An Introduction to the Study of Gastrotricha, with a Taxonomic Key to Families and Genera of the Group

 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Extraction
2.3. Morphological Analysis
2.4. Taxonomic Key
3. Results
Key to Families and Genera of Gastrotricha
- 1a
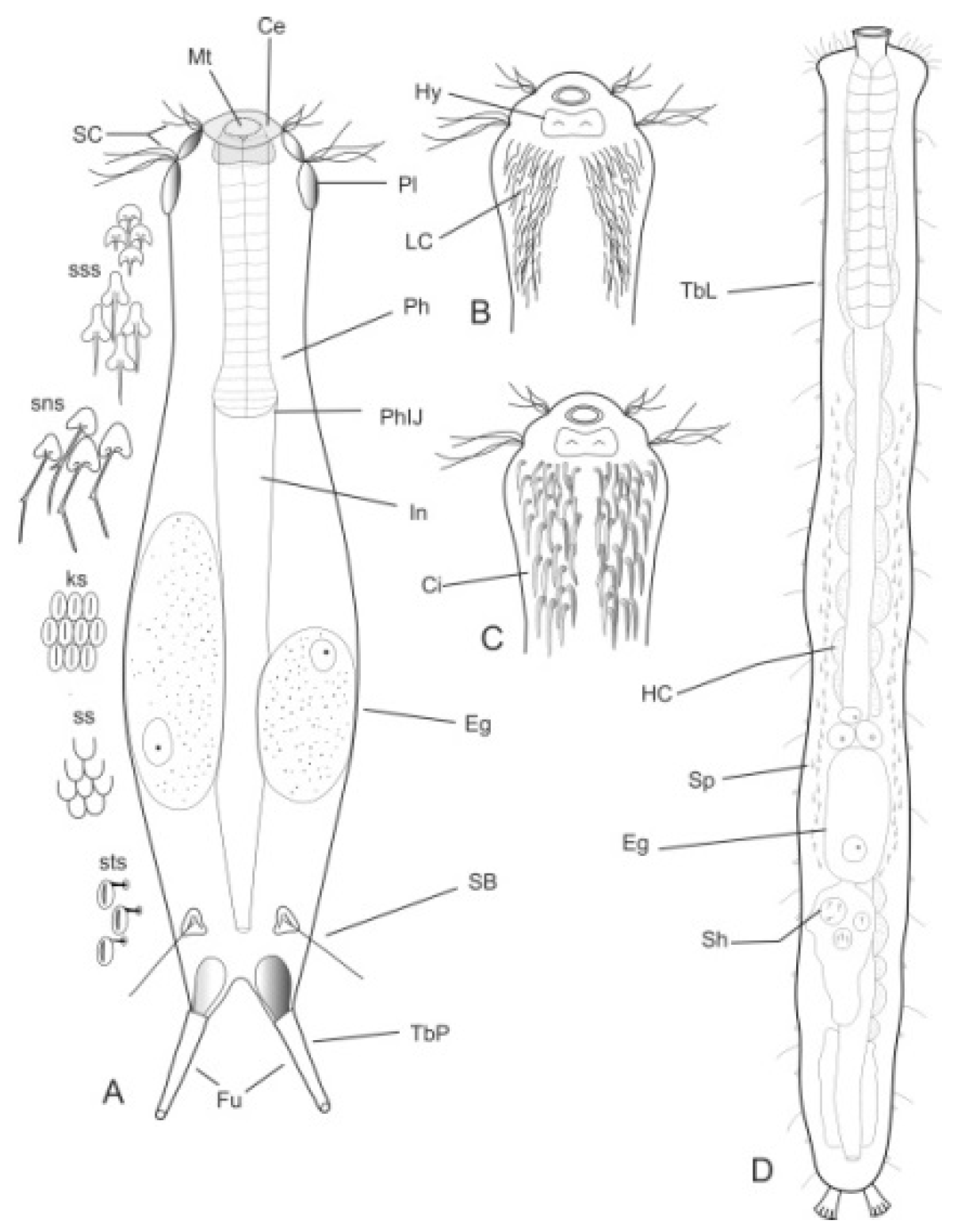
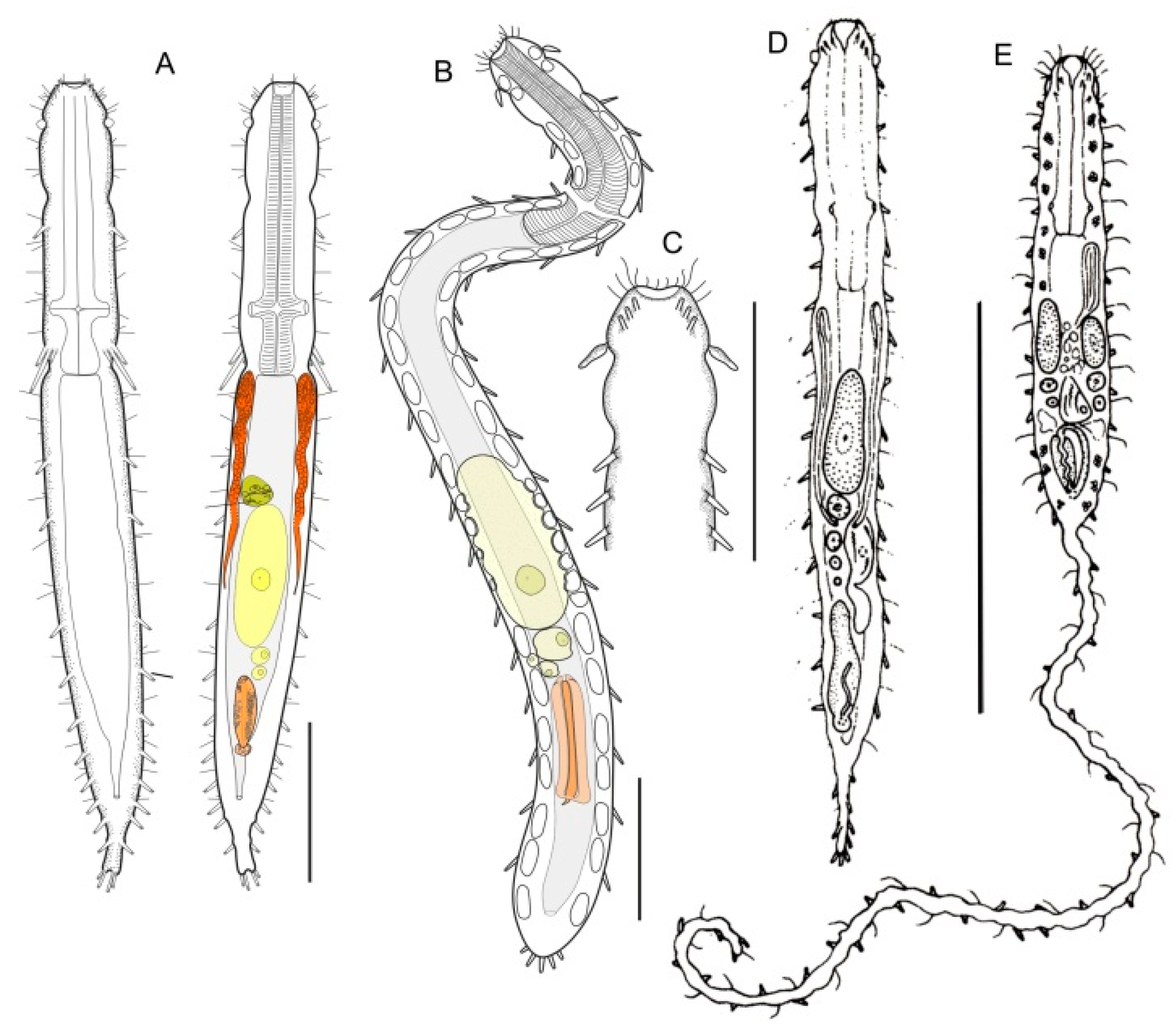
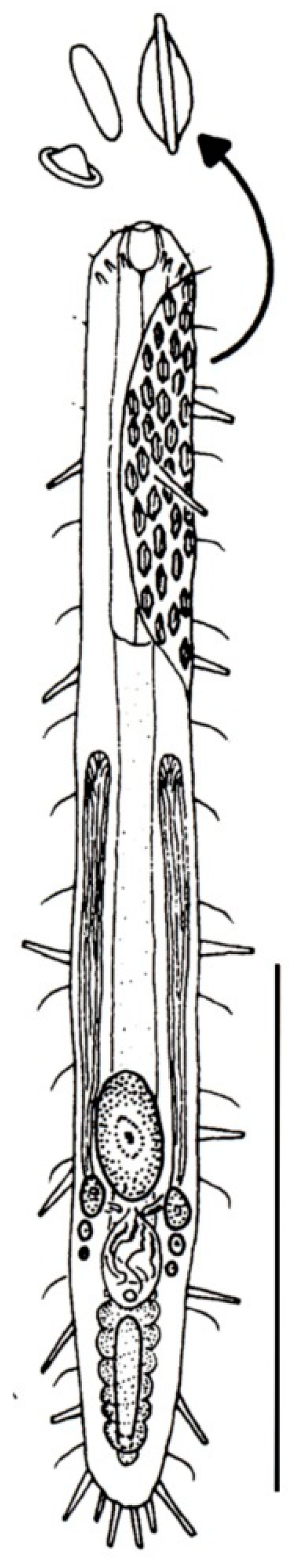
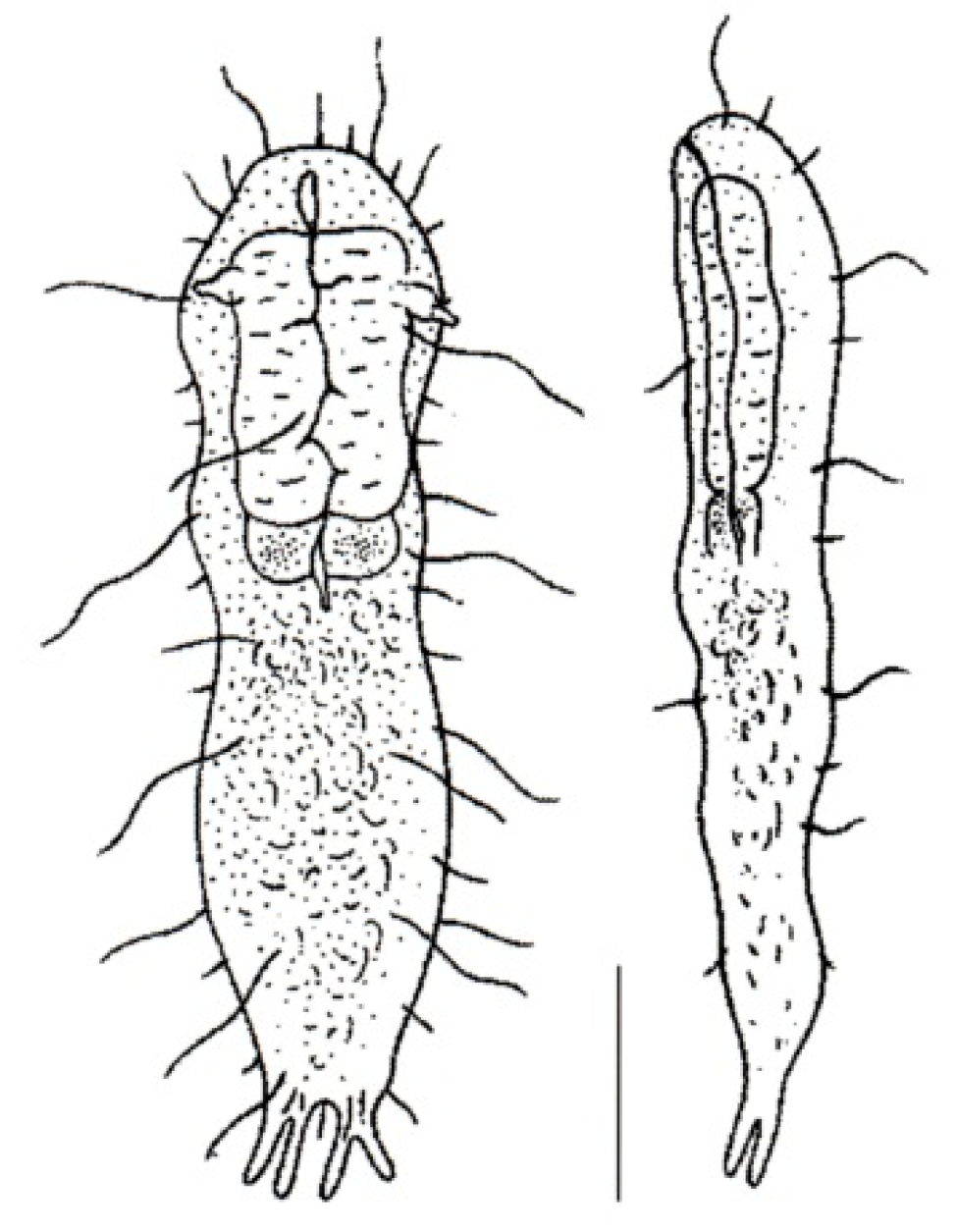
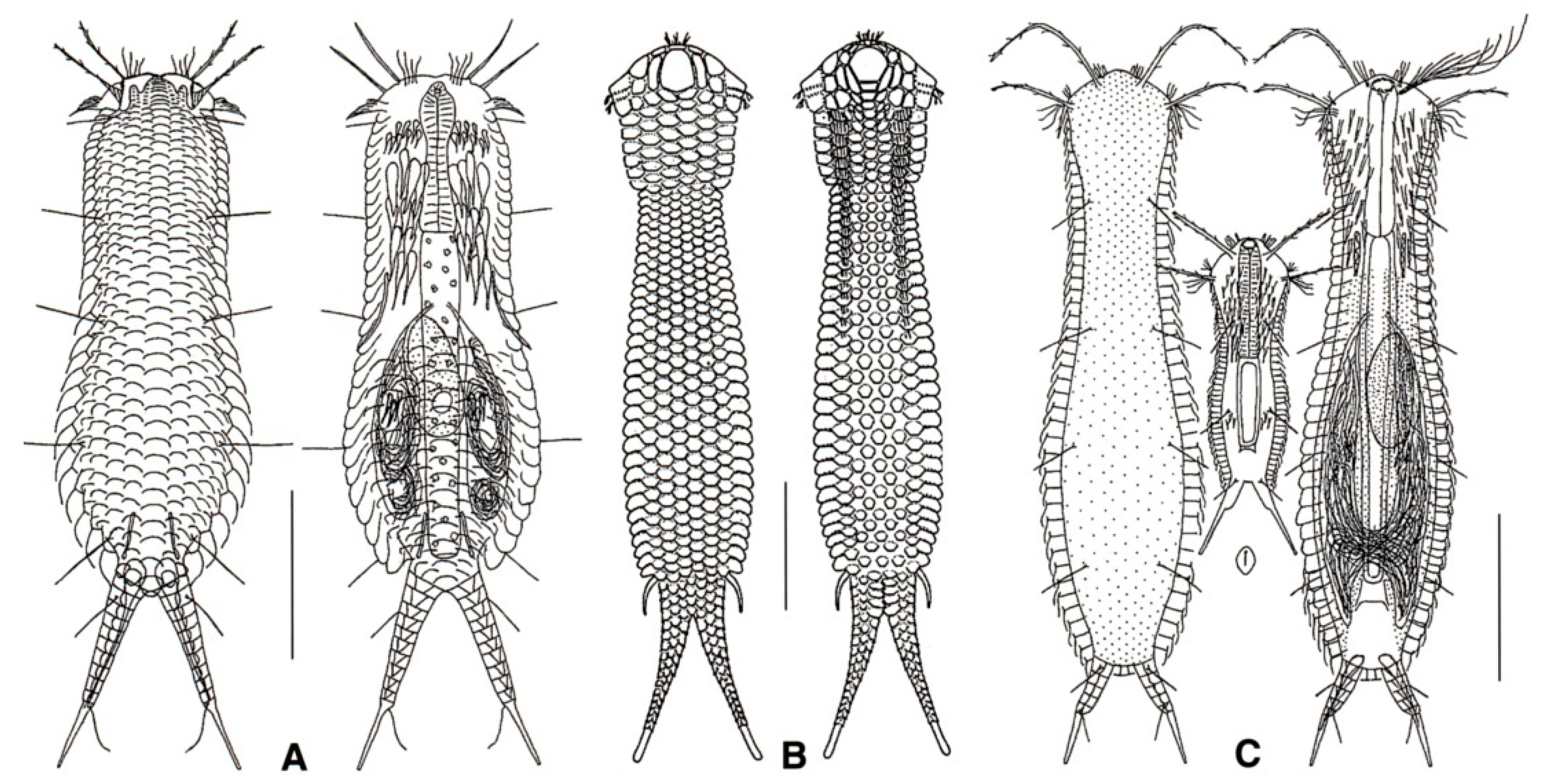
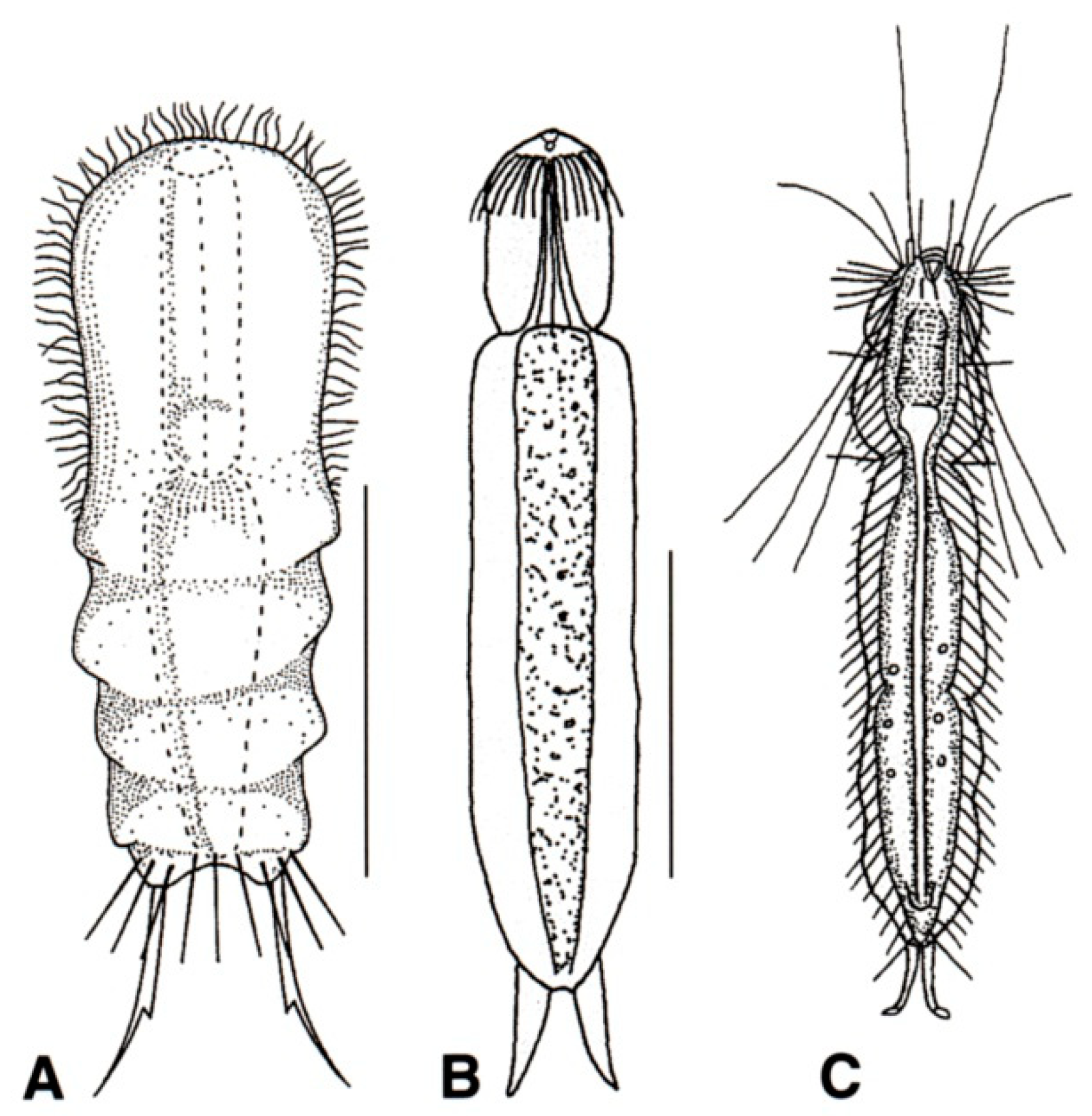
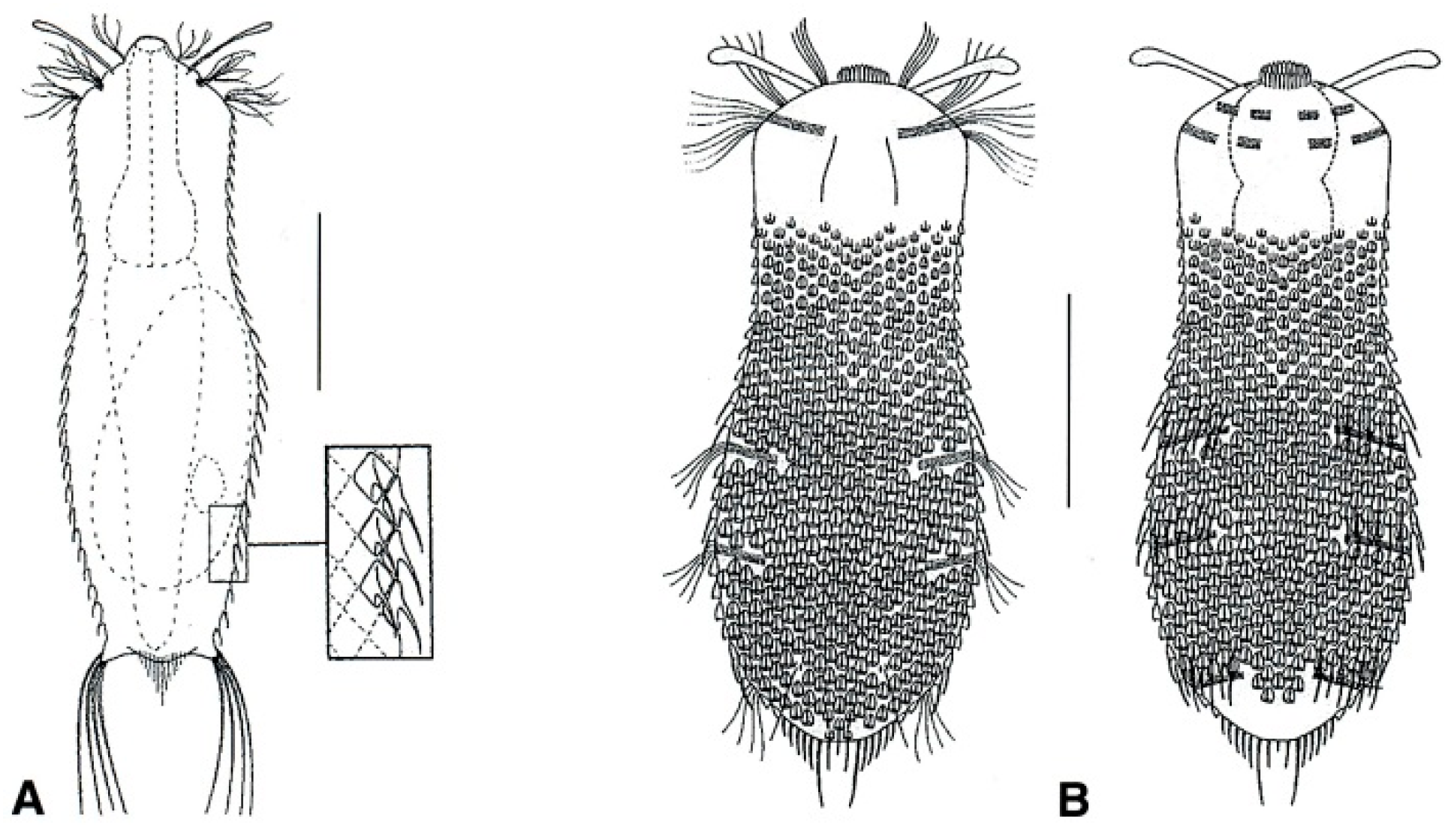
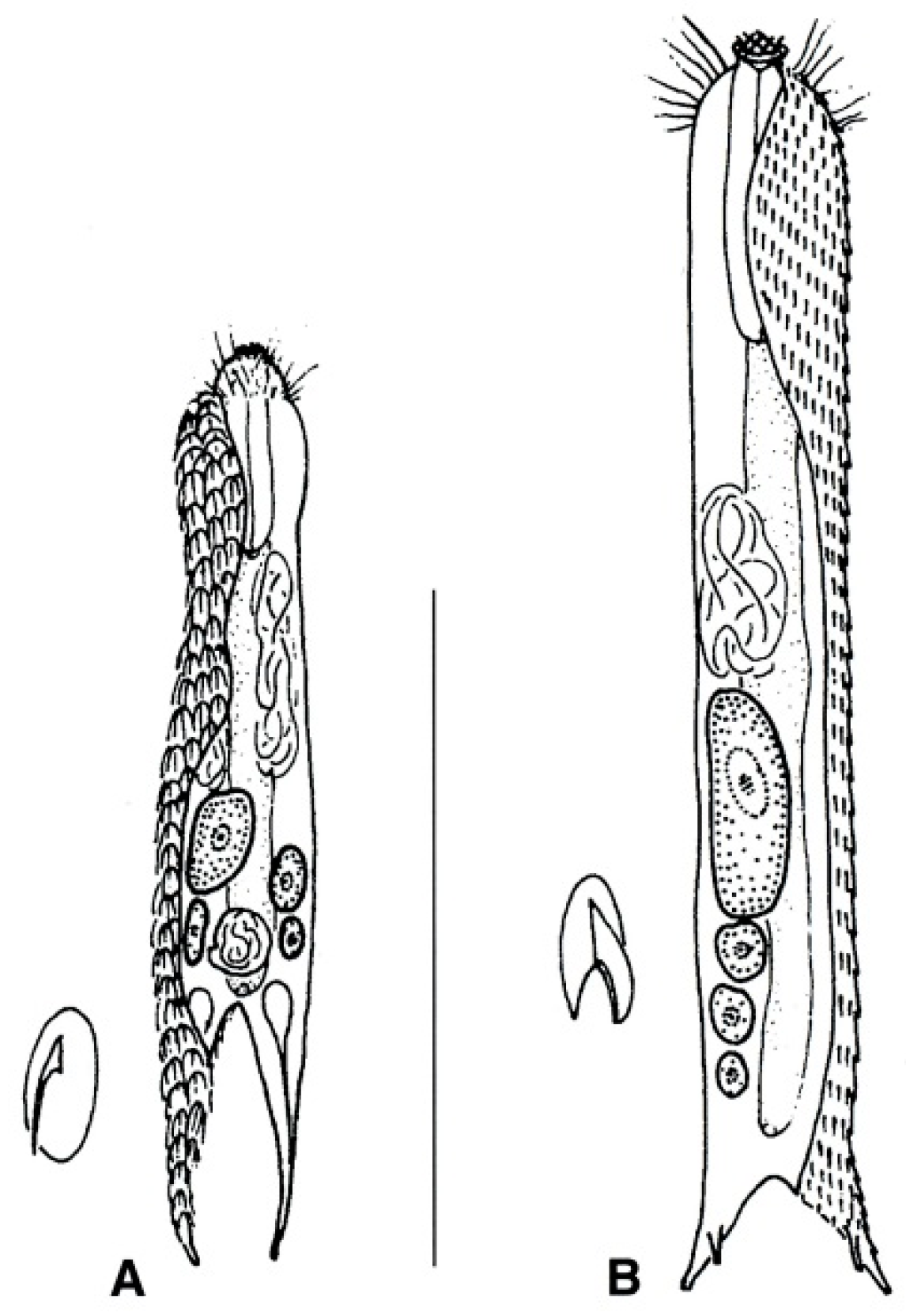
- Body flask-, bottle- or tenpin-shaped; posterior body region usually furcate (furca), less often rounded off or bifurcate; cuticle usually forming ornamentations such as scales and spines; TbA, TbD, and TbL absent; TbP present, numbering 2 (rarely 4 or 0) at the distal end of the furcal rami; mouth narrow; pharynx lacking pores. Freshwater, marine, and brackish: periphytic, epibenthic, and interstitial, occasionally semiplanktonic. Order CHAETONOTIDA, Suborder PAUCITUBULATINA (Figure 1A–C) ……………….…….... 38
- 1b
- Different from the above. …………….………..…………………………………..……....…….... 2

