Distributional Patterns and Ecological Determinants of Bat Occurrence Inside Caves: A Broad Scale Meta-Analysis


Abstract
1. Introduction
2. Methods
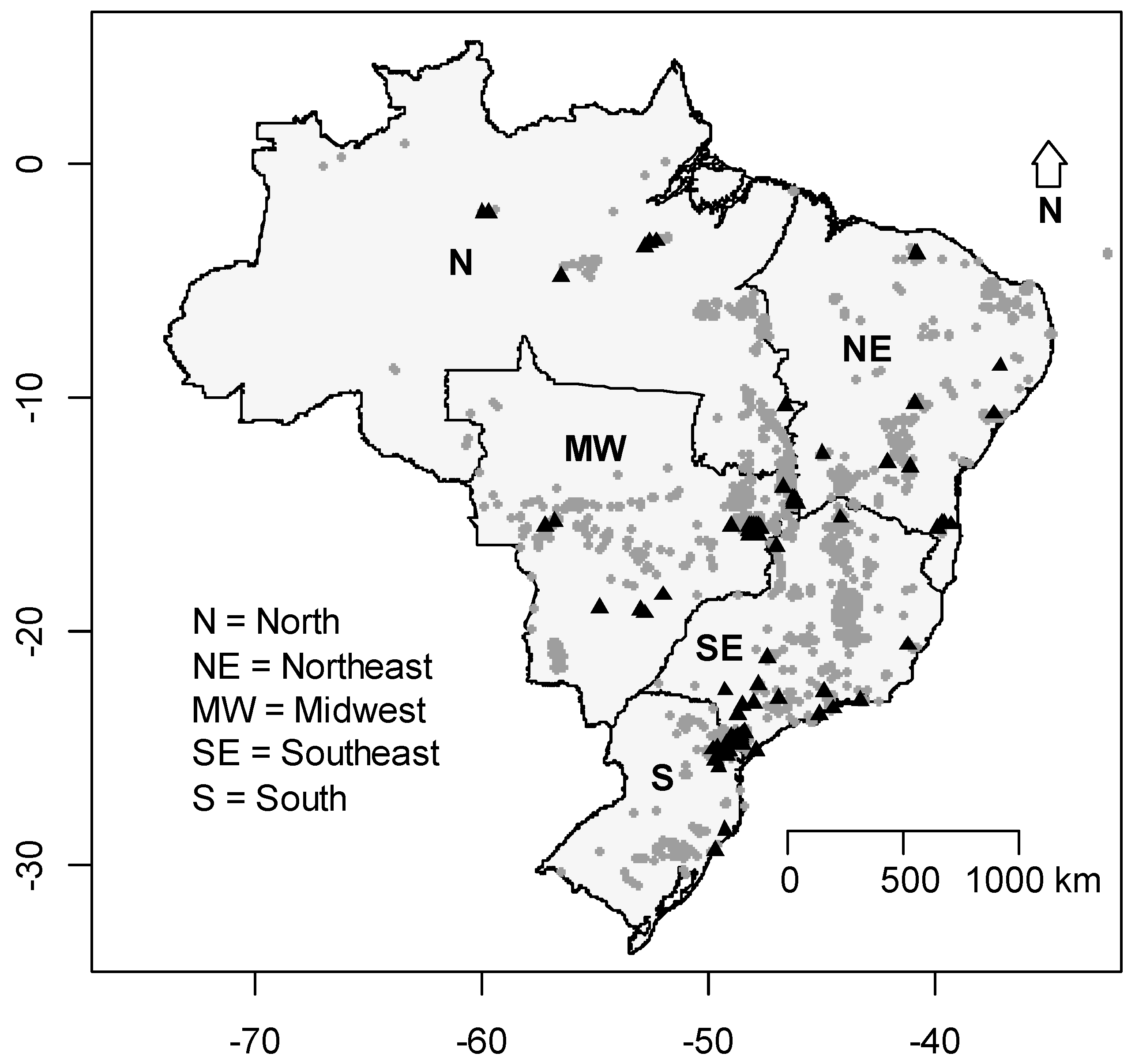
2.1. Study Area
2.2. Literature Review
2.3. Statistical Analysis
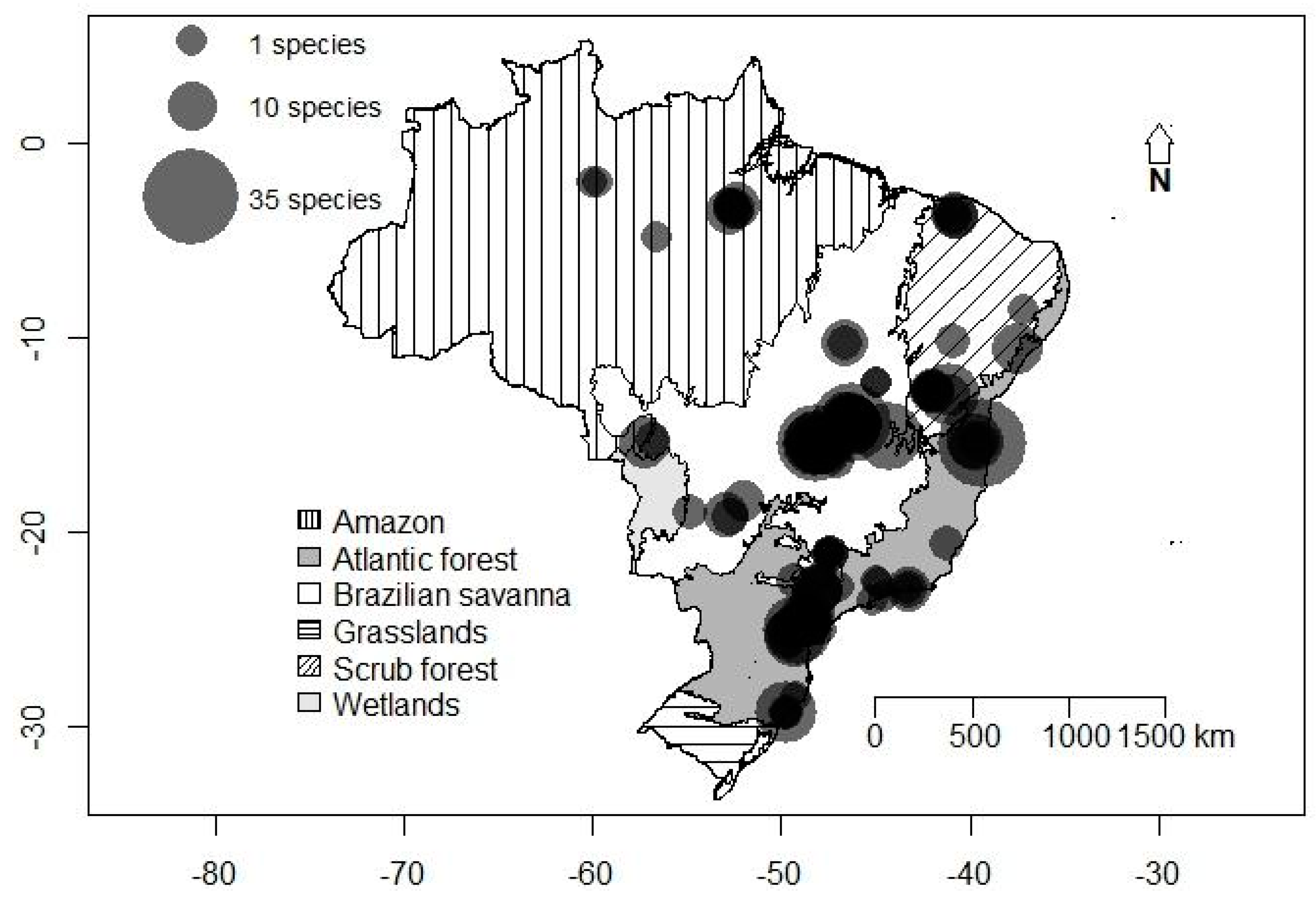
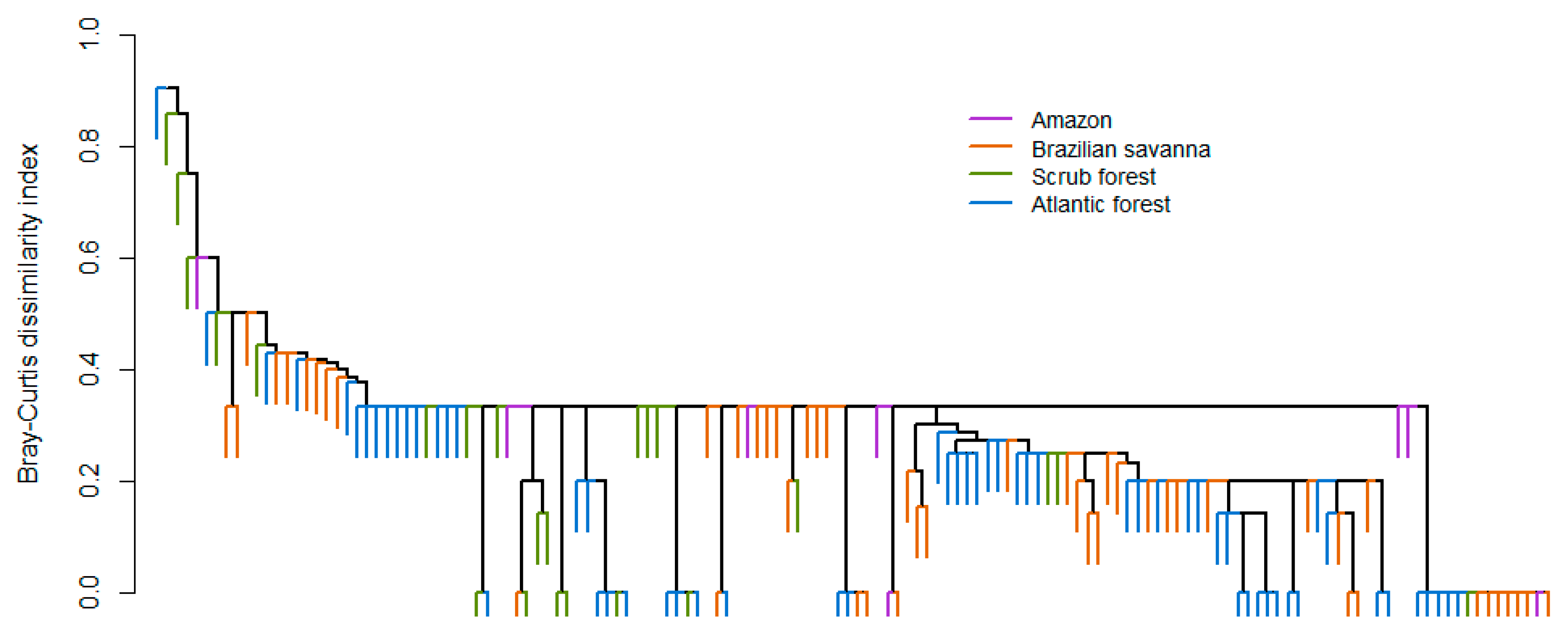
3. Results
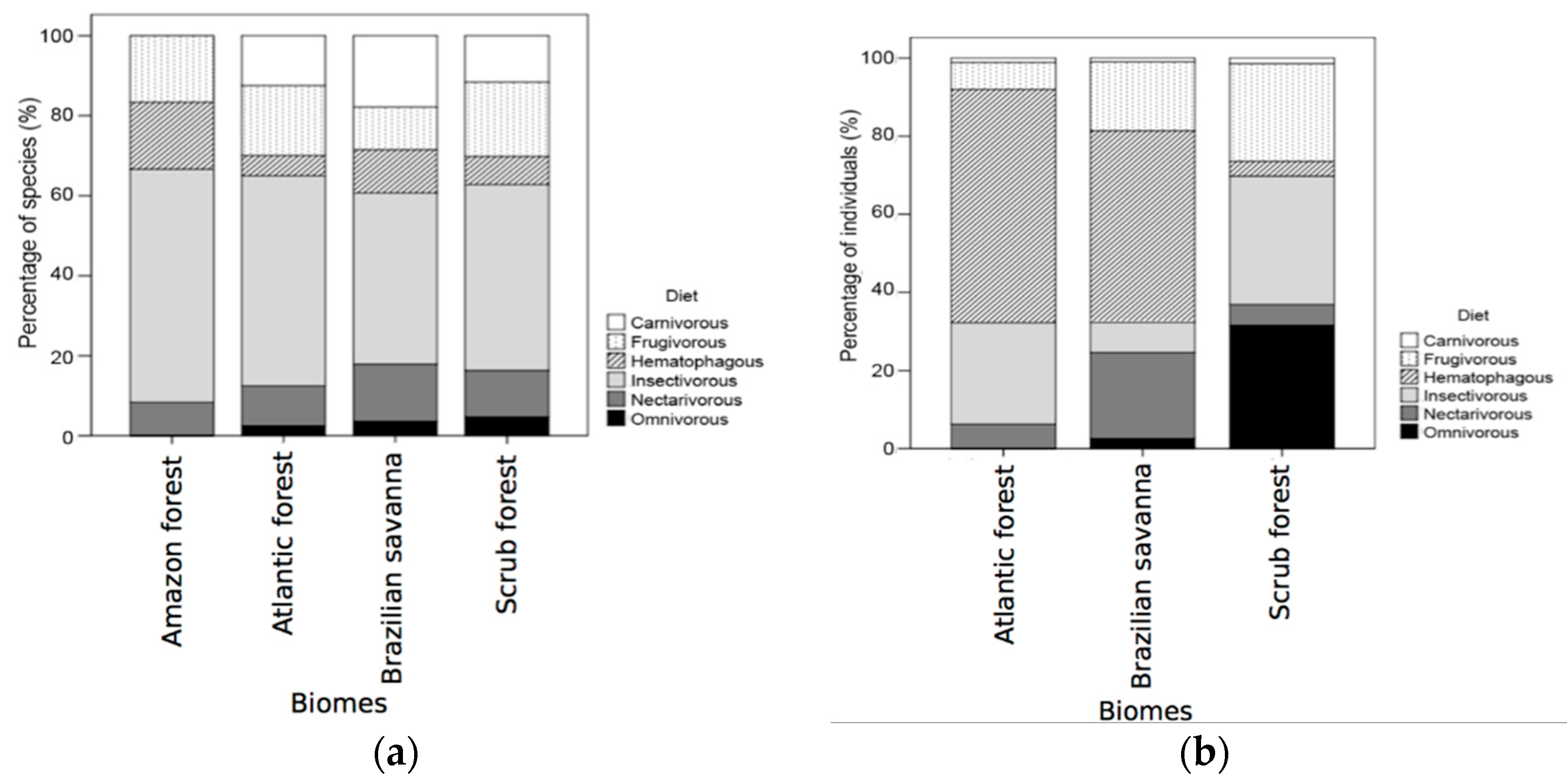
3.1. General Patterns of Cave Use
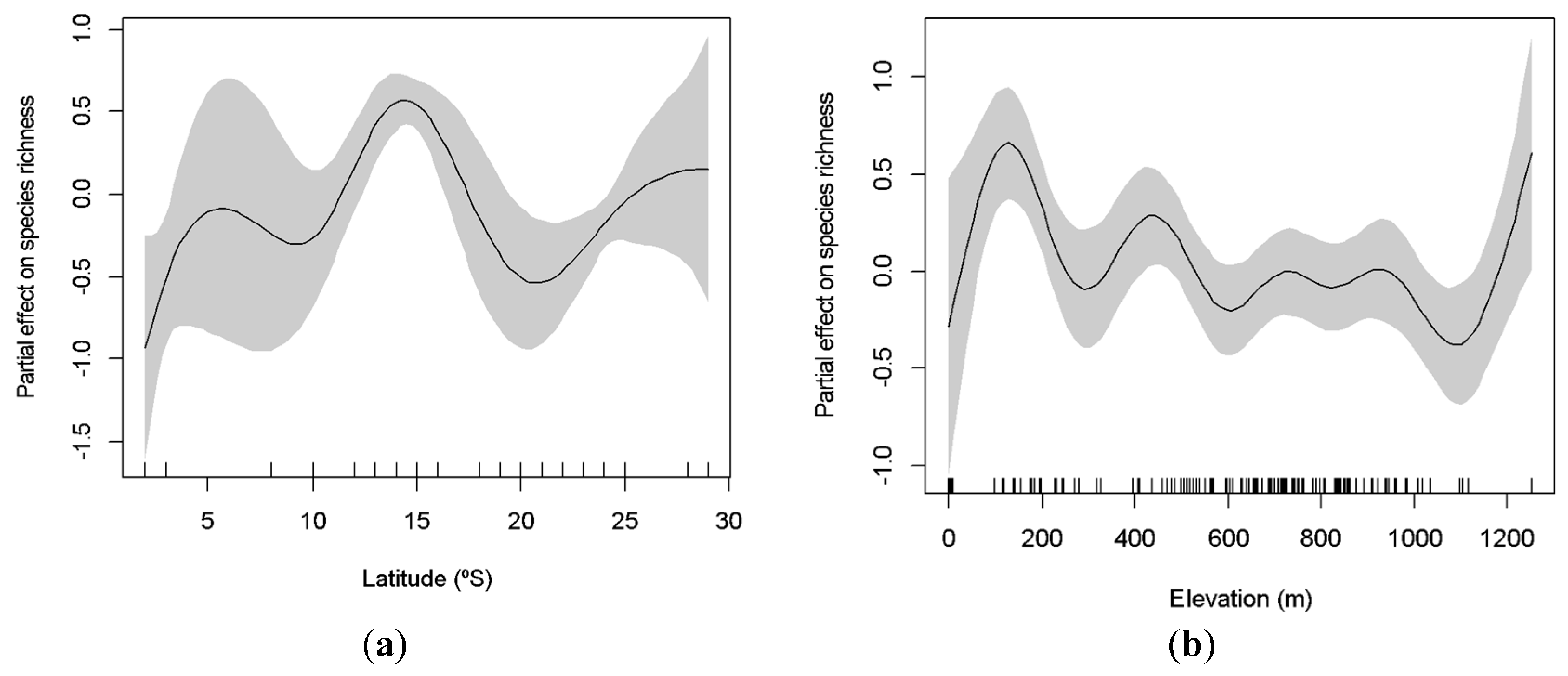
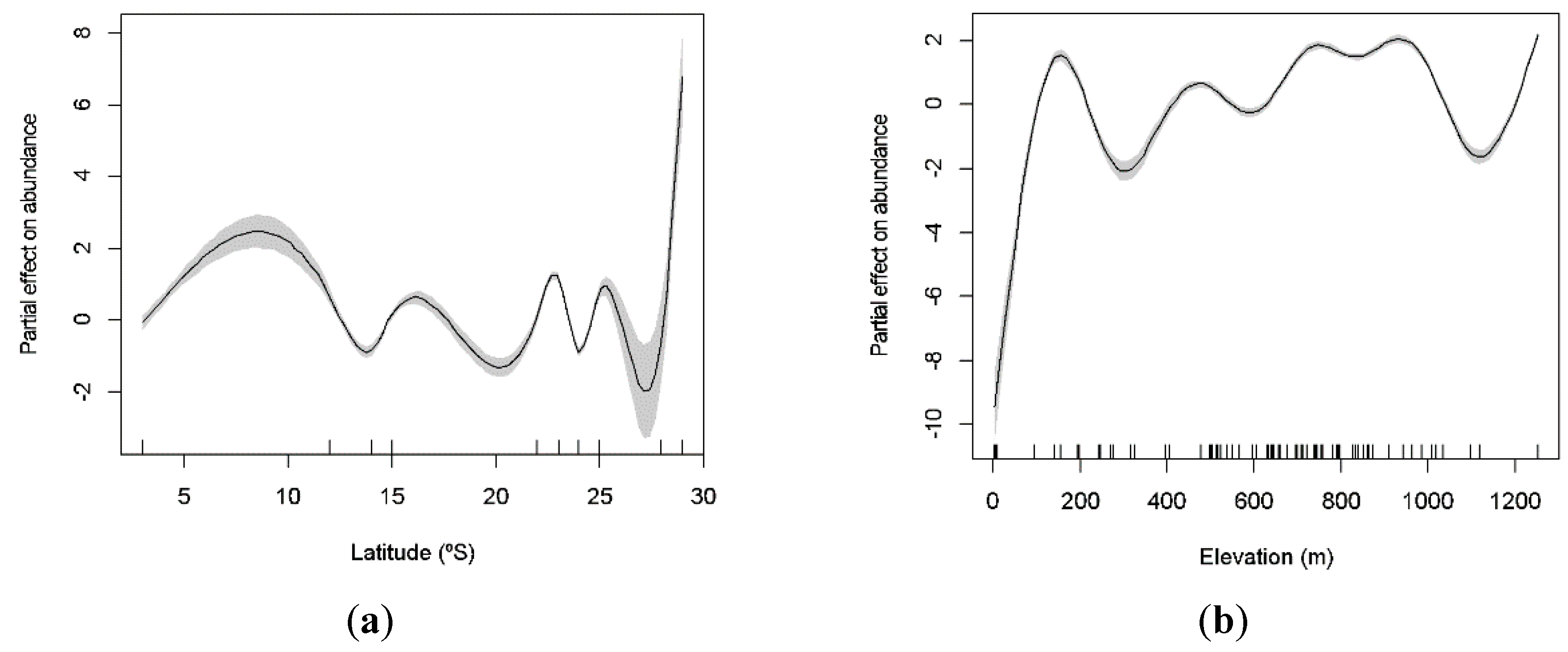
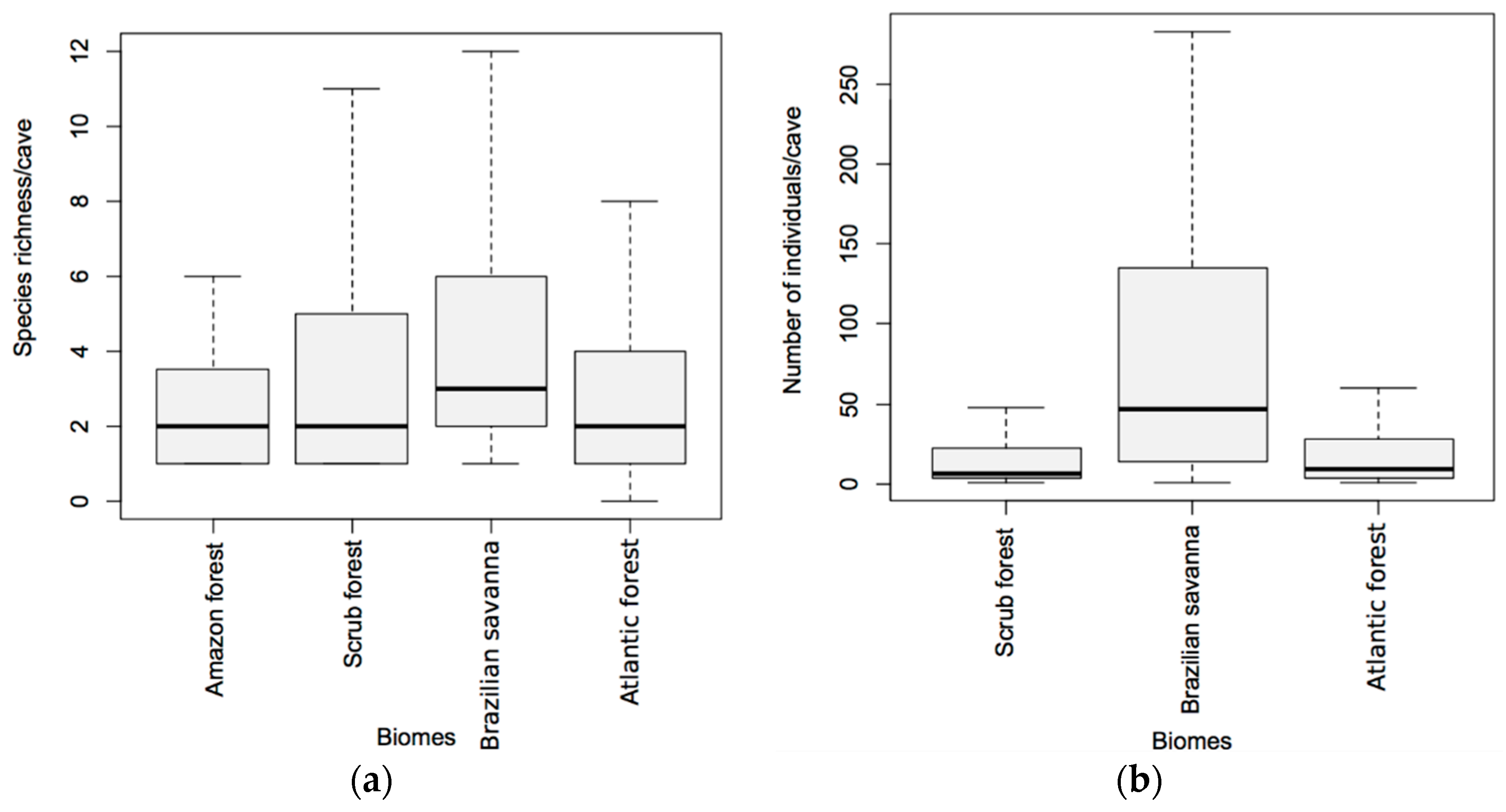
3.2. Latitudinal and Elevational Trends and Biome Effects on Assemblage Parameters
3.3. The Deficit of Knowledge and Sampling Bias
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Darwin, C. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, 1st ed.; John Murray: London, UK, 1859; p. 559. [Google Scholar]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Ricklefs, R.E. A comprehensive framework for global patterns in biodiversity. Ecol. Lett. 2004, 7, 1–15. [Google Scholar] [CrossRef]
- Shurin, J.B.; Allen, E.G. Effects of Competition, Predation, and Dispersal on Species Richness at Local and Regional Scales. Am. Nat. 2001, 158, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Willis, K.J.; Field, R. Scale and species richness: Towards a general, hierarchical theory of species diversity. J. Biogeogr. 2001, 28, 453–470. [Google Scholar] [CrossRef]
- Pianka, E.R. Latitudinal gradients in species diversity: A review of concepts. Am. Nat. 1966, 100, 33–46. [Google Scholar] [CrossRef]
