Abrupt Change in a Subtidal Rocky Reef Community Coincided with a Rapid Acceleration of Sea Water Warming

 ,
,  and
and 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
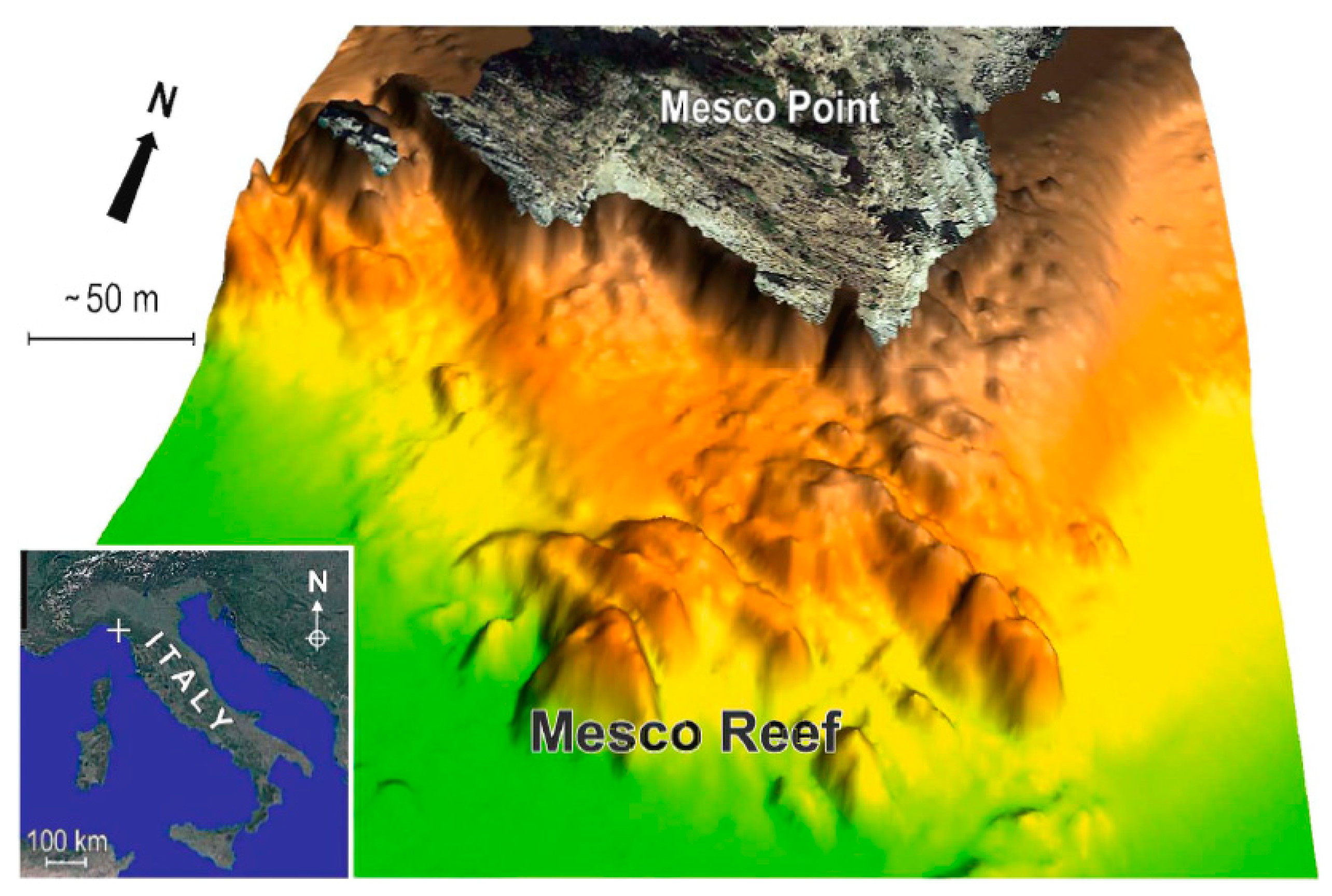
2.1. Study Area and Field Activity
2.2. Temperature Data
2.3. Data Management
3. Results
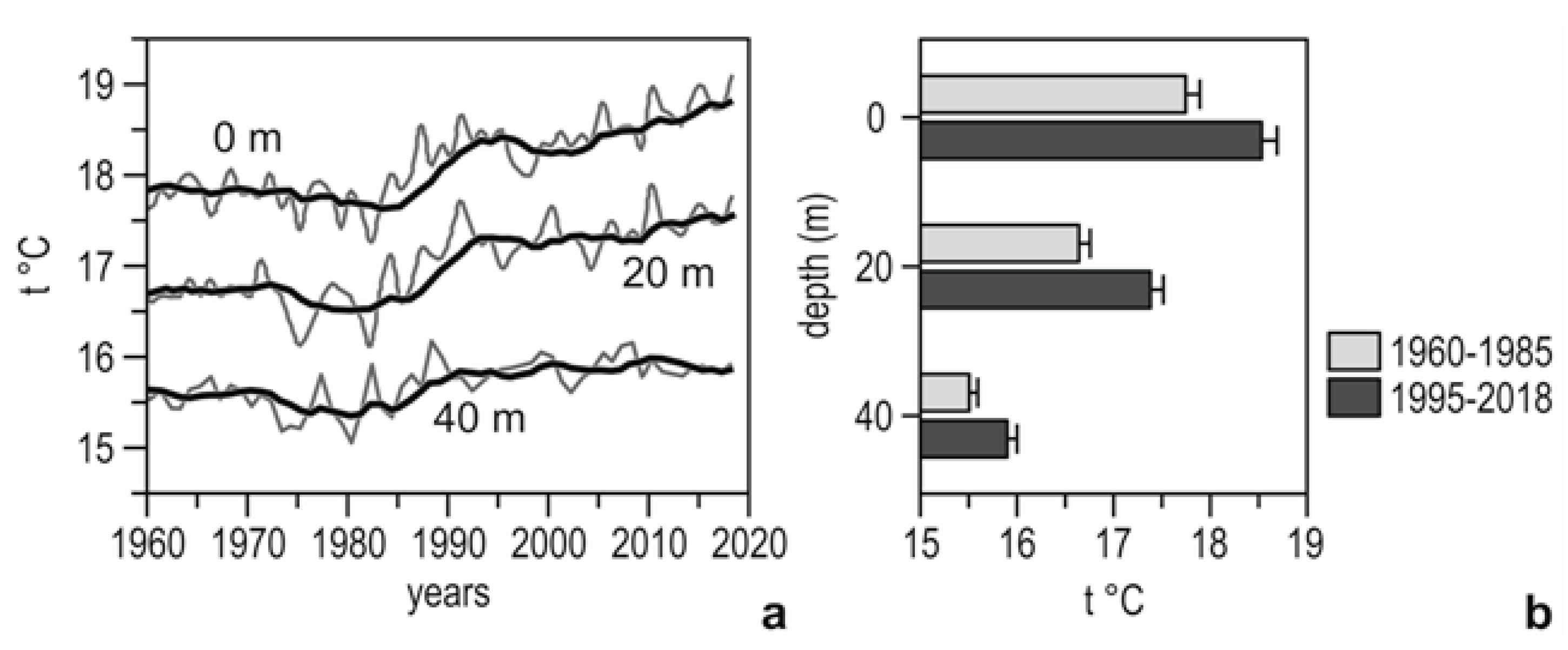
3.1. Sea Water Temperature
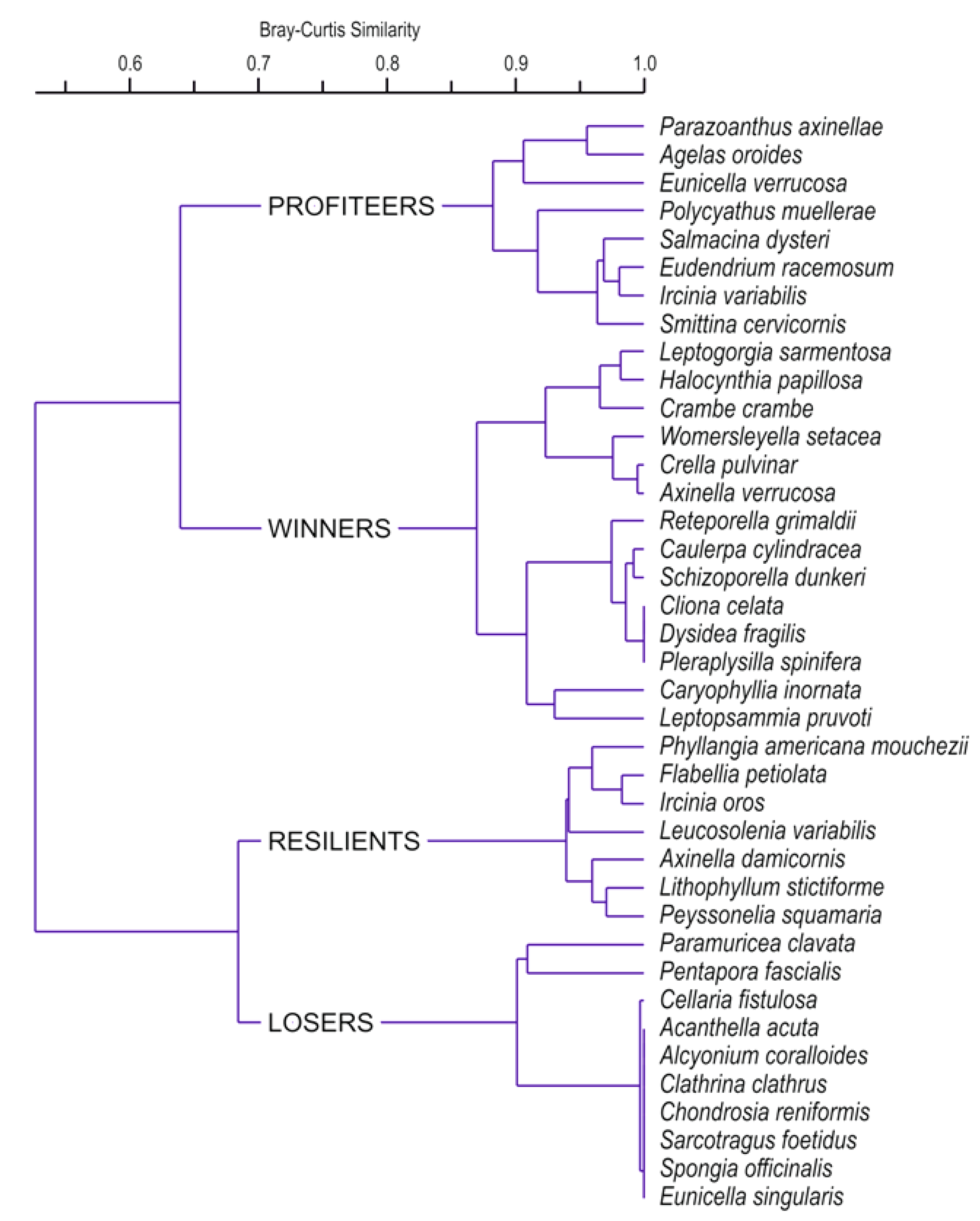
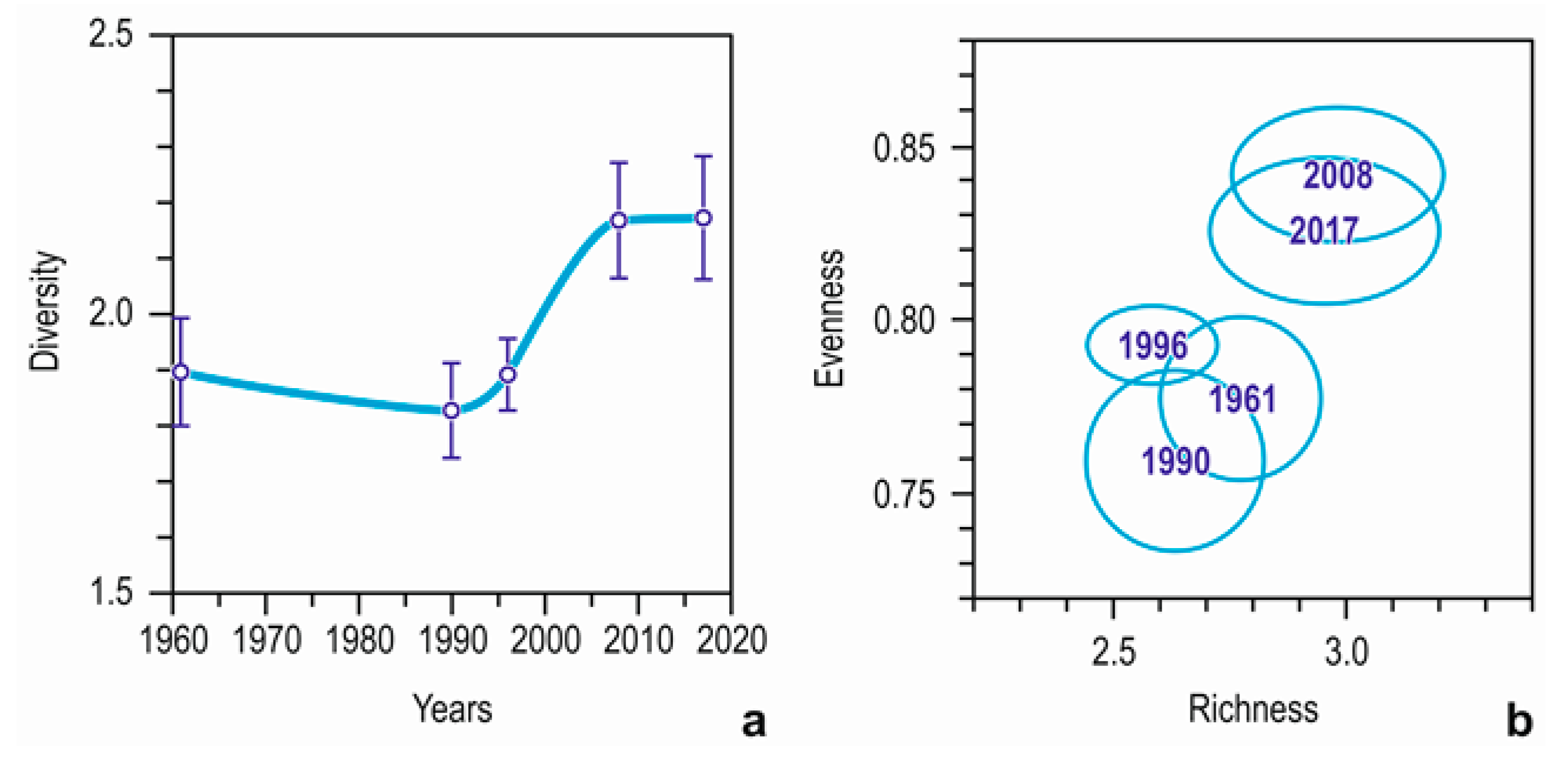
3.2. Species and Species Groups
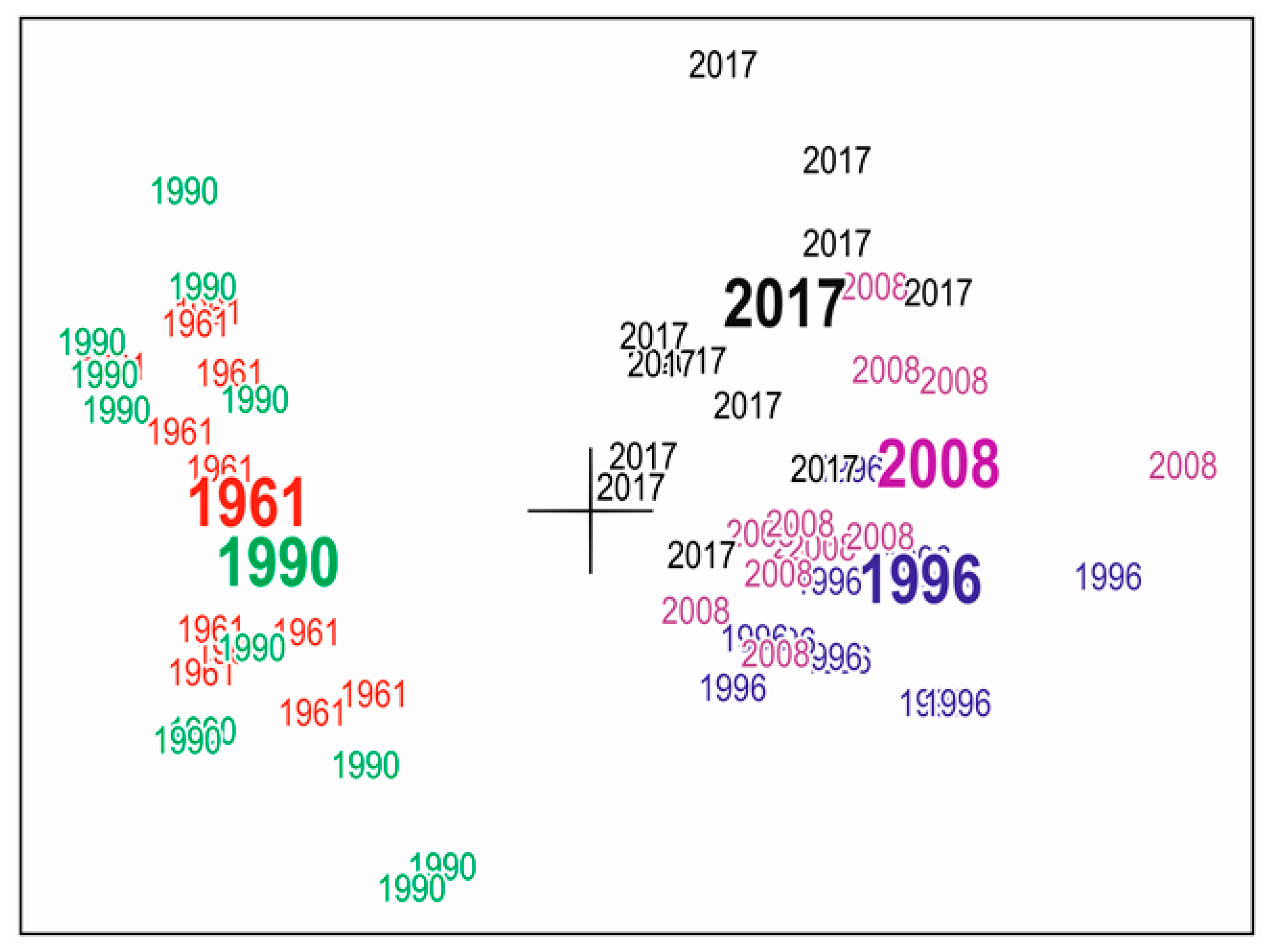
3.3. Community Composition
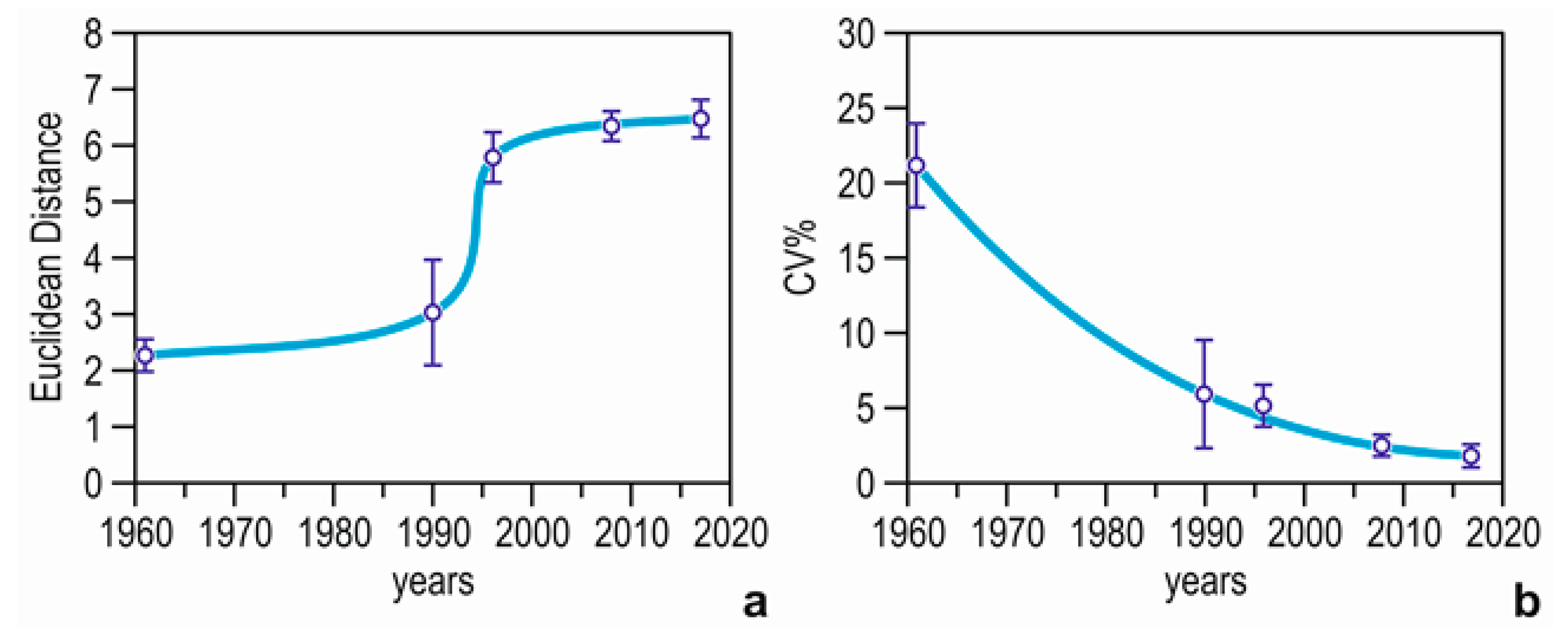
3.4. Community Structure
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Walther, G.R. Community and ecosystem responses to recent climate change. Philos. Trans. R. Soc. B 2010, 365, 2019–2024. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Poloczanska, E.S. The effect of climate change across ocean regions. Front. Mar. Sci. 2017, 4, 361. [Google Scholar] [CrossRef]
- Harley, C.D.G.; Hughes, A.R.; Hultgren, K.M.; Miner, B.G.; Sorte, C.J.B.; Thornber, C.S.; Rodriguez, L.F.; Tomanek, L.; Williams, S.L. The impacts of climate change in coastal marine systems. Ecol. Lett. 2006, 9, 228–241. [Google Scholar] [CrossRef]
- Beaugrand, G.; Conversi, A.; Atkinson, A.; Cloern, J.; Chiba, S.; Fonda-Umani, S.; Kirby, R.R.; Greene, C.H.; Goberville, E.; Otto, S.A.; et al. Prediction of unprecedented biological shifts in the global ocean. Nat. Clim. Chang. 2019, 9, 237–243. [Google Scholar] [CrossRef]
- Smale, D.A.; Taylor, J.D.; Coombs, S.H.; Moore, G.; Cunliffe, M. Community responses to seawater warming are conserved across diverse biological groupings and taxonomic resolutions. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170534. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C. Climate change and biological response in Mediterranean Sea ecosystems—A need for broad-scale and long-term research. Ocean Chall. 2004, 13, 32–36. [Google Scholar]
- Doney, S.C.; Ruckelshaus, M.; Duffy, J.E.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N.; et al. Climate change impacts on marine ecosystems. Annu. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef]
