Roles of Arbuscular Mycorrhizal Fungi on Plant Growth and Performance: Importance in Biotic and Abiotic Stressed Regulation

 ,
,  ,
,
Abstract

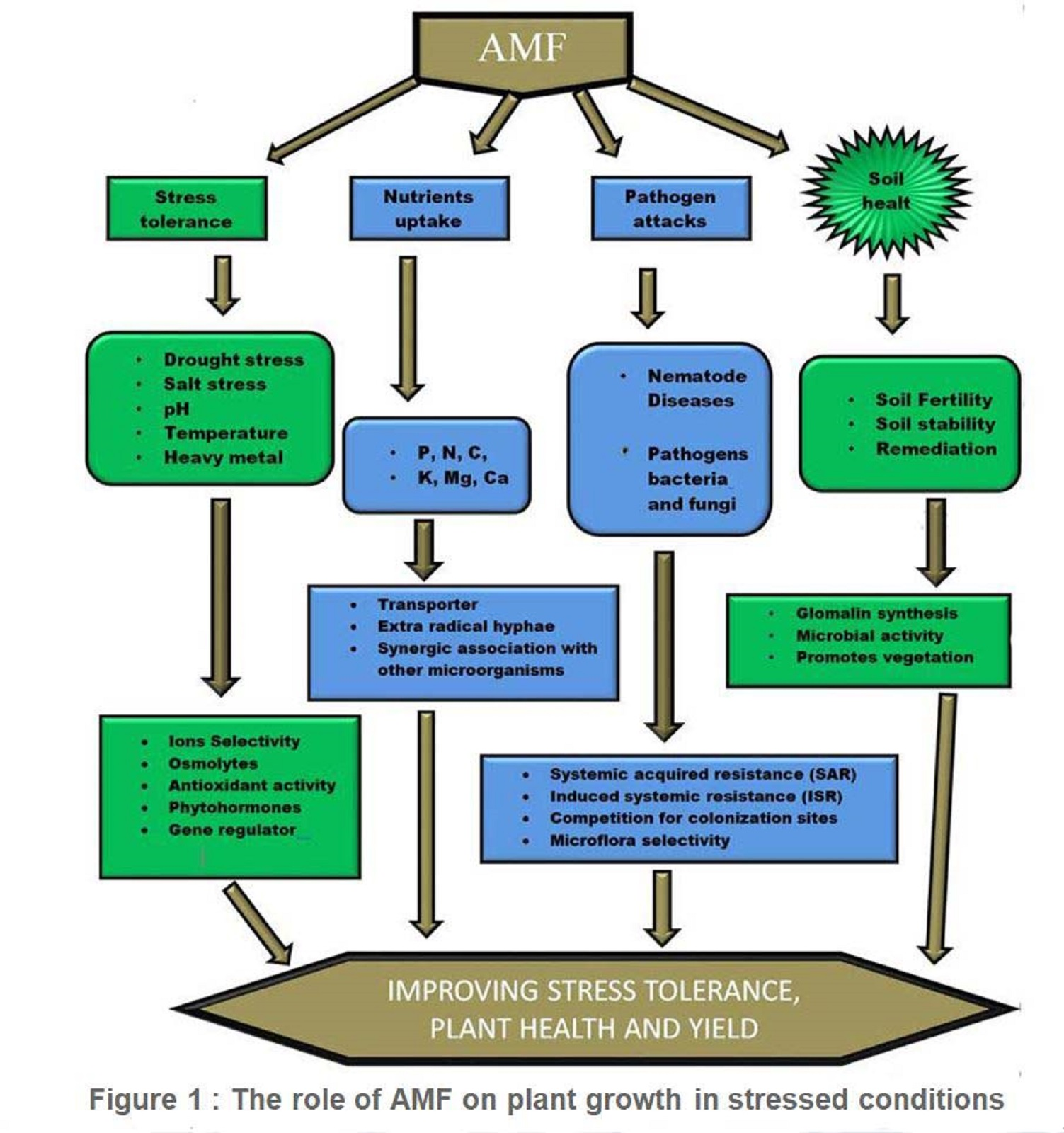
1. Introduction
2. Contribution of Arbuscular Mycorrhizal Fungi to Plant Nutrition and Growth
3. Role of Arbuscular Mycorrhizal Fungi in Alleviation of Abiotic Stresses in Plants
3.1. AMF and Plant Drought Tolerance

{kind=link}
| Host Plants | AMF Strains | Responses Related to AMF Inoculation | References |
|---|---|---|---|
| Zea mays L. Solanum lycopersicum L. | Rhizophagus irregularis (Błaszk., Wubet, Renker & Buscot) C. Walker & A. Schüßler 2010 | Enhanced apoplastic water flow | [69] |
| S. lycopersicum L. | F. mosseae R. irregularis | Increased plant height and biomass, intrinsic water use efficiency (iWUE) index, stomatal density, capacity to absorb CO2 and proline concentrations, and reduced hydrogen peroxide, leaf and root ABA contents | [71] |
| Lycopersicon. esculentum L. | Glomus clarum T.H. Nicolson & N.C. Schenck 1979 | Improved leaf area, dry mass, stomatal conductance, photosynthetic activity, and root hydraulic conductivity | [72] |
| L. esculatum L. | R. irregularis | Increased plant height, number of primary branches, flowers, and fruits, shoot and root dry matter, N and P contents, fruit yields, leaf relative water content (RWC), water use efficiency (WUE) and quality of fruits (less acidity and quantities of ascorbic acid and total soluble solids) | [73] |
| S. lycopersicum Lactuca sativa Linn. | R. irregularis | Improved shoot dry weight, stomatal conductance, photosystem II efficiency, ABA and strigolactone contents | [74] |
| Triticum aestivum L. | R. fasciculatus F. mosseae | Enhanced stomatal conductance and leaf osmotic adjustment | [75] |
| Lavandula spica L. | R. irregularis F. mossea | Increased biomass, N, K and water contents, and reduced antioxidant compounds (glutathione, ascorbate and H2O2) | [76] |
| Allium cepa L. | Glomus etunicatus W.N. Becker & Gerd. 1977 | Improved fresh and dry weights and phosphorus nutrition | [77] |
| Trifolium repens L. | R. irregularis | Enhanced dry weight, nutrients content (P, K, Ca, Mg, Zn and B), relative water content, proline concentrations, and glutathione reductase activity | [78] |
| Pistacia vera L. | G. etunicatum | Increased shoot and root weights, leaf area, total chlorophyll, and flavonoids contents, nutrient concentrations (P, N, K, Ca, Fe, Zn, and Cu), soluble sugar, proline, and soluble proteins contents, CAT and POD activities | [79] |
| L. esculatum L. Capsicum annuum L. | Rhizophagus irregularis Rhizophagus fasciculatus (Thaxt.) C. Walker & A. Schüßler, 2010 | Improved biomass, root length, shoot length, and chlorophyll contents, and reduced proline concentration | [86] |
3.2. AMF and Plant Flooding Tolerance
3.3. AMF and Plant Tolerance to Extreme Temperatures
3.4. AMF and Plant Tolerance to Salinity
3.5. AMF and Plant Tolerance to Heavy Metals
4. Role of AM Fungi in Alleviation of Biotic Stresses in Plants
| Host Plants | Disease or Pathogen | AMF Strains | Responses Related to AMF Inoculation | References |
|---|---|---|---|---|
| Solanum lycopersicum L. | M. incognita | F. mosseae | Induced systemic resistance against both the sedentary nematode Meloidogyne incognita and the migratory nematode Pratylenchus penetrans | [144] |
| L. esculentum | Fusarium oxysporum f. sp. lycopersici | Glomus sp. | Production of antimicrobial compounds from the mycorrhizal root that arrested the mycelial growth of the fungal pathogen Reduced the disease incidence Increased the plant growth, dry weight, N, P, K content, chlorophyll content and yield of the plant | [145] |
| Glycine max (L.) Merr. | Macrophomina phaseolina (Tassi) Goid 1947 | R. irregularis | Improve plant height and the number of functional leaves | [147] |
| Solanum lycopersicum L. | M. javanica | F. mosseae | Caused also a reduction in galling, nematode reproduction and morphometric parameters of females in tomato plants inoculated with | [151] |
| Alternaria solani Sorauer 1896 Fusarium oxysporum | F. mosseae | Alleviated tomato diseases | [155,156] | |
| Saccharum officinarum L. | Striga hermonthica Del Benth 1836 | G. etunicatum, Scutellospora fulgida Koske & C. Walker 1986, G. margarita | Stimulated plant growth, plant biomass and physiological parameters of plants in the presence of Striga | [165] |
| Solanum lycopersicum L. | Cladosporium fulvum Cooke 1883 | F. mosseae | Higher resistance against subsequent pathogen infection higher fresh and dry weight increases in total chlorophyll contents and net photosynthesis rate | [166] |
| Astragalus adsurgens var. Shanxi Yulin | Erysiphe pisi DC 1805 | C. etunicatum, G. versiforme, F. mosseae | Increased the shoot and root growth of standing milkvetch even though their presence in the roots increased susceptibility to powdery mildew. | [167] |
| Capsicum annum | Pythium aphanidermatum (Edson) Fitzp 1923 | Glomus sp. | Mycelial growth of the fungal pathogen reduced the disease incidence and increased the growth and yield of crop plants | [168] |
| Cicer arietinum L. | Fusarium wilt | Glomus hoi S.M. Berch & Trappe 1985, R. fasciculatum | Increased total contents of P and N in treated plants | [169] |
| Cucumis melo L. | Fusarium wilt | F. mosseae | Greatest capacity for reduction of disease incidence | [170] |
| Arachis hypogaea L. | Sclerotium rolfsii Sacc 1911 | R. fasciculatum, Gigaspora margarita, Cucumis melo L., A. laevis, and Sclerocystis dussii (Pat.) Höhn. 1910 | Eliminated the damaging effects of S. rolfsii | [171] |
| Solanum melongena L. Cucumis sativus L. | Verticillium dahliae Kleb 1913 Pseudomonas lacrymans (Smith and Bryan) Carsner 1970 | G. versiforme | Alleviated wilt symptoms caused by V. dahliae | [172] |
| S. tuberosum | Potato virus Y (PVY) | R. irregularis | Milder symptoms and significant stimulation of shoot growth were observed in PVY-infected plants inoculated | [173] |
| Nicotiana tabacum L. | Tobacco mosaic virus (TMV) Cucumber green mottle mosaic virus (CGMMV) | R. irregularis | Showed reduced disease symptoms and virus titer if compared to non-mycorrhizal plants | [174] |
| Zea mays L. | Striga hermonthica Del Benth 1836 | G. etunicatum, Scutellospora fulgida Koske & C. Walker 1986, G. margarita | Reduced Striga plant incidence, plant biomass, and phosphate content | [175] |
| Sorghum bicolor (L.) Moench | S. hermonthica | F. mosseae | Improved the performance of sorghum | [176] |
5. Interaction between AMF and Other Beneficial Soil Microorganisms
5.1. Interaction between AMF and Nitrogen Fixing Bacteria
5.1.1. Interaction between AMF and Rhizobia
5.1.2. Interaction between AMF and Frankia
5.2. Interaction between AMF and Plant Growth Promoting Rhizobacteria
5.3. Benefits of the Tripartite Symbiosis (AMF, Nitrogen Fixing Bacteria, PGPR, or Ectomycorrhizal Fungi)
5.4. Interaction between AMF and Mycorrhization Helper Bacteria
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2010; ISBN 978-0-08-055934-6. [Google Scholar]
- Kivlin, S.N.; Hawkes, C.V.; Treseder, K.K. Global diversity and distribution of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2011, 43, 2294–2303. [Google Scholar] [CrossRef]
- Wang, B.; Qiu, Y.-L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.S.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef]
- Schüβler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: Phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.V.; Gryganskyi, A.; et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, Z.A.; Pichtel, J. Mycorrhizae: An Overview. In Mycorrhizae: Sustainable Agriculture and Forestry; Springer Science and Business Media LLC: Berlin, Germany, 2008; pp. 1–35. [Google Scholar]
- Johns, C.D. Agricultural Application of Mycorrhizal Fungi to Increase Crop Yields, Promote Soil Health and Combat Climate Change. Future Directions International. 2014. Available online: https://www.futuredirections.org.au/publication/agricultural-application-of-mycorrhizal-fungi-to-increase-crop-yields-promote-soil-health-and-combat-climate-change/ (accessed on 11 August 2020).
