scholar]
- Cubo, J.; Casinos, A. Biomechanical significance of cross-sectional geometry of avian long bones. Eur. J. Morphol. 1998, 36, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Nudds, R.L. Wing-bone length allometry in birds. J. Avian Biol. 2007, 38, 515–519. [Google Scholar] [CrossRef]
- Sullivan, T.N.; Meyers, M.A.; Arzt, E. Scaling of bird wings and feathers for efficient flight. Sci. Adv. 2019, 5, eaat4269. [Google Scholar] [CrossRef]
- Sievwright, H.; Macleod, N. Eigensurface analysis, ecology, and modelling of morphological adaptation in the falconiform humerus (Falconiformes: Aves). Zool. J. Linn. Soc. 2012, 165, 390–419. [Google Scholar] [CrossRef][Green Version]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. A field comes of age: Geometric morphometrics in the 21st century. Hystrix 2013, 24, 7. [Google Scholar]
- Prum, R.O.; Berv, J.S.; Dornburg, A.; Field, D.J.; Townsend, J.P.; Lemmon, E.M.; Lemmon, A.R. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 2015, 526, 569–573. [Google Scholar] [CrossRef]
- Marugán-Lobón, J.; Buscalioni, A.D. Geometric morphometrics in macroevolution: Morphological diversity of the skull in modern avian form in contrast to some theropods dinosaurs. In Morphometrics: Applications in Biology and Paleontology; Elewa, A., Ed.; Springer: New York, NY, USA, 2004; pp. 157–171. [Google Scholar]
- Hume, J.P.; Steel, L. Fight club: A unique weapon in the wing of the solitaire, Pezophaps solitaria (Aves: Columbidae), an extinct flightless bird from Rodrigues, Mascarene Islands. Biol. J. Linn. Soc. 2013, 110, 32–44. [Google Scholar] [CrossRef]
- Clarke, J.A. Morphology, phylogenetic taxonomy, and systematics of Ichthyornis and Apatornis (Avialae: Ornithurae). Bull. Am. Mus. Nat. Hist. 2004, 286, 1–179. [Google Scholar] [CrossRef]
- Porras-Múzquiz, H.G.; Chatterjee, S.; Lehman, T.M. The carinate bird Ichthyornis from the Upper Cretaceous of Mexico. Cretac. Res. 2014, 51, 148–152. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.5. 2011. Available online: http://mesquiteproject.org (accessed on 1 April 2020).
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A resource for timelines, timetrees, and divergence times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef] [PubMed]
- Del Hoyo, J.; Elliott, A.; Sargatal, J.; Christie, D.A.; Kirwan, G. (Eds.) Handbook of the Birds of the World Alive; Lynx Edicions: Barcelona, Spain, 2020; Available online: http://www.hbw.com/ (accessed on 1 April 2020).
- Myers, P.; Espinosa, R.; Parr, C.S.; Jones, T.; Hammond, G.S.; Dewey, T.A. The Animal Diversity Web (online). 2020. Available online: https://animaldiversity.org (accessed on 1 April 2020).
- Serrano, F.J.; Palmqvist, P.; Sanz, J.L. Multivariate analysis of neognath skeletal measurements: Implications for body mass estimation in Mesozoic birds. Zool. J. Linn. Soc. 2015, 173, 929–955. [Google Scholar] [CrossRef]
- Smith, N.A. Evolution of body mass in the Pan-Alcidae (Aves, Charadriiformes): The effects of combining neontological and paleontological data. Paleobiology 2016, 42, 8–26. [Google Scholar] [CrossRef]
- Howard, H. A review of the extinct avian genus, Mancalla. Los Angel. Cty. Mus. Contrib. Sci. 1970, 203, 1–12. [Google Scholar]
- van Heteren, A.H.; van Dierendonck, R.C.; van Egmond, M.A.; Sjang, L.; Kreuning, J. Neither slim nor fat: Estimating the mass of the dodo (Raphus cucullatus, Aves, Columbiformes) based on the largest sample of dodo bones to date. PeerJ 2017, 5, e4110. [Google Scholar] [CrossRef]
- Rohlf, F.J. TPS Dig v.2.25; Department of Ecology and Evolution, State University of New York: Stony Brook, NY, USA, 2016; Available online: http://life.bio.sunysb.edu/morph/ (accessed on 1 April 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; Version 3.4.1; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://cran.R-project.org (accessed on 1 April 2020).
- Adams, D.C.; Collyer, M.; Kaliontzopoulou, A.; Sherratt, E. Geomorph: Software for Geometric Morphometric Analyses. 2016. Available online: https://cran.r-project.org/web/packages/geomorph/index.html (accessed on 1 April 2020).