- 2a (1b)
- Body vermiform; cuticle naked, not forming scales and/or spines; TbA and TbD absent; TbL present in the form of numerous papillae along each side; TbP, some per side, fused at their bases forming two adhesive structures; mouth narrow, pharynx lacking pores. Uncommon; marine: interstitial. Order CHAETONOTIDA, Suborder MULTITUBULATINA, NEODASYIDAE …………………………………………………….……... Neodasys (Figure 1D)
- 2b (1b)
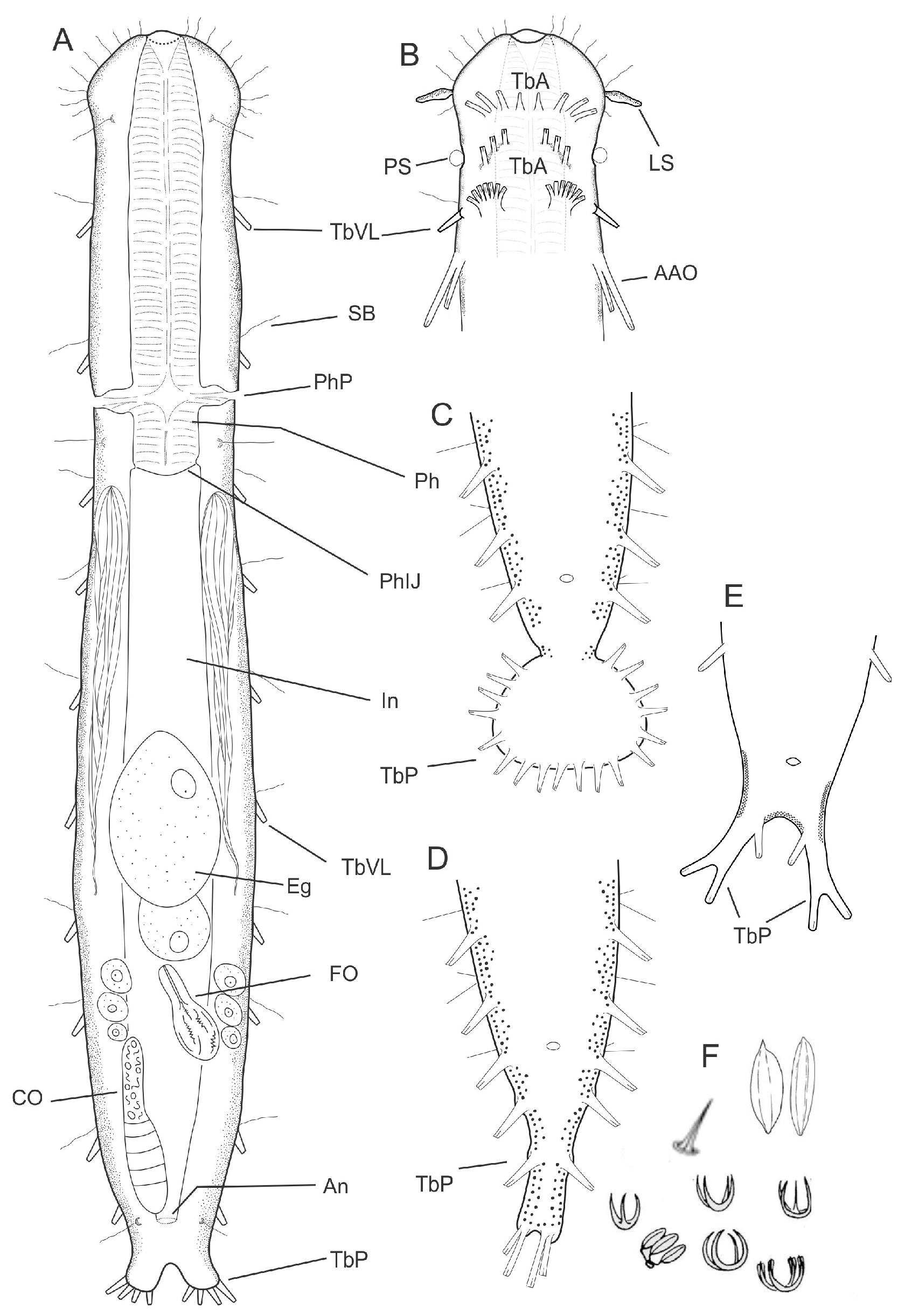
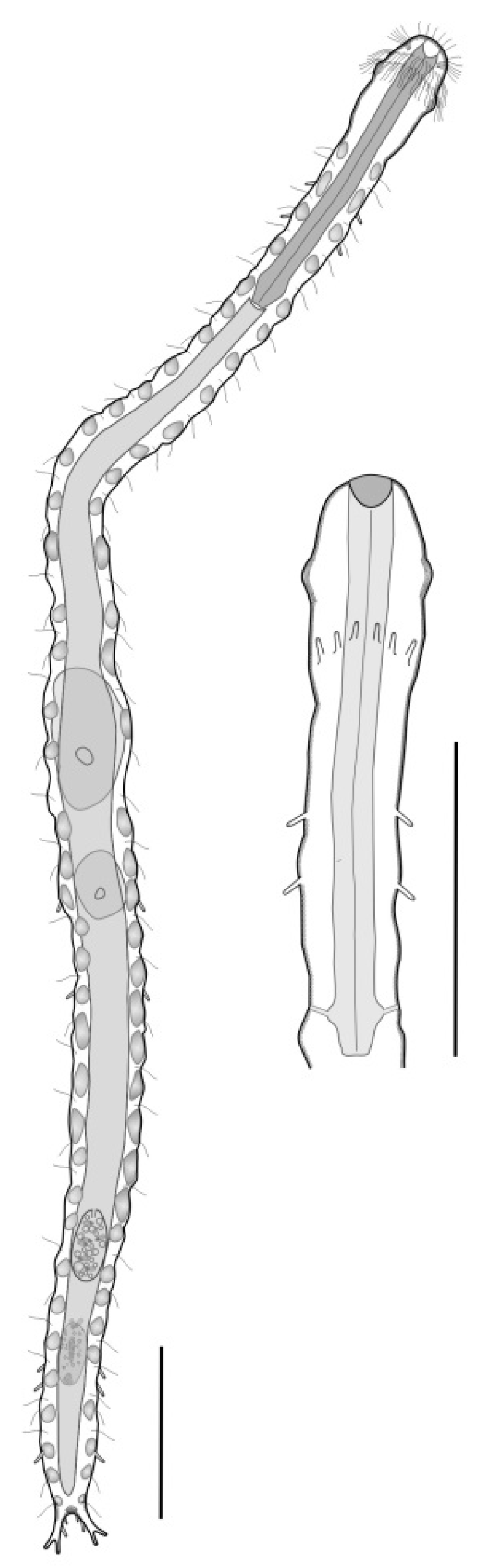
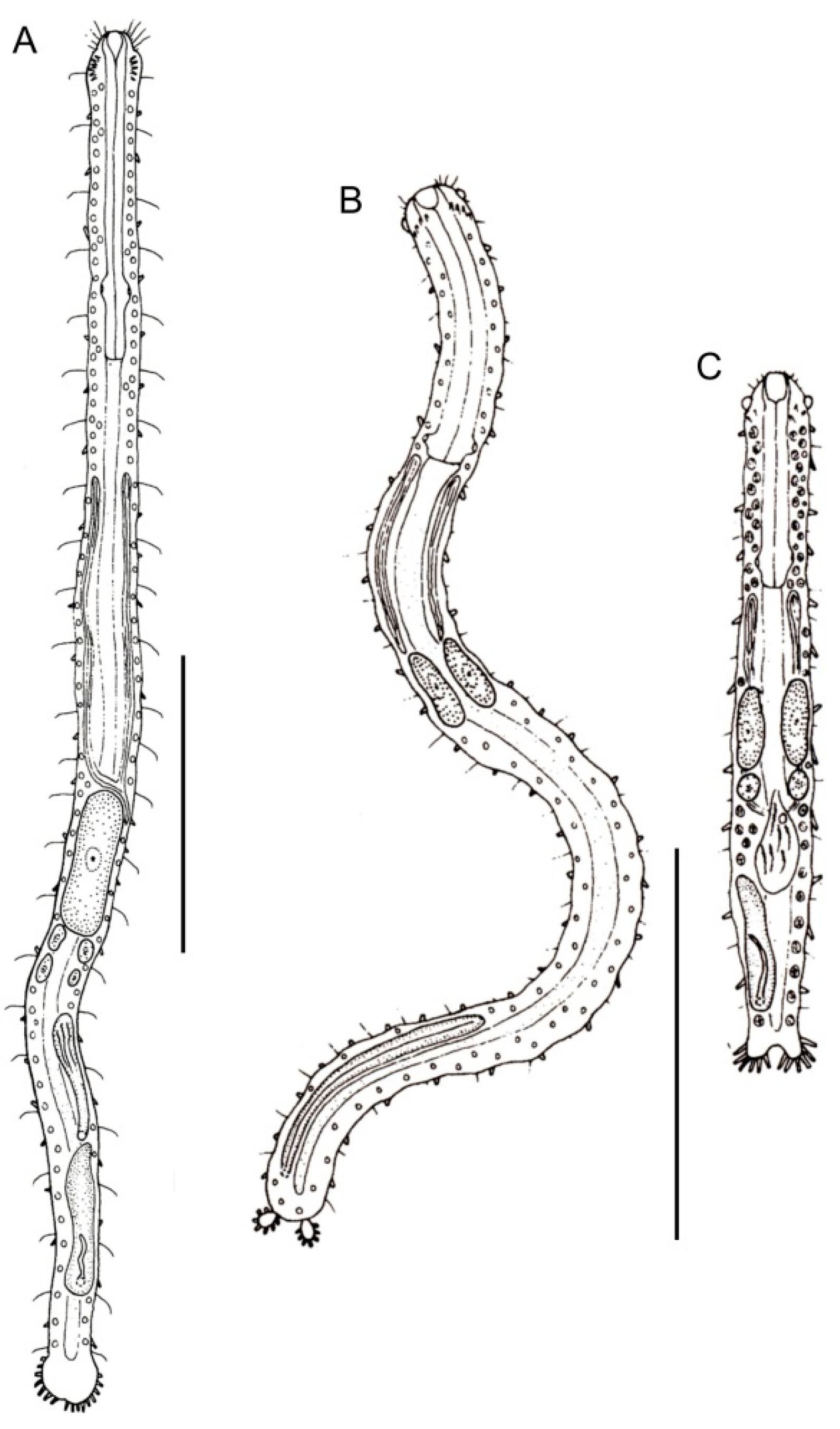
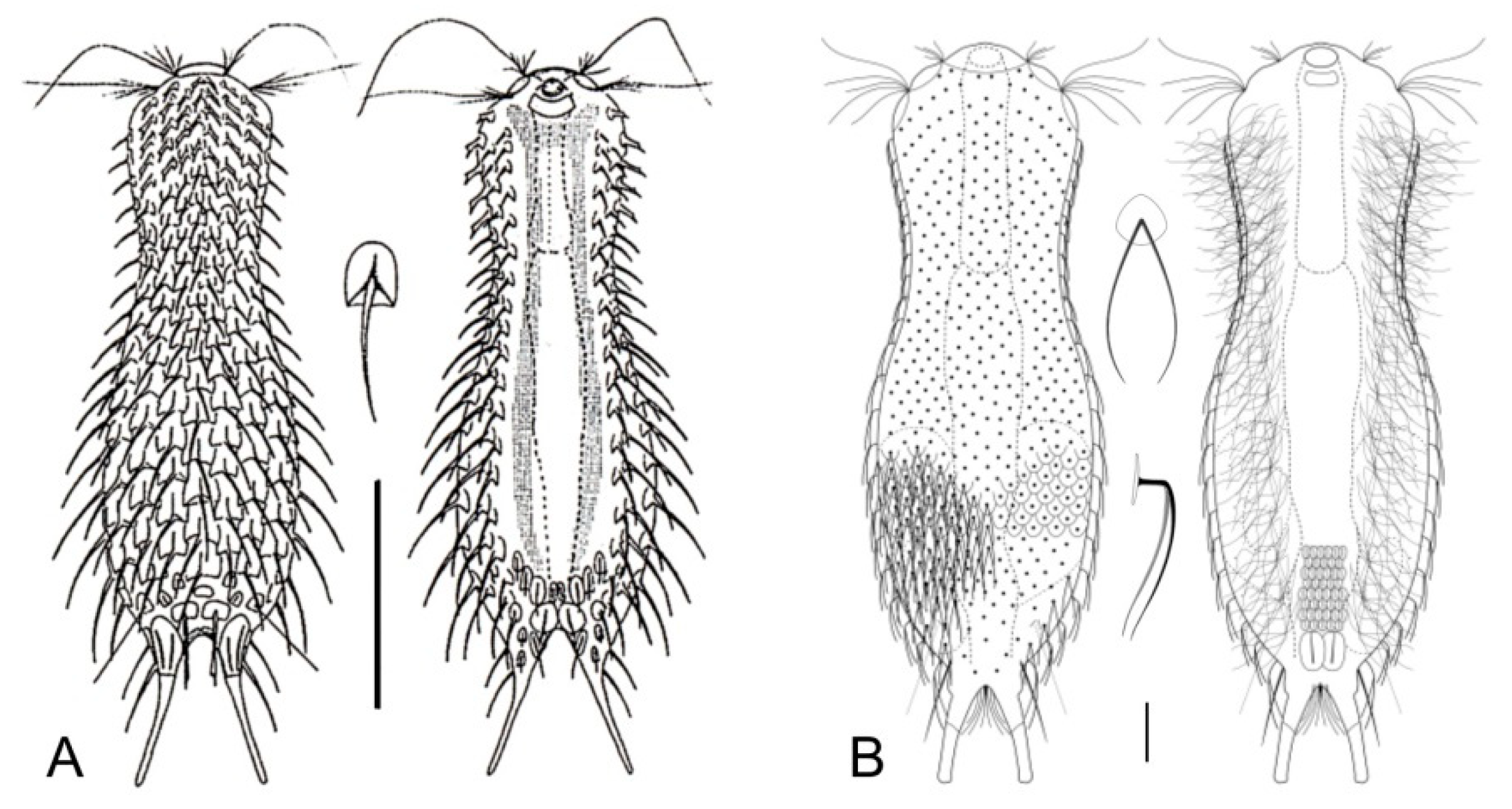
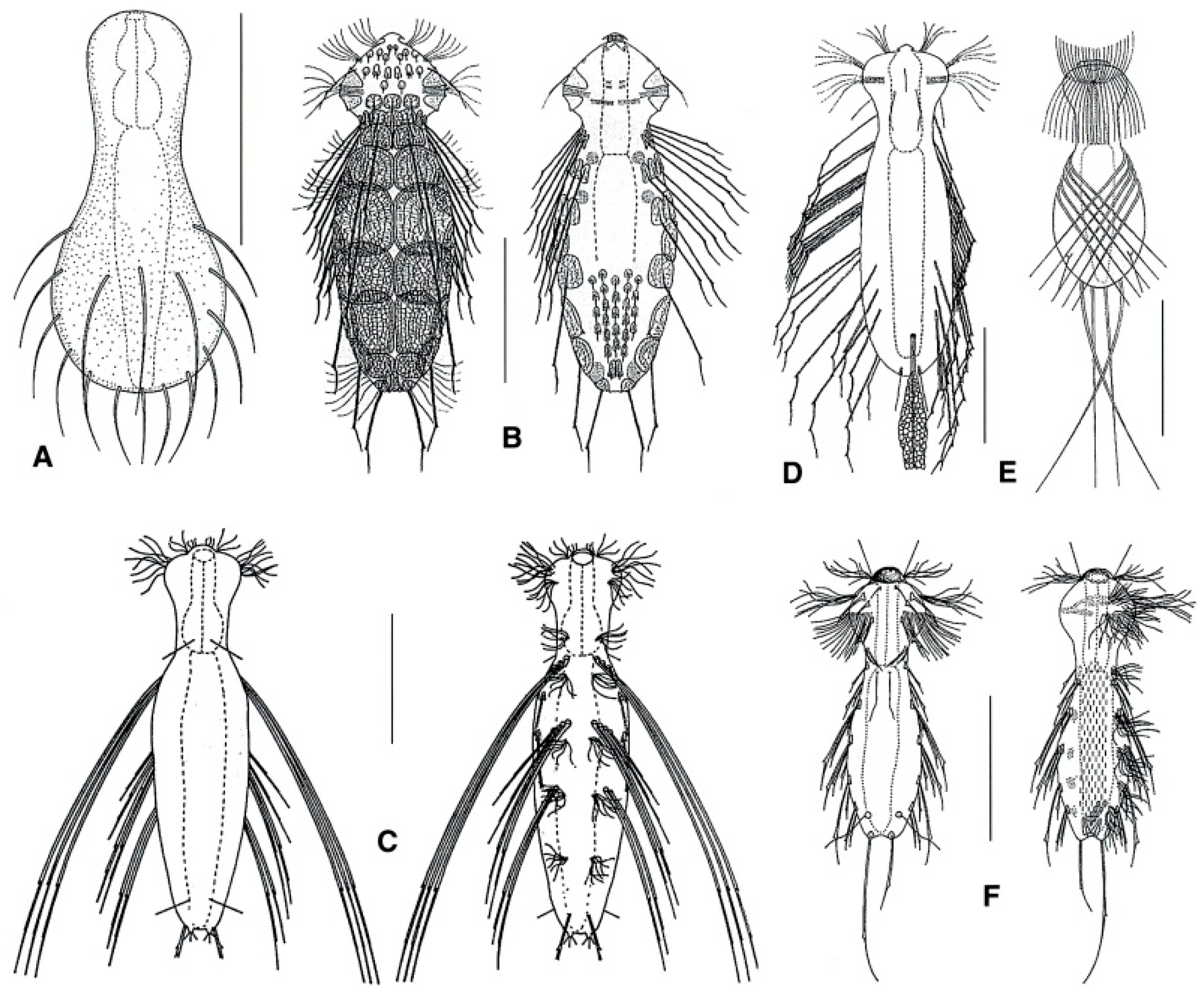
- Body usually vermiform, occasionally tenpin-shaped; cuticle naked or forming ornamentations such as plates and multi-pointed hooks; TbA, TbL, and TbP present, usually numerous; TbD present in several taxa; mouth opening narrow to broad; pharyngeal pores usually present. Marine and brackish, rarely fresh water: interstitial. Order MACRODASYIDA (Figure 2) ........... 3