- Lubchenco, J.; Navarrete, S.A.; Tissot, B.N.; Castilla, J.C. Possible ecological responses to global climate change: Nearshore benthic biota of northeastern Pacific coastal ecosystems. In Earth System Responses to Global Change; Mooney, H.A., Fuentes, E.R., Kronberg, B.I., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 147–166. [Google Scholar]
- Southward, A.J.; Boalch, G.T. The effect of changing climate on marine life: Past events and future predictions. In Man and the Maritime Environment; Fisher, S., Ed.; Maritime Studies; University of Exeter Press: Exeter, UK, 1994; pp. 101–143. [Google Scholar]
- Bianchi, C.N. Climate change and biological response in the marine benthos. Proc. It. Ass. Oceanol. Limnol. 1997, 12, 3–20. [Google Scholar]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Edwards, M.; Richardson, A.J. Impact of climate change on marine pelagic phenology and trophic mismatch. Nature 2004, 430, 881–884. [Google Scholar] [CrossRef]
- Perry, A.L.; Low, P.J.; Ellis, J.R.; Reynolds, J.D. Climate change and distribution shifts in marine fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef]
- Helmuth, B.; Mieszkowska, N.; Moore, P.; Hawkins, S.J. Living on the edge of two changing worlds: Forecasting the responses of rocky intertidal ecosystems to climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 373–404. [Google Scholar] [CrossRef]
- de Madron, X.D.; Guieu, C.; Sempéré, R.; Conan, P.; Cossa, D.; D’Ortenzio, F.; Estournel, C.; Gazeau, F.; Rabouille, C.; Stemmann, L.; et al. Marine ecosystems’ responses to climatic and anthropogenic forcings in the Mediterranean. Prog. Oceanogr. 2011, 91, 97–166. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C.; Chiantore, M.; Montefalcone, M.; Parravicini, V.; Rovere, A. Mediterranean Sea biodiversity between the legacy from the past and a future of change. In Life in the Mediterranean Sea: A Look at Habitat Changes; Stambler, N., Ed.; Nova Science Publishers: New York, NY, USA, 2012; pp. 1–55. [Google Scholar]
- Marbà, N.; Jordà, G.; Agustí, S.; Girard, C.; Duarte, C.M. Footprints of climate change on Mediterranean Sea biota. Front. Mar. Sci. 2015, 2, 56. [Google Scholar] [CrossRef]
- Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 2006, 33, 08707. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C. Marine biodiversity of the Mediterranean Sea: Situation, problems and prospects for future research. Mar. Pollut. Bull. 2000, 40, 367–376. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef]