- Dryden, I.L.; Mardia, K. Statistical Analysis of Shape; Wiley: Chichester, UK, 1998; p. 347. [Google Scholar]
- Gunz, P.; Mitteroecker, P. Semilandmarks: A method for quantifying curves and surfaces. Hystrix 2013, 24, 103–109. [Google Scholar]
- Collyer, M.L.; Sekora, D.J.; Adams, D.C. A method for analysis of phenotypic change for phenotypes described by high-dimensional data. Heredity 2015, 115, 357–365. [Google Scholar] [CrossRef]
- Revell, L.J. Phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Maddison, W.P. Squared-change parsimony reconstructions of ancestral states for continuous-valued characters on a phylogenetic tree. Syst. Biol. 1991, 40, 304–314. [Google Scholar] [CrossRef]
- Kulemeyer, C.; Asbahr, K.; Gunz, P.; Frahnert, S.; Bairlein, F. Functional morphology and integration of corvid skulls–a 3D geometric morphometric approach. Front. Zool. 2009, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Cubo, J.; Casinos, A. Scaling of skeletal element mass in birds. Belg. J. Zool. 1994, 124, 127–137. [Google Scholar]
- Baier, D.B.; Gatesy, S.M.; Jenkins, F.A. A critical ligamentous mechanism in the evolution of avian flight. Nature 2007, 445, 307–310. [Google Scholar] [CrossRef]
- Tobalske, B.W.; Warrick, D.R.; Jackson, B.E.; Dial, K.P. Morphological and Behavioral Correlates of Flapping Flight. In Living Dinosaurs: The Evolutionary History of Modern Birds; Dyke, G., Kaiser, G., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2011; pp. 257–281. [Google Scholar]
- Serrano, F.J.; Chiappe, L.M. Aerodynamic modelling of a Cretaceous bird reveals thermal soaring capabilities during early avian evolution. J. R. Soc. Interface 2017, 14, 20170182. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, M.; Goslow, G.E. Analysis of vertebrate structure, 5th ed.; John Wiley & Sons: New York, NY, USA, 2001; p. 656. [Google Scholar]
- Alexander, R.M. Principles of Animal Locomotion; Princeton University Press: Princeton, NJ, USA, 2003; p. 384. [Google Scholar]
- Chai, P.; Millard, D. Flight and size constraints: Hovering performance of large hummingbirds under maximal loading. J. Exp. Biol. 1997, 200, 2757–2763. [Google Scholar]
- Videler, J.J. Avian Flight. In Oxford Ornithology Series; Birkhead, T.R., Ed.; Oxford University Press: Oxford, UK, 2006; p. 288. [Google Scholar]
- Karkhu, A.A. Morphological divergence within the order Apodiformes as revealed by the structure of the humerus. Natur. Hist. Mus. Los Angel. Co. Sci. Ser. 1992, 36, 379–384. [Google Scholar]
- Savile, D.B.O. Adaptive evolution in the avian wing. Evolution 1957, 11, 212–224. [Google Scholar] [CrossRef]
- Rayner, J.M. Form and function in avian flight. In Current Ornithology; Springer: Berlin/Heidelberg, Germany, 1988; pp. 1–66. [Google Scholar]
- Turner, A. Swallows and Martins (Hirundinidae). In Handbook of the Birds of the World Alive; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Lynx Edicions: Barcelona, Spain, 2020; Available online: https://www.hbw.com/node/52303 (accessed on 28 February 2020).