- Stevens, G.C. The latitudinal gradient in geographical range: How so many species co-exist in the tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal gradients of biodiversity: pattern, process, scale, and synthesis. Ann. Rev. Ecol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Santelices, B. Phytogeographic characterization of the temperate coast of Pacific South America. Phycologia 1980, 19, 1–12. [Google Scholar] [CrossRef]
- Kouki, J.; Niemelä, P.; Viitasaari, M. Reversed latitudinal gradient in species richness of sawflies (Hymenoptera, Symphyta). Ann. Zool. Fenn. 1994, 31, 83–88. [Google Scholar] [CrossRef]
- Buckley, H.L.; Miller, T.E.; Ellison, A.M.; Gotelli, N.J. Reverse latitudinal trends in species richness of pitcher-plant food webs. Ecol. Lett. 2003, 6, 825–829. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Species diversity gradients: We know more and less than we thought. J. Mammal. 1992, 73, 715–730. [Google Scholar] [CrossRef]
- Qian, H. Large-scale biogeographic patterns of vascular plant richness in North America: An analysis at the generic level. J. Biogeogr. 1998, 25, 829–836. [Google Scholar] [CrossRef]
- Hillebrand, H. On the generality of the latitudinal diversity gradient. Am. Nat. 2004, 163, 192–211. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, G.; Ehrlich, P.R. Global mammal distributions, biodiversity hotspots, and conservation. Proc. Natl. Acad. Sci. USA 2006, 103, 19374–19379. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Orme, C.D.L.; Davies, R.G.; Burgess, M.; Eigenbrod, F.; Pickup, N.; Olson, V.A.; Webster, A.J.; Ding, T.; Rasmussen, P.C.; Ridgely, R.S.; et al. Global hotspots of species richness are not congruent with endemism or threat. Nature 2005, 436, 1016–1019. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Kelt, D.A.; Sun, Z.; Liu, H.; Hu, L.; Ren, H.; Wen, J. Global variation in elevational diversity. Nature 2013, 3, 3007. [Google Scholar] [CrossRef]
- McCain, C.M. Elevational gradients in diversity of small mammals. Ecology 2005, 86, 366–372. [Google Scholar] [CrossRef]
- McCain, C.M. Could temperature and water availability drive elevational species richness patterns? A global case study for bats. Glob. Ecol. Biogeogr. 2007, 16, 1–13. [Google Scholar] [CrossRef]
- Soares, C.; Brito, J.C. Environmental correlates for species richness among amphibians and reptiles in a climate transition area. Biodivers. Conserv. 2007, 16, 1087–1102. [Google Scholar] [CrossRef]
- Price, B.; Kutt, A.S.; McAlpine, C.A. The importance of fine-scale savanna heterogeneity for reptiles and small mammals. Biol. Conserv. 2010, 143, 2504–2513. [Google Scholar] [CrossRef]
- Al-Shami, S.A.; Rawi, C.S.M.; Ahmad, A.; Madrus, M.R.; Mutairi, K.A. Importance of regional diversity and environmental conditions on local species richness of aquatic macro-invertebrates in tropical forested streams. J. Trop. Ecol. 2014, 30, 335–346. [Google Scholar] [CrossRef]
- Ferger, S.W.; Schleuning, M.; Hemp, A.; Howell, K.M.; Bohning-Gaese, K. Food resources and vegetation structure mediate climatic effects on species richness of birds. Glob. Ecol. Biogeogr. 2014, 23, 541–549. [Google Scholar] [CrossRef]