- 3a (2b)
- Marine or brackish. ...……………………...……….……………………………………………… 4
- 3b (2b)
- Freshwater. ...…………...…………………....……………….......…………………....……..……37
- 4a (3a)
- Body tenpin-shaped; head well discernible, including most of pharynx; TbD absent; posterior body region lobed, furcate, or bifurcate. Cuticle bare or developing thickenings and ridges. ..……………………………………………………………………………...……...… 5
- 4b (3a)
- Body vermiform, head usually indistinct or, when distinct, includes only part of pharynx; cuticle naked or developing spines and/or scales. …...……….………………….…….....…... 9
- 5a (4a)
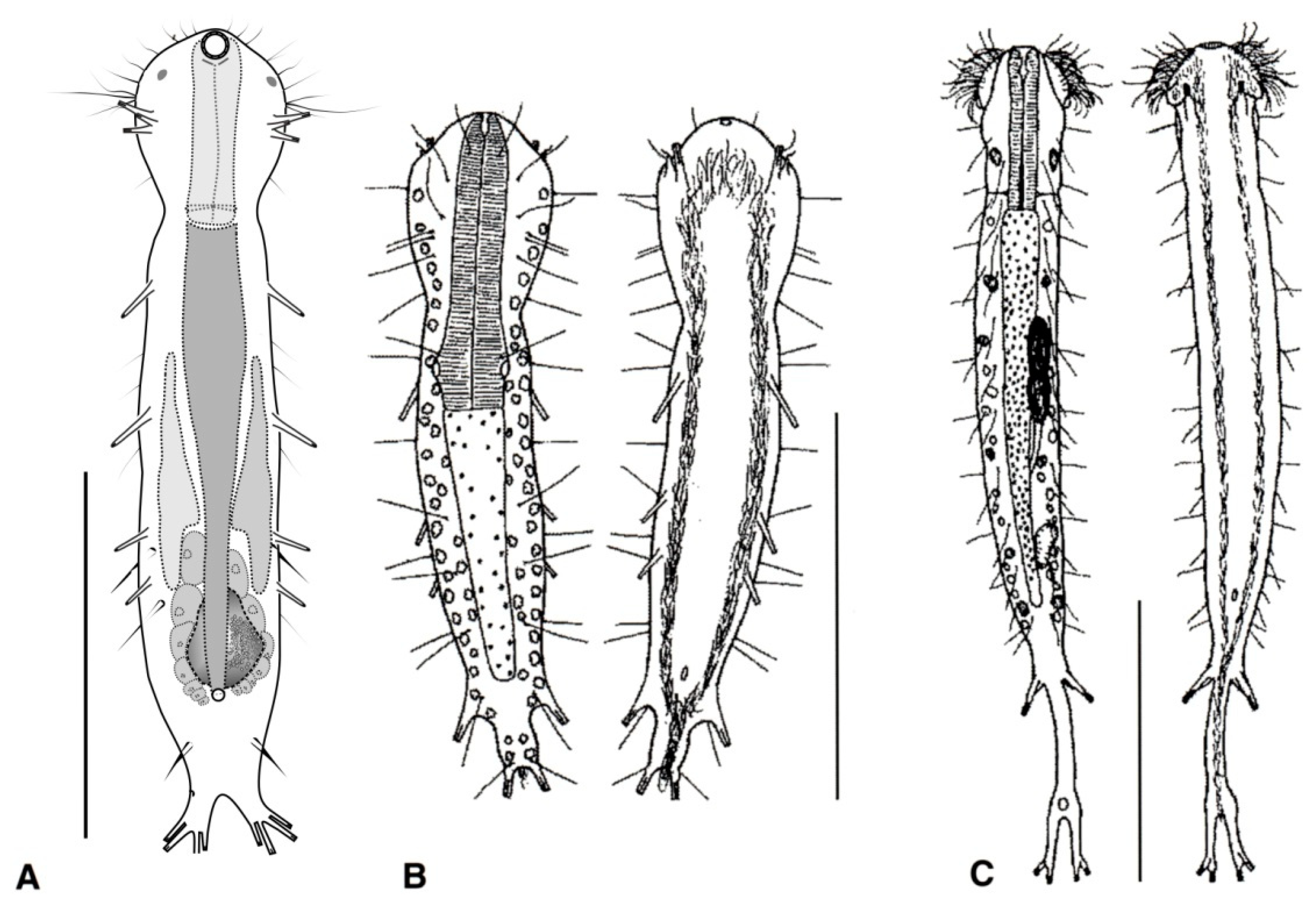
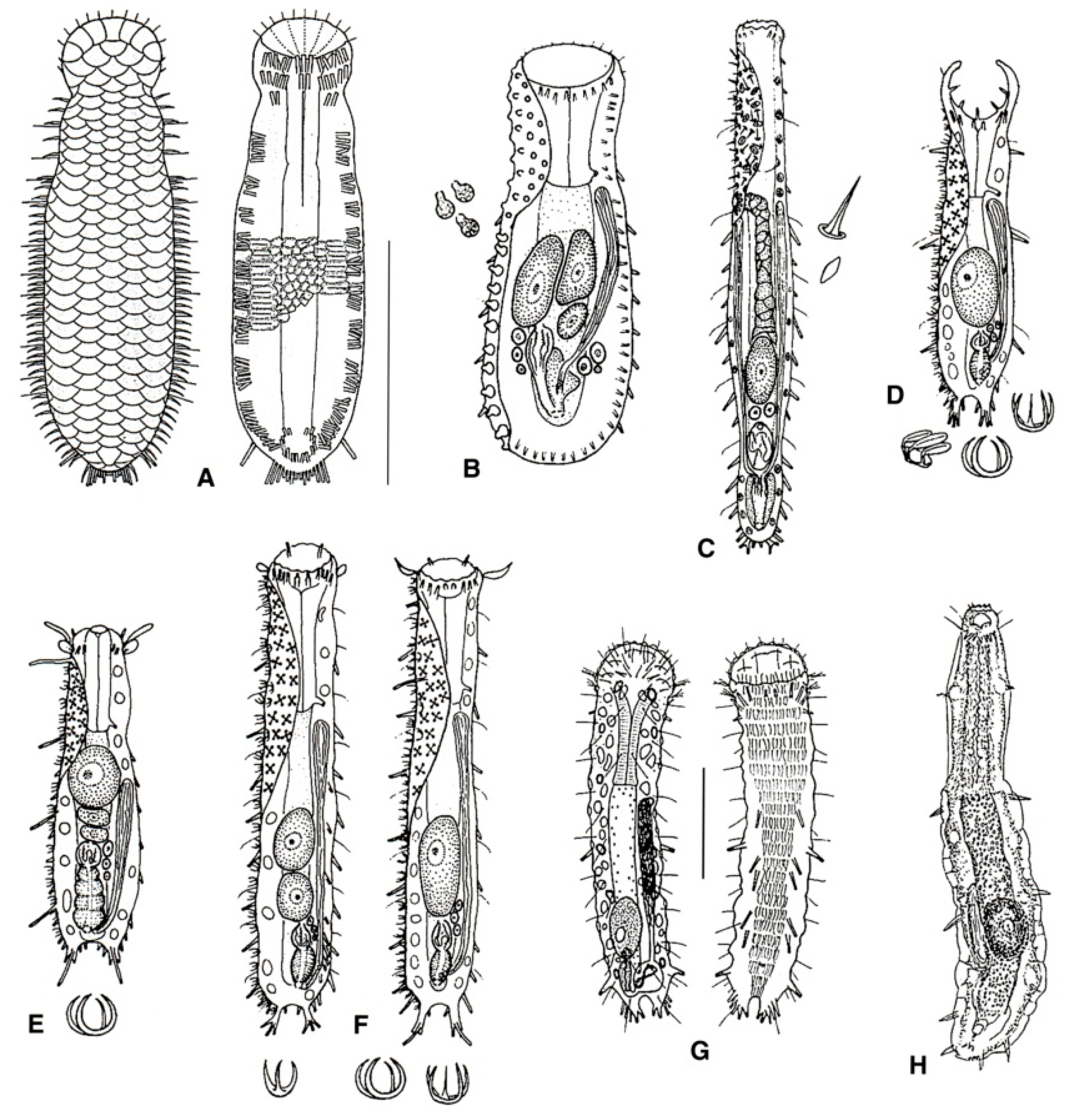
- Cuticle naked; dorsal side of the trunk naked; chordoid organ not present. Common to rare; marine and brackish: interstitial. DACTYLOPODOLIDAE (Figure 3) ….………..…...……. 6
- 5b (4a)
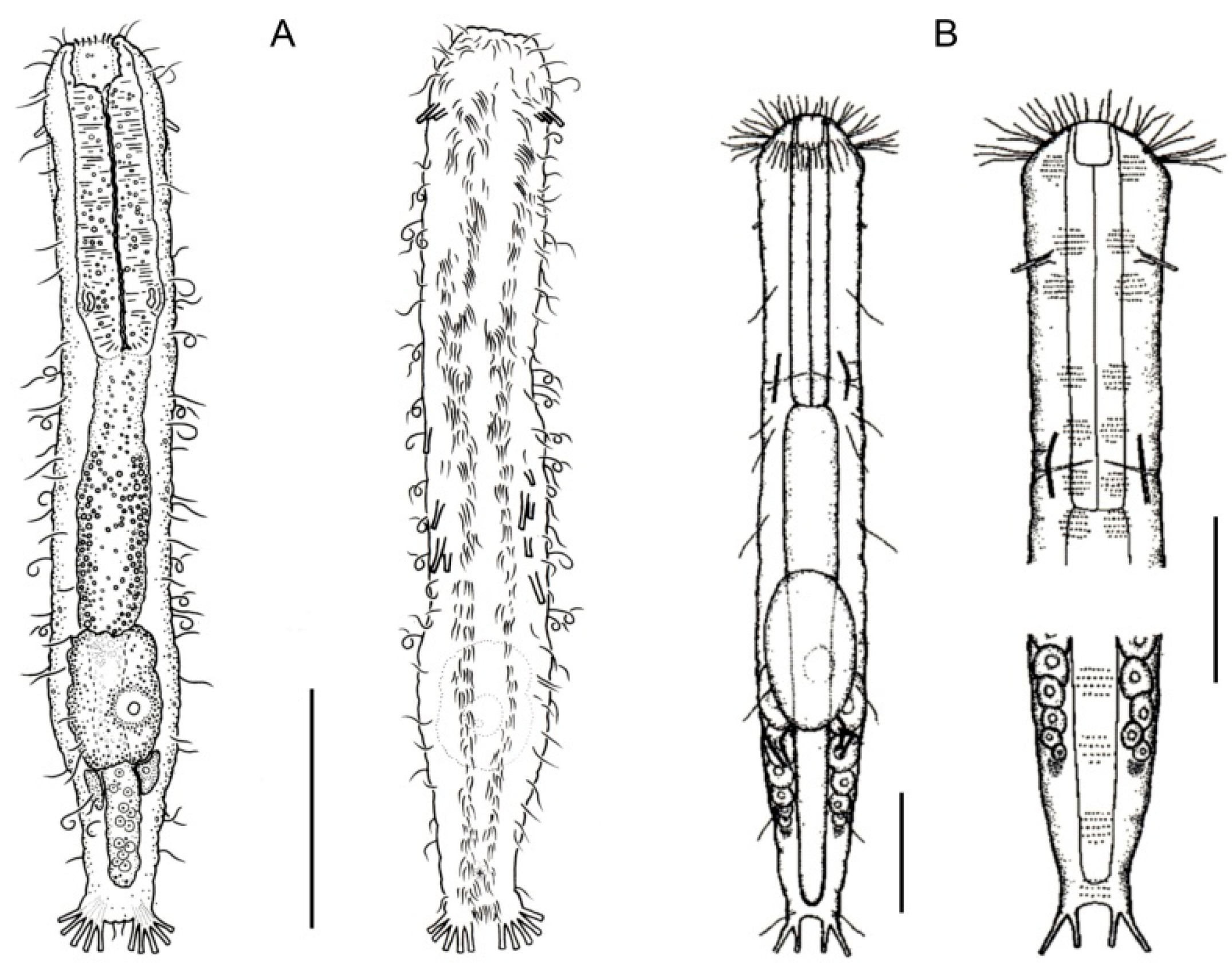
- Cuticle often developing thickenings and ridges; if naked, the trunk bears, on the dorsal side, long rod-like structures; chordoid organ present. Rare; marine: interstitial. XENODASYIDAE (Figure 4) ……………………………………..………………….…....…..… 8