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Genet. 2008, 6, 763–775. [Google Scholar] [CrossRef]
- Nakmee, P.S.; Techapinyawat, S.; Ngamprasit, S. Comparative potentials of native arbuscular mycorrhizal fungi to improve nutrient uptake and biomass of Sorghum bicolor Linn. Agric. Nat. Resour. 2016, 50, 173–178. [Google Scholar] [CrossRef]
- Posta, K.; Duc, N.H. Benefits of Arbuscular Mycorrhizal Fungi Application to Crop Production under Water Scarcity. Drought Detect. Solut. 2020. Available online: https://www.intechopen.com/books/drought-detection-and-solutions/benefits-of-arbuscular-mycorrhizal-fungi-application-to-crop-production-under-water-scarcity (accessed on 11 August 2020). [CrossRef]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads improve yield, quality and nutritional value of tomato: A field study. Mycorrhiza 2016, 27, 1–11. [Google Scholar] [CrossRef]
- Gamalero, E.; Trotta, A.; Massa, N.; Copetta, A.; Martinotti, M.G.; Berta, G. Impact of two fluorescent pseudomonads and an arbuscular mycorrhizal fungus on tomato plant growth, root architecture and P acquisition. Mycorrhiza 2003, 14, 185–192. [Google Scholar] [CrossRef]
- Kim, S.J.; Eo, J.-K.; Lee, E.-H.; Park, H.; Eom, A.-H. Effects of Arbuscular Mycorrhizal Fungi and Soil Conditions on Crop Plant Growth. Mycobiology 2017, 45, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Parihar, P.; Bora, M. Effect of mycorrhiza (Glomus mosseae) on morphological and biochemical properties of Ashwagandha (Withania somnifera) (L.) Dunal. J. Appl. Nat. Sci. 2018, 10, 1115–1123. [Google Scholar] [CrossRef]
- Al-Hmoud, G.; Al-Momany, A. Effect of Four Mycorrhizal Products on Squash Plant Growth and its Effect on Physiological Plant Elements. Adv. Crop. Sci. Technol. 2017, 5, 1–6. [Google Scholar] [CrossRef]
- Gogoi, P. Differential effect of some arbuscular mycorrhizal fungi on growth of Piper longum L. (Piperaceae). Indian J. Sci. Technol. 2011, 4, 119–125. [Google Scholar] [CrossRef]
- Ibijbijen, J.; Urquiaga, S.; Ismaili, M.; Alves, B.J.R.; Boddey, R.M. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition and nitrogen fixation of three varieties of common beans (Phaseolus vulgaris). New Phytol. 1996, 134, 353–360. [Google Scholar] [CrossRef]
- Miller, S.P.; Sharitz, R.R. Manipulation of flooding and arbuscular mycorrhiza formation influences growth and nutrition of two semiaquatic grass species. Funct. Ecol. 2000, 14, 738–748. [Google Scholar] [CrossRef]
- Ban, D.; Ban, S.G.; Oplanić, M.; Horvat, J.; Novak, B.; Žanić, K.; Žnidarčič, D. Growth and Yield Response of Watermelon to in-row Plant Spacings and Mycorrhiza. Chil. J. Agric. Res. 2011, 71, 497–502. [Google Scholar] [CrossRef][Green Version]
- Rodríguez-Romero, A.S.; Guerra, M.S.P.; Jaizme-Vega, M.D.C. Effect of arbuscular mycorrhizal fungi and rhizobacteria on banana growth and nutrition. Agron. Sustain. Dev. 2005, 25, 395–399. [Google Scholar] [CrossRef][Green Version]
- Berta, G.; Trotta, A.; Fusconi, A.; Hooker, J.E.; Munro, M.; Atkinson, D.; Giovannetti, M.; Morini, S.; Fortuna, P.; Tisserant, B.; et al. Arbuscular mycorrhizal induced changes to plant growth and root system morphology in Prunus cerasifera. Tree Physiol. 1995, 15, 281–293. [Google Scholar] [CrossRef]
- Jansa, J.; Forczek, S.T.; Rozmoš, M.; Püschel, D.; Bukovská, P.; Hršelová, H. Arbuscular mycorrhiza and soil organic nitrogen: Network of players and interactions. Chem. Biol. Technol. Agric. 2019, 6, 10. [Google Scholar] [CrossRef]
- Song, Z.; Bi, Y.; Zhang, J.; Gong, Y.; Yang, H. Arbuscular mycorrhizal fungi promote the growth of plants in the mining associated clay. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.A.; Smith, F.A. Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth: New Paradigms from Cellular to Ecosystem Scales. Annu. Rev. Plant. Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.F. Linking water and nutrients through the vadose zone: A fungal interface between the soil and plant systems. J. Arid. Land 2011, 3, 155–163. [Google Scholar] [CrossRef]
- Allen, M.F. Influence of vesicular-arbuscular mycorrhizae on water movement through Bouteloua gracilis (H.B.K.) lag ex steud. New Phytol. 1982, 91, 191–196. [Google Scholar] [CrossRef]
- Faber, B.A.; Zasoski, R.J.; Munns, D.N.; Shackel, K. A method for measuring hyphal nutrient and water uptake in mycorrhizal plants. Can. J. Bot. 1991, 69, 87–94. [Google Scholar] [CrossRef]
- Ruth, B.; Khalvati, M.; Schmidhalter, U. Quantification of mycorrhizal water uptake via high-resolution on-line water content sensors. Plant. Soil 2011, 342, 459–468. [Google Scholar] [CrossRef]
- Kaya, C.; Higgs, D.; Kirnak, H.; Taş, I. Mycorrhizal colonisation improves fruit yield and water use efficiency in watermelon (Citrullus lanatus Thunb.) grown under well-watered and water-stressed conditions. Plant. Soil 2003, 253, 287–292. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Li, G.-H.; Zou, Y.N. Roles of arbuscular mycorrhizal fungi on growth and nutrient acquisition of peach (Prunus persica L. Batsch) seedlings. J. Anim. Plant. Sci. 2011, 21, 746–750. [Google Scholar]
- Trouvelot, S.; Bonneau, L.; Redecker, D.; Van Tuinen, D.; Adrian, M.; Wipf, D. Arbuscular mycorrhiza symbiosis in viticulture: A review. Agron. Sustain. Dev. 2015, 35, 1449–1467. [Google Scholar] [CrossRef]
- Ravnskov, S.; Jakobsen, I. Functional compatibility in arbuscular mycorrhizas measured as hyphal P transport to the plant. New Phytol. 1995, 129, 611–618. [Google Scholar] [CrossRef]
- Farmer, M.; Li, X.; Feng, G.; Zhao, B.; Chatagnier, O.; Gianinazzi, S.; Gianinazzi-Pearson, V.; Van Tuinen, D. Molecular monitoring of field-inoculated AMF to evaluate persistence in sweet potato crops in China. Appl. Soil Ecol. 2007, 35, 599–609. [Google Scholar] [CrossRef]
- Jansa, J.; Smith, F.A.; Smith, S.E. Are there benefits of simultaneous root colonization by different arbuscular mycorrhizal fungi? New Phytol. 2008, 177, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Zangaro, W.; Nisizaki, S.M.A.; Domingos, J.C.B.; Nakano, E.M. Mycorrhizal response and successional status in 80 woody species from south Brazil. J. Trop. Ecol. 2003, 19, 315–324. [Google Scholar] [CrossRef]
- Vandresen, J.; Nishidate, F.R.; Torezan, J.M.D.; Zangara, W. Inoculação de fungos micorrízicos arbusculares e adubação na formação e pós-transplante de mudas de cinco espécies arbóreas nativas do sul do Brasil. Acta Bot. Bras. 2007, 21, 753–765. [Google Scholar] [CrossRef]
- Tahat, M.; Kamaruzaman, S.; Radziah, O.; Kadir, J.; Masdek, H. Plant Host Selectivity for Multiplication of Glomus mosseae Spore. Int. J. Bot. 2008, 4, 466–470. [Google Scholar] [CrossRef][Green Version]
- Golubkina, N.A.; Krivenkov, L.; Sękara, A.; Vasileva, V.; Tallarita, A.; Caruso, G. Prospects of Arbuscular Mycorrhizal Fungi Utilization in Production of Allium Plants. Plants 2020, 9, 279. [Google Scholar] [CrossRef]
- Coccina, A.; Cavagnaro, T.R.; Pellegrino, E.E.; Ercoli, L.; McLaughlin, M.; Watts-Williams, S.J. The mycorrhizal pathway of zinc uptake contributes to zinc accumulation in barley and wheat grain. BMC Plant. Biol. 2019, 19, 133. [Google Scholar] [CrossRef] [PubMed]
- Conversa, G.; Lazzizera, C.; Chiaravalle, A.E.; Miedico, O.; Bonasia, A.; La Rotonda, P.; Elia, A. Selenium fern application and arbuscular mycorrhizal fungi soil inoculation enhance Se content and antioxidant properties of green asparagus (Asparagus officinalis L.) spears. Sci. Hortic. 2019, 252, 176–191. [Google Scholar] [CrossRef]
- Luo, W.; Li, J.; Ma, X.; Niu, H.; Hou, S.; Wu, F. Effect of arbuscular mycorrhizal fungi on uptake of selenate, selenite, and selenomethionine by roots of winter wheat. Plant. Soil 2019, 438, 71–83. [Google Scholar] [CrossRef]
- Watts-Williams, S.J.; Gilbert, S.E. Arbuscular mycorrhizal fungi affect the concentration and distribution of nutrients in the grain differently in barley compared with wheat. Plants People Planet 2020. [Google Scholar] [CrossRef]
- Pellegrino, E.; Bedini, S. Enhancing ecosystem services in sustainable agriculture: Biofertilization and biofortification of chickpea (Cicer arietinum L.) by arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2014, 68, 429–439. [Google Scholar] [CrossRef]