- Kent, R.; Bar-Massada, A.; Carmel, Y. Bird and mammal species composition in distinct geographic regions and their relationships with environmental factors across multiple spatial scales. Ecol. Evol. 2014, 4, 1963–1971. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Huang, Z.Y.X.; Chen, B.J.W.; Zhang, M.; Liu, M. Can local landscape attributes explain species richness patterns at macroecological scales? Glob. Ecol. Biogeogr. 2014, 23, 436–445. [Google Scholar] [CrossRef]
- Fenton, M.B.; Simmons, N.B. Bats: A world of science and mystery. University of Chicago PressMuscarella, R.; Fleming, T.H. The role of frugivorous bats in tropical forest succession. Biol. Rev. 2007, 82, 573–590. [Google Scholar] [CrossRef]
- Kalka, M.B.; Smith, A.R.; Kalko, E.K.V. Bats limit arthropods and herbivory in a tropical forest. Science 2008, 320, 71. [Google Scholar] [CrossRef] [PubMed]
- Fleming, T.H.; Geiselman, C.; Kress, W.J. The evolution of bat pollination: A phylogenetic perspective. Ann. Bot. 2009, 104, 1017–1043. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, D.M. Diversity of New World mammals: universality of the latitudinal gradients of species and bauplans. J. Mammal. 1995, 76, 322–334. [Google Scholar] [CrossRef]
- Stevens, R.D.; Willig, M.R.; Strauss, R.E. Latitudinal gradients in the phenetic diversity of New World bat communities. Oikos 2006, 112, 41–50. [Google Scholar] [CrossRef]
- Curran, M.; Kopp, M.; Beck, J.; Fahr, J. Species diversity of bats along an altitudinal gradient on Mount Mulanje, southern Malawi. J. Trop. Ecol. 2012, 28, 243–253. [Google Scholar] [CrossRef]
- Sánchez, M.S.; Giannini, N.P. Altitudinal patterns in two syntopic species of Sturnira (Mammalia: Chiroptera: Phyllostomidae) in the montane rain forests of Argentina. Biotropica 2014, 46, 1–5. [Google Scholar] [CrossRef]