- Lejeusne, C.; Chevaldonné, P.; Pergent-Martini, C.; Boudouresque, C.F.; Pérez, T. Climate change effects on a miniature ocean: The highly diverse, highly impacted Mediterranean Sea. Trends Ecol. Evol. 2010, 25, 250–260. [Google Scholar] [CrossRef]
- Turley, C.M. The changing Mediterranean Sea, a sensitive ecosystem? Prog. Oceanogr. 1999, 44, 387–400. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C. Global sea warming and “tropicalization” of the Mediterranean Sea: Biogeographic and ecological aspects. Biogeographia 2003, 24, 319–327. [Google Scholar] [CrossRef]
- Bianchi, C.N. Biodiversity issues for the forthcoming tropical Mediterranean Sea. Hydrobiologia 2007, 580, 7–21. [Google Scholar] [CrossRef]
- Hampton, S.E.; Scheuerell, M.D.; Church, M.J.; Melack, J.M. Long-term perspectives in aquatic research. Limnol. Oceanogr. 2019, 64, S2–S10. [Google Scholar] [CrossRef]
- Muelbert, J.H.; Nidzieko, N.J.; Acosta, A.T.R.; Beaulieu, S.E.; Bernardino, A.F.; Boikova, E.; Bornman, T.G.; Cataletto, B.; Deneudt, K.; Eliason, E.; et al. ILTER–The international long-term ecological research network as a platform for global coastal and ocean observation. Front. Mar. Sci. 2019, 6, 527. [Google Scholar] [CrossRef]
- Calvo, E.; Simó, R.; Coma, R.; Ribes, M.; Pascual, J.; Sabatés, A.; Gili, J.M.; Pelejero, C. Effects of climate change on Mediterranean marine ecosystems: The case of the Catalan Sea. Clim. Res. 2011, 50, 1–29. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Azzola, A.; Bertolino, M.; Betti, F.; Bo, M.; Cattaneo-Vietti, R.; Cocito, S.; Montefalcone, M.; Morri, C.; Oprandi, A.; et al. Consequences of the marine climate and ecosystem shift of the 1980-90s on the Ligurian Sea biodiversity (NW Mediterranean). Eur. Zool. J. 2019, 86, 458–487. [Google Scholar]
- Gatti, G.; Bianchi, C.N.; Montefalcone, M.; Venturini, S.; Diviacco, G.; Morri, C. Observational information on a temperate reef community helps understanding the marine climate and ecosystem shift of the 1980–90s. Mar. Pollut. Bull. 2017, 114, 528–538. [Google Scholar] [CrossRef]
- Conversi, A.; Fonda Umani, S.; Peluso, T.; Molinero, J.C.; Santojanni, R.; Edwards, M. The Mediterranean Sea regime shift at the end of the 1980s, and intriguing parallelism with other European basins. PLoS ONE 2010, 5, e10633. [Google Scholar] [CrossRef]
- Reid, P.C.; Hari, R.E.; Beaugrand, G.; Livingstone, D.M.; Marty, C.; Straile, D.; Barichivich, J.; Goberville, E.; Adrian, R.; Aono, Y.; et al. Global impacts of the 1980s regime shift. Glob. Chang. Biol. 2016, 22, 682–703. [Google Scholar] [CrossRef]
- Alheit, J.; Gröger, J.; Licandro, P.; McQuinn, I.H.; Pohlmann, T.; Tsikliras, A.C. What happened in the mid-1990s? The coupled ocean-atmosphere processes behind climate-induced ecosystem changes in the Northeast Atlantic and the Mediterranean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2019, 159, 130–142. [Google Scholar] [CrossRef]
- Gatti, G.; Bianchi, C.N.; Parravicini, V.; Rovere, A.; Peirano, A.; Montefalcone, M.; Massa, F.; Morri, C. Ecological change, sliding baselines and the importance of historical data: Lessons from combining observational and quantitative data on a temperate reef over 70 years. PLoS ONE 2015, 10, e0118581. [Google Scholar] [CrossRef]
- Tortonese, E.; Faraggiana, R. Osservazioni biologiche nell’insenatura di Levanto (Riviera Ligure). Natura 1937, 28, 51–72. [Google Scholar]
- Roghi, G. La foresta viola: Tre anni di ricerche biologiche a cinquanta metri di profondità sulla secca del Mesco. Mondo Sommerso 1963, 5, 110–115. [Google Scholar]
- Rossi, L. Il coralligeno di Punta Mesco (La Spezia). Ann. Mus. Civ. Stor. Nat. Genova 1965, 75, 144–180. [Google Scholar]
- Tunesi, L.; Molinari, A.; Salvati, E. Fish assemblage of the marine protected area of Cinque Terre (NW Mediterranean Sea): First characterization and assessment by visual census. Chem. Ecol. 2006, 22 (Suppl. 1), 245–253. [Google Scholar] [CrossRef]
- Azzola, A.; Bianchi, C.N.; Morri, C.; Montefalcone, M. Evolution of coralligenous shoals at Punta Mesco: 60 years of history. In Proceedings of the 50° Congresso della Società Italiana di Biologia Marina, Livorno, Italy, 10–14 June 2019; pp. 184–187. [Google Scholar]
- Peirano, A.; Sassarini, M. Analisi delle caratteristiche distributive di alcune facies di substrato duro dei fondali delle Cinque Terre (Mar Ligure). Oebalia 1992, 17, 523–528. [Google Scholar]
- Peirano, A.; Salvati, E.; Bianchi, C.N.; Morri, C. Long-term change in the subtidal epibenthic assemblages of Punta Mesco (Ligurian Sea, Italy) as assessed through underwater photography. PMNHS Newsl. 2000, 5, 9–12. [Google Scholar]
- Bianchi, C.N.; Peirano, A.; Salvati, E.; Morri, C. Assessing interannual and decadal changes in marine epibenthic assemblages through UW photography: An example from Punta Mesco, Ligurian Sea. Arch. Oceanogr. Limnol. 2001, 22, 83–86. [Google Scholar]
- Roghi, F.; Parravicini, V.; Montefalcone, M.; Rovere, A.; Morri, C.; Peirano, A.; Salvati, E.; Firpo, M.; Bianchi, C.N. Decadal Evolution of a Temperate Reef Ecosystem under the Influence of Climate Change. In Reefs in a Changing Environment; Kaandorp, J., Meesters, E., Osinga, R., Verreth, J., Wijgerde, T., Eds.; ISRS: Wageningen, The Netherlands, 2010; pp. 77–78. [Google Scholar]
- Roghi, F.; Parravicini, V.; Montefalcone, M.; Rovere, A.; Morri, C.; Peirano, A.; Firpo, M.; Bianchi, C.N. Decadal evolution of a coralligenous ecosystem under the influence of human impacts and climate change. Biol. Mar. Mediterr. 2010, 17, 59–62. [Google Scholar]
- Prieur, L. Physical historical data on the Ligurian Sea from the Villefranche-sur-Mer Observatory. In Tracking Long-Term Hydrological Change in the Mediterranean Sea; Millot, C., Briand, F., Eds.; Workshop Series No. 16; CIESM: Monaco, 2002; pp. 37–40. [Google Scholar]
- Parravicini, V.; Mangialajo, L.; Mousseau, L.; Peirano, A.; Morri, C.; Francour, P.; Kulbicki, M.; Bianchi, C.N. Climate change and warm-water species at the north-western boundary of the Mediterranean Sea. Mar. Ecol. 2015, 36, 897–909. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Caroli, F.; Guidetti, P.; Morri, C. Seawater warming at the northern reach for southern species: Gulf of Genoa, NW Mediterranean. J. Mar. Biol. Assoc. UK 2018, 98, 1–12. [Google Scholar] [CrossRef]
- Astraldi, M.; Bianchi, C.N.; Gasparini, G.P.; Morri, C. Climatic fluctuations, current variability and marine species distribution: A case study in the Ligurian Sea (North-West Mediterranean). Oceanol. Acta 1995, 18, 139–149. [Google Scholar]
