Anthropogenic Impacts on Coral Reef Harpacticoid Copepods

 ,
,
Abstract
1. Introduction
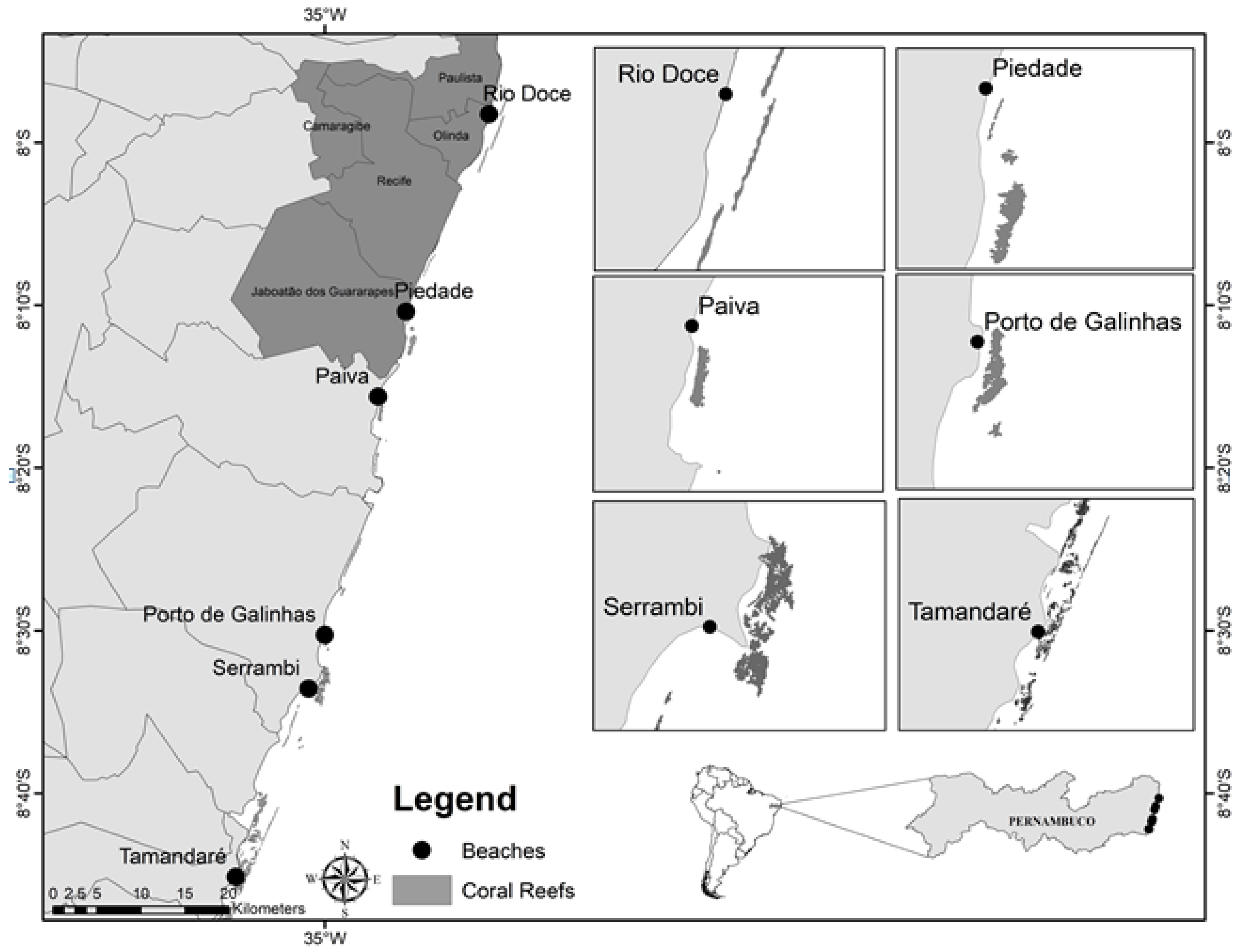
2. Materials and Methods
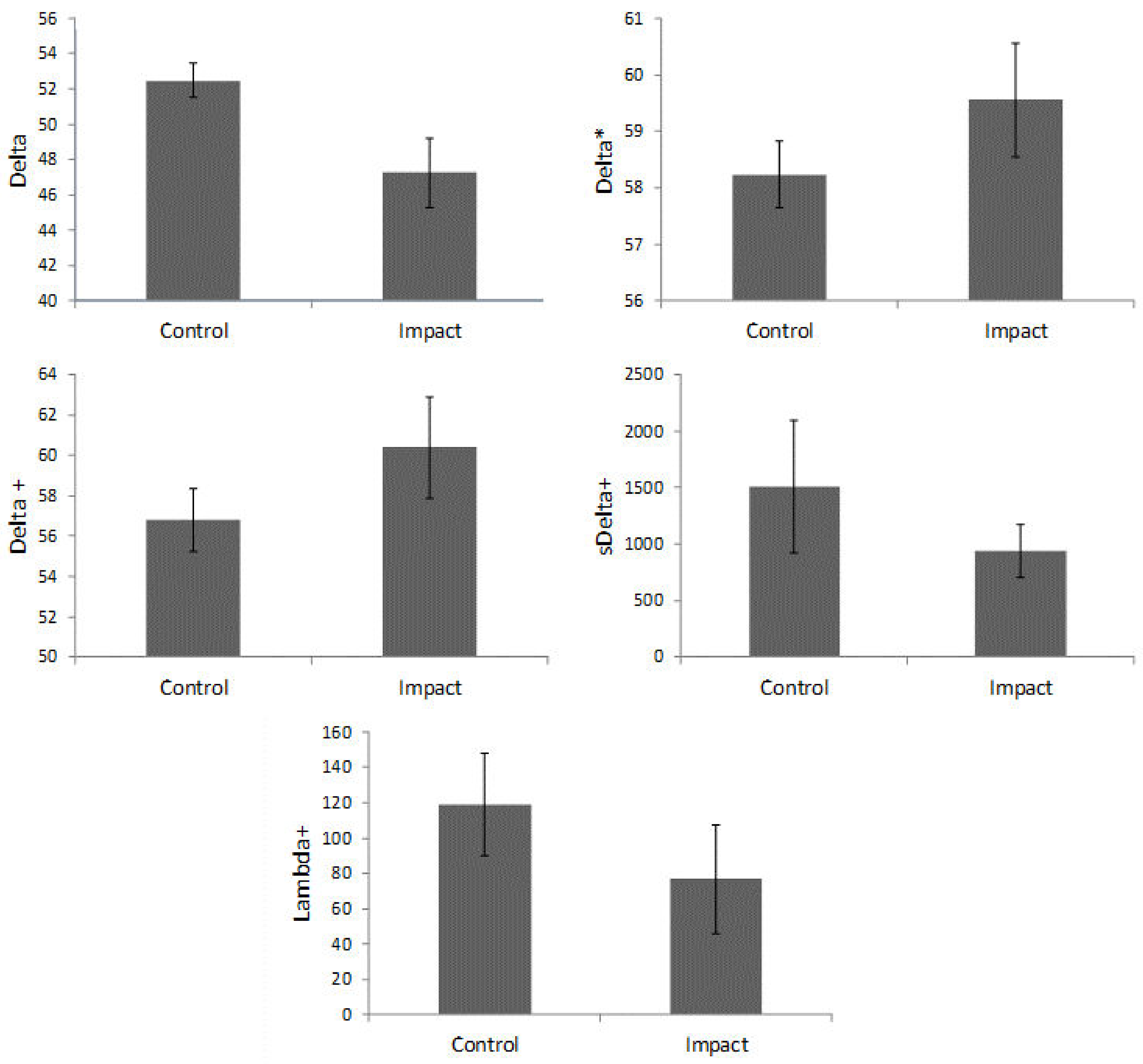
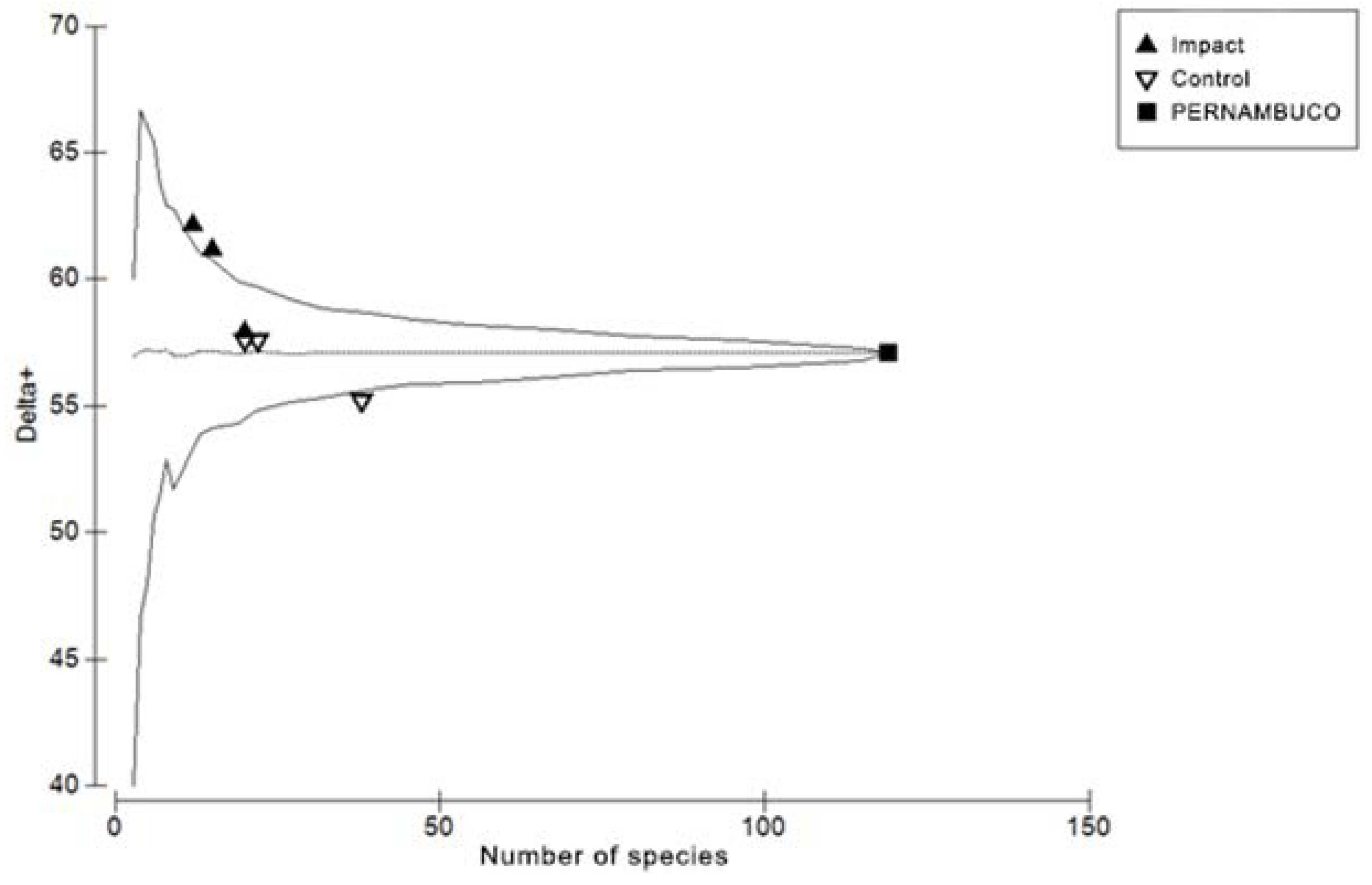
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, A.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [PubMed]
- Zeppilli, D.; Sarrazin, J.; Leduc, D.; Arbizu, P.M.; Fontaneto, D.; Fontanier, C.; Gooday, A.J.; Kristensen, R.M.; Ivanenko, V.N.; Sørensen, M.V.; et al. Is the meiofauna a good indicator for climate change and anthropogenic impacts? Mar. Biodiv. 2015, 45, 505–535. [Google Scholar] [CrossRef]
- Borja, A.; Muxika, I. Do benthic indicator tools respond to all impact sources? The case of AMBI (AZTI Marine Biotic Index). In Proceedings of the Workshop Indicators of Stress in the Marine Benthos, Torregrande-Oristano, Italy, 8–9 October 2004; Magni, P., Hyland, J., Manzella, G., Rumohr, H., Viaroli, P., Zenetos, A., Eds.; Intergovernmental Oceanographic Commission: Paris, France, 2005; pp. 15–18. [Google Scholar]