- 6a (5a)
- Head simple or bearing two sensorial tentacles; cuticular covering bare; posterior body region bilobed; TbL present. Regionally common; marine: interstitial. .......... Dactylopodola (Figure 3A)
- 6b (5a)
- Head simple or with crenulated lateral lobes; cuticular covering bare; posterior body region bifurcate; TbL absent. ………………………….………………………………………………….. 7
- 7a (6b)
- Head simple, cuticle naked. Rare; marine: interstitial. …………….. Dendropodola (Figure 3B)
- 7b (6b)
- Head with elongate crenulated lateral lobes. Uncommon; marine: interstitial. ………………………………………………………………………………. Dendrodasys (Figure 3C)
- 8a (5b)
- Trunk region without tentacles, but presenting dented lateral sides; posterior body region furcate; distal rami, each showing a small TbP. Rare; marine: interstitial. ......................................................................................................................... Xenodasys (Figure 4A)
- 8b (5b)
- Trunk region bearing numerous tentacles; lateral sides of the trunk region parallel, lacking indentations; posterior region furcate; each ramus showing an adhesive pad at the end. Rare; marine: interstitial. ………………………………………..………........ Chordodasiopsis (Figure 4B)

- 9a (4b)
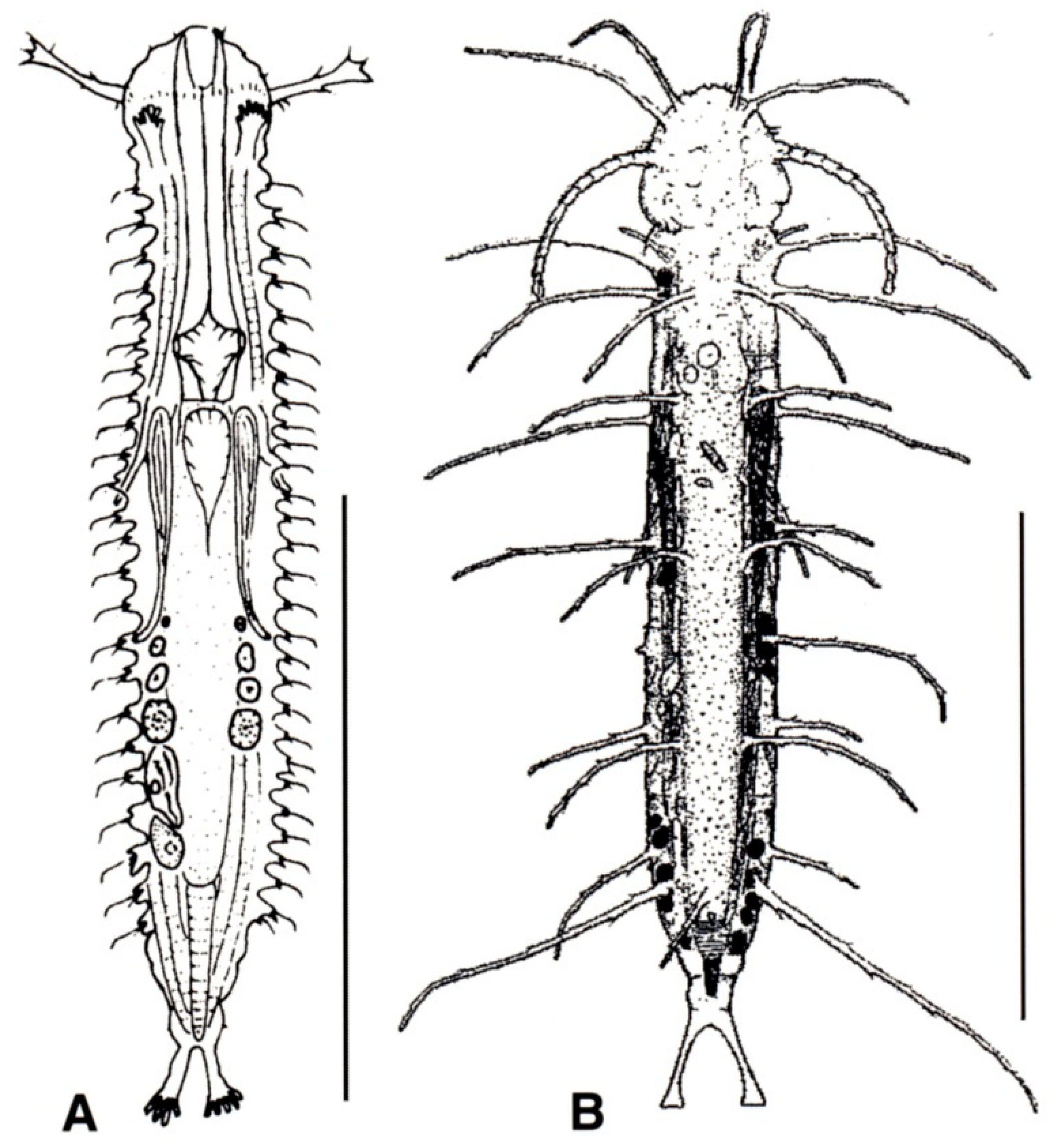
- TbA, usually 4 or more per side, occasionally 2 or 3, at the end of extensible fleshy base (Figure 2B); pharynx with pores located at the base. ................................................................. 10
- 9b (4b)
- TbA, generally 1 to 3 per side, occasionally 4 or more, arising singly and directly from the body surface; pharynx with pores at the base or in the middle. .............................................. 16
- 10a (9a)
- Head generally well demarcated posteriorly by a furrow; posterior body region tapered into a medial process, truncated, rounded, or broadly expanded, but never two-lobed. CEPHALODASYIDAE (partim) (Figure 5) .................................................................................. 11
- 10b (9a)
- Head normally not clearly delimited; posterior body region two-lobed. TURBANELLIDAE (partim) (Figure 6) ........................................................................................................................... 12