- Bianchi, C.N.; Corsini-Foka, M.; Morri, C.; Zenetos, A. Thirty years after: Dramatic change in the coastal marine ecosystems of Kos Island (Greece), 1981–2013. Mediterr. Mar. Sci. 2014, 15, 482–497. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Cocito, S.; Diviacco, G.; Dondi, N.; Fratangeli, F.; Montefalcone, M.; Parravicini, V.; Rovere, A.; Sgorbini, S.; Vacchi, M.; et al. The park never born: Outcome of a quarter of a century of inaction on the sea-floor integrity of a proposed but not established Marine Protected Area. Aquat. Conserv. 2018, 28, 1209–1228. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 1–264. [Google Scholar]
- Abbiati, M.; Bianchi, C.N.; Castelli, A.; Giangrande, A.; Lardicci, C. Distribution of Polychaetes on hard substrates of the midlittoral-infralittoral transition zone, western Mediterranean. Ophelia 1991, 5, 421–432. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PaSt: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Cattaneo-Vietti, R.; Cappanera, V.; Castellano, M.; Povero, P. Yield and catch changes in a Mediterranean small tuna trap: A climate change effect? Mar. Ecol. 2015, 36, 155–166. [Google Scholar] [CrossRef]
- Morri, C.; Bianchi, C.N. Recent changes in biodiversity in the Ligurian Sea (NW Mediterranean): Is there a climatic forcing? In Mediterranean Ecosystems: Structure and Processes; Faranda, F.M., Guglielmo, L., Spezie, G., Eds.; Springer: Milano, Italy, 2001; pp. 375–384. [Google Scholar]
- Montefalcone, M.; Parravicini, V.; Bianchi, C.N. Quantification of coastal ecosystem resilience. In Treatise on Estuarine and Coastal Science; Wolanski, E., McLusky, D.S., Eds.; Ecohydrology and Restoration; Academic Press: Waltham, MA, USA, 2011; pp. 49–70. [Google Scholar]
- Ponti, M.; Perini, R.A.; Ventra, V.; Grech, D.; Abbiati, M. Ecological shifts in Mediterranean coralligenous assemblages related to gorgonian forest loss. PLoS ONE 2014, 9, e102782. [Google Scholar] [CrossRef]
- Rossi, S.; Isla, E.; Bosch-Belmar, M.; Galli, G.; Gori, A.; Gristina, M.; Ingrosso, G.; Milisenda, G.; Piraino, S.; Rizzo, L.; et al. Changes of energy fluxes in marine animal forests of the Anthropocene: Factors shaping the future seascape. ICES J. Mar. Sci. in press. [CrossRef]
- Boudouresque, C.F. Recherches sur les concepts de biocénose et de continuum au niveau de peuplements benthiques sciaphiles. Vie Milieu 1970, 21, 103–136. [Google Scholar]
- Nielsen, T.F.; Sand-Jensen, K.; Dornelas, M.; Bruun, H.H. More is less: Net gain in species richness, but biotic homogenization over 140 years. Ecol. Lett. 2019, 22, 1650–1657. [Google Scholar] [CrossRef]
- Baskin, Y. Winners and losers in a changing world. Bioscience 1998, 48, 788–792. [Google Scholar] [CrossRef]
- Zenetos, A.; Çinar, M.E.; Pancucci-Papadopoulou, M.A.; Harmelin, J.G.; Furnari, G.; Andaloro, F.; Bellou, N.; Streftaris, N.; Zibrowius, H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Medit. Mar. Sci. 2005, 6, 63–118. [Google Scholar] [CrossRef]
- Airoldi, L.; Rindi, F.; Cinelli, F. Structure, seasonal dynamics and reproductive phenology of a filamentous turf assemblage on a sediment influenced, rocky subtidal shore. Bot. Mar. 1995, 38, 227–273. [Google Scholar] [CrossRef]
- Bussotti, S.; Conti, M.; Guidetti, P.; Martini, F.; Matricardi, G. First record of Caulerpa racemosa (Forsskål) J. Agardh along the coast of Genoa (North-Western Mediterranean). Doriana 1996, 6, 1–5. [Google Scholar]
- Montefalcone, M.; Morri, C.; Parravicini, V.; Bianchi, C.N. A tale of two invaders: Divergent spreading kinetics of the alien green algae Caulerpa taxifolia and Caulerpa cylindracea. Biol. Invasions 2015, 17, 2717–2728. [Google Scholar] [CrossRef]
- Morri, C.; Montefalcone, M.; Gatti, G.; Vassallo, P.; Paoli, C.; Bianchi, C.N. An alien invader is the cause of homogenization in the recipient ecosystem: A simulation-like approach. Diversity 2019, 11, 146. [Google Scholar] [CrossRef]
- Fridley, J.D.; Stachowicz, J.J.; Naeem, S.; Sax, D.F.; Seabloom, E.W.; Smith, M.D.; Stohlgren, T.J.; Tilman, D.; Von Holle, B. The invasion paradox: Reconciling pattern and process in species invasions. Ecology 2007, 88, 3–17. [Google Scholar] [CrossRef]
- Guidetti, P.; Milazzo, M.; Bussotti, S.; Molinari, A.; Murenu, M.; Pais, A.; Spanò, N.; Balzano, R.; Agardy, T.; Boero, F.; et al. Italian marine reserve effectiveness: Does enforcement matter? Biol. Conserv. 2008, 141, 699–709. [Google Scholar] [CrossRef]
- Roberts, C.M.; O’Leary, B.C.; McCauley, D.J.; Cury, P.M.; Duarte, C.M.; Lubchenco, J.; Pauly, D.; Sáenz-Arroyo, A.; Sumaila, U.R.; Wilson, R.W.; et al. Marine reserves can mitigate and promote adaptation to climate change. Proc. Natl. Acad. Sci. USA 2017, 114, 6167–6175. [Google Scholar] [CrossRef]
- Giakoumi, S.; Pey, A.; Di Franco, A.; Francour, P.; Kizilkaya, Z.; Arda, Y.; Raybaud, V.; Guidetti, P. Exploring the relationships between marine protected areas and invasive fish in the world’s most invaded sea. Ecol. Appl. 2019, 29, 01809. [Google Scholar] [CrossRef]
- Wolff, B.A.; Duggan, S.B.; Clements, W.H. Resilience and regime shifts: Do novel communities impede ecological recovery in a historically metal-contaminated stream? J. Appl. Ecol. in press. [CrossRef]
- Shaltout, M.; Omstedt, A. Recent sea surface temperature trends and future scenarios for the Mediterranean Sea. Oceanologia 2014, 56, 411–443. [Google Scholar] [CrossRef]
- Adloff, F.; Somot, S.; Sevault, F.; Jordà, G.; Aznar, R.; Déqué, M.; Herrmann, M.; Marcos, M.; Dubois, C.; Padorno, E.; et al. Mediterranean Sea response to climate change in an ensemble of twenty first century scenarios. Clim. Dyn. 2015, 45, 2775–2802. [Google Scholar] [CrossRef]
- McDonald, T. Changes over time and changes yet to come. Ecol. Manag. Restor. 2019, 20, 181. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianchi, C.N.; Azzola, A.; Parravicini, V.; Peirano, A.; Morri, C.; Montefalcone, M. Abrupt Change in a Subtidal Rocky Reef Community Coincided with a Rapid Acceleration of Sea Water Warming. Diversity 2019, 11, 215. https://doi.org/10.3390/d11110215
Bianchi CN, Azzola A, Parravicini V, Peirano A, Morri C, Montefalcone M. Abrupt Change in a Subtidal Rocky Reef Community Coincided with a Rapid Acceleration of Sea Water Warming. Diversity. 2019; 11(11):215. https://doi.org/10.3390/d11110215
Chicago/Turabian StyleBianchi, Carlo Nike, Annalisa Azzola, Valeriano Parravicini, Andrea Peirano, Carla Morri, and Monica Montefalcone. 2019. "Abrupt Change in a Subtidal Rocky Reef Community Coincided with a Rapid Acceleration of Sea Water Warming" Diversity 11, no. 11: 215. https://doi.org/10.3390/d11110215
APA StyleBianchi, C. N., Azzola, A., Parravicini, V., Peirano, A., Morri, C., & Montefalcone, M. (2019). Abrupt Change in a Subtidal Rocky Reef Community Coincided with a Rapid Acceleration of Sea Water Warming. Diversity, 11(11), 215. https://doi.org/10.3390/d11110215