- Lehmann, A.; Veresoglou, S.D.; Leifheit, E.F.; Rillig, M.C. Arbuscular mycorrhizal influence on zinc nutrition in crop plants—A meta-analysis. Soil Biol. Biochem. 2014, 69, 123–131. [Google Scholar] [CrossRef]
- Burleigh, S.H.; Bechmann, I.E. Plant nutrient transporter regulation in arbuscular mycorrhizas. Plant. Soil 2002, 244, 247–251. [Google Scholar] [CrossRef]
- Allen, J.W.; Shachar-Hill, Y. Sulfur Transfer through an Arbuscular Mycorrhiza. Plant. Physiol. 2008, 149, 549–560. [Google Scholar] [CrossRef]
- Sieh, D.; Watanabe, M.; Devers, E.A.; Brueckner, F.; Hoefgen, R.; Krajinski, F. The arbuscular mycorrhizal symbiosis influences sulfur starvation responses of Medicago truncatula. New Phytol. 2012, 197, 606–616. [Google Scholar] [CrossRef]
- Giovannetti, M.; Tolosano, M.; Volpe, V.; Kopriva, S.; Bonfante, P. Identification and functional characterization of a sulfate transporter induced by both sulfur starvation and mycorrhiza formation in Lotus japonicus. New Phytol. 2014, 204, 609–619. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Cavagnaro, T.R.; Watts-Williams, S.J. The effects of soil phosphorus and zinc availability on plant responses to mycorrhizal fungi: A physiological and molecular assessment. Sci. Rep. 2019, 9, 14880. [Google Scholar] [CrossRef]
- Ma, J.; Janouskova, M.; Ye, L.; Bai, L.; Dong, R.; Yan, Y.; Yu, X.; Zou, Z.; Li, Y.; He, C. Role of arbuscular mycorrhiza in alleviating the effect of cold on the photosynthesis of cucumber seedlings. Photosynthetica 2019, 57, 86–95. [Google Scholar] [CrossRef]
- Ma, X.; Luo, W.; Li, J.; Wu, F. Arbuscular mycorrhizal fungi increase both concentrations and bioavilability of Zn in wheat (Triticum aestivum L) grain on Zn-spiked soils. Appl. Soil Ecol. 2019, 135, 91–97. [Google Scholar] [CrossRef]
- Gorzelak, M.A.; Asay, A.K.; Pickles, B.; Simard, S.W. Inter-Plant communication through mycorrhizal networks mediates complex adaptive behaviour in plant communities. Aob Plants 2015, 7, 7. [Google Scholar] [CrossRef]
- Ingraffia, R.; Amato, G.; Frenda, A.S.; Giambalvo, D. Impacts of arbuscular mycorrhizal fungi on nutrient uptake, N2 fixation, N transfer, and growth in a wheat/faba bean intercropping system. PLoS ONE 2019, 14, e0213672. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, I.; Fontaine, J.; Sahraoui, A.L.-H. Arbuscular mycorrhizal fungal responses to abiotic stresses: A review. Phytochemistry 2016, 123, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Del Val, C.; Barea, J.M.; Azcón-Aguilar, C. Assessing the tolerance to heavy metals of arbuscular mycorrhizal fungi isolated from sewage sludge-contaminated soils. Appl. Soil Ecol. 1999, 11, 261–269. [Google Scholar] [CrossRef]
- Del Val, C.; Barea, J.M.; Azcón-Aguilar, C. Diversity of Arbuscular Mycorrhizal Fungus Populations in Heavy-Metal-Contaminated Soils. Appl. Environ. Microbiol. 1999, 65, 718–723. [Google Scholar] [CrossRef]
- Weissenhorn, I.; Glashoff, A.; Leyval, C.; Berthelin, J. Differential tolerance to Cd and Zn of arbuscular mycorrhizal (AM) fungal spores isolated from heavy metal-polluted and unpolluted soils. Plant. Soil 1994, 167, 189–196. [Google Scholar] [CrossRef]
- Millar, N.S.; Bennett, A.E. Stressed out symbiotes: Hypotheses for the influence of abiotic stress on arbuscular mycorrhizal fungi. Oecologia 2016, 182, 625–641. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A.; Klironomos, J.N.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I.R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Bennett, A.E.; Classen, A. Climate change influences mycorrhizal fungal–plant interactions, but conclusions are limited by geographical study bias. Ecology 2020, 101. [Google Scholar] [CrossRef]
- Kapoor, R.; Evelin, H.; Mathur, P.; Giri, B. Arbuscular Mycorrhiza: Approaches for Abiotic Stress Tolerance in Crop Plants for Sustainable Agriculture. In Plant Acclimation to Environmental Stress; Tuteja, N., Singh Gill, S., Eds.; Springer: New York, NY, USA, 2013; pp. 359–401. ISBN 978-1-4614-5001-6. [Google Scholar]
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J.M. Salinity stress alleviation using arbuscular mycorrhizal fungi. A review. Agron. Sustain. Dev. 2011, 32, 181–200. [Google Scholar] [CrossRef]
- Hoeksema, J.D.; Chaudhary, V.B.; Gehring, C.A.; Johnson, N.C.; Karst, J.; Koide, R.T.; Pringle, A.; Zabinski, C.; Bever, J.D.; Moore, J.C.; et al. A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. Ecol. Lett. 2010, 13, 394–407. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.-S.P. Response of plants to water stress. Front. Plant. Sci. 2014, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Balestrini, R.; Lumini, E. Focus on mycorrhizal symbioses. Appl. Soil Ecol. 2018, 123, 299–304. [Google Scholar] [CrossRef]
- Bernardo, L.; Carletti, P.; Badeck, F.; Rizza, F.; Morcia, C.; Ghizzoni, R.; Rouphael, Y.; Colla, G.; Terzi, V.; Lucini, L. Metabolomic responses triggered by arbuscular mycorrhiza enhance tolerance to water stress in wheat cultivars. Plant. Physiol. Biochem. 2019, 137, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Augé, R.M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 2001, 11, 3–42. [Google Scholar] [CrossRef]
- Bárzana, G.; Aroca, R.; Paz, J.A.; Chaumont, F.; Martínez-Ballesta, M.C.; Carvajal, M.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal symbiosis increases relative apoplastic water flow in roots of the host plant under both well-watered and drought stress conditions. Ann. Bot. 2012, 109, 1009–1017. [Google Scholar] [CrossRef]
- Chitarra, W.; Maserti, B.; Gambino, G.; Guerrieri, E.; Balestrini, R. Arbuscular mycorrhizal symbiosis-mediated tomato tolerance to drought. Plant. Signal. Behav. 2016, 11, e1197468. [Google Scholar] [CrossRef]
- Chitarra, W.; Pagliarani, C.; Maserti, B.; Lumini, E.; Siciliano, I.; Cascone, P.; Schubert, A.; Gambino, G.; Balestrini, R.; Guerrieri, E. Insights on the Impact of Arbuscular Mycorrhizal Symbiosis on Tomato Tolerance to Water Stress1. Plant. Physiol. 2016, 171, 1009–1023. [Google Scholar] [CrossRef]
- Dell’Amico, J.; Torrecillas, A.; Rodríguez, P.; Morte, A.; Sánchez-Blanco, M.J. Responses of tomato plants associated with the arbuscular mycorrhizal fungus Glomus clarum during drought and recovery. J. Agric. Sci. 2002, 138, 387–393. [Google Scholar] [CrossRef]
- Subramanian, K.; Santhanakrishnan, P.; Balasubramanian, P. Responses of field grown tomato plants to arbuscular mycorrhizal fungal colonization under varying intensities of drought stress. Sci. Hortic. 2006, 107, 245–253. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Aroca, R.; Zamarreño, Á.M.; Molina, S.; Andreo-Jimenez, B.; Porcel, R.; García-Mina, J.M.; Ruyter-Spira, C.; López-Ráez, J.A. Arbuscular mycorrhizal symbiosis induces strigolactone biosynthesis under drought and improves drought tolerance in lettuce and tomato. PlantCell Environ. 2015, 39, 441–452. [Google Scholar] [CrossRef]
- Allen, M.F.; Boosalis, M.G. Effects of two species of VA mycorrhizal fungi on drought tolerance of winter wheat. New Phytol. 1983, 93, 67–76. [Google Scholar] [CrossRef]
- Marulanda, A.; Porcel, R.; Barea, J.M.; Azcón, R. Drought Tolerance and Antioxidant Activities in Lavender Plants Colonized by Native Drought-tolerant or Drought-sensitive Glomus Species. Microb. Ecol. 2007, 54, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Nelsen, C.E.; Safir, G.R. Increased drought tolerance of mycorrhizal onion plants caused by improved phosphorus nutrition. Planta 1982, 154, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, N.; Armada, E.; Duque, E.; Roldan, A.; Azcón, R. Contribution of arbuscular mycorrhizal fungi and/or bacteria to enhancing plant drought tolerance under natural soil conditions: Effectiveness of autochthonous or allochthonous strains. J. Plant. Physiol. 2015, 174, 87–96. [Google Scholar] [CrossRef]
- Abbaspour, H.; Saeidi-Sar, S.; Afshari, H.; Abdel-Wahhab, M.; Abdel-Wahhab, M.A. Tolerance of Mycorrhiza infected Pistachio (Pistacia vera L.) seedling to drought stress under glasshouse conditions. J. Plant. Physiol. 2012, 169, 704–709. [Google Scholar] [CrossRef]