- 11a (10a)
- Head surrounded by very thick and dense sensory cilia; a couple of accessory adhesive organs present near the PhIJ, laterally directed; each organ comprising 3–4 tubes of unequal length; a couple of club-shaped gravireceptor organs on the dorsal side of the posterior cephalic region may be present. Rare; marine: interstitial. ……...….. Pleurodasys (Figure 5B)
- 11b (10a)
- Cephalic sensory cilia and accessory adhesive organs described above are absent. Regionally common; marine and brackish: interstitial. ................... Cephalodasys (Figure 5A)
- 12a (10b)
- Head showing elongate lateral tentacles. …………………………………….….....………… 13
- 12b (10b)
- Head without tentacles, occasionally with conical lobes. ……………………...….......…… 14
- 13a (12a)
- TbL numerous. Uncommon; marine: interstitial. ……………..…..…. Dinodasys (Figure 6A)
- 13b (12a)
- TbL lacking, paired TbV inserted just past the PhIJ. Rare; marine: interstitial. ................. ............................................................................................................. Pseudoturbanella (Figure 6B)
- 14a (12b)
- Paired accessory adhesive organs in the anterior pharyngeal region; organs are posteriorly directed, and each is made up of two tubes of unequal lengths. Common; marine and brackish: interstitial. ........………….…..…………………..…..…… Paraturbanella (Figure 6C)
- 14b (12b)
- Accessory adhesive tubes described above are either absent or present in different body regions. ………………………………………………………………………………………...…... 15
- 15a (14b)
- Accessory adhesive tubes not present. Common; marine and brackish: interstitial. ………….……………………………………………………………….……Turbanella (Figure 6E)
- 15b (14b)
- Accessory adhesive tubes present, close to the PhIJ. Rare; marine: interstitial. …………………..…..………………...........................................…... Prostobuccantia (Figure 6D)

- 16a (9b)
- Pharynx with pores far from the base; posterior body region unilobed, ovoidal in shape, or tapering off. MACRODASYIDAE (Figure 7) …......................................................................…. 17
- 16b (9b)
- Pharynx with pores at the base; posterior end of body not tapering off ……………...….… 20
- 17a (16a)
- Head bearing a lateral leaf-like sensorial organ; posterior body region unilobed, ovoidal in shape. Rare; marine: interstitial. …….…………….…………………. Thaidasys (Figure 7B,C)
- 17b (16a)
- Head bearing lateral piston pit sensorial organs; posterior body region tapering into a medial process. ...……………………………………..…………………………………………… 18
- 18a (17b)
- Posterior process in the form of a long tail. Regionally common; marine: interstitial and epibenthic. …………………………………………….……………….……. Urodasys (Figure 7E)
- 18b (17b)
- Posterior process short or in the form of a short tail. ………………….……………….…… 19
- 19a (18b)
- Frontal organ posterior to the largest egg; spermatozoa filiform. Common; marine: interstitial. …..……………….…..……..………………………..…..…... Macrodasys (Figure 7D)
- 19b (18b)
- Frontal organ anterior to the largest egg; spermatozoa stout. Uncommon; marine: interstitial. ……………………...……..…………………………….…... Kryptodasys (Figure 7A)
- 20a (16b)
- Cuticle forming ornamentations such as hooks, papillae, scales, or thickenings. ..……..... 21
- 20b (16b)
- Cuticle smooth, without ornamentation such those reported above. ...……….……..….… 27

- 21a (20a)
- ) Presence of elongate scales; mouth narrow; pharynx without pores. Uncommon; marine: interstitial. LEPIDODASYIDAE ................................................…….... Lepidodasys (Figure 8)
- 21b (20a)
- Presence of variously spined hooks, large scales, or papillae; mouth opening generally broad; pharyngeal pores present. THAUMASTODERMATIDAE (partim) (Figure 9) ……………... 22
- 22a (21b)
- Presence of papillae or large scales. ...…………………..……………………….……………. 23
- 22b (21b)
- Presence of uni- or multi-spined hooks. ……………………….………………………….…. 24

- 23a (22a)
- Cuticle with large scales, but not papillae; on either side of the body a single row of wide spines present. Regionally common; marine: interstitial. ..........….. Diplodasys (Figure 9A)
- 23b (22a)
- Cuticle with papillae, but not scales or spines. Uncommon; marine: interstitial. ....................………………………...…………………………..……..….... Oregodasys (Figure 9B)
- 24a (22b)
- Cuticle with hooks showing a single spine; right and left testicles present; Common; marine: interstitial. .....…….……...…….…………………..……....… Acanthodasys (Figure 9C)
- 24b (22b)
- Cuticle with hooks showing more than one spine; a single testicle on the right-hand body side. ……………………………………………………………………………………………….... 25
- 25a (24b)
- Anterior body region showing conspicuous, grasping structures on either side of the mouth funnel (buccal palps); hooks bearing 5, 4, or 3 spines (penta-, tetra-, or triancres). Common; marine: interstitial. …………………………………….. Pseudostomella (Figure 9D)
- 25b (24b)
- Anterior body region without buccal palps; hooks showing 5, 4, 3, or 2 spines (penta-, tetra-, tri-, or biancres). ..…………………………………………..…………………………..…. 26
- 26a (25b)
- Head bearing two pairs of sensoria tentacles on the lateral sides; mouth narrow, hooks with four spines. Common; marine: interstitial. ………….….... Thaumastoderma (Figure 9E)
- 26b (25b)
- Head bearing no or one pair of sensorial tentacles on the lateral sides; hooks with 5, 4, 3, or 2 spines. Very common; marine: interstitial. ………............. Tetranchyroderma (Figure 9F)

- 27a (20b)
- Male apparatus absent (i.e., parthenogenetic); TbA, two groups of three tubes per side; TbL, four or five per side, TbP up to five per side. Rare; marine: interstitial. REDUDASYIDAE (partim) ……………………………………………………………….. Anandrodasys (Figure 10A)
- 27b (20b)
- These characteristics not combined. ..……………..……...………………..………………… 28

- 28a (27b)
- TbA, several to many, arranged in two tufts; TbL absent. Rare; marine: interstitial. TURBANELLIDAE (partim) …………………………..…………….. Desmodasys (Figure 6F)
- 28b (27b)
- TbA, few to many, but not arranged in tufts; TbL normally present or, if absent, then TbA few in number. …………....…………………………………………………..……….………….. 29
- 29a (28b)
- TbA, few; TbL few; body elongate (to about 1 mm in length) and narrow; posterior end in the form of two distinct pedicles. HUMMONDASYIDAE. …….... Hummondasys(Figure 11)
- 29b (28b)
- These characteristics not combined. ……………….…………………….…….……….…….. 30

- 30a (29b)
- TbA, few to many; TbL and TbP, numerous (more than 10 per side); mouth narrow (< 0.4 × head width); posterior body region in the form of a large round lobe or clearly two-lobed. PLANODASYIDAE (Figure 12) ……………………………………………..……..……...……….. 31
- 30b (29b)
- TbA or TbL numbering fewer than six tubes per side; oral opening narrow to broad, and if narrow, then posterior body region not clearly two-lobed. ...……………………………... 33
- 31a (30a)
- TbA, present in low numbers; body very long (up to 3.5 mm in length) and rather narrow; posterior body region ending as a large lobe. Uncommon; marine: interstitial. ……………………………………………………………………….…… Megadasys (Figure 12A)
- 31b (30a)
- Posterior body region distinctly two-lobed. ………………………..……..…………………. 32

- 32a (31b)
- Posterior lobes in the form of oval appendages; most anterior TbA arranged transversely; caudal organ elongate. Rare; marine: interstitial. .……………...…… Planodasys (Figure 12B)
- 32b (31b)
- Posterior lobes in the form of furcate extensions; most anterior TbA arranged longitudinally; caudal organ ovate. Uncommon; marine: interstitial. ……………………... Crasiella (Figure 12C)
- 33a (30b)
- Oral opening, narrow (< 0.4 × head width); right and left testicles present. CEPHALODASYIDAE (partim) (Figure 5) ………………………………….……………..…... 34
- 33b (30b)
- Oral opening broad (> 0.6 × head width) or, if narrow, leading to a large buccal cavity surrounded by an oral hood; a single testicle, on the right-hand side. THAUMASTODERMATIDAE (partim) (Figure 9) ….…………………………...………………. 36
- 34a (33a)
- Total body length > 1 mm; TbA, one per side; TbL in form of numerous papillae along the body sides. Uncommon; marine: interstitial. ……………………… Dolichodasys (Figure 5D)
- 34b (33a)
- Total body length < 1 mm. …………………………………..…………….…………………… 35
- 35a (34b)
- TbA, 1–4 tubes per side, arranged in two groups; TbL, 0–6 tubes per side. Uncommon; marine: interstitial. ....................................................................................... Paradasys (Figure 5E)
- 35b (34b)
- TbA, few to several per side; TbL, several to many. Common; marine: interstitial. ……………….....……………….......………………………………....…. Mesodasys (Figure 5C)
- 36a (33b)
- Oral opening, broad; locomotor cilia extending over the entire ventral surface; male genital pore not surrounded by cuticular plates. Common; marine: interstitial. ……….…………………………………………………………......… Ptychostomella(Figure 9G)
- 36b (33b)
- Oral opening, narrow, leading to a large buccal cavity covered by an oral hood; ventral locomotor cilia restricted to the pharyngeal region; male genital pore surrounded by cuticular plates. Very rare (possibly extinct); marine: interstitial …... Hemidasys (Figure 9H)
- 37a (3b)
- Total body length 300–400 μm; TbA, 1–2 per side; pharyngeal pores present. Rare; interstitial. REDUDASYIDAE (partim) .………………..…….……..… Redudasys (Figure 10B)
- 37b (3b)
- Body length up to 220 μm; TbA, one per side; pharyngeal pores absent. Rare; interstitial. INCERTAE SEDIS. …………………………..…….………………..…… Marinellina (Figure 13)

- 38a (1a)
- 38b (1a)
- Ventral locomotor ciliation formed by single cilia, occurring in longitudinal bands or tufts, never composed of cirri (Figure 1B). Freshwater, marine, and brackish. ............................... 41
- 39a (38a)
- Cirri of the head and pharyngeal regions of two different sizes, with 1–2 transverse rows of small and short cirri anteriorly followed by transverse rows of big and longer cirri; frontal portion of pharynx with a swelling (bulb). Common; marine and brackish: interstitial. ..……………………………………….…….....….. Heteroxenotrichula (Figure 14A)
- 39b (38a)
- Cirri, all of similar size, pharynx without anterior swelling (bulb). ...………………..…… 40
- 40a (39b)
- Ovary and eggs present, testicles and spermatozoa absent; head clearly distinct; scales on the dorsal side, flat; scales of the lateral mid-trunk, pedunculated; a pair of lateral spines at the base of the furcal branches. Common; marine: interstitial. ..... Draculiciteria (Figure 14B)
- 40b (39b)
- Testicles and spermatozoa present; head in general not clearly defined; scales of the lateral mid-trunk bearing a stalk or flat; if stalked, similar to the dorsal scales. Common; marine and brackish: interstitial. ………………..……………….…. Xenotrichula (Figure 14C)

- 41a (38b)
- Posterior body region furcate or bifurcate; caudal rami with or without TbP. ……..…..... 42
- 41b (38b)
- Posterior body region rounded or truncated; perhaps showing two caudal protuberances or spines. ………………………………………………………………………………………………... 60
- 42a (41a)
- Posterior body region bifurcate, bearing four TbP or two TbP and two spiniform cuticular processes; elsewhere, cuticle smooth, not forming scales or spines. Rare; freshwater: interstitial or periphytic/epibenthic. DICHAETURIDAE …………………….….….. Dichaetura (Figure 15A)
- 42b (41a)
- Posterior body region furcate; cuticle smooth or forming spines and/or scales. ..……...... 43
- 43a (42b)
- Body cuticle smooth; caudal rami with TbP, sickle-shaped; cilia of the head not arranged into tufts. Very rare; freshwater: semiplanktonic or hyperbenthic. PROICHTHYDIIDAE (Figure 15B,C) ……………………………………..…………………………….…………..…….. 44
- 43b (42b)
- Body cuticle generally forming spines and scales; caudal rami with or without TbP; if present, caudal rami and TbP generally straight, short to very long; cilia of the head emerging as tufts or forming a continuous band around the elongate, muzzle-like frontal end. .…………………………………………………………………………………………....…… 45
- 44a (43a)
- Cilia of the head arranged as a transverse row of small elements on the dorsal side; locomotor cilia limited to head and neck, emerging as separate tufts. Freshwater: hyperbenthic …………………………………………...…….………Proichthydium (Figure 15B)
- 44b (43a)
- Cilia of the head emerging mostly from the lateral sides as single, short to very long elements; locomotor cilia distributed in two bands that run from under the head to the posterior trunk region. Freshwater: semiplanktonic. …....................................…… Proichthydioides (Figure 15C)