- Avila-Cabadilla, L.D.; Stoner, K.E.; Nassar, J.M.; Espírito-Santo, M.M.; Alvarez-Añorve, M.Y.; Aranguren, C.I.; Henry, M.; González-Carcacía, J.A.; Falcão, L.A.D.; Sanchez-Azofeifa, G.A. Phyllostomid bat occurrence in successional stages of Neotropical dry forests. PLoS ONE 2014, 9, e84572. [Google Scholar] [CrossRef] [PubMed]
- Arita, H.T. Conservation biology of the cave bats of Mexico. J. Mammal. 1993, 74, 693–702. [Google Scholar] [CrossRef]
- Furman, A.; Özgül, A. Distribution of cave-dwelling bats and conservation status of underground habitats in the Istanbul area. Ecol. Res. 2002, 17, 69–77. [Google Scholar] [CrossRef]
- Niu, H.; Wang, N.; Liu, J. Distribution and underground habitats of cave-dwelling bats in China. Anim. Conservat. 2007, 10, 470–477. [Google Scholar] [CrossRef]
- Glover, A.M.; Altringham, J.D. Cave selection and use by swarming bat species. Biol. Conservat. 2008, 141, 1493–1504. [Google Scholar] [CrossRef]
- Rydell, J.; Entwistle, A.; Racey, R. Timing of foraging flights of three species of bats in relation to insect activity and predation risk. Oikos 1996, 76, 243–252. [Google Scholar] [CrossRef]
- Sedgeley, J.A.; O’Donnell, C.F.J. Factors influencing the selection of roost cavities by a temperate rainforest bat (Vespertilionidae: Chalinobolus tuberculatus) in New Zealand. J. Zool. 1999, 249, 437–446. [Google Scholar] [CrossRef]
- Soriano, P.J.; Ruiz, A.; Arends, A. Physiological responses to ambient temperature manipulation by three species of bats from Andean cloud forests. J. Mammal. 2002, 83, 445–457. [Google Scholar] [CrossRef]
- Vasseur, D.A.; DeLong, J.P.; Gilbert, B.; Greig, H.S.; Harley, C.D.; McCann, K.S.; Savage, V.; Tunney, T.D.; O’Connor, M.I. Increased temperature variation poses a greater risk to species than climate warming. Proc. R. Soc. Lond. B Biol. Sci. 2014, 281, 20132612. [Google Scholar] [CrossRef] [PubMed]
- Ohlemüller, R.; Wilson, J.B. Vascular plant species richness along latitudinal and altitudinal gradients: A contribution from New Zealand temperate rainforests. Ecol. Lett. 2000, 3, 262–266. [Google Scholar] [CrossRef]
- Hamilton, I.M.; Barclay, R.M. Patterns of daily torpor and day-roost selection by male and female big brown bats (Eptesicus fuscus). Can. J. Zool. 1994, 72, 744–749. [Google Scholar] [CrossRef]
- Vonhof, M.J.; Barclay, R.M. Roost-site selection and roosting ecology of forest-dwelling bats in southern British Columbia. Can. J. Zool. 1996, 74, 1797–1805. [Google Scholar] [CrossRef]
- Callahan, E.V.; Drobney, R.D.; Clawson, R.L. Selection of summer roosting sites by Indiana bats (Myotis sodalis) in Missouri. J. Mammal. 1997, 78, 818–825. [Google Scholar] [CrossRef]
- Arita, H.T.; Vargas, J.A. Natural history, interspecific association, and incidence of the cave bats of Yucatan, Mexico. Southwest. Nat. 1995, 40, 29–37. [Google Scholar]
- Grytnes, J.A.; McCain, C.M. Elevational trends in biodiversity. In Encyclopedia of Biodiversity, 1st ed.; Levin, S.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 1, pp. 1–8. ISBN 9780122268656. [Google Scholar]
- MMA—Ministério do Meio Ambiente. Available online: http://mma.gov.br/ (accessed on 5 June 2016).
- Auler, A.; Farrant, A.R. A brief introduction to karst and caves in Brazil. Proc. Univ. Bristol Speleol. Soc. 1996, 20, 187–200. [Google Scholar]
- CECAV—Database of Caves of Brazil. Available online: http://www.icmbio.gov.br/cecav/downloads/mapas.html (accessed on 15 July 2014).
- Simmons, N.B. Chiroptera. In Mammal Species of the World–A Taxonomic and Geographic Reference, 3rd ed.; Wilson, D.E., Reeder, D.M., Eds.; Hopkins University Press: Baltimore, MD, USA, 2005; Volume 1, pp. 312–529. ISBN 978-0801882210. [Google Scholar]
- Nogueira, M.R.; de Lima, I.P.; Moratelli, R.; da Cunha Tavares, V.; Gregorin, R.; Peracchi, A.L. Checklist of Brazilian bats, with comments on original records. Check List 2014, 10, 808–821. [Google Scholar] [CrossRef]
- Simmons, N.B.; Voss, R.S. The mammals of Paracou, French Guiana, a Neotropical lowland rainforest fauna. Part 1, Bats. Bull. Am. Mus. Nat. Hist. 1998, 237, 1–218. [Google Scholar]
- Reis, N.R.; Peracchi, A.L.; Pedro, W.A.; Lima, I.P. Morcegos do Brasil, 1st ed.; Univesidade Estadual de Londrina: Londrina, Basil, 2007; ISBN 978-85-906395-1-0. [Google Scholar]
- Emmons, L.H.; Feer, F. Neotropical Rainforest Mammals: A Field Guide, 2nd ed.; The University of Chicago Press: Chicago, IL, USA, 1997; ISBN 978-0226207216. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: www.R-project.org (accessed on 4 April 2018).
- Wood, S. Generalized Additive Models—An Introduction with R; Chapman & Hall: London, UK, 2006; ISBN 978-1584884743. [Google Scholar]
- Crawley, M.J. The R Book; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2007. [Google Scholar]
- O’Hara, R.B.; Kotze, D.J. Do not log-transform count data. Methods Ecol. Evol. 2010, 1, 118–122. [Google Scholar] [CrossRef]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002; ISBN 978-0521009768. [Google Scholar]
- Pinheiro, J.C.; Bates, D.M. Fitting linear mixed-effects models. In Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2000; pp. 133–199. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan’—Community Ecology Package. R package version 1.15-1. Available online: http://vegan.r-forge.r-project.org/ (accessed on 4 January 2013).