- Osonubi, O.; Mulongoy, K.; Awotoye, O.O.; Atayese, M.O.; Okali, D.U.U. Effects of ectomycorrhizal and vesicular-arbuscular mycorrhizal fungi on drought tolerance of four leguminous woody seedlings. Plant. Soil 1991, 136, 131–143. [Google Scholar] [CrossRef]
- Ouledali, S.; Ennajeh, M.; Ferrandino, A.; Khemira, H.; Schubert, A.; Secchi, F. Influence of arbuscular mycorrhizal fungi inoculation on the control of stomata functioning by abscisic acid (ABA) in drought-stressed olive plants. S. Afr. J. Bot. 2019, 121, 152–158. [Google Scholar] [CrossRef]
- De Ollas, C.; Dodd, I.C. Physiological impacts of ABA-JA interactions under water-limitation. Plant. Mol. Biol. 2016, 91, 641–650. [Google Scholar] [CrossRef]
- Yosefi, M.; Sharafzadeh, S.; Bazrafshan, F.; Zare, M.; Amiri, B. Application of jasmonic acid can mitigate water deficit stress in cotton through yield-related physiological properties. Acta Agrobot. 2018, 71. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Li, W.; Nguyen, K.H.; Fujita, M.; Tran, L.-S.P. Strigolactones in plant adaptation to abiotic stresses: An emerging avenue of plant research. PlantCell Environ. 2018, 41, 2227–2243. [Google Scholar] [CrossRef] [PubMed]
- Padmavathi, T.; Dikshit, R.; Seshagiri, S. Influence of Rhizophagus spp. And Burkholderia seminalison the Growth of Tomato (Lycopersicon esculatum) and Bell Pepper (Capsicum annuum) under Drought Stress. Commun. Soil Sci. Plant. Anal. 2016, 47, 1975–1984. [Google Scholar] [CrossRef]
- Khanam, D. Influence of Flooding on the Survival of Arbuscular Mycorrhiza. Bangladesh J. Microbiol. 2008, 25, 111–114. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Y.; Qiu, Q.; Xin, G.; Yang, Z.; Shi, S. Flooding Greatly Affects the Diversity of Arbuscular Mycorrhizal Fungi Communities in the Roots of Wetland Plants. PLoS ONE 2011, 6, e24512. [Google Scholar] [CrossRef]
- Bao, X.; Wang, Y.; Li, S.; Olsson, P.A. Arbuscular mycorrhiza under water—Carbon‒phosphorus exchange between rice and arbuscular mycorrhizal fungi under different flooding regimes. Soil Biol. Biochem. 2019, 129, 169–177. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Fougnies, L.; Renciot, S.; Müller, F.; Plenchette, C.; Prin, Y.; De Faria, S.M.; Bouvet, J.M.; Sylla, S.N.; Dreyfus, B.; Bâ, A.M. Arbuscular mycorrhizal colonization and nodulation improve flooding tolerance in Pterocarpus officinalis Jacq. seedlings. Mycorrhiza 2006, 17, 159–166. [Google Scholar] [CrossRef]
- Wang, Y.; Qiu, Q.; Yang, Z.; Hu, Z.; Tam, N.; Xin, G. Arbuscular mycorrhizal fungi in two mangroves in South China. Plant. Soil 2009, 331, 181–191. [Google Scholar] [CrossRef]
- Solís-Rodríguez, U.R.J.; Ramos-Zapata, J.; Hernández-Cuevas, L.; Salinas-Peba, L.; Guadarrama, P. Arbuscular mycorrhizal fungi diversity and distribution in tropical low flooding forest in Mexico. Mycol. Prog. 2020, 19, 195–204. [Google Scholar] [CrossRef]
- Zhu, X.-C.; Song, F.-B.; Liu, S.-Q.; Liu, T.-D. Effects of arbuscular mycorrhizal fungus on photosynthesis and water status of maize under high temperature stress. Plant. Soil 2011, 346, 189–199. [Google Scholar] [CrossRef]
- Caradonia, F.; Francia, E.; Morcia, C.; Ghizzoni, R.; Moulin, L.; Terzi, V.; Ronga, D. Arbuscular Mycorrhizal Fungi and Plant Growth Promoting Rhizobacteria Avoid Processing Tomato Leaf Damage during Chilling Stress. Agronomy 2019, 9, 299. [Google Scholar] [CrossRef]
- Zhu, X.; Song, F.; Liu, F. Arbuscular Mycorrhizal Fungi and Tolerance of Temperature Stress in Plants. In Arbuscular Mycorrhizas and Stress Tolerance of Plants; Springer Science and Business Media LLC: Berlin, Germany, 2017; Volume 33, pp. 163–194. [Google Scholar]
- Latef, A.A.H.A.; Chaoxing, H. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci. Hortic. 2011, 127, 228–233. [Google Scholar] [CrossRef]
- Mathur, S.; Jajoo, A. Arbuscular mycorrhizal fungi protects maize plants from high temperature stress by regulating photosystem II heterogeneity. Ind. Crop. Prod. 2020, 143, 111934. [Google Scholar] [CrossRef]
- Hajiboland, R.; Joudmand, A.; Aliasgharzad, N.; Tolrá, R.; Poschenrieder, C. Arbuscular mycorrhizal fungi alleviate low-temperature stress and increase freezing resistance as a substitute for acclimation treatment in barley. Crop. Pasture Sci. 2019, 70, 218–233. [Google Scholar] [CrossRef]
- Bender, S.F.; Plantenga, F.; Neftel, A.; Jocher, M.; Oberholzer, H.-R.; Köhl, L.; Giles, M.; Daniell, T.J.; Van Der Heijden, M.G. Symbiotic relationships between soil fungi and plants reduce N2O emissions from soil. ISME J. 2013, 8, 1336–1345. [Google Scholar] [CrossRef]
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular Mycorrhizal Fungi as Natural Biofertilizers: Let’s Benefit from Past Successes. Front. Microbiol. 2016, 6. [Google Scholar] [CrossRef]
- Gemma, J.N.; Koske, R.E. Seasonal Variation in Spore Abundance and Dormancy of Gigaspora Gigantea and in Mycorrhizal Inoculum Potential of a Dune Soil. Mycology 1988, 80, 211–216. [Google Scholar] [CrossRef]
- Maya, M.A.; Matsubara, Y.-I. Influence of arbuscular mycorrhiza on the growth and antioxidative activity in cyclamen under heat stress. Mycorrhiza 2013, 23, 381–390. [Google Scholar] [CrossRef]
- Chu, X.; Fu, J.; Sun, Y.; Xu, Y.; Miao, Y.; Xu, Y.; Hu, T. Effect of arbuscular mycorrhizal fungi inoculation on cold stress-induced oxidative damage in leaves of Elymus nutans Griseb. S. Afr. J. Bot. 2016, 104, 21–29. [Google Scholar] [CrossRef]
- Yamato, M.; Ikeda, S.; Iwase, K. Community of arbuscular mycorrhizal fungi in a coastal vegetation on Okinawa island and effect of the isolated fungi on growth of sorghum under salt-treated conditions. Mycorrhiza 2008, 18, 241–249. [Google Scholar] [CrossRef]
- Beltrano, J.; Ruscitti, M.; Arango, M.; Ronco, M. Effects of arbuscular mycorrhiza inoculation on plant growth, biological and physiological parameters and mineral nutrition in pepper grown under different salinity and p levels. J. Soil Sci. Plant. Nutr. 2013, 13, 123–141. [Google Scholar] [CrossRef]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef]
- Amanifar, S.; Khodabandeloo, M.; Fard, E.M.; Askari, M.S.; Ashrafi, M. Alleviation of salt stress and changes in glycyrrhizin accumulation by arbuscular mycorrhiza in liquorice (Glycyrrhiza glabra) grown under salinity stress. Environ. Exp. Bot. 2019, 160, 25–34. [Google Scholar] [CrossRef]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Influence of arbuscular mycorrhizal fungi and salinity on growth, biomass, and mineral nutrition of Acacia auriculiformis. Biol. Fertil. Soils 2003, 38, 170–175. [Google Scholar] [CrossRef]
- Djighaly, P.I.; Diagne, N.; Ngom, M.; Ngom, D.; Hocher, V.; Fall, D.; Diouf, D.; Laplaze, L.; Svistoonoff, S.; Champion, A. Selection of arbuscular mycorrhizal fungal strains to improve Casuarina equisetifolia L. and Casuarina glauca Sieb. tolerance to salinity. Ann. Sci. 2018, 75, 72. [Google Scholar] [CrossRef]
- Hajiboland, R.; Aliasgharzad, N.; Laiegh, S.F.; Poschenrieder, C. Colonization with arbuscular mycorrhizal fungi improves salinity tolerance of tomato (Solanum lycopersicum L.) plants. Plant. Soil 2009, 331, 313–327. [Google Scholar] [CrossRef]
- Daei, G.; Ardekani, M.; Rejali, F.; Teimuri, S.; Miransari, M. Alleviation of salinity stress on wheat yield, yield components, and nutrient uptake using arbuscular mycorrhizal fungi under field conditions. J. Plant. Physiol. 2009, 166, 617–625. [Google Scholar] [CrossRef]
- Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Wirth, S.; Egamberdieva, D. The Interaction between Arbuscular Mycorrhizal Fungi and Endophytic Bacteria Enhances Plant Growth of Acacia gerrardii under Salt Stress. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Li, J.; Meng, B.; Chai, H.; Yang, X.; Song, W.; Li, S.; Lu, A.; Zhang, T.; Sun, W. Arbuscular Mycorrhizal Fungi Alleviate Drought Stress in C3 (Leymus chinensis) and C4 (Hemarthria altissima) Grasses via Altering Antioxidant Enzyme Activities and Photosynthesis. Front. Plant. Sci. 2019, 10, 10. [Google Scholar] [CrossRef]
- Wani, S.H.; Kumar, V.; Khare, T.; Guddimalli, R.; Parveda, M.; Solymosi, K.; Suprasanna, P.; Kishor, P.B.K. Engineering salinity tolerance in plants: Progress and prospects. Planta 2020, 251, 76. [Google Scholar] [CrossRef] [PubMed]
- Talaat, N.B.; Shawky, B. Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) plants exposed to salinity. Environ. Exp. Bot. 2014, 98, 20–31. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant. Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wu, N.; Meng, S.; Wu, F.; Liu, T. Arbuscular mycorrhizal fungi (AMF) enhance the tolerance of Euonymus maackii Rupr. at a moderate level of salinity. PLoS ONE 2020, 15, e0231497. [Google Scholar] [CrossRef]
- Estrada, B.; Aroca, R.; Maathuis, F.J.M.; Barea, J.M.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal fungi native from a Mediterranean saline area enhance maize tolerance to salinity through improved ion homeostasis. PlantCell Environ. 2013, 36, 1771–1782. [Google Scholar] [CrossRef]
- Pedranzani, H.; Rodríguez-Rivera, M.; Gutierrez, M.; Porcel, R.; Hause, B.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal symbiosis regulates physiology and performance of Digitaria eriantha plants subjected to abiotic stresses by modulating antioxidant and jasmonate levels. Mycorrhiza 2015, 26, 141–152. [Google Scholar] [CrossRef]
- Al-Karaki, G.N. Nursery inoculation of tomato with arbuscular mycorrhizal fungi and subsequent performance under irrigation with saline water. Sci. Hortic. 2006, 109, 1–7. [Google Scholar] [CrossRef]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Improved Tolerance of Acacia nilotica to Salt Stress by Arbuscular Mycorrhiza, Glomus fasciculatum may be Partly Related to Elevated K/Na Ratios in Root and Shoot Tissues. Microb. Ecol. 2007, 54, 753–760. [Google Scholar] [CrossRef]
- Soliman, A.S.; Shanan, N.T.; Massoud, O.N.; Swelim, D.M. Improving salinity tolerance of Acacia saligna (Labill.) plant by arbuscular mycorrhizal fungi and Rhizobium inoculation. Afr. J. Biotechnol. 2012, 11, 1259–1266. [Google Scholar] [CrossRef]
- Santander, C.; Sanhueza, M.; Olave, J.; Borie, F.; Valentine, A.; Cornejo, P. Arbuscular Mycorrhizal Colonization Promotes the Tolerance to Salt Stress in Lettuce Plants through an Efficient Modification of Ionic Balance. J. Soil Sci. Plant. Nutr. 2019, 19, 321–331. [Google Scholar] [CrossRef]
- Diouf, D.; Duponnois, R.; Ba, A.T.; Neyra, M.; Lesueur, D. Symbiosis of Acacia auriculiformis and Acacia mangium with mycorrhizal fungi and Bradyrhizobium spp. improves salt tolerance in greenhouse conditions. Funct. Plant. Biol. 2005, 32, 1143–1152. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Wang, M.; Li, Y.; Wu, A.; Huang, J. Effects of arbuscular mycorrhizal fungi on growth and nitrogen uptake of Chrysanthemum morifolium under salt stress. PLoS ONE 2018, 13, e0196408. [Google Scholar] [CrossRef]
- Tian, C.; Feng, G.; Li, X.; Zhang, F. Different effects of arbuscular mycorrhizal fungal isolates from saline or non-saline soil on salinity tolerance of plants. Appl. Soil Ecol. 2004, 26, 143–148. [Google Scholar] [CrossRef]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.-B.F.; Aldehaish, H.A.; Egamberdieva, D.; Allah, E.A. Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 2018, 25, 1102–1114. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.; Allah, E.A.; Alqarawi, A.A.; Wirth, S.; Egamberdieva, D. Comparing symbiotic performance and physiological responses of two soybean cultivars to arbuscular mycorrhizal fungi under salt stress. Saudi J. Biol. Sci. 2016, 26, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, U.; Regvar, M.; Bothe, H. Arbuscular mycorrhiza and heavy metal tolerance. Phytochemistry 2007, 68, 139–146. [Google Scholar] [CrossRef]
- Wang, F. Occurrence of arbuscular mycorrhizal fungi in mining-impacted sites and their contribution to ecological restoration: Mechanisms and applications. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1–57. [Google Scholar] [CrossRef]
- Kelkar, T.S.; Bhalerao, S.A. Beneficiary Effect of Arbuscular Mycorrhiza to Trigonella Foenum-Graceum in Contaminated Soil by Heavy Metal. Res. J. Recent Sci. 2013, 2, 29–32. [Google Scholar]
- Kaldorf, M.; Kuhn, A.; Schröder, W.; Hildebrandt, U.; Bothe, H. Selective Element Deposits in Maize Colonized by a Heavy Metal Tolerance Conferring Arbuscular Mycorrhizal Fungus. J. Plant. Physiol. 1999, 154, 718–728. [Google Scholar] [CrossRef]
- Gaur, A.; Adholeya, A. Prospects of arbuscular mycorrhizal fungi in phytoremediation of heavy metal contaminated soils. Curr. Sci. 2004, 86, 528–534. [Google Scholar]
- Gong, X.; Tian, D.Q. Study on the effect mechanism of Arbuscular Mycorrhiza on the absorption of heavy metal elements in soil by plants. IOP Conf. Ser. Earth Environ. Sci. 2019, 267, 052064. [Google Scholar] [CrossRef]
- Audet, P.; Charest, C. Heavy metal phytoremediation from a meta-analytical perspective. Environ. Pollut. 2007, 147, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, J.; Yue, F.; Yan, X.; Wang, F.; Bloszies, S.; Wang, Y. Effects of arbuscular mycorrhizal inoculation and biochar amendment on maize growth, cadmium uptake and soil cadmium speciation in Cd-contaminated soil. Chemosphere 2018, 194, 495–503. [Google Scholar] [CrossRef] [PubMed]
- González-Guerrero, M.; Escudero, V.; Saéz, Á.; Tejada-Jiménez, M. Transition Metal Transport in Plants and Associated Endosymbionts: Arbuscular Mycorrhizal Fungi and Rhizobia. Front. Plant. Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Tamayo, E.; Ferrol, N.; Gómez-Gallego, T.; Azcón-Aguilar, C. Genome-wide analysis of copper, iron and zinc transporters in the arbuscular mycorrhizal fungus Rhizophagus irregularis. Front. Plant. Sci. 2014, 5, 5. [Google Scholar] [CrossRef]
- Jiang, Q.-Y.; Zhuo, F.; Long, S.-H.; Zhao, H.-D.; Yang, D.-J.; Ye, Z.-H.; Li, S.-S.; Jing, Y.-X. Can arbuscular mycorrhizal fungi reduce Cd uptake and alleviate Cd toxicity of Lonicera japonica grown in Cd-added soils? Sci. Rep. 2016, 6, 21805. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.; Allah, E.A.; Alqarawi, A.A.; Al Huqail, A.A.; Egamberdieva, D.; Wirth, S. Alleviation of cadmium stress in Solanum lycopersicum L. by arbuscular mycorrhizal fungi via induction of acquired systemic tolerance. Saudi J. Biol. Sci. 2016, 23, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Lingua, G.; Franchin, C.; Todeschini, V.; Castiglione, S.; Biondi, S.; Burlando, B.; Parravicini, V.; Torrigiani, P.; Berta, G. Arbuscular mycorrhizal fungi differentially affect the response to high zinc concentrations of two registered poplar clones. Environ. Pollut. 2008, 153, 137–147. [Google Scholar] [CrossRef]
- Li, X.; Christie, P. Changes in soil solution Zn and pH and uptake of Zn by arbuscular mycorrhizal red clover in Zn-contaminated soil. Chemosphere 2001, 42, 201–207. [Google Scholar] [CrossRef]
- Vos, C.; Tesfahun, A.; Panis, B.; De Waele, D.; Elsen, A. Arbuscular mycorrhizal fungi induce systemic resistance in tomato against the sedentary nematode Meloidogyne incognita and the migratory nematode Pratylenchus penetrans. Appl. Soil Ecol. 2012, 61, 1–6. [Google Scholar] [CrossRef]
- Kumari, S.M.P.; Prabina, B.J. Protection of Tomato, Lycopersicon esculentum from Wilt Pathogen, Fusarium oxysporum f.sp. lycopersici by Arbuscular Mycorrhizal Fungi, Glomus sp. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 1368–1378. [Google Scholar] [CrossRef]
- Nguvo, K.J.; Gao, X. Weapons hidden underneath: Bio-Control agents and their potentials to activate plant induced systemic resistance in controlling crop Fusarium diseases. J. Plant. Dis. Prot. 2019, 126, 177–190. [Google Scholar] [CrossRef]
- Spagnoletti, F.N.; Cornero, M.; Chiocchio, V.; Lavado, R.S.; Roberts, I.N. Arbuscular mycorrhiza protects soybean plants against Macrophomina phaseolina even under nitrogen fertilization. Eur. J. Plant. Pathol. 2020, 156, 839–849. [Google Scholar] [CrossRef]
- Pozo, M.J.; Azcón-Aguilar, C. Unraveling mycorrhiza-induced resistance. Curr. Opin. Plant. Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef]