- Leao, Z.M.; Kikuchi, R.K. A relic coral fauna threatened by global changes and human activities, Eastern Brazil. Mar. Pollut. Bull. 2005, 51, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Defeo, O.; Rueda, M. Spatial structure, sampling design and abundance estimates in sandy beach macroinfauna: Some warnings and new perspectives. Mar. Biol. 2002, 140, 1215–1225. [Google Scholar] [CrossRef]
- Van Andel, T.H. Science at Sea: Tales of an Old Ocean; W.H. Freeman & Co. Ltd.: San Francisco, CA, USA, 1981; ISBN 9780716713647. [Google Scholar]
- Gingold, R.; Moens, T.; Rocha-Olivares, A. Assessing the Response of Nematode Communities to Climate Change-Driven Warming: A Microcosm Experiment. PLoS ONE 2013, 8, e66653. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Sarmento, V.C.; Souza, T.P.; Esteves, A.M.; Santos, P.J.P. Effects of seawater acidification on a coral reef meiofauna community. Coral Reefs 2015, 34, 955–966. [Google Scholar] [CrossRef]
- Valença, A.P.M.; Santos, P.J. Macrobenthic community for assessment of estuarine health in tropical areas (Northeast, Brazil): Review of macrofauna classification in ecological groups and application of AZTI Marine Biotic Index. Mar. Pollut. Bull. 2012, 64, 1809–1820. [Google Scholar] [CrossRef] [PubMed]
- Myers, N. Mass extinction and evolution. Science 1997, 278, 597–598. [Google Scholar] [CrossRef]
- Baskin, Y. Winners and losers in a changing world. BioScience 1998, 48, 788–792. [Google Scholar] [CrossRef]
- Fabricius, K.E.; Langdon, C.; Uthicke, S.; Humphrey, C.; Noonan, S.; De’ath, G.; Lough, J.M. Losers and winners in coral reefs acclimatized to elevated carbon dioxide concentrations. Nat. Clim. Chang. 2011, 1, 165–169. [Google Scholar] [CrossRef]
- Tabarelli, M.; Carlos, A.P.; Felipe, P.L.M. The ‘few winners and many losers’ paradigm revisited: Emerging prospects for tropical forest biodiversity. Biol. Conserv. 2012, 155, 136–140. [Google Scholar] [CrossRef]
- McKinney, M.L.; Lockwood, J.L. Biotic homogenization: A few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 1999, 14, 450–453. [Google Scholar] [CrossRef]
- Leão, Z.M.N.; Dominguez, J.M.L. Tropical coast of Brazil. Mar. Pollut. Bull. 2000, 41, 112–122. [Google Scholar] [CrossRef]
- Ferreira, M.N.; Rosso, S. Effects of human trampling on a rocky shore fauna on the Sao Paulo coast, southeastern Brazil. Braz. J. Biol. 2009, 69, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Leão, Z.M.N.; Kikuchi, R.K.P.; Oliveira, M.D.M.; Vasconcellos, V. Status of Eastern Brazilian coral reefs in time of climate changes. Pan-Am. J. Aquat. Sci. 2011, 5, 224–235. [Google Scholar]
- Moberg, F.; Folke, C. Ecological goods and services of coral reef ecosystems. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Dunning, K.H. Ecosystem services and community based coral reef management institutions in post blast-fishing Indonesia. Ecosyst. Serv. 2015 16, 319–332.
- Sarmento, V.C.; Barreto, A.F.; Santos, P.J. The response of meiofauna to human trampling on coral reefs. Sci. Mar. 2011, 75, 559–570. [Google Scholar] [CrossRef]
- Leão, Z.M.; Kikuchi, R.K.; Ferreira, B.P.; Neves, E.G.; Sovierzoski, H.H.; Oliveira, M.D.; Maida, M.; Correia, M.D.; Johnsson, R. Brazilian coral reefs in a period of global change: A synthesis. Braz. J. Oceanogr. 2016, 64, 97–116. [Google Scholar] [CrossRef]
- Mirto, S.; Danovaro, R. Meiofaunal colonisation on artificial substrates: A tool for biomonitoring the environmental quality on coastal marine systems. Mar. Pollut. Bull. 2004, 48, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Bishop, M.J. Artificial sampling units: A tool for increasing the sensitivity of tests for impact in soft sediments. Environ. Monit. Assess. 2005, 107, 203–220. [Google Scholar] [CrossRef] [PubMed]
- De Troch, M.; Vandepitte, L.M.; Raes, M.; Suárez-Morales, E.; Vincx, M. A field colonization experiment with meiofauna and seagrass mimics: Effect of time, distance and leaf surface area. Mar. Biol. 2005, 148, 73–86. [Google Scholar] [CrossRef]
- Rule, M.J.; Smith, S.D.A. Depth-associated patterns in the development of benthic assemblages on artificial substrata deployed on shallow, subtropical reefs. J. Exp. Mar. Biol. Ecol. 2007, 345, 38–51. [Google Scholar] [CrossRef]
- Hicks, G.R.; Geoffrey, R.F.; Bruce, C. The ecology of marine meiobenthic harpacticoid copepods. Oceanogr. Mar. Biol. Anu. Rev. 1983, 21, 67–175. [Google Scholar]
- Coull, B.C. Ecology of the marine meiofauna. In Introduction to the Study of Meiofauna; Higgins, R.P., Thiel, H., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1988; Volume 132, pp. 18–38. ISBN 0-87474-488-1. [Google Scholar]
- Danovaro, R.; Scopa, M.; Gambi, C.; Franschetti, S. Trophic importance of subtidal metazoan meiofauna: Evidence from in situ exclusion experiments on soft and rocky substrates. Mar. Biol. 2007, 152, 339–350. [Google Scholar] [CrossRef]
- Danovaro, R.; Corinaldesi, C.; Dell’Anno, A.; Snelgrove, P.V.R. The deep-sea under global change. Curr. Biol. 2017, 27, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Sarmento, V.C.; Santos, P.J.P. Trampling on coral reefs: Tourism effects on harpacticoid copepods. Coral Reefs 2012, 31, 135–146. [Google Scholar] [CrossRef]
- Costa, A.B.H.P.; Valença, A.P.M.C.; Santos, P.J.P. Is meiofauna community structure in Artificial Substrate Units a good tool to assess anthropogenic impact in estuaries? Mar. Poll. Bull. 2016, 110, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Warwick, R.M.; Clarke, K.R. New “biodiversity” measures reveal a decrease in taxonomic distinctness with increasing stress. Mar. Ecol. Prog. Ser. 1995, 129, 301–305. [Google Scholar] [CrossRef]
- Piepenburg, D.; Voss, J.; Gutt, J. Assemblages of sea stars (Echinodermata: Asteroidea) and brittle stars (Echinodermata: Ophiuroidea) in the Weddell Sea (Antarctica) and off Northeast Greenland (Arctic): A comparison of diversity and abundance. Polar Biol. 1997, 17, 305–322. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. A taxonomic distinctness index and its statistical properties. J. Appl. Ecol. 1998, 35, 523–531. [Google Scholar] [CrossRef]
- Hall, S.J.; Greenstreet, S.P. Taxonomic distinctness and diversity measures: Responses in marine fish communities. Mar. Ecol. Prog. Ser. 1998, 166, 227–229. [Google Scholar] [CrