Dactylonectria and Ilyonectria Species Causing Black Foot Disease of Andean Blackberry (Rubus Glaucus Benth) in Ecuador


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates from Symptomatic Plants
2.2. Morphological Characterization
2.3. Molecular Identification and Phylogenetic Analysis
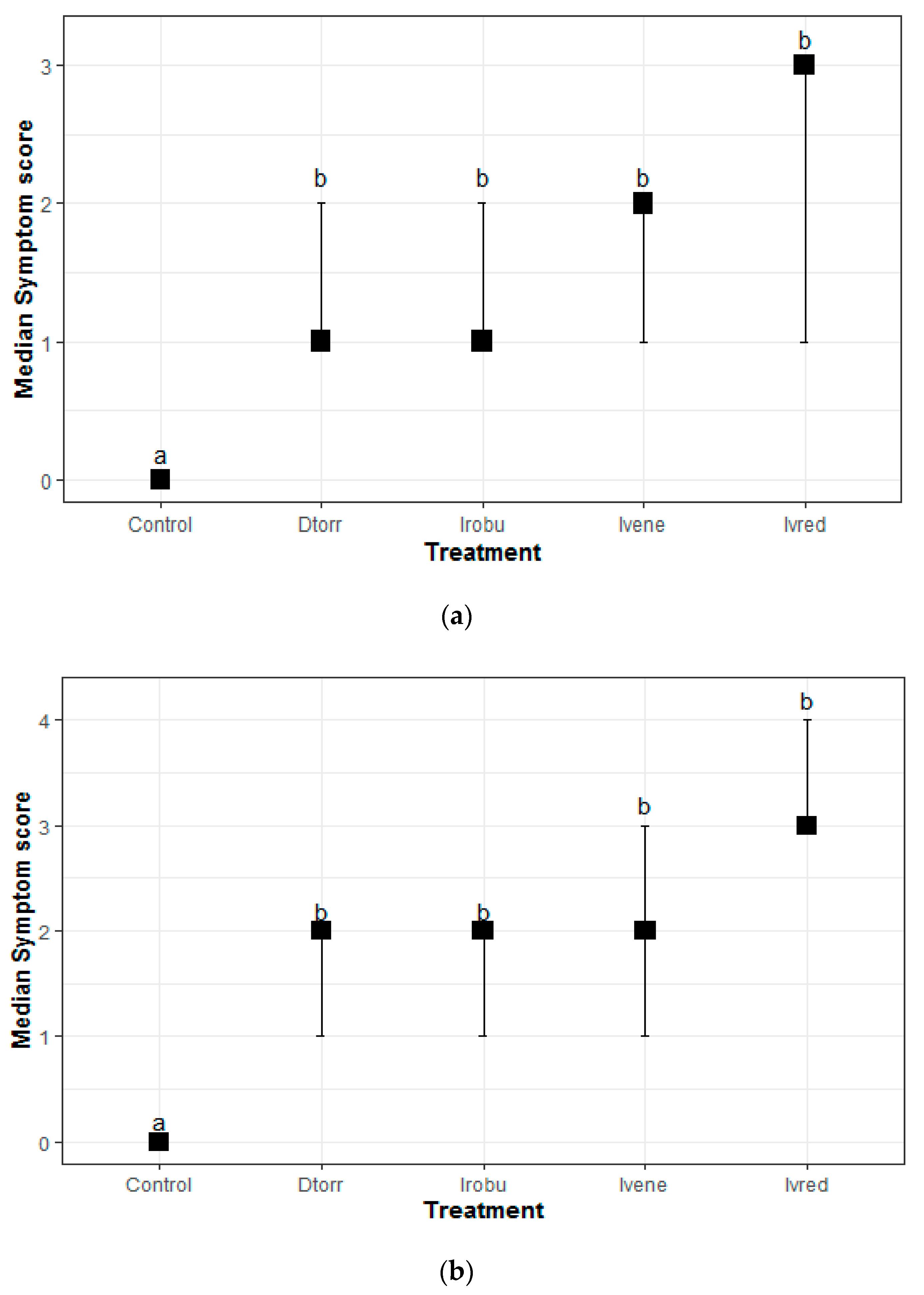
2.4. Pathogenicity Tests
3. Results
3.1. Isolates
3.2. Morphological Characterization
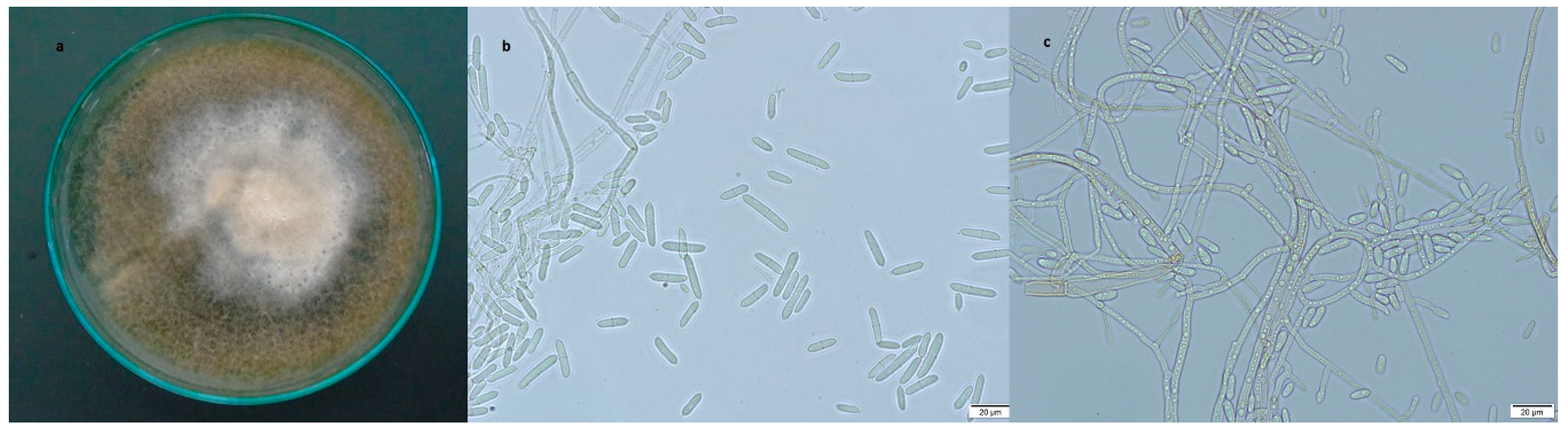
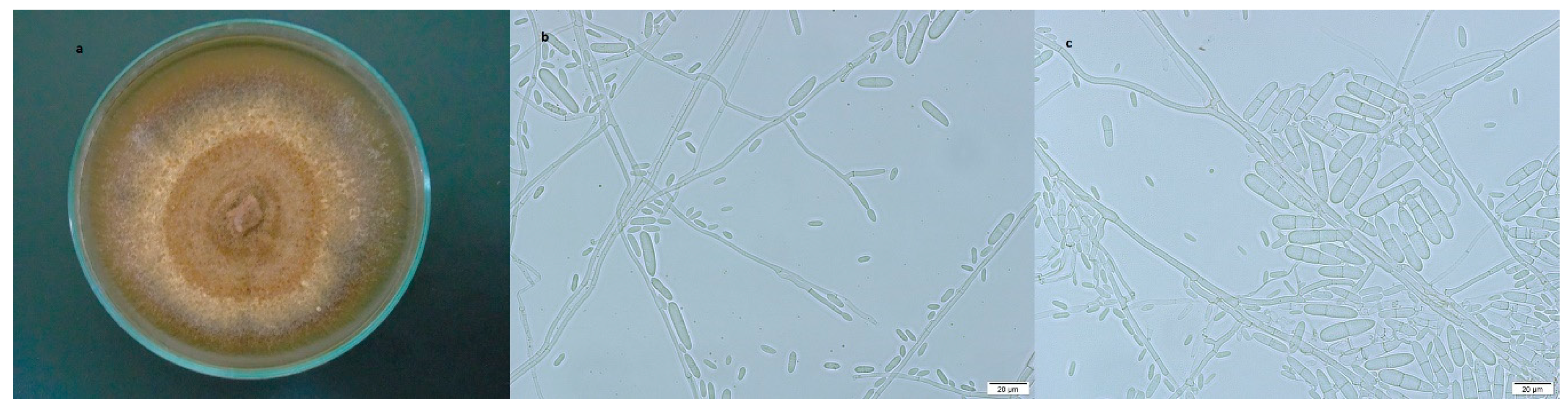
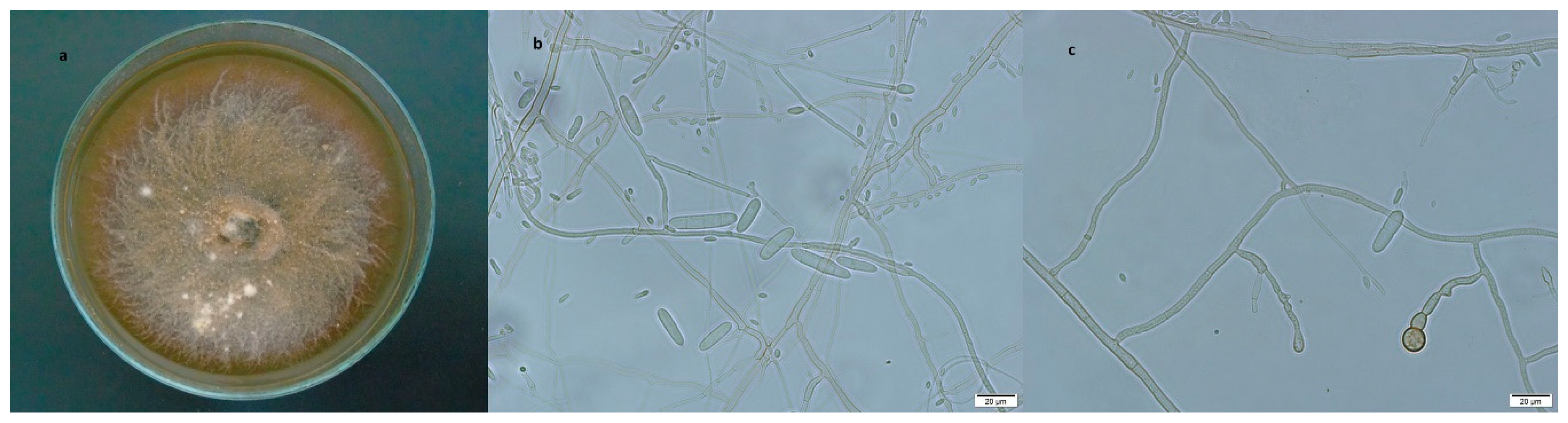
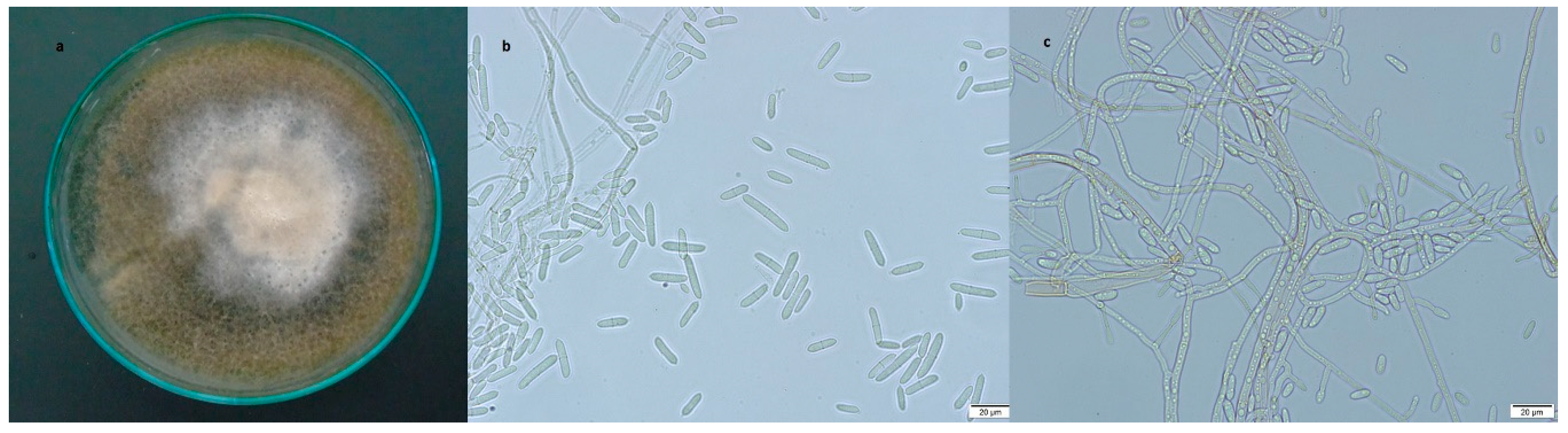
3.2.1. Dactylonectria torresensis
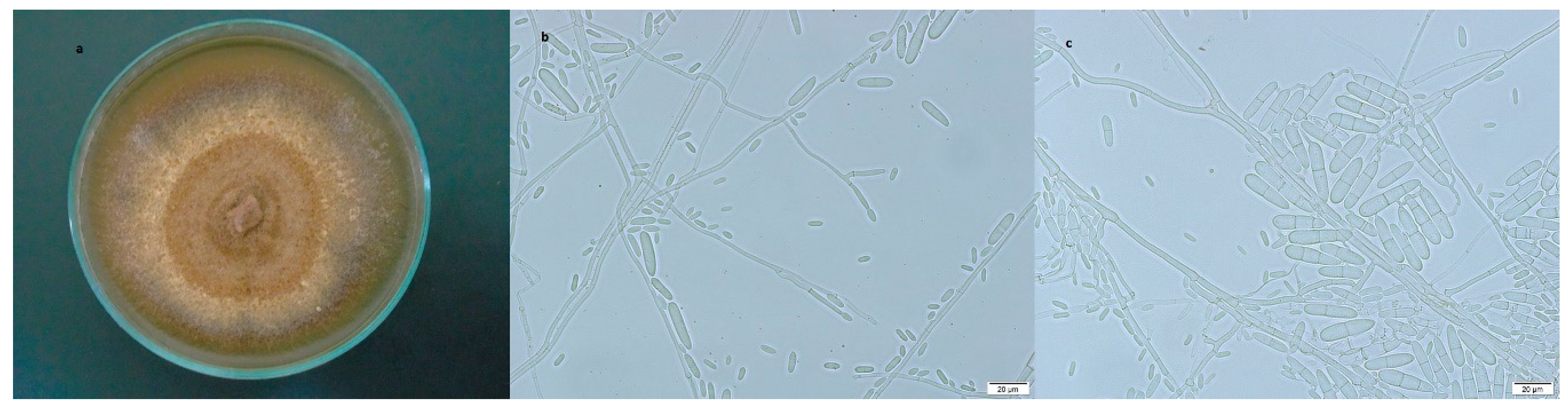
3.2.2. Ilyonectria robusta
3.2.3. Ilyonectria venezuelensis
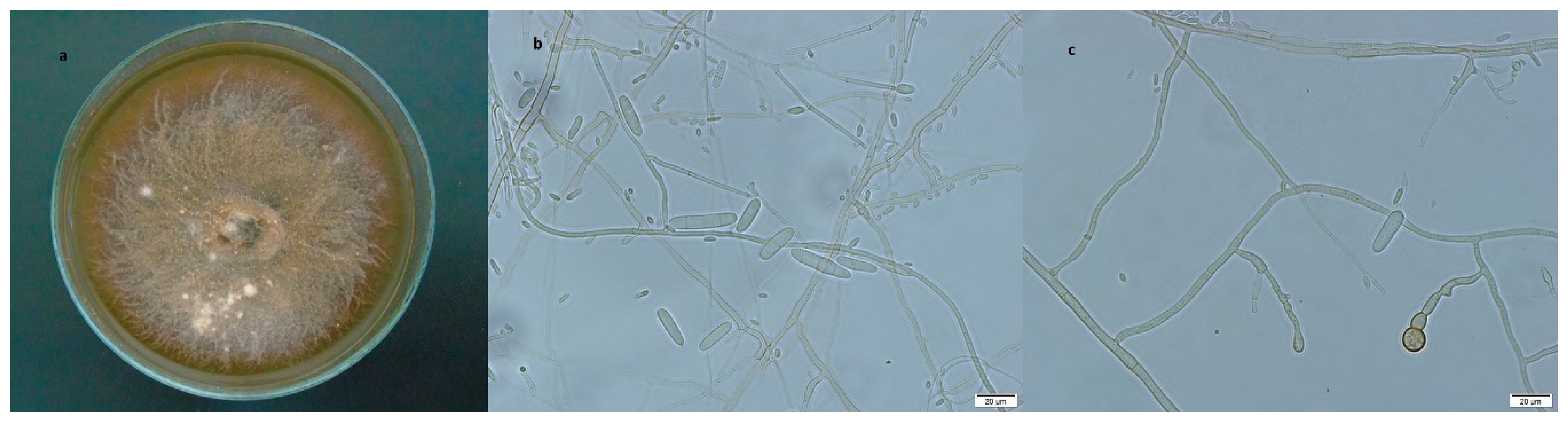
3.2.4. Ilyonectria vredehoekensis
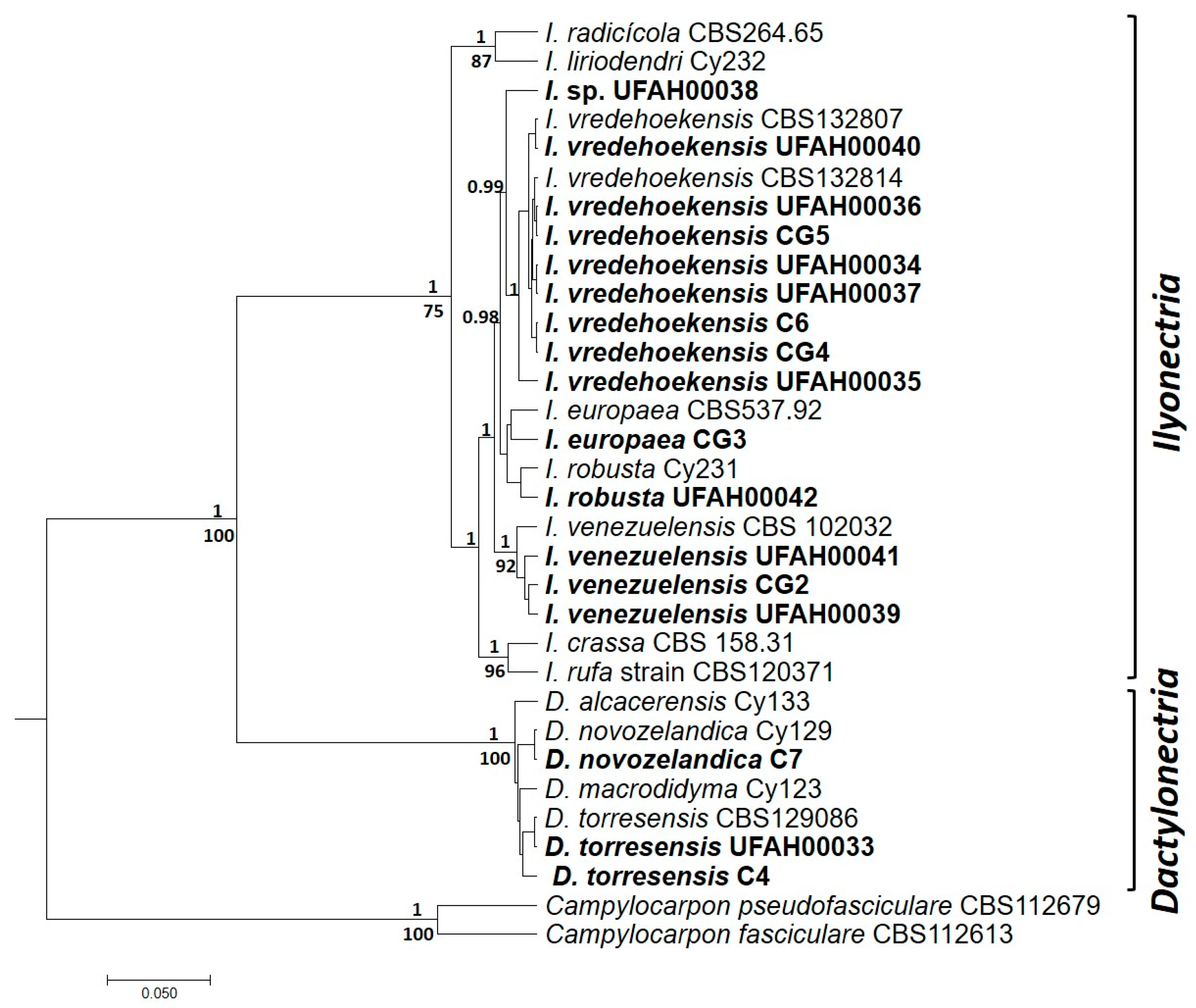
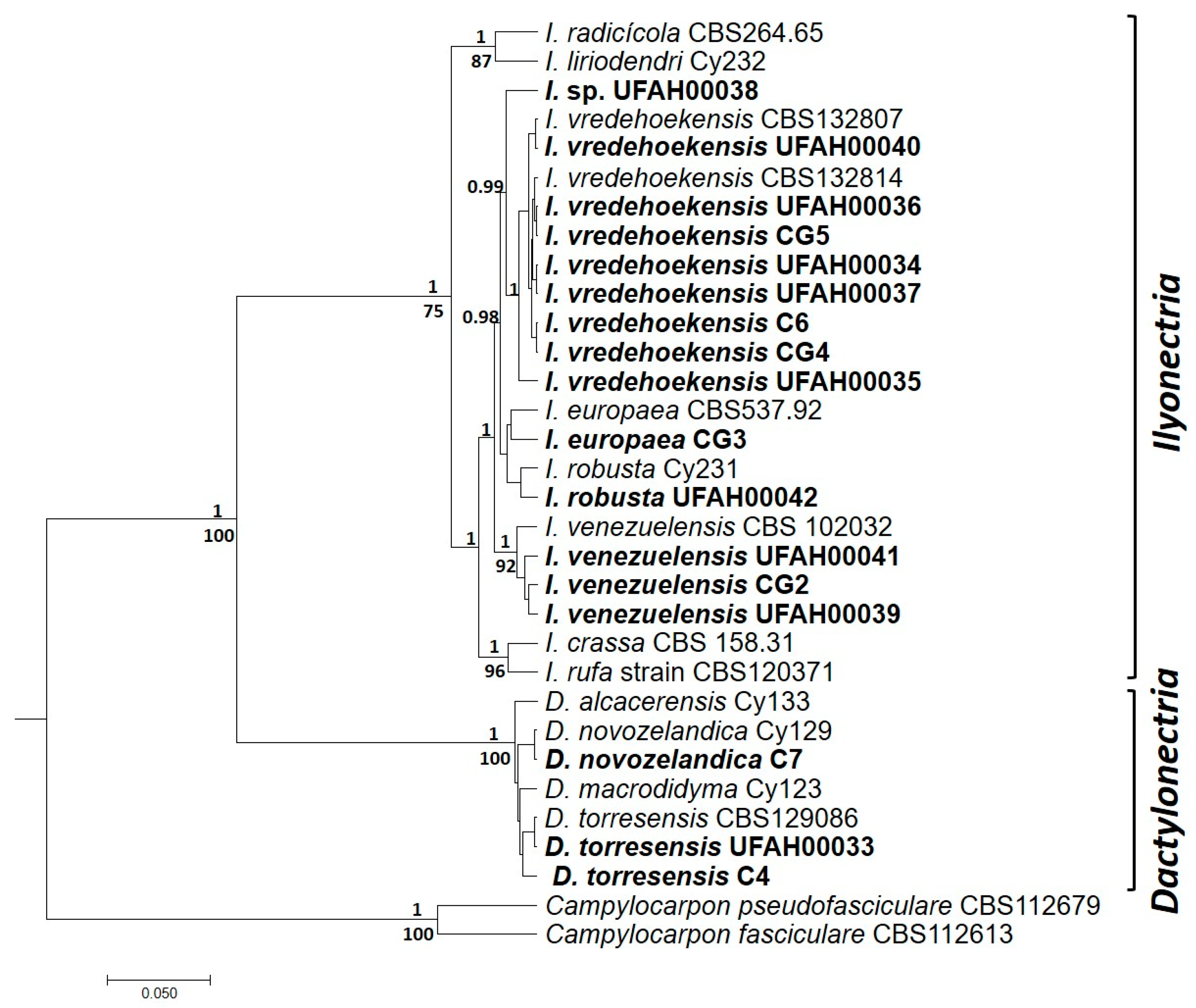
3.3. Molecular Identification and Phylogenetic Analysis
3.4. Pathogenicity Tests
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chaverri, P.; Salgado, C.; Hirooka, Y.; Rossman, A.; Samuels, G. Delimitation of Neonectria and Cylindrocarpon (Nectriaceae, Hypocreales, Ascomycota) and related genera with Cylindrocarpon-like anamorphs. Stud. Mycol. 2011, 68, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Samuels, G.; Brayford, D. Variation in Nectria radicicola and its anamorph, Cylindrocarpon destructans. Mycol. Res. 1990, 94, 433–442. [Google Scholar] [CrossRef]
- Halleen, F.; Schroers, H.; Groenewald, J.; Crous, P. Novel species of Cylindrocarpon (Neonectria) and Campylocarpon gen. nov. associated with black foot disease of grapevines (Vitis spp.). Stud. Mycol. 2004, 50, 431–455. [Google Scholar]
- Halleen, F.; Schroers, H.; Groenewald, J.; Rego, C.; Oliveira, H.; Crous, P. Neonectria liriodendri sp. nov., the main causal agent of black foot disease of grapevines. Stud. Mycol. 2006, 55, 227–234. [Google Scholar] [CrossRef]
- Maluta, D.; Larignon, P. Pied-noir: Mieux veut prèvenir. Viticulture 1991, 11, 71–72. [Google Scholar]
- Weber, R.; Entrop, A. Dactylonectria torresensis as the main component of the black root rot complex of strawberries and raspberries in northern Germany. Erwerbs-Obstbau 2017, 59, 157–169. [Google Scholar] [CrossRef]
- Cedeño, L.; Carrera, C.; Quintero, K.; Pino, H.; Espinoza, W. Cylindrocarpon destructans var. destructans and neonectria discophora var. rubi associated with black foot rot on blackberry (Rubus glaucus Benth.) in Merida, Venezuela. Interciencia 2004, 29, 455–460. [Google Scholar]
- Cabral, A.; Groenewald, J.; Rego, C.; Oliveira, H.; Crous, P. Cylindrocarpon root rot: Multi-gene analysis reveals novel species within the Ilyonectria radicicola species complex. Mycol. Prog. 2012, 11, 655–688. [Google Scholar] [CrossRef]
- Lombard, L.; Bezuidenhout, C.M.; Crous, P.W. Ilyonectria black foot rot associated with Proteaceae. Australas. Plant Pathol. 2013, 42, 337–349. [Google Scholar] [CrossRef]
- Cabral, A.; Nascimento, T.; Oliveira, H.; Groenewald, J.Z.; Crous, P.W.; Spatafora, J.W. Multi-gene analysis and morphology reveal novel Ilyonectria species associated with black foot disease of grapevines. Fungal Biol. 2012, 116, 62–80. [Google Scholar] [CrossRef]
- Brayford, D.; Honda, B.; Mantiri, F.; Samuels, G. Neonectria and Cylindrocarpon: The Nectria mammoidea group and species lacking microconidia. Mycologia 2004, 96, 572–597. [Google Scholar] [CrossRef] [PubMed]
- Lombard, L.; van der Merwe, N.; Groenwald, J.; Crous, P. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [PubMed]
- Lombard, L.; Van Der Merwe, N.; Groenewald, J.; Crous, P. Lineages in Nectriaceae: Re-evaluating the generic status of Ilyonectria and allied genera. Phytopathol. Mediterr. 2014, 53, 515–532. [Google Scholar]
- Crous, P.; Groenewald, J.; Risède, J.; Simoneau, P.; Hywel-Jones, N. Calonectria species and their Cylindrocladium anamorphs: Species with sphaeropedunculate vesicles. Stud. Mycol. 2004, 50, 415–430. [Google Scholar]
- O’Donnell, K.; Kistler, H.; Cigelnik, E.; Ploetz, R. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Flores, J.; Marek, S.; Orquera, G.; Walker, N. Molecular identification and multilocus phylogeny of ophiosphaerella species associated with spring dead spot of bermudagrass. Crop Sci. 2017, 57, S249–S261. [Google Scholar] [CrossRef]
- Edgar, R. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Drummond, A.; Suchard, M.; Dong, X.; Rambaut, A. Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Alaniz, S.; León, M.; Vicent, A.; García-Jiménez, J.; Abad-Campos, P.; Armengol, J. Characterization of Cylindrocarpon Species Associated with Black Foot Disease of Grapevine in Spain. Plant Dis. 2007, 91, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Úrbez-Torres, J.R.; Haag, P.; Bowen, P.; O’Gorman, D.T. Grapevine trunk diseases in British Columbia: Incidence and characterization of the fungal pathogens associated with black foot disease of grapevine. Plant Dis. 2014, 98, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Aiello, D.; Guarnaccia, V.; Epifani, F.; Perrone, G.; Polizzi, G. “Cylindrocarpon” and Ilyonectria Species Causing Root and Crown Rot Disease of Potted Laurustinus Plants in Italy. J. Phytopathol. 2015, 163, 675–680. [Google Scholar] [CrossRef]
- Kluge, E. Pathogenität gegenüber kiefernsämlingen und toxinbildung bei Cylindrocarpon radicicola Wr. J. Phytopathol. 1966, 55, 368–388. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Species | Year | Province/Location | Accession Number | |
|---|---|---|---|---|---|
| TEF | HIS3 | ||||
| CG2 | Ilyonectria venezuelensis | 2014 | Bolivar/Guaranda | MG852016 | MG852002 |
| CG3 | Ilyonectria europaea | 2014 | Bolivar/Guaranda | MG852017 | MG852003 |
| CG4 | Ilyonectria vredehoekensis | 2014 | Bolivar/Guaranda | - | MG852004 |
| CG5 | I. vredehoekensis | 2014 | Bolivar/Guaranda | - | MG852005 |
| C4 | Dactylonectria torresensis | 2014 | Tungurahua/Ambato | MG852018 | MG851999 |
| C6 | I. vredehoekensis | 2014 | Tungurahua/Ambato | - | MG852000 |
| C7 | Dactylonectria novozelandica | 2014 | Tungurahua/Ambato | MG852019 | MG852001 |
| UFAH00033 | D. torresensis | 2016 | Tungurahua/Tisaleo | KY114517 | MG852015 |
| UFAH00034 | I. vredehoekensis | 2017 | Bolivar/Chillanes | MG852020 | MG852014 |
| UFAH00035 | I. vredehoekensis | 2017 | Bolivar/Chillanes | MG852021 | MG852006 |
| UFAH00036 | I. vredehoekensis | 2017 | Bolivar/Chillanes | MG852022 | MG852013 |
| UFAH00037 | I. vredehoekensis | 2017 | Tungurahua/Píllaro | - | MG852010 |
| UFAH00038 | Ilyonectria sp. | 2017 | Tungurahua/Ambato | - | MG852009 |
| UFAH00039 | I. venezuelensis | 2017 | Tungurahua/Tisaleo | MG852023 | MG852012 |
| UFAH00040 | I. vredehoekensis | 2017 | Bolivar/Chillanes | - | MG852008 |
| UFAH00041 | I. venezuelensis | 2017 | Tungurahua/Ambato | MG852024 | MG852011 |
| UFAH00042 | Ilyonectria robusta | 2017 | Tungurahua/Cevallos | - | MG852007 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez, J.; Iturralde, P.; Koch, A.; Tello, C.; Martinez, D.; Proaño, N.; Martínez, A.; Viera, W.; Ayala, L.; Flores, F. Dactylonectria and Ilyonectria Species Causing Black Foot Disease of Andean Blackberry (Rubus Glaucus Benth) in Ecuador. Diversity 2019, 11, 218. https://doi.org/10.3390/d11110218
Sánchez J, Iturralde P, Koch A, Tello C, Martinez D, Proaño N, Martínez A, Viera W, Ayala L, Flores F. Dactylonectria and Ilyonectria Species Causing Black Foot Disease of Andean Blackberry (Rubus Glaucus Benth) in Ecuador. Diversity. 2019; 11(11):218. https://doi.org/10.3390/d11110218
Chicago/Turabian StyleSánchez, Jessica, Paola Iturralde, Alma Koch, Cristina Tello, Dennis Martinez, Natasha Proaño, Anibal Martínez, William Viera, Ligia Ayala, and Francisco Flores. 2019. "Dactylonectria and Ilyonectria Species Causing Black Foot Disease of Andean Blackberry (Rubus Glaucus Benth) in Ecuador" Diversity 11, no. 11: 218. https://doi.org/10.3390/d11110218
APA StyleSánchez, J., Iturralde, P., Koch, A., Tello, C., Martinez, D., Proaño, N., Martínez, A., Viera, W., Ayala, L., & Flores, F. (2019). Dactylonectria and Ilyonectria Species Causing Black Foot Disease of Andean Blackberry (Rubus Glaucus Benth) in Ecuador. Diversity, 11(11), 218. https://doi.org/10.3390/d11110218