- Currie, D.J.; Mittelbach, G.G.; Cornell, H.V.; Field, R.; Guegan, J.F.; Hawkins, B.A.; Kaufman, D.M.; Kerr, J.T.; Oberdorff, T.; O’ Brien, E.; et al. Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol. Lett. 2004, 7, 1121–1134. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Schemske, D.W.; Cornell, H.V.; Allen, A.P.; Brown, J.M.; Bush, M.B.; Harrison, S.P.; Hurlbert, A.H.; Knowlton, N.; Lessios, H.A.; et al. Evolution and the latitudinal diversity gradient: speciation, extinction and biogeography. Ecol. Lett. 2007, 10, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Durán, A. Metabolic rates and thermal conductance in 4 species of Neotropical bats roosting in hot caves. Comp. Biochem. Physiol. A 1995, 110, 347–355. [Google Scholar] [CrossRef]
- Baudinette, R.V.; Churchill, S.K.; Christian, K.A.; Nelson, J.E.; Hudson, P.J. Energy, water balance and the roost microenvironment in three Australian cave-dwelling bats (Microchiroptera). J. Comp. Physiol. B 2000, 170, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Graham, G.L. Bats vs. birds: Comparisons among Peruvian volant vertebrate faunas along an elevational gradient. J. Biogeogr. 1990, 17, 657–668. [Google Scholar] [CrossRef]
- Cisneros, L.M.; Burgio, K.R.; Dreiss, L.M.; Klingbeil, B.T.; Patterson, B.D.; Presley, S.J.; Willig, M.R. Multiple dimensions of bat biodiversity along an extensive tropical elevational gradient. J. Anim. Ecol. 2014, 83, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.D.; Law, B.S.; French, K.O. Vegetation structure influences the vertical stratification of open- and edge-space aerial-foraging bats in harvested forests. For. Ecol. Manag. 2009, 258, 2090–2100. [Google Scholar] [CrossRef]
- Bolívar-Cimé, B.; Laborde, J.; MacSwiney, M.C.; Muñoz-Robles, C.; Tun-Garrido, J. Response of phytophagous bats to patch quality and landscape attributes in fragmented tropical semi-deciduous forest. Acta Chiropterol. 2013, 15, 399–409. [Google Scholar] [CrossRef]
- Aguirre, L.F.; Lens, L.; Matthysen, E. Patterns f roost use by bats in a Neotropical savanna: implications for conservation. Biol. Conserv. 2013, 111, 435–443. [Google Scholar] [CrossRef]
- Avila-Flores, R.; Medellín, R. Ecological, taxonomic, and physiological correlates of cave use by Mexican bats. J. Mammal. 2004, 85, 675–687. [Google Scholar] [CrossRef]
- Paglia, A.P.; Fonseca, G.A.B.; Rylands, A.B.; Herrmann, G.; Aguiar, L.M.S.; Chiarello, A.G.; Leite, Y.L.R.; Costa, L.P.; Siciliano, S.; Kierulff, M.C.M.; et al. Annotated Checklist of Brazilian Mammals, 2nd ed.; Conservação Internacional do Brasil: Belo Horizonte, Brazil, 2012; ISBN 978-1-934151-49-5. [Google Scholar]
- Kunz, T.H. Roosting ecology of bats. In Ecology of Bats, 1st ed.; Kunz, T.H., Ed.; Plenum Press: New York, NY, USA, 1982; Volume 1, pp. 1–55. ISBN 9780306409509. [Google Scholar]
- Elgar, M.A.; Harvey, H. Basal metabolic rates in mammals: allometry, phylogeny and ecology. Funct. Ecol. 1987, 1, 25–36. [Google Scholar] [CrossRef]
- Lyman, C.P. Thermoregulation and metabolism in bats. In Biology of Bats, 1st ed.; Wimsatt, W.A., Ed.; Academic Press: New York, NY, USA, 2012; Volume 1, pp. 301–330. ISBN 978-0124313187. [Google Scholar]
- McNab, B.K. The influence of food habits on the energetics of eutherian mammals. Ecol. Monogr. 1986, 56, 1–19. [Google Scholar] [CrossRef]
- Speakman, J.R.; Webb, P.I.; Racey, P.A. Effects of disturbance on the energy expenditure of hibernating bats. J. Appl. Ecol. 1991, 28, 1087–1104. [Google Scholar] [CrossRef]
- Mann, S.L.; Steidl, R.J.; Dalton, V.M. Effects of cave tours on breeding Myotis velifer. J. Wildl. Manag. 2002, 66, 618–624. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Abundance (% of Individuals Belonging to the Families) | Richness of Species that Had Their Abundance Registered | Total Richness Registered | Richness of Species Registered in Brazil (% of Species of the Family that Use Caves as a Roost) |
|---|---|---|---|---|
| Phyllostomidae | 8693 (89.9) | 35 | 39 | 92 (42.4) |
| Furipteridae | 490 (5.1) | 1 | 1 | 1 (100.0) |
| Vespertilionidae | 170 (1.7) | 5 | 10 | 28 (35.7) |
| Mormoopidae | 129 (1.3) | 2 | 5 | 3 (100.0) |
| Emballonuridae | 92 (0.9) | 3 | 4 | 17 (23.5) |
| Natalidae | 79 (0.8) | 1 | 1 | 1 (100.0) |
| Noctilionidae | 10 (0.0) | 1 | 1 | 2 (50.0) |
| Molossidae | 2 (0.0) | 1 | 10 | 29 (34.5) |
| Thyropteridae | 1 (0.0) | 1 | 1 | 5 (20.0) |
| Total | 9666 (100.0) | 50 | 72 | 178 (40.4) |
| Feeding Guild | Total Richness | Richness with the Abundance Registered | Individuals | Biomass (g) |
|---|---|---|---|---|
| Hematophagous | 4 | 3 | 4011 | 130,703.5 |
| Omnivorous | 2 | 2 | 546 | 48,840.0 |
| Frugivorous | 12 | 11 | 1182 | 29,694.5 |
| Nectarivorous | 7 | 7 | 1907 | 24,793.4 |
| Insectivorous | 41 | 21 | 1823 | 16,796.8 |
| Carnivorous | 6 | 6 | 179 | 8193.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Oliveira, H.F.M.; Oprea, M.; Dias, R.I. Distributional Patterns and Ecological Determinants of Bat Occurrence Inside Caves: A Broad Scale Meta-Analysis. Diversity 2018, 10, 49. https://doi.org/10.3390/d10030049
De Oliveira HFM, Oprea M, Dias RI. Distributional Patterns and Ecological Determinants of Bat Occurrence Inside Caves: A Broad Scale Meta-Analysis. Diversity. 2018; 10(3):49. https://doi.org/10.3390/d10030049
Chicago/Turabian StyleDe Oliveira, Hernani Fernandes Magalhães, Monik Oprea, and Raphael Igor Dias. 2018. "Distributional Patterns and Ecological Determinants of Bat Occurrence Inside Caves: A Broad Scale Meta-Analysis" Diversity 10, no. 3: 49. https://doi.org/10.3390/d10030049
APA StyleDe Oliveira, H. F. M., Oprea, M., & Dias, R. I. (2018). Distributional Patterns and Ecological Determinants of Bat Occurrence Inside Caves: A Broad Scale Meta-Analysis. Diversity, 10(3), 49. https://doi.org/10.3390/d10030049