- Jung, R.E.; Zembic, A.; Pjetursson, B.E.; Zwahlen, M.; Thoma, D.S. Systematic review of the survival rate and the incidence of biological, technical, and aesthetic complications of single crowns on implants reported in longitudinal studies with a mean follow-up of 5 years. Clin. Oral Implant. Res. 2012, 23, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.D.; Neal, A.L.; Van Wees, S.A.; Ton, J. Mycorrhiza-induced resistance: More than the sum of its parts? Trends Plant. Sci. 2013, 18, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, Z.A.; Mahmood, I. Effect of a plant growth promoting bacterium, an AM fungus and soil types on the morphometrics and reproduction of Meloidogyne javanica on tomato. Appl. Soil Ecol. 1998, 8, 77–84. [Google Scholar] [CrossRef]
- Akhtar, M.S.; Siddiqui, Z.A. Glomus intraradices, Pseudomonas alcaligenes, and Bacillus pumilus: Effective agents for the control of root-rot disease complex of chickpea (Cicer arietinum L.). J. Gen. Plant. Pathol. 2007, 74, 53–60. [Google Scholar] [CrossRef]
- Hoffmann, D.; Vierheilig, H.; Schausberger, P. Arbuscular mycorrhiza enhances preference of ovipositing predatory mites for direct prey-related cues. Physiol. Èntomol. 2010, 36, 90–95. [Google Scholar] [CrossRef]
- Vos, C.; Claerhout, S.; Mkandawire, R.; Panis, B.; De Waele, D.; Elsen, A. Arbuscular mycorrhizal fungi reduce root-knot nematode penetration through altered root exudation of their host. Plant. Soil 2011, 354, 335–345. [Google Scholar] [CrossRef]
- Song, Y.; Chen, D.; Lu, K.; Sun, Z.; Zeng, R. Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front. Plant. Sci. 2015, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- El-Khallal, S.M. Induction and modulation of resistance in tomato plants against Fusarium wilt disease by bioagent fungi (arbuscular mycorrhiza) and/or hormonal elicitors (Jasmonic acid & Salicylic acid): 2-Changes in the antioxidant enzymes, phenolic compounds and pathogen related- proteins. Aust. J. Basic Appl. Sci. 2007, 1, 717–732. [Google Scholar]
- Akhtar, M.S.; Siddiqui, Z.A.; Wiemken, A. Arbuscular Mycorrhizal Fungi and Rhizobium to Control Plant Fungal Diseases. Altern. Farming Syst. Biotechnol. Drought Stress Ecol. Fertil. 2010, 263–292. [Google Scholar] [CrossRef]
- Gallou, A.; Mosquera, H.P.L.; Cranenbrouck, S.; Suárez, J.P.; Declerck, S. Mycorrhiza induced resistance in potato plantlets challenged by Phytophthora infestans. Physiol. Mol. Plant. Pathol. 2011, 76, 20–26. [Google Scholar] [CrossRef]
- Koricheva, J.; Gange, A.C.; Jones, T. Effects of mycorrhizal fungi on insect herbivores: A meta-analysis. Ecology 2009, 90, 2088–2097. [Google Scholar] [CrossRef] [PubMed]
- Gange, A.C.; Brown, V.K.; Aplin, D.M. Multitrophic links between arbuscular mycorrhizal fungi and insect parasitoids. Ecol. Lett. 2003, 6, 1051–1055. [Google Scholar] [CrossRef]
- Gange, A.C.; West, H.M. Interactions between arbuscular mycorrhizal fungi and foliar-feeding insects in Plantago lanceolata L. New Phytol. 1994, 128, 79–87. [Google Scholar] [CrossRef]
- Atera, E.A.; Itoh, K.; Azuma, T.; Ishii, T. Farmers’ perspectives on the biotic constraint of Striga hermonthica and its control in western Kenya. Weed Biol. Manag. 2012, 12, 53–62. [Google Scholar] [CrossRef]
- Jones, N.; Madhura, A.; Prashant, S.; Ramesh, B.; Jagadeesh, K.; Asha, A. Evaluation of arbuscular mycorrhizal fungi for suppression of Striga hermonthica, a parasitic weed in sorghum. In Proceedings of the Biennial Conference on Emerging Challenges in Weed Management, Jabalpur, India, 15–17 February 2014; p. 227. [Google Scholar]
- Lendzemo, V.W.; Van Ast, A.; Kuyper, T.W. Can Arbuscular Mycorrhizal Fungi Contribute to Striga Management on Cereals in Africa? Outlook Agric. 2006, 35, 307–311. [Google Scholar] [CrossRef]
- Manjunatha, H.P.; Jones Nirmalnath, P.; Chandranath, H.T.; Shiney, A.; Jagadeesh, K.S. Field evalualtion of native arbuscular mycorrhizal fungi in the management of Striga in sugarcane (Saccharum officinarum L.). J. Pharm. Phytochem. 2018, 7, 2496–2500. [Google Scholar]
- Wang, Y.-Y.; Yin, Q.-S.; Qu, Y.; Li, G.-Z.; Hao, L. Arbuscular mycorrhiza-mediated resistance in tomato against Cladosporium fulvum -induced mould disease. J. Phytopathol. 2017, 166, 67–74. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, X.; Gao, P.; Li, Y.; Christensen, M.J.; Duan, T. Arbuscular mycorrhiza fungi increased the susceptibility of Astragalus adsurgens to powdery mildew caused by Erysiphe pisi. Mycology 2018, 9, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.M.P.; Srimeena, N. Arbuscular Mycorrhizal Fungi (AMF) Induced Defense Factors against the Damping-off Disease Pathogen, Pythium aphanidermatum in Chilli (Capsicum annum). Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 2243–2248. [Google Scholar] [CrossRef]
- Singh, P.K.; Singh, M.; Vyas, D. Biocontrol of Fusarium Wilt of Chickpea using Arbuscular Mycorrhizal Fungi and Rhizobium leguminosorum Biovar. Caryologia 2010, 63, 349–353. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Roldan, A.; Pascual, J.A. Interaction between arbuscular mycorrhizal fungi and Trichoderma harzianum under conventional and low input fertilization field condition in melon crops: Growth response and Fusarium wilt biocontrol. Appl. Soil Ecol. 2011, 47, 98–105. [Google Scholar] [CrossRef]
- Kulkarni, S.A.; Kulkarni, S.; Sreenivas, M.N. Interaction Between Vesicular-Arbuscular (V-A) Mycorrhizae and Sclerotium rolfsii Sacc. in Groundnut. J. Farm Sci. 2011, 10, 919–921. [Google Scholar]
- Jones, N.; Krishnaraj, P.; Kulkarni, J.; Patil, A.; Laxmipathy, R.; Vasudeva, R. Diversity of arbuscular mycorrhizal fungi in different ecological zones of northern Karnataka. Eco Environ. Cons. 2011, 18, 1053–1058. [Google Scholar]
- Thiem, D.; Szmidt-Jaworska, A.; Baum, C.; Muders, K.; Niedojadło, K.; Hrynkiewicz, K. Interactive physiological response of potato (Solanum tuberosum L.) plants to fungal colonization and Potato virus Y (PVY) infection. Acta Mycol. 2014, 1, 291–303. [Google Scholar] [CrossRef]
- Stolyarchuk, I.M.; Shevchenko, T.P.; Polischuk, V.P.; Kripka, A.V. Virus infection course in different plant species under influence of arbuscular mycorrhiza. Microbiology 2009, 3, 70–75. [Google Scholar] [CrossRef]
- Othira, J.O. Effectiveness of arbuscular mycorrhizal fungi in protection of maize (Zea mays L.) against witchweed (Striga hermonthica Del Benth) infestation. J. Agric. Biotechnol. Sustain. Dev. 2012, 4, 37–44. [Google Scholar] [CrossRef]
- Isah, K.; Kumar, N.; Lagoke, S.O.; Atayese, M. Management of Striga hermonthica on sorghum (Sorghum bicolor) using arbuscular mycorrhizal fungi (Glomus mosae) and NPK fertilizer levels. Pak. J. Biol. Sci. 2013, 16, 1563–1568. [Google Scholar] [CrossRef][Green Version]
- Wilson, G.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, L.; Palla, M.; Agnolucci, M.; Avio, L.; Sbrana, C.; Turrini, A.; Giovannetti, M. Arbuscular Mycorrhizal Fungi and Associated Microbiota as Plant Biostimulants: Research Strategies for the Selection of the Best Performing Inocula. Agronomy 2020, 10, 106. [Google Scholar] [CrossRef]
- Nacoon, S.; Jogloy, S.; Riddech, N.; Mongkolthanaruk, W.; Kuyper, T.W.; Boonlue, S. Interaction between Phosphate Solubilizing Bacteria and Arbuscular Mycorrhizal Fungi on Growth Promotion and Tuber Inulin Content of Helianthus tuberosus L. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, M.M.; César, S.; Azcón, R.; Barea, J.M. Interactions between arbuscular mycorrhizal fungi and other microbial inoculants (Azospirillum, Pseudomonas, Trichoderma) and their effects on microbial population and enzyme activities in the rhizosphere of maize plants. Appl. Soil Ecol. 2000, 15, 261–272. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A. Synergistic Effects of the Inoculation with Plant Growth-Promoting Rhizobacteria and an Arbuscular Mycorrhizal Fungus on the Performance of Wheat. Turk. J. Agric. 2007, 31, 355–362. [Google Scholar]
- Diagne, N.; Baudoin, E.; Svistoonoff, S.; Ouattara, C.; Diouf, D.; Kane, A.; Ndiaye, C.; Noba, K.; Bogusz, D.; Franche, C.; et al. Effect of native and allochthonous arbuscular mycorrhizal fungi on Casuarina equisetifolia growth and its root bacterial community. Arid. Land Res. Manag. 2017, 32, 212–228. [Google Scholar] [CrossRef]
- Nanjundappa, A.; Bagyaraj, D.J.; Saxena, A.K.; Kumar, M.; Chakdar, H. Interaction between arbuscular mycorrhizal fungi and Bacillus spp. in soil enhancing growth of crop plants. Fungal Biol. Biotechnol. 2019, 6, 1–10. [Google Scholar] [CrossRef]
- Barea, J.M.; Azcón, R.; Azcón-Aguilar, C. Mycorrhizosphere interactions to improve plant fitness and soil quality. Antonie Leeuwenhoek 2002, 81, 343–351. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Tang, T.-T.; Xie, M.-M.; Chen, S.-M.; Zhang, S.-M. Effects of Arbuscular Mycorrhizal Fungi and Rhizobia on Physiological Activities in White Clover (Trifolium repens). Biotechnology 2019, 18, 49–54. [Google Scholar] [CrossRef][Green Version]
- Larimer, A.L.; Clay, K.; Bever, J.D. Synergism and context dependency of interactions between arbuscular mycorrhizal fungi and rhizobia with a prairie legume. Ecology 2014, 95, 1045–1054. [Google Scholar] [CrossRef]
- Xavier, L. Response of lentil under controlled conditions to co-inoculation with arbuscular mycorrhizal fungi and rhizobia varying in efficacy. Soil Biol. Biochem. 2002, 34, 181–188. [Google Scholar] [CrossRef]
- Wang, X.; Pan, Q.; Chen, F.; Yan, X.; Liao, H. Effects of co-inoculation with arbuscular mycorrhizal fungi and rhizobia on soybean growth as related to root architecture and availability of N and P. Mycorrhiza 2010, 21, 173–181. [Google Scholar] [CrossRef]
- Bagyaraj, D.J.; Manjunath, A.; Patil, R.B. Interaction between a vesicular-arbuscular mycorrhiza and rhizobium and their effects on soybean in the field. New Phytol. 1979, 82, 141–145. [Google Scholar] [CrossRef]
- Xavier, L.J.C.; Germida, J.J. Selective interactions between arbuscular mycorrhizal fungi and Rhizobium leguminosarum bv. viceae enhance pea yield and nutrition. Biol. Fertil. Soils 2003, 37, 261–267. [Google Scholar] [CrossRef]
- Abd-Alla, M.H.; El-Enany, A.-W.E.; Nafady, N.A.; Khalaf, D.M.; Morsy, F.M. Synergistic interaction of Rhizobium leguminosarum bv. viciae and arbuscular mycorrhizal fungi as a plant growth promoting biofertilizers for faba bean (Vicia faba L.) in alkaline soil. Microbiol. Res. 2014, 169, 49–58. [Google Scholar] [CrossRef]
- Jin, L.; Sun, X.; Wang, X.; Shen, Y.; Hou, F.; Chang, S.; Wang, C. Synergistic interactions of arbuscular mycorrhizal fungi and rhizobia promoted the growth of Lathyrus sativus under sulphate salt stress. Symbiosis 2010, 50, 157–164. [Google Scholar] [CrossRef]
- Chatarpaul, L.; Chakravarty, P.; Subramaniam, P. Studies in tetrapartite symbioses. Plant. Soil 1989, 118, 145–150. [Google Scholar] [CrossRef]
- Herrera, M.A.; Salamanca, C.P.; Barea, J.M. Inoculation of Woody Legumes with Selected Arbuscular Mycorrhizal Fungi and Rhizobia to Recover Desertified Mediterranean Ecosystems. Appl. Environ. Microbiol. 1993, 59, 129–133. [Google Scholar] [CrossRef]
- Orfanoudakis, M.Z.; Hooker, J.E.; Wheeler-Jones, C.T. Early interactions between arbuscular mycorrhizal fungi and Frankia during colonisation and root nodulation of Alnus glutinosa. Symbiosis 2004, 36, 69–82. [Google Scholar]
- Orfanoudakis, M.; Wheeler, C.T.; Hooker, J.E. Both the arbuscular mycorrhizal fungus Gigaspora rosea and Frankia increase root system branching and reduce root hair frequency in Alnus glutinosa. Mycorrhiza 2009, 20, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.S.; Castro, P.M.L.; Dodd, J.; Vosátka, M. Synergistic effect of Glomus intraradices and Frankia spp. on the growth and stress recovery of Alnus glutinosa in an alkaline anthropogenic sediment. Chemosphere 2005, 60, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Andrade, D.D.S.; Leal, A.C.; Ramos, A.L.M.; De Goes, K.C.G.P. Growth of Casuarina cunninghamiana inoculated with arbuscular mycorrhizal fungi and Frankia actinomycetes. Symbiosis 2015, 66, 65–73. [Google Scholar] [CrossRef]
- Chonglu, Z.; Mingqin, G.; Yu, C.; Fengzhen, W. Inoculation of Casuarina with Ectomycorrhizal Fungi, Vesicular-Arbuscular Mycorrhizal Fungi and Frankia. In Mycorrhizas for Plantation Forestry in Asia—ACIAR. In Proceedings of the International Symposium and Workshop, Kaiping, China, 7–11 November 2014; p. 122. [Google Scholar]
- Wheeler, C.; Tilak, M.; Scrimgeour, C.; Hooker, J.; Handley, L. Effects of symbiosis with Frankia and arbuscular mycorrhizal fungus on the natural abundance of 15N in four species of Casuarina. J. Exp. Bot. 2000, 51, 287–297. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Visser, S.; Danielson, R.M.; Parkinson, D. Field performance of Elaeagnus commutata and Shepherdia canadensis (Elaeagnaceae) inoculated with soil containing Frankia and vesicular–arbuscular mycorrhizal fungi. Can. J. Bot. 1991, 69, 1321–1328. [Google Scholar] [CrossRef]
- Cely, M.V.T.; Siviero, M.A.; Emiliano, J.; Spago, F.R.; Freitas, V.F.; Barazetti, A.R.; Goya, E.T.; Lamberti, G.D.S.; Dos Santos, I.M.O.; De Oliveira, A.G.; et al. Inoculation of Schizolobium parahyba with Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Increases Wood Yield under Field Conditions. Front. Plant. Sci. 2016, 7. [Google Scholar] [CrossRef]
- Vafadar, F.; Amooaghaie, R.; Otroshy, M. Effects of plant-growth-promoting rhizobacteria and arbuscular mycorrhizal fungus on plant growth, stevioside, NPK, and chlorophyll content of Stevia rebaudiana. J. Plant. Interact. 2013, 9, 128–136. [Google Scholar] [CrossRef]
- Karthikeyan, B.; Abitha, B.; Henry, A.J.; Sa, T.; Joe, M.M. Interaction of Rhizobacteria with Arbuscular Mycorrhizal Fungi (AMF) and Their Role in Stress Abetment in Agriculture. In Fungal Biology; Springer Science and Business Media LLC: Berlin, Germany, 2016; pp. 117–142. [Google Scholar]
- Aalipour, H.; Nikbakht, A.; Etemadi, N.; Rejali, F.; Soleimani, M. Biochemical response and interactions between arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria during establishment and stimulating growth of Arizona cypress (Cupressus arizonica G.) under drought stress. Sci. Hortic. 2020, 261, 108923. [Google Scholar] [CrossRef]
- Tank, N.; Saraf, M. Phosphate solubilization, exopolysaccharide production and indole acetic acid secretion by rhizobacteria isolated from Trigonella foenum-graecum. Indian J. Microbiol. 2003, 43, 37–40. [Google Scholar]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Shah, M.A. Induction of Osmoregulation and Modulation of Salt Stress in Acacia gerrardii Benth. by Arbuscular Mycorrhizal Fungi and Bacillus subtilis (BERA 71). Biomed Res. Int. 2016, 2016, 1–11. [Google Scholar] [CrossRef]
- Lesueur, D.; Duponnois, R. Relations between rhizobial nodulation and root colonization of Acacia crassicarpa provenances by an arbuscular mycorrhizal fungus, Glomus intraradices Schenk and Smith or an ectomycorrhizal fungus, Pisolithus tinctorius Coker & Couch. Ann. Sci. 2005, 62, 467–474. [Google Scholar] [CrossRef][Green Version]
- Rajendran, K.; Devaraj, P. Biomass and nutrient distribution and their return of Casuarina equisetifolia inoculated with biofertilizers in farm land. Biomass Bioenergy 2004, 26, 235–249. [Google Scholar] [CrossRef]
- Chilvers, G.A.; Lapeyrie, F.F.; Horan, D.P. Ectomycorrhizal vs Endomycorrhizal fungi within the same root system. New Phytol. 1987, 107, 441–448. [Google Scholar] [CrossRef]
- Duponnois, R.; Diédhiou, S.; Chotte, J.L.; Sy, M.O. Relative importance of the endomycorrhizal and (or) ectomycorrhizal associations in Allocasuarina and Casuarina genera. Can. J. Microbiol. 2003, 49, 281–287. [Google Scholar] [CrossRef]
- Tian, C.; He, X.; Zhong, Y.; Chen, J. Effect of inoculation with ecto- and arbuscular mycorrhizae and Rhizobium on the growth and nitrogen fixation by black locust, Robinia pseudoacacia. New For. 2003, 25, 125–131. [Google Scholar] [CrossRef]
- Elumalai, S.; Raaman, N. In vitro synthesis of Frankia and mycorrhiza with Casuarina equisetifolia and ultrastructure of root system. Indian J. Exp. Biol. 2009, 47, 289–297. [Google Scholar]