- Johansson, L.C.; Aldrin, B.S.W. Kinematics of diving Atlantic puffins (Fratercula arctica L.): Evidence for an active upstroke. J. Exp. Biol. 2002, 205, 371–378. [Google Scholar]
- Mayr, G. Tertiary plotopterids (Aves, Plotopteridae) and a novel hypothesis on the phylogenetic relationships of penguins (Spheniscidae). J. Zool. Syst. 2005, 43, 67–71. [Google Scholar] [CrossRef]
- Raikow, R.J.; Bicanovsky, L.; Bledsoe, A.H. Forelimb joint mobility and the evolution of wing-propelled diving in birds. Auk 1988, 105, 446–451. [Google Scholar] [CrossRef]
- Louw, G.J. Functional anatomy of the penguin flipper. J. S. Afr. Vet. Assoc. 1992, 63, 113–120. [Google Scholar]
- Lovvorn, J.R.; Liggins, G.A. Interactions of body shape, body size and stroke-acceleration patterns in costs of underwater swimming by birds. Funct. Ecol. 2002, 16, 106–112. [Google Scholar] [CrossRef]
- Livezey, B.C. Morphometrics of flightlessness in the Alcidae. Auk 1988, 105, 681–698. [Google Scholar] [CrossRef]
- Smith, N.A. Taxonomic revision and phylogenetic analysis of the flightless Mancallinae (Aves, Pan-Alcidae). ZooKeys 2011, 91, 1–116. [Google Scholar] [CrossRef] [PubMed]
- Elliott, K.H.; Ricklefs, R.E.; Gaston, A.J.; Hatch, S.A.; Speakman, J.R.; Davoren, G.K. High flight costs, but low dive costs, in auks support the biomechanical hypothesis for flightlessness in penguins. Proc. Natl. Acad. Sci. USA 2013, 110, 9380–9384. [Google Scholar] [CrossRef] [PubMed]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Hartmann, K.; Mooers, A.O. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.J.; Wood, J.R.; Scofield, R.P.; Llamas, B.; Cooper, A. Ancient mitochondrial genome reveals unsuspected taxonomic affinity of the extinct Chatham duck (Pachyanas chathamica) and resolves divergence times for New Zealand and sub-Antarctic brown teals. Mol. Phylogenet. Evol. 2014, 70, 420–428. [Google Scholar] [CrossRef]
- Livezey, B.C. Flightlessness in the Galápagos cormorant (Compsohalieus [Nannopterum] harrisi): Heterochrony, giantism and specialization. Zool. J. Linn. Soc. 1992, 105, 155–224. [Google Scholar] [CrossRef]
- Wilson, R.P.; Hustler, K.; Ryan, P.G.; Burger, A.E.; Noldeke, E.C. Diving birds in cold water: Do Archimedes and Boyle determine energetic costs? Am. Nat. 1992, 140, 179–200. [Google Scholar] [CrossRef]
- Halsey, L.G.; Blackburn, T.M.; Butler, P.J. A comparative analysis of the diving behaviour of birds and mammals. Funct. Ecol. 2006, 20, 889–899. [Google Scholar] [CrossRef]
- Wilson, R.P.; Vargas, F.H.; Steinfurth, A.; Riordan, P.; Ropert-Coudert, Y.; Macdonald, D.W. What grounds some birds for life? Movement and diving in the sexually dimorphic Galapagos cormorant. Ecol. Monogr. 2008, 78, 633–652. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Linear Model | adj R2 | p-value | slope |
| CS | all birds | 0.811 | <0.001 | 0.423 |
| flighted | 0.907 | <0.001 | 0.484 | |
| flightless | 0.329 | 0.03 | 0.138 | |
| HL | all birds | 0.824 | <0.001 | 0.423 |
| flighted | 0.915 | <0.001 | 0.482 | |
| flightless | 0.340 | 0.027 | 0.145 | |
| HW | all birds | 0.906 | <0.001 | 0.365 |
| flighted | 0.937 | <0.001 | 0.379 | |
| flightless | 0.436 | 0.012 | 0.291 | |
| Variable | PGLS | adj R2 | p-value | |
| CS | Mb | 0.370 | <0.001 | |
| flighted-flightless | 0.099 | <0.001 | ||
| Mb: flighted-flightless | 0.259 | <0.001 | ||
| HL | Mb | 0.387 | <0.001 | |
| flighted-flightless | 0.092 | <0.001 | ||
| Mb: flighted-flightless | 0.260 | <0.001 | ||
| HW | Mb | 0.495 | <0.001 | |
| flighted-flightless | 0.016 | 0.014 | ||
| Mb: flighted-flightless | 0.148 | <0.001 |
| Linear Model | adj R2 | p-value |
| CS | 0.256 | <0.001 |
| Mb | 0.121 | <0.001 |
| PGLS | adj R2 | p-value |
| CS | 0.058 | <0.001 |
| flighted-flightless | 0.056 | <0.001 |
| CS: flighted-flightless | 0.012 | 0.108 |
| Mb | 0.022 | 0.016 |
| flighted-flightless | 0.057 | <0.001 |
| Mb: flighted-flightless | 0.014 | 0.074 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serrano, F.J.; Costa-Pérez, M.; Navalón, G.; Martín-Serra, A. Morphological Disparity of the Humerus in Modern Birds. Diversity 2020, 12, 173. https://doi.org/10.3390/d12050173
Serrano FJ, Costa-Pérez M, Navalón G, Martín-Serra A. Morphological Disparity of the Humerus in Modern Birds. Diversity. 2020; 12(5):173. https://doi.org/10.3390/d12050173
Chicago/Turabian StyleSerrano, Francisco J., Mireia Costa-Pérez, Guillermo Navalón, and Alberto Martín-Serra. 2020. "Morphological Disparity of the Humerus in Modern Birds" Diversity 12, no. 5: 173. https://doi.org/10.3390/d12050173
APA StyleSerrano, F. J., Costa-Pérez, M., Navalón, G., & Martín-Serra, A. (2020). Morphological Disparity of the Humerus in Modern Birds. Diversity, 12(5), 173. https://doi.org/10.3390/d12050173