Hydroxymethylfurfural Affects Caged Honey Bees (Apis mellifera carnica)



Abstract
1. Introduction
2. Materials and Methods
2.1. Toxicological Tests
2.2. Immunohistochemical Analyses
2.3. Semi-Quantitative Analysis of Cell Death
2.4. Nosema Ceranae Spore Counts
3. Results
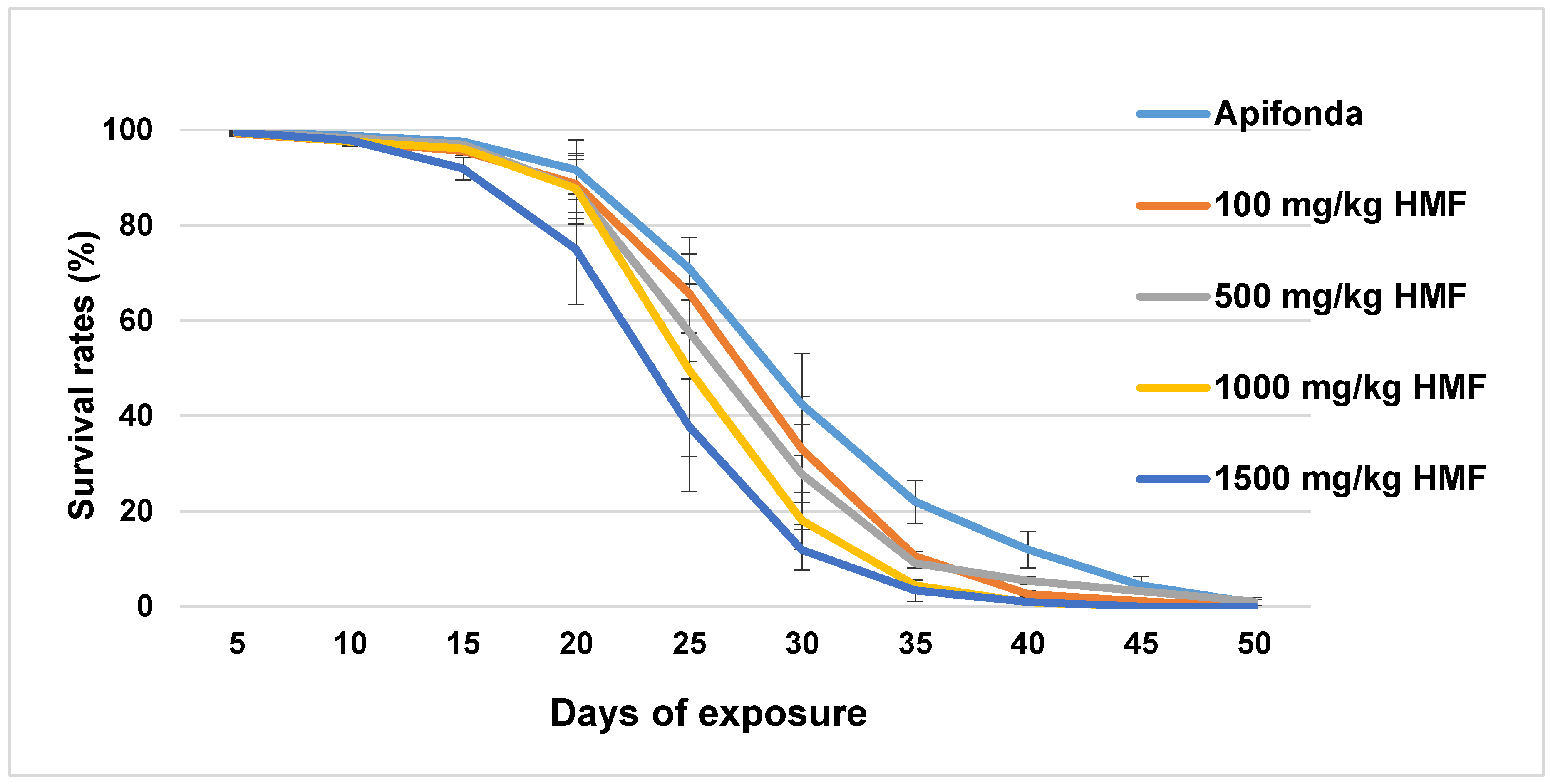
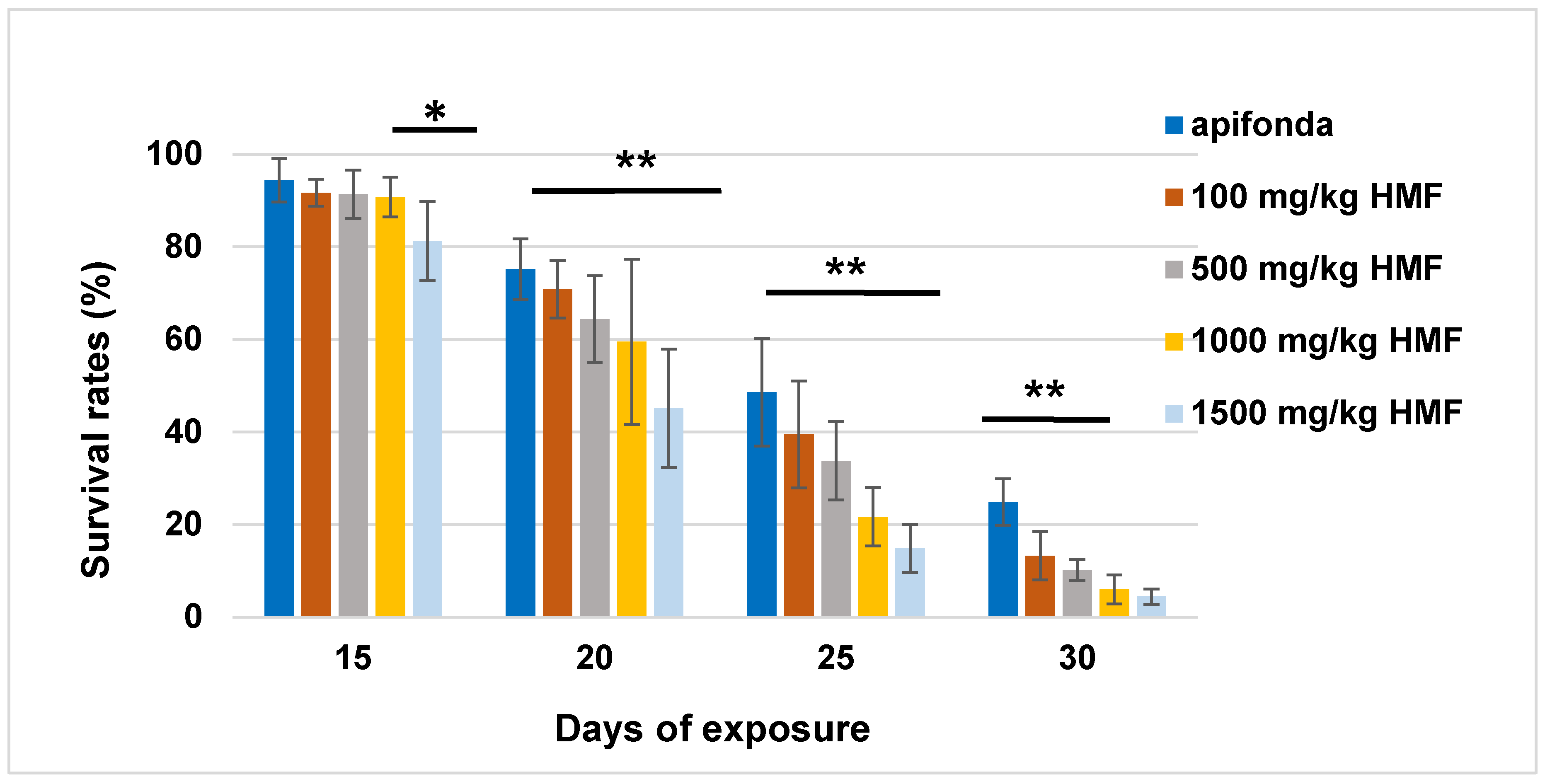
3.1. Longevity of Bees
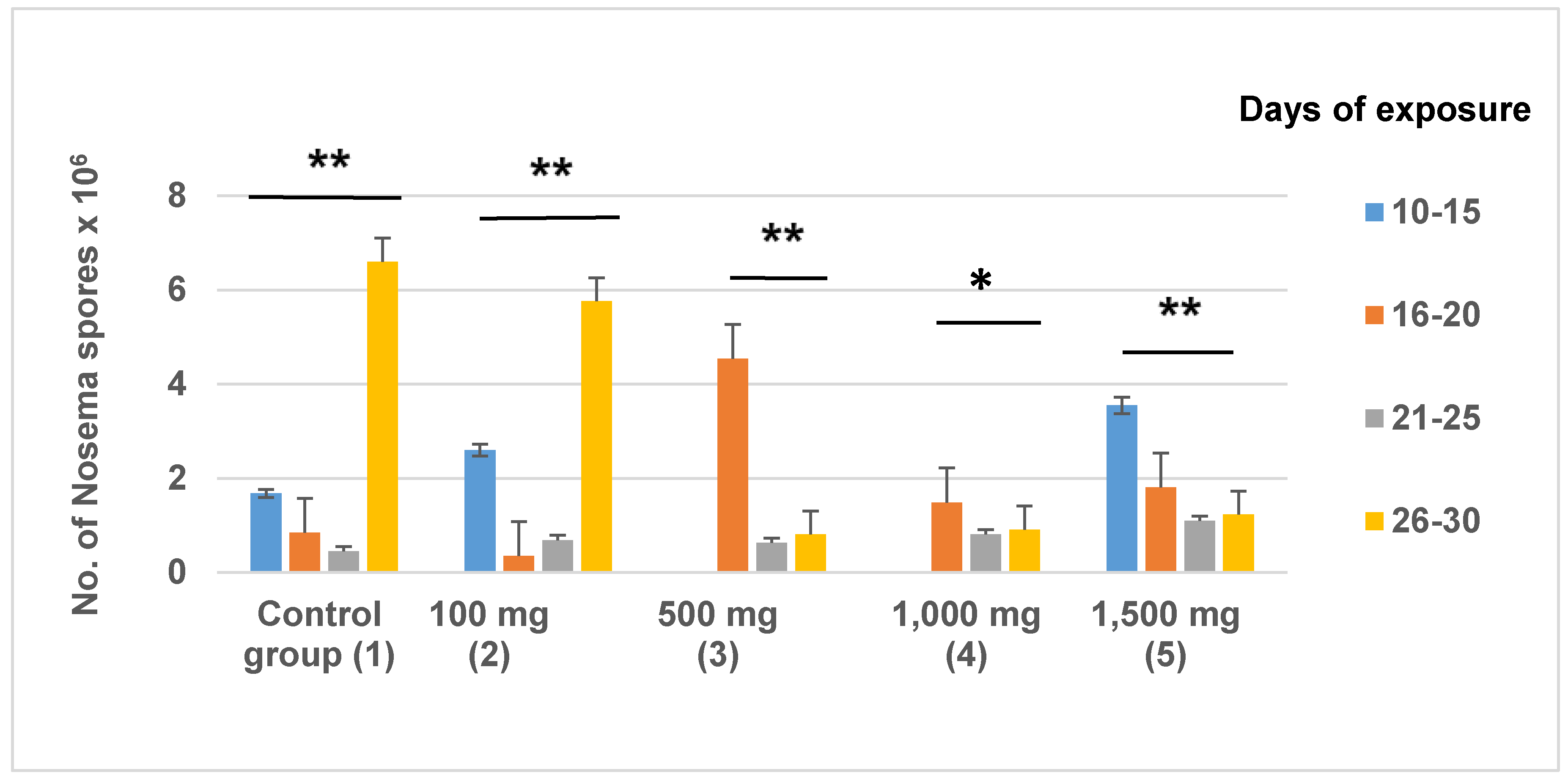
3.2. Nosema Ceranae Spores
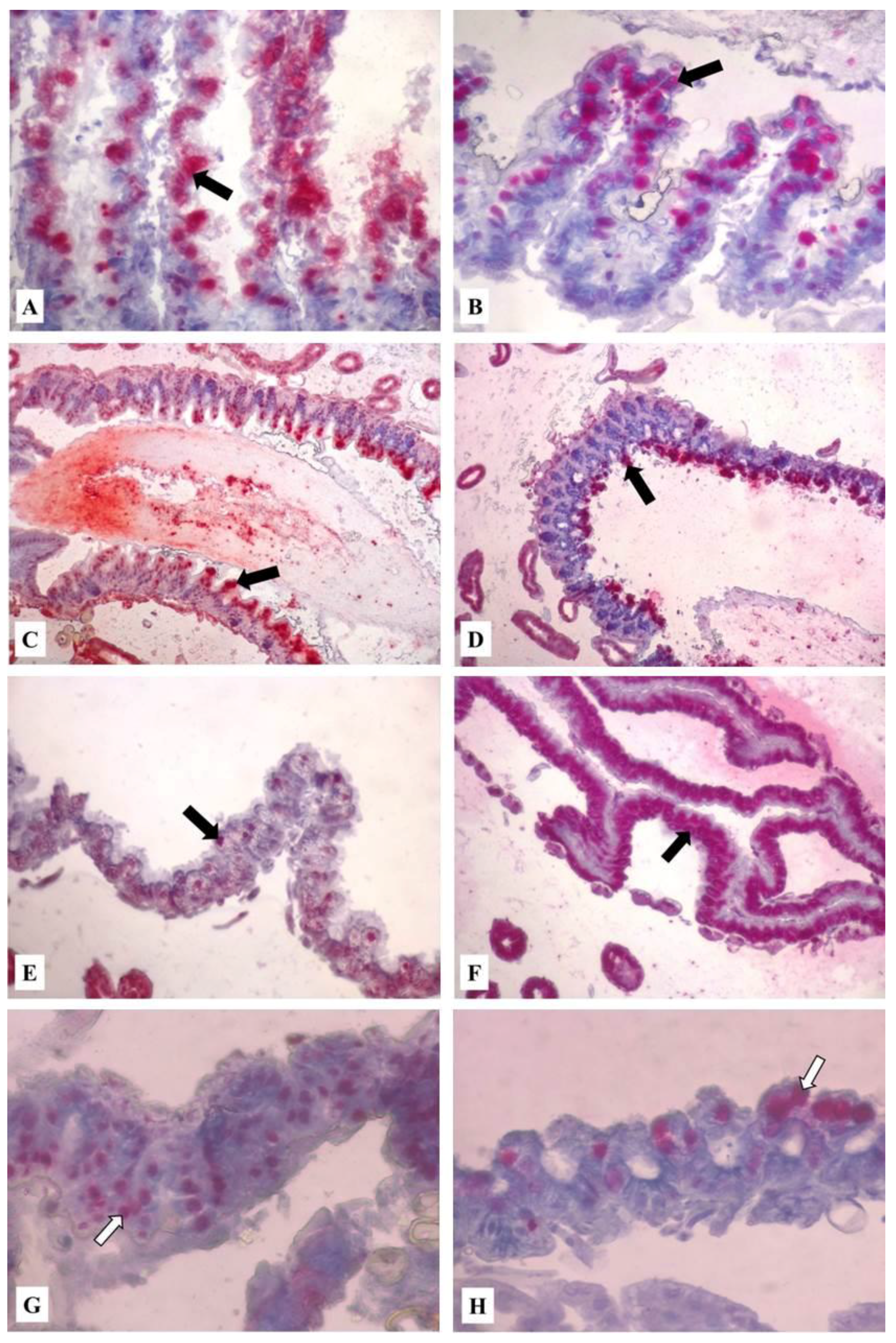
3.3. Immunohistochemical Analyses
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, J.W., Jr.; Doner, L. Honey Composition and Properties. In Beesource Beekeeping; U.S. Government Printing Office: Washington, DC, USA, 1980; pp. 82–91. [Google Scholar]
- Hermersdörfer, H.J. Lipp: Der Honig. 3. völlig neubearbeitete und erweiterte Auflage. 205 Seiten, 66 Abbildungen, 8 Farbtafeln und 30 Tabellen. Verlag Eugen Ulmer, Stuttgart 1994. Preis: 88,-DM. Food/Nahrung 1995, 39, 540. [Google Scholar]
- Shalumova, T.; Tanski, J.M. 5-(Hy-droxy-meth-yl)furan-2-carbaldehyde. Acta Cryst. Sect. E Struct. Rep. Online 2010, 66, o2266. [Google Scholar] [CrossRef]
- Teixidó, E.; Santos, F.J.; Puignou, L.; Galceran, M.T. Analysis of 5-hydroxymethylfurfural in foods by gas chromatography—Mass spectrometry. J. Chromatogr. A 2006, 1135, 85–90. [Google Scholar] [CrossRef]
- Basumallick, L.; Rohrer, J. Determination of Hydroxymethylfurfural in Honey and Biomass. Thermo Sci. Dionex Appl. Note Update 2001, 10, 6. [Google Scholar]
- Lee, H.S.; Nagy, S. Relative reactivities of sugars in the formation of 5-hydroxymethylfurfural in sugar-catalyst model systems1. J. Food Process. Preserv. 1990, 14, 171–178. [Google Scholar] [CrossRef]
- Gökmen, V.; Açar, Ö.Ç.; Köksel, H.; Acar, J. Effects of dough formula and baking conditions on acrylamide and hydroxymethylfurfural formation in cookies. Food Chem. 2007, 104, 1136–1142. [Google Scholar] [CrossRef]
- Morales, F.J. Hydroxymethylfurfural (HMF) and Related Compounds. In Process-Induced Food Toxicants: Occurrence, Formation, Mitigation, and Health Risks; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 978-0-470-07475-6. [Google Scholar]
- Tosi, E.; Ciappini, M.; Ré, E.; Lucero, H. Honey thermal treatment effects on hydroxymethylfurfural content. Food Chem. 2002, 77, 71–74. [Google Scholar] [CrossRef]
- De Ribeiro, R.O.R.; da Carneiro, C.S.; Mársico, E.T.; Cunha, F.L.; Conte, C.A., Jr.; Mano, S.B. Influence of the time/temperature binomial on the hydroxymethylfurfural content of floral honeys subjected to heat treatment. Ciência Agrotecnologia 2012, 36, 204–209. [Google Scholar] [CrossRef]
- Jachimowicz, T.; Sherbiny, G.E. Zur problematik der verwendung von invertzucker für die bienenfütterung. Apidologie 1975, 6, 121–143. [Google Scholar] [CrossRef]
- Bailey, L. The Effect of Acid-Hydrolysed Sucrose on Honeybees. J. Apic. Res. 1966, 5, 127–136. [Google Scholar] [CrossRef]
- Kammerer, F.X. Aktueller Standder Erkenntnisse über die Fütterung vonBienen mit Zucker. Imkerfreund 1989, 1, 12–14. [Google Scholar]
- LeBlanc, B.W.; Eggleston, G.; Sammataro, D.; Cornett, C.; Dufault, R.; Deeby, T.; Cyr, E. Formation of Hydroxymethylfurfural in Domestic High-Fructose Corn Syrup and Its Toxicity to the Honey Bee (Apis mellifera). J. Agric. Food Chem. 2009, 57, 7369–7376. [Google Scholar] [CrossRef] [PubMed]
- Van der Zee, R.; Pisa, L. Bijensterfte 2009-10 en toxische invertsuikersiroop. NCB Rapp. 2010, 2, 1–15. [Google Scholar]
- Stabentheiner, A.; Kovac, H.; Brodschneider, R. Honeybee Colony Thermoregulation—Regulatory Mechanisms and Contribution of Individuals in Dependence on Age, Location and Thermal Stress. PLoS ONE 2010, 5, e8967. [Google Scholar] [CrossRef] [PubMed]
- Gates, B.N. The Temperature of the Bee Colony; U.S. Department of Agriculture: Washington, DC, USA, 1914.
- Evans, J.D.; Chen, Y.P.; di Prisco, G.; Pettis, J.; Williams, V. Bee cups: Single-use cages for honey bee experiments. J. Apic. Res. 2009, 48, 300–302. [Google Scholar] [CrossRef]
- Gregorc, A.; Alburaki, M.; Werle, C.; Knight, P.R.; Adamczyk, J. Brood removal or queen caging combined with oxalic acid treatment to control varroa mites (Varroa destructor) in honey bee colonies (Apis mellifera). Apidologie 2017, 48, 821–832. [Google Scholar] [CrossRef]
- Gregorc, A.; Sampson, B.; Knight, P.R.; Adamczyk, J. Diet quality affects honey bee (Hymenoptera: Apidae) mortality under laboratory conditions. J. Apic. Res. 2019, 58, 492–493. [Google Scholar] [CrossRef]
- Statistical Graphics Corporation. Statgraphics Version 5: Statistical Graphics System; STSC Inc.: Rockville, MD, USA, 1991; ISBN 978-0-926683-06-8. [Google Scholar]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in fish: Proposal for a protocol to assess aquatic pollution. J. Fish Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef]
- Gregorc, A.; Silva-Zacarin, E.C.M.; Carvalho, S.M.; Kramberger, D.; Teixeira, E.W.; Malaspina, O. Effects of Nosema ceranae and thiametoxam in Apis mellifera: A comparative study in Africanized and Carniolan honey bees. Chemosphere 2016, 147, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Higes, M.; García-Palencia, P.; Martín-Hernández, R.; Meana, A. Experimental infection of Apis mellifera honeybees with Nosema ceranae (Microsporidia). J. Invertebr. Pathol. 2007, 94, 211–217. [Google Scholar] [CrossRef]
- Ceksteryte, V.; Racys, J. The quality of syrups used for bee feeding before winter and their suitability for bee wintering. J. Apic. Sci. 2006, 5, 5–14. [Google Scholar]
- Krainer, S.; Brodschneider, R.; Vollmann, J.; Crailsheim, K.; Riessberger-Gallé, U. Effect of hydroxymethylfurfural (HMF) on mortality of artificially reared honey bee larvae (Apis mellifera carnica). Ecotoxicology 2016, 25, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Okuda, K.; de Almeida, F.; Mortara, R.A.; Krieger, H.; Marinotti, O.; Tania Bijovsky, A. Cell death and regeneration in the midgut of the mosquito, Culex quinquefasciatus. J. Insect Physiol. 2007, 53, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Rost-Roszkowska, M.M.; Poprawa, I.; Klag, J.; Migula, P.; Mesjasz-Przybyłowicz, J.; Przybyłowicz, W. Degeneration of the midgut epithelium in Epilachna cf. nylanderi (Insecta, Coccinellidae): Apoptosis, autophagy, and necrosis. Can. J. Zool. 2008, 86, 1179–1188. [Google Scholar]
- Prochazkova, P.; Roubalova, R.; Dvorak, J.; Tlaskalova-Hogenova, H.; Cermakova, M.; Tomasova, P.; Sediva, B.; Kuzma, M.; Bulant, J.; Bilej, M.; et al. Microbiota, Microbial Metabolites, and Barrier Function in A Patient with Anorexia Nervosa after Fecal Microbiota Transplantation. Microorganisms 2019, 7, 338. [Google Scholar] [CrossRef]
- Chaffey, N.; Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular biology of the cell. 4th edn. Ann. Bot. 2003, 91, 401. [Google Scholar] [CrossRef]
- Hill, C.A.; Pinnock, D.E. Histopathological Effects ofBacillus thuringiensison the Alimentary Canal of the Sheep Louse, Bovicola ovis. J. Invertebr. Pathol. 1998, 72, 9–20. [Google Scholar] [CrossRef]
- James, E.R.; Green, D.R. Infection and the origins of apoptosis. Cell Death Differ. 2002, 9, 355–357. [Google Scholar] [CrossRef]
- Rost-Roszkowska, M.M.; Machida, R.; Fukui, M. The role of cell death in the midgut epithelium in Filientomon takanawanum (Protura). Tissue Cell 2010, 42, 24–31. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age of Bees (Days) | Treatment Groups | ||||
|---|---|---|---|---|---|
| 1 (control) | 2 (100 mg HMF) | 3 (500 mg HMF) | 4 (1000 mg HMF) | 5 (1500 mg HMF) | |
| 5 | I | I | I | IV | IV |
| 10 | II | I | IV | III | I |
| 15 | II | III | IV | IV | IV |
| 20 | IV | I | IV | II | III |
| 25 | I | I | IV | III | I |
| 30 | x | x | III | I | I |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gregorc, A.; Jurišić, S.; Sampson, B. Hydroxymethylfurfural Affects Caged Honey Bees (Apis mellifera carnica). Diversity 2020, 12, 18. https://doi.org/10.3390/d12010018
Gregorc A, Jurišić S, Sampson B. Hydroxymethylfurfural Affects Caged Honey Bees (Apis mellifera carnica). Diversity. 2020; 12(1):18. https://doi.org/10.3390/d12010018
Chicago/Turabian StyleGregorc, Aleš, Snežana Jurišić, and Blair Sampson. 2020. "Hydroxymethylfurfural Affects Caged Honey Bees (Apis mellifera carnica)" Diversity 12, no. 1: 18. https://doi.org/10.3390/d12010018
APA StyleGregorc, A., Jurišić, S., & Sampson, B. (2020). Hydroxymethylfurfural Affects Caged Honey Bees (Apis mellifera carnica). Diversity, 12(1), 18. https://doi.org/10.3390/d12010018