- Gunasekera, D.; Gunawardana, D.; Jayasinghearachchi, H. Diversity of Actinomycetes in Nitrogen Fixing Root Nodules of Casuarina equisetifolia and its Impact on Plant Growth. Int. J. Multidiscip. Stud. 2016, 3, 17. [Google Scholar] [CrossRef]
- Duponnois, R. Mycorrhiza Helper Bacteria: Their Ecological Impact in Mycorrhizal Symbiosis. Handb. Microb. Biofertil. 2006, 117, 231–250. [Google Scholar]
- Frey-Klett, P.; Garbaye, J.; Tarkka, M. The mycorrhiza helper bacteria revisited. New Phytol. 2007, 176, 22–36. [Google Scholar] [CrossRef]
- Rigamonte, T.A.; Pylro, V.S.; Duarte, G.F. The role of mycorrhization helper bacteria in the establishment and action of ectomycorrhizae associations. Braz. J. Microbiol. 2010, 41, 832–840. [Google Scholar] [CrossRef]
| Host Plants | AMF Strains | Responses Related to AMF Inoculation | References |
|---|---|---|---|
| Panicum hemitomon Schult. Schultes Leersia hexandra Schwartz | Acaulospora trappei R.N. Ames & Linderman 1976, Scutellospora heterogama, A Acaulospora laevis Gerd. & Trappe 1974, Glomus leptotichum N.C. Schenck & G.S. Sm 1982, G. etunicatum and Glomus gerdemannii S.L. Rose, B.A. Daniels & Trappe 1979 | Improve phosphorus (P) nutrition Greater tissue P concentrations | [19] |
| Pterocarpus officinalis (Jacq.) | Glomus intraradices | Improve plant growth and P acquisition in leaves | [91] |
| Aster tripolium L. | Glomus geosporum T.H. Nicolson & Gerd.) C. Walker 1982 | Higher concentrations of soluble sugars and proline | [93] |
| Host Plants | AMF Strains | Responses Related to AMF Inoculation | References |
|---|---|---|---|
| Cucumis sativus L. | R. irregularis | Increases the photosynthetic efficiency of cold-stressed cucumber seedlings by protecting their photosynthetic apparatus against light-induced damage and increasing their carbon sink. | [51] |
| Zea mays L. | Funneliformis (Glomus) species. | Regulated photosystem (PS) II heterogeneity | [98] |
| Hordeum vulgare L. | G. versiforme R. irregularis | Increasing the survival rate, alleviation of low-temperature stress | [99] |
| Cyclamen persicum Mill. | R. fasciculatum | Enhanced biomass production and heat stress response Increase activity of antioxidative enzymes such as superoxide dismutase and ascorbate peroxidase | [103] |
| Elymus nutans Griseb. | F. mosseae | Less oxidative damage Promoted plant growth and enhanced the level of chlorophyll and antioxidant compounds such as glutathione and soluble sugars | [104] |
| Host Plants | AMF Strains | Responses Related to AMF Inoculation | References |
|---|---|---|---|
| Lycopersicon esculentum L. | F. mosseae | Increased plant growth, fruit weight, and yield, chlorophyll content, concentrations of P and K, antioxidant enzymes activities (SOD, CAT, POD, and APX), and reduced Na concentration in leaves | [97] |
| Capsicum annuum L. | R. irregularis | Increased leaf area, mineral content, proline, sugars, and cell membrane integrity, and reduced shoot content of Na | [106] |
| Acacia auriculiformis A. Cunn. ex Benth. | R. fasciculatus Glomus macrocarpum Tul. & C. Tul. 1845 | Increased root and shoot weights, and greater nutrient acquisition, changes in root morphology, and electrical conductivity of the soil | [109] |
| Solanum lycopersicum L. | R. irregularis | Enhanced shoot and root dry weights, chlorophyll and proline concentrations, nutrient uptake (P, Ca, and K), stomatal conductance, the activity of ROS scavenging enzymes (APX, CAT, POD, and SOD) and protecting photochemical processes of PSII | [111] |
| Triticum aestivum L. | G. etunicatum F. mosseae R. irregularis | Increased plant growth, nutrient uptake and grain yield, and reduced concentrations of Na+ and Cl− | [112] |
| Zea mays L. | R. irregularis (isolate EEZ 58) R. irregularis (Ri CdG) S. constrictum (Trappe) Sieverd., G.A. Silva & Oehl 2011 (Sc CdG) Claroideoglomus etunicatum (W.N. Becker & Gerd.) C. Walker & A. Schüßler, 2010 (Ce CdG) (Ce CdG) | Improved K+ and Na+ homeostasis, shoot and root dry weights, K concentration in shoots, and reduced Cl and Na contents in shoots | [119] |
| Digitaria eriantha Steud. | R. irregularis | Increased stomatal conductance, antioxidant enzymes activities (CAT et APX), jasmonate content, and reduced root and shoot hydrogen peroxide accumulation | [120] |
| Lycopersicon esculentum Mill. Cv. Marriha | F. mosseae | Improved plant biomass, fruit fresh yield and shoot contents of P, K, Cu, Fe and Zn, and reduced shoot Na concentrations | [121] |
| Acacia nilotica Willd. | R. fasciculatum | Improved root and shoot biomass, and nutrient concentrations (P, Zn, K, and Cu), and Na concentration | [122] |
| Acacia saligna (Labill.) H.L. Wendl. | AMF | Improved plant growth and dry weight, nodulation parameters, chlorophylls, carbohydrates, proline and nutrient contents (N, P, K and Ca) and reduced Na concentrations | [123] |
| Acacia auriculiformis Acacia mangium | R. irregularis | Enhanced plant growth and nodulation, and nutrient contents (P, N) | [125] |
| Gossypium arboreum L. | F. mosseae (isolate GM1) F. mosseae (isolate GM2) | Increased biomass and phophorus concentrations | [127] |
| Cucumis sativus L. | C. etunicatum R. irregularis F. mosseae | Enhancing the biomass, synthesis of pigments, activity of antioxidant enzymes, including superoxide dismutase, catalase, ascorbate peroxidase, and glutathione reductase, and the content of ascorbic acid Enhancing jasmonic acid, salicylic acid and several important mineral elements (K, Ca, Mg, Zn, Fe, Mn, and Cu) Reducing the uptake of deleterious ions like Na+ | [128] |
| Glycine max L. Merrill | C. etunicatum R. irregularis F. mosseae | Protected soybean genotypes from salt-induced membrane damage Reduced the production of hydrogen peroxide and lipid peroxidation Improved plant growth and symbiotic performance by stimulating the endogenous level of auxins that contribute to improved root systems and nutrient acquisition under salt stress | [129] |
| Host Plants | AMF Strains | Responses Related to AMF Inoculation | References |
|---|---|---|---|
| Trigonella foenum-graceum L. | F. mosseae | Better plant performance | [132] |
| Zea mays L. | Glomus isolates | Improved dry weight and contents of essential elements (K, P, and Mg), and distinct differences in the cellular distribution of heavy metals and essential elements | [133] |
| Lonicera japonica Thunb. | G. versiforme R. intraradices | Decreased Cd concentrations in shoots and roots, Reduced Cd concentrations in shoots but increased Cd concentrations in roots | [140] |
| Solanum lycopersicum L. | F. mosseae (syn. Glomus mosseae), R. intraradices (syn. Glomus intraradices) C. etunicatum (syn. Glomus etunicatum) | AMF reduced the production of malonaldehyde and hydrogen peroxide by mitigating oxidative stress. AMF strengthened the plant’s defense system and provide efficient protection against Cd stress | [141] |
| Populus alba Villafranca Populus nigra Jean Pourtet | F. mosseae or R. irregularis | Alleviation of Cu and Zn phytotoxicity | [142] |
| Trifolium pratense L. | Glomus mosseae | Decreases in Zn uptake and in root and shoot concentrations | [143] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of Arbuscular Mycorrhizal Fungi on Plant Growth and Performance: Importance in Biotic and Abiotic Stressed Regulation. Diversity 2020, 12, 370. https://doi.org/10.3390/d12100370
Diagne N, Ngom M, Djighaly PI, Fall D, Hocher V, Svistoonoff S. Roles of Arbuscular Mycorrhizal Fungi on Plant Growth and Performance: Importance in Biotic and Abiotic Stressed Regulation. Diversity. 2020; 12(10):370. https://doi.org/10.3390/d12100370
Chicago/Turabian StyleDiagne, Nathalie, Mariama Ngom, Pape Ibrahima Djighaly, Dioumacor Fall, Valérie Hocher, and Sergio Svistoonoff. 2020. "Roles of Arbuscular Mycorrhizal Fungi on Plant Growth and Performance: Importance in Biotic and Abiotic Stressed Regulation" Diversity 12, no. 10: 370. https://doi.org/10.3390/d12100370
APA StyleDiagne, N., Ngom, M., Djighaly, P. I., Fall, D., Hocher, V., & Svistoonoff, S. (2020). Roles of Arbuscular Mycorrhizal Fungi on Plant Growth and Performance: Importance in Biotic and Abiotic Stressed Regulation. Diversity, 12(10), 370. https://doi.org/10.3390/d12100370