- 45a (43b)
- 45b (43b)
- Cilia of the head organized in a band, encircling a muzzle-like frontal end; TbP numbering two or four. Uncommon to rare, marine: epibenthic or interstitial. MUSELLIFERIDAE (Figure 20) …………………………………………………………………………………………………………. 67
- 46a (45a)
- TbP at the end of the furcal rami absent. Rare; freshwater: epibenthic. …. Undula (Figure 16A)
- 46b (45a)
- TbP at the end of the furcal rami present. ...……………………………………....……...…... 47
- 47b (46b)
- Furcal base narrow (pedunculated); caudal rami segmented; cephalion and hypostomion extremely large; scales without a keel, notch, or spine. Rare; freshwater: epibenthic. …………………………………………………………...……………….. Cephalionotus (Figure 16B)
- 47b (46b)
- These characteristics not combined. ……………………………………….…..……...……… 48
- 48a (47b)
- Furcal rami very long (up to one-third of the total body length), multi-segmented, bare or with tiny spines or scales. Common, freshwater: periphytic and epibenthic. ………………………………………………………………………...… Polymerurus (Figure 16C)
- 48b (47b)
- Furcal rami from very short to mid length, not segmented, scales or spines limited to the proximal portion or lacking altogether. ………………………………………..…..………….... 49
- 49a (48b)
- Cuticular covering bare (or mostly bare) or made up of scales lacking spines; seldomly, some spines may be present at the base of the furca. .……………..…………....…………..… 50
- 49b (48b)
- Cuticular covering including scales bearing spines (spined scales) and/or a keel (spined, keeled scales and keeled scales, respectively); spines from long to very short, bearing 1–2 indentations laterally (notched spines), or simple. ………………………………………………... 57

- 50a (49a)
- Cuticular covering bare, rarely a few scales and/or spines at base of the furca may be present. …………………………………………………………………………………………...... 51
- 50b (49a)
- Cuticular covering wholly or prevalently made of spineless scales. ….…….……...……... 53
- 51a (50a)
- Cuticle completely bare, very thick, obviously distinguishable from the underlying epidermal layer. Rare; freshwater: interstitial. ……...……...………..…. Arenotus (Figure 16D)
- 51b (50a)
- Cuticle thin, mostly bare, except for perhaps two terminal scales at the end ventral interciliary field; occasionally, weak striations along the body or few spines and/or scales at the furcal base may be present. Common; freshwater, rarely marine or brackish water: periphytic, epibenthic, and interstitial. …………………………………………………………………………………..…… 52
- 52a (51b)
- Furcal base pedunculated; locomotor cilia distributed in separated tufts. Uncommon; marine: interstitial. ………….……..………………..………....…. Caudichthydium (Figure 16E)
- 52b (51b)
- Furcal base not pedunculated; locomotor cilia mostly forming two longitudinal bands. Common; freshwater, rarely brackish or marine: epibenthic, periphytic, and interstitial. ………………………………………………………………….……….…Ichthydium (Figure 16F)
- 53a (50b)
- Scales, small, keeled, or stalked. ………………………………………………………………. 54
- 53b (50b)
- Scales, large and bare, round, rhomboidal, or polygonal in shape. ..………………….…... 55
- 54a (53a)
- Most scales with a stalk; occasionally, few scales may lack a stalk and bear a keel or a spine instead. Common; freshwater, brackish, and marine: epibenthic, periphytic, and interstitial. ……………………………………………………………………...…….. Aspidiophorus (Figure 16G)
- 54b (53a)
- Numerous keeled scales; occasionally, few scales may bear a spine. Common; freshwater, brackish, and marine: periphytic, epibenthic, and interstitial. ...Heterolepidoderma (Figure 16H)
- 55a (53b)
- Scales polygonal in shape. Common; freshwater, rarely brackish or marine: interstitial, epibenthic, and periphytic. ……………………………..……….....Lepidodermella (Figure 16I)
- 55b (53b)
- Scales rhomboidal or circular in shape. ………………………….………………………...… 56

- 56a (55b)
- Scales circular. Rare; freshwater: periphytic. ..................................... Fluxiderma (Figure 16L)
- 56b (55b)
- Scales rhomboidal. Rare; freshwater: periphytic. ................. Rhomballichthys (Figure 16M)
- 57a (49b)
- Scales of the ventral interciliary field similar in shape to the scales of the dorsal side; scales of the dorsal side possessing a double edge anteriorly, with or without a spine but always deprived of a keel; several pairs of thin spines of increasing length at the lateral sides of the furcal base. Rather common; freshwater: periphytic and epibenthic. ……………………………………………………………………….... Lepidochaetus (Figure 16N)
- 57b (49b)
- Scales of the ventral interciliary field dissimilar in shape from scales of the dorsal side; scales of the dorsal side with a single edge anteriorly, keeled or keeled and spined…………………... 58
- 58a (57b)
- Scales lateral to the ventral locomotor cilia with spines bearing lamellae (hydrofoil scales); scales of the dorsal side bearing a keel; seldom presence of 1–5 scales bearing spines. Common; marine and brackish: interstitial. ……………….... Halichaetonotus (Figure 16O)
- 58b (57b)
- Scales bearing spines with lamellae normally absent; if present, dorsal scales spined. .…..…. 59
- 59a (58b)
- Dorsal scales round to suboval, without keels and/or notches but carrying distally bifurcating hairlike spines. Rare; freshwater: epibenthic……...…. Bifidochaetus (Figure 17B)
- 59b (58b)
- These characteristics not combined. Very common; freshwater, marine, and brackish: epibenthic, periphytic, and interstitial. ………...……………....…… Chaetonotus (Figure 17A)
- 60a (41b)
- Posterior body region rounded-off or truncated with paired lateral projections; head bearing a pair of rod- or club-shaped tentacles; trunk bearing small, spined scales; rarely, trunk scales restricted to a small patch on the ventral side. Uncommon to rare; freshwater: hyperbenthic and semiplanktonic. NEOGOSSEIDAE (Figure 18) ………….…….…………. 61
- 60b (41b)
- Posterior body region rounded or truncated, occasionally with a very short caudal lobe or paired postero-lateral protuberance; head without tentacles; body scales reduced or absent; trunk bearing very long and movable spines arranged into groups. Uncommon to rare; freshwater: hyperbenthic, epibenthic, and semiplanktonic. DASYDYTIDAE (Figure 19) ….…. 62

- 61a (60a)
- Posterior body region truncated, showing two lateral projections bearing a tuft of long spines; trunk with fine spined scales. Uncommon: epibenthic and semiplanktonic. ...................................................................................................................... Neogossea (Figure 18A)
- 61b (60a)
- Posterior body region rounded, with a central group of spines and no lateral projections; trunk with keeled scales, seldom reduced to a small group on the ventral side. Rare: epibenthic and semiplanktonic. …............................……………………..…………….…. Kijanebalola (Figure 18B)
- 62a (60b)
- Trunk region bearing long, scattered spines on the dorsal side or two caudal spines only; body scales absent; locomotor cilia arranged in two longitudinal bands; pharynx bearing two robust swellings (bulbs). Rare: epibenthic and semiplanktonic. …………………………………………...……………………….... Anacanthoderma (Figure 19A)
- 62b (60b
- ) Trunk region bearing long, lateral spines arranged into columns or groups; dorsal spines present or absent; locomotor cilia arranged in tufts; pharynx bearing a single swelling or cylindrical. ……………………………………………………………………………….…...…… 63
- 63a (62b)
- Lateral spines, simple or with notches; if present, scales large, elliptical in shape, and few in number; pharynx cylindrical (i.e., without bulbs). …………….……………...………….… 64
- 63b (62b)
- Lateral spines with a single lateral notch and bifurcate apex, or with 2–3 lateral notches and pointed apex; if present, numerous, small, keeled scales; pharynx bearing a swelling at the posterior end. ………………………………………………………………………………………… 65

- 64a (63a)
- Trunk showing dorsal spines; two pairs of caudal spines; all spines show a noticeable lateral notch; dorsal scales, rather large and of peculiar lace-like appearance. Rare: epibenthic, periphytic, and semiplanktonic. ……………….……... Ornamentula (Figure 19B)
- 64b (63a)
- Trunk lacking dorsal spines; a single pair of caudal spines or none; if very long, the lateral spines are thick and bent basally, becoming thinner and thinner distally; lateral notch present or absent; where present, body scales are small and feebly keeled. ………...……... 66
- 65a (65b)
- Lateral spines, robust, showing pointed tip and 2–3 lateral notches; body scales lacking; posterior body region showing two bristled protuberances on the sides. Uncommon: semiplanktonic and epibenthic.……………………..…….……………Stylochaeta (Figure 19C)
- 65b (63b)
- Lateral spines, almost straight, showing a bifurcate tip and a single lateral notch; body scales present; posterior body region rounded. Uncommon: semiplanktonic, periphytic, and epibenthic ………………………….………………………………...……Dasydytes (Figure 19D)
- 66a (63b)
- Caudal spines absent or present; if present, in general of different length; lateral spines, straight, of medium length; ventral, S-shaped, jumping spines lacking. Rare: semiplanktonic and epibenthic. ......................…………………….......................................... Setopus (Figure 19F)
- 66b (64b)
- Caudal spines absent; lateral spines very long, strongly bent crossing over the dorsal side; ventral S-shaped jumping spines present. Rare: semiplanktonic and epibenthic. ……………………………………………………………….………….…. Haltidytes (Figure 19E)
- 67a (45b)
- Furcal rami each with a single TbP; body scales bearing fine spines but lacking keels. Rare; marine: interstitial or infaunal. Uncommon: epibenthic and interstitial. .……………………………………………………………………..………. Musellifer (Figure 20A)
- 67b (45b)
- Furcal rami each with two TbP; body scales, keeled. Rare: interstitial. ...................................................................................................................... Diuronotus (Figure 20B)

4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Todaro, M.A. Meiofauna from the Meloria Shoals: Gastrotricha, biodiversity and seasonal dynamics. Biol. Mar. Medit. 1998, 5, 587–590. [Google Scholar]
- Gray, J.S. The effects of pollution on sand meiofauna communities. Thalass. Jugosl. 1971, 7, 79–86. [Google Scholar]
- Coull, B.S. Long-term variability of estuarine meiobenthos: An 11 year study. Mar. Ecol. Prog. Ser. 1985, 24, 205–218. [Google Scholar] [CrossRef]
- Todaro, M.A.; Fleeger, J.W.; Hummon, W.D. Marine gastrotrichs from the sand beaches of the northern Gulf of Mexico: Species list and distribution. Hydrobiologia 1995, 310, 107–117. [Google Scholar] [CrossRef]
- Hochberg, R. Spatiotemporal size-class distribution of Turbanella mustela (Gastrotricha: Macrodasyida) on a northern California beach and its effect on tidal suspension. Pacific Sci. 1999, 53, 50–60. [Google Scholar]
- Nesteruk, T. Density and biomass of Gastrotricha in sediments of different types of standing waters. Hydrobiologia 1996, 24, 205–208. [Google Scholar] [CrossRef]
- Ruppert, E.E. Gastrotricha. In Microscopic Anatomy of Invertebrates, Aschelminthes; Harrison, F.W., Ruppert, E.E., Eds.; Wiley-Liss: New York, NY, USA, 1991; Volume 4, pp. 41–109. [Google Scholar]
- Hochberg, R.; Litvaitis, M.K. A muscular double helix in gastrotricha. Zool. Anz. 2001, 240, 61–68. [Google Scholar] [CrossRef]
- Todaro, M.A.; Telford, M.J.; Lockyer, A.E.; Littlewood, D.T.J. Interrelationships of the Gastrotricha and their place among the Metazoa inferred from 18S rRNA genes. Zool. Scr. 2006, 35, 251–259. [Google Scholar] [CrossRef]
- Struck, T.H.; Wey-Fabrizius, A.R.; Golombek, A.; Hering, L.; Weigert, A.; Bleidorn, C.; Klebow, S.; Iakovenko, N.; Hausdorf, B.; Petersen, M.; et al. Platyzoan paraphyly based on phylogenomic data supports a noncoelomate ancestry of Spiralia. Mol. Biol. Evol. 2014, 31, 1833–1849. [Google Scholar] [CrossRef]
- Egger, B.; Lapraz, F.; Müller, S.; Dessimoz, C.; Girstmair, J.; Skunca, N.; Rawlinson, K.A.; Cameron, C.B.; Beli, E.; Todaro, M.A.; et al. A Transcriptomic-phylogenomic analysis of the evolutionary relationships of flatworms. Curr. Biol. 2015, 25, 1–7. [Google Scholar] [CrossRef]
- Todaro, M.A.; Dal Zotto, M.; Jondelius, U.; Hochberg, R.; Hummon, W.D.; Kånneby, T.; Rocha, C.E.F. Gastrotricha: a marine sister for a freshwater puzzle. PLoS ONE 2012, 7, e31740. [Google Scholar] [CrossRef] [PubMed]
- Todaro, M.A.; Leasi, F.; Hochberg, R. A new species, genus and family of marine Gastrotricha from Jamaica, with a phylogenetic analysis of Macrodasyida based on molecular data. Syst. Biodiv. 2014, 12, 473–488. [Google Scholar] [CrossRef]
- Todaro, M.A.; Dal Zotto, M.; Leasi, F. An integrated morphological and molecular approach to the description and systematisation of a novel genus and species of Macrodasyida (Gastrotricha). PLoS ONE 2015, 10, e0130278. [Google Scholar] [CrossRef] [PubMed]
- Todaro, M.A.; Dal Zotto, M.; Kånneby, T.; Hochberg, R. Integrated data analysis allows the establishment of a new, cosmopolitan genus of marine Macrodasyida (Gastrotricha). Sci. Rep. 2019, 9, 7989. [Google Scholar] [CrossRef] [PubMed]
- Leasi, F.; Todaro, M.A. The muscular system of Musellifer delamarei (Renaud-Mornant, 1968) and other chaetonotidans with implications for the phylogeny and systematisation of the Paucitubulatina (Gastrotricha). Biol. J. Linn. Soc. 2008, 94, 379–398. [Google Scholar] [CrossRef]
- Kånneby, T.; Todaro, M.A.; Jondelius, U. Phylogeny of Chaetonotidae and other Paucitubulatina (Gastrotricha: Chaetonotida) and the colonization of aquatic ecosystems. Zool. Scr. 2013, 42, 88–105. [Google Scholar] [CrossRef]
- Guidi, L.; Todaro, M.A.; Ferraguti, M.; Balsamo, M. Reproductive system and spermatozoa ultrastructure support the phylogenetic proximity of Megadasys and Crasiella (Gastrotricha, Macrodasyida). Contr. Zool. 2014, 83, 119–131. [Google Scholar] [CrossRef]
- Kånneby, T.; Todaro, M.A. The phylogenetic position of Neogosseidae (Gastrotricha: Chaetonotida) and the origin of planktonic Gastrotricha. Org. Divers. Evol. 2015, 6, 1–12. [Google Scholar]
- Balsamo, M.; Grilli, P.; Guidi, L.; d’Hondt, J.L. Gastrotricha: Biology, ecology and systematics. Families Dasydytidae, Dichaeturidae, Neogosseidae, Proichthydiidae. In Identification Guides to the Plankton and Benthos of Inland Waters; Dumont, H.J.F., Ed.; Backhuys Publisher: Leiden, The Netherlands, 2014; Volume 24, pp. 1–187. [Google Scholar]
- Kånneby, T.; Hochberg, R. Phylum Gastrotricha. In Thorp and Covich’s Freshwater Invertebrates: Ecology and General Biology; Thorp, J., Rogers, D.C., Eds.; Elsevie Academic Press: Amsterdam, The Netherlands, 2015; Volume 1, pp. 211–223. [Google Scholar]
- Kieneke, A.; Schmidt-Rhaesa, A. Gastrotricha and Gnathifera. In Handbook of Zoology; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany, 2015; Volume 3, pp. 1–134. [Google Scholar]
- Gastrotricha Web Portal. Available online: http://www.gastrotricha.unimore.it/ (accessed on 25 June 2019).
- World Register of Marine Species (WoRMS). Available online: http://www.marinespecies.org/ (accessed on 25 June 2019).
- Hummon, W.D.; Todaro, M.A.; Balsamo, M.; Tongiorgi, P. Effects of pollution on marine Gastrotricha in the northwestern Adriatic Sea. Mar. Pollut. Bull. 1990, 21, 241–243. [Google Scholar] [CrossRef]
- Todaro, M.A.; Rocha, C.E.F. Diversity and distribution of marine Gastrotricha along the northern beaches of the state of Sao Paulo (Brazil), with description of a new species of Macrodasys (Macrodasyida, Macrodasyidae). J. Nat. Hist. 2004, 38, 1605–1634. [Google Scholar] [CrossRef]
- Todaro, M.A.; Rocha, C.E.F. Further data on marine gastrotrichs from the State of São Paulo and the first records from the State of Rio de Janeiro (Brazil). Meiofauna Mar. 2005, 14, 27–31. [Google Scholar]
- Hummon, W.D. Gastrotricha. In The Light and Smith Manual: Intertidal Invertebrates from Central California to Oregon; Carlton, J.T., Ed.; University of California Press: Berkeley, CA, USA, 2007; pp. 267–268. [Google Scholar]
- Todaro, M.A.; Leasi, F.; Bizzarri, N.; Tongiorgi, P. Meiofauna densities and gastrotrich community composition in a Mediterranean sea cave. Mar. Biol. 2006, 149, 1079–1091. [Google Scholar] [CrossRef]
- Sergeeva, N.G.; Ürkmez, D.; Todaro, M.A. Significant occurrence of Musellifer profundus Vivier, 1974 (Gastrotricha, Chaetonotida) in the Black Sea. Check List 2019, 15, 219–224. [Google Scholar] [CrossRef]
- Pfannkuche, O.; Thiel, H. Sampling processing. In Introduction to the Study of Meiofauna; Higgins, R.P., Thiel, H., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1988; pp. 134–145. [Google Scholar]
- Todaro, M.A. Contribution to the study of the Mediterranean meiofauna: Gastrotricha from the Island of Ponza, Italy. Boll. Zool. 1992, 59, 321–333. [Google Scholar] [CrossRef]
- Hochberg, R.; Litvaitis, M.K. Hexamethyldisilazane for scanning electron microscopy of Gastrotricha. Biotech. Histochem. 2000, 75, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Chang, C.Y. Two new marine gastrotrichs of the genus Ptychostomella (Macrodasyida, Thaumastodermatidae) from South Korea. Zool. Sci. 2003, 20, 481–489. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Todaro, M.A.; Hummon, W.D. An overview and a dichotomous key to genera of the phylum Gastrotricha. Meiofauna Mar. 2008, 16, 3–20. [Google Scholar]
- Kolicka, M.; Dabert, M.; Dabert, J.; Kånneby, T.; Kisielewski, J. Bifidochaetus, a new Arctic genus of freshwater Chaetonotida (Gastrotricha) from Spitsbergen revealed by an integrative taxonomic approach. Invert. Syst. 2016, 30, 398–419. [Google Scholar] [CrossRef]
- Garraffoni, A.R.S.; Araujo, T.Q.; Lourenço, A.P.; Guidi, L.; Balsamo, M. A new genus and new species of freshwater Chaetonotidae (Gastrotricha: Chaetonotida) from Brazil with phylogenetic position inferred from nuclear and mitochondrial DNA sequences. Syst. Biodiv. 2017, 15, 49–62. [Google Scholar] [CrossRef]
- Balsamo, M.; Todaro, M.A. Gastrotricha. In Freshwater Meiofauna Biology and Ecology; Rundle, S.D., Robertson, A.I., Schmid-Araya, J.M., Eds.; Backhuys Publisher: Leiden, The Netherlands, 2002; pp. 45–61. [Google Scholar]
- Pfannkuche, O. Gastrotricha. In Introduction to the Study of Meiofauna; Higgins, R.P., Thiel, H., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1988; pp. 302–311. [Google Scholar]
- Todaro, M.A.; Perissinotto, R.; Bownes, S.J. Two new marine Gastrotricha from the Indian Ocean coast of South Africa. Zootaxa 2015, 3905, 193–208. [Google Scholar] [CrossRef]
- Hummon, W.D.; Todaro, M.A.; Tongiorgi, P. Italian Marine Gastrotricha: II. One new genus and ten new species of Macrodasyida. Boll. Zool. 1993, 60, 109–127. [Google Scholar] [CrossRef]
- Hummon, W.D.; Todaro, M.A.; Tongiorgi, P.; Balsamo, M. Italian marine Gastrotricha: V. Four new and one redescribed species of Macrodasyida in the Dactylopodolidae and Thaumastodermatidae. Ital. J. Zool. 1998, 65, 109–119. [Google Scholar] [CrossRef]
- Rieger, R.M.; Ruppert, E.E.; Rieger, G.E.; Schoepfer-Sterrer, C. On the fine structure of gastrotrichs, with description of Chordodasys antennatus sp. n. Zool. Scr. 1974, 3, 219–237. [Google Scholar] [CrossRef]
- Hummon, W.D. Gastrotricha of the North Atlantic Ocean: 1. Twenty four new and two redescribed species of Macrodasyida. Meiofauna Mar. 2008, 16, 117–174. [Google Scholar]
- Todaro, M.A.; Dal Zotto, M.; Bownes, S.J.; Perissinotto, R. Two new interesting species of Macrodasyida (Gastrotricha) from KwaZulu-Natal (South Africa). Proc. Biol. Soc. Wash. 2017, 130, 139–154. [Google Scholar] [CrossRef]
- Evans, W.A.; Hummon, W.D. A new genus and species of Gastrotricha from the Atlantic coast of Florida, U.S.A. Trans. Am. Microsc. Soc. 1991, 110, 321–327. [Google Scholar] [CrossRef]
- Clausen, C. Gastrotricha Macrodasyida from the Tromsø region, northern Norway. Sarsia 2000, 85, 357–384. [Google Scholar] [CrossRef]
- Luporini, P.; Magagnini, G.; Tongiorgi, P. Contribution a la connaissance des gastrotriches des cotes de Toscane. Cah. Biol. Mar. 1971, 12, 433–455. [Google Scholar]
- Claparède, E. Miscellaneous zoologiques. III. Type d’un nouveau genere de gastrotriches. Ann. Sci. Nat. Zool. 1867, 8, 16–23. [Google Scholar]
- Kieneke, A.; Rothe, B.H.; Schmidt-Rhaesa, A. Record and description of Anandrodasys agadasys (Gastrotricha: Redudasyidae) from Lee Stocking Island (Bahamas), with remarks on populations from different geographic areas. Meiofauna Mar. 2013, 20, 39–48. [Google Scholar]
- Kisielewski, J. Two new interesting genera of Gastrotricha (Macrodasyida and Chaetonotida) from the Brazilian freshwater psammon. Hydrobiologia 1987, 153, 23–30. [Google Scholar] [CrossRef]
- Ruttner-Kolisko, A. Rheomorpha neiswestnovae und Marinellina flagellata, zwei phylogeneticsh interessante Wurmtypen aus dem Susswasserpsammon. Österr. Zool. Z. 1955, 6, 55–69. [Google Scholar]
- Luporini, P.; Magagnini, G.; Tongiorgi, P. Chaetonotoid gastrotrichs of the Tuscan Coast. Boll. Zool. 1973, 40, 31–40. [Google Scholar] [CrossRef]
- Balsamo, M. Gastrotrichi. In Guide C.N.R. per IL Riconoscimento Delle Specie Animali Delle Acque Interne Italiane; Consiglio Nazionale delle Ricerche AQ/1/199; CNR (Centro Nazionale Ricerche): Roma, Italy, 1983; Volume 7, pp. 547–571. [Google Scholar]
- Cordero, E.H. Notes sur les Gastrotriches. Physis 1918, 4, 241–244. [Google Scholar]
- Sudzuki, M. The Gastrotricha of Japan which live in the capillary water of the interstitial system: II. Bull. Biogeogr. Soc. Japan 1971, 27, 37–41. [Google Scholar]
- Kisielewski, J. Inland-water Gastrotricha from Brazil. Ann. Zool. (Warsaw) 1991, 43, 1–168. [Google Scholar]
- Mock, H. Chaetonotoidea (Gastrotricha) from the North Sea Island of Sylt. Mikrofauna. Meeres. 1980, 78, 1–107. [Google Scholar]
- Schwank, P. Gastrotricha und Nemertini. In Süsswasserfauna von Mittleuropas; Brauer, A., Ed.; G. Fisher Verlag: Stuttgart, Germany, 1990; Volume 3, pp. 1–252. [Google Scholar]
- Schrom, H. Nordadriatische Gastrotrichen. Helgoländer Wiss. Meeresunters. 1972, 23, 286–351. [Google Scholar] [CrossRef]
- Hummon, W.D.; Balsamo, M.; Todaro, M.A. Italian marine Gastrotricha: I. Six new and one redescribed species of Chaetonotida. Boll. Zool. 1992, 59, 499–516. [Google Scholar] [CrossRef]
- Kånneby, T. A redescription of Chaetonotus (Primochaetus) veronicae Kånneby, 2013 (Gastrotricha: Chaetonotidae). Zootaxa 2015, 4027, 442–446. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Todaro, M.A.; Sibaja-Cordero, J.A.; Segura-Bermúdez, O.A.; Coto-Delgado, G.; Goebel-Otárola, N.; Barquero, J.D.; Cullell-Delgado, M.; Dal Zotto, M. An Introduction to the Study of Gastrotricha, with a Taxonomic Key to Families and Genera of the Group. Diversity 2019, 11, 117. https://doi.org/10.3390/d11070117
Todaro MA, Sibaja-Cordero JA, Segura-Bermúdez OA, Coto-Delgado G, Goebel-Otárola N, Barquero JD, Cullell-Delgado M, Dal Zotto M. An Introduction to the Study of Gastrotricha, with a Taxonomic Key to Families and Genera of the Group. Diversity. 2019; 11(7):117. https://doi.org/10.3390/d11070117
Chicago/Turabian StyleTodaro, M. Antonio, Jeffrey Alejandro Sibaja-Cordero, Oscar A. Segura-Bermúdez, Génesis Coto-Delgado, Nathalie Goebel-Otárola, Juan D. Barquero, Mariana Cullell-Delgado, and Matteo Dal Zotto. 2019. "An Introduction to the Study of Gastrotricha, with a Taxonomic Key to Families and Genera of the Group" Diversity 11, no. 7: 117. https://doi.org/10.3390/d11070117
APA StyleTodaro, M. A., Sibaja-Cordero, J. A., Segura-Bermúdez, O. A., Coto-Delgado, G., Goebel-Otárola, N., Barquero, J. D., Cullell-Delgado, M., & Dal Zotto, M. (2019). An Introduction to the Study of Gastrotricha, with a Taxonomic Key to Families and Genera of the Group. Diversity, 11(7), 117. https://doi.org/10.3390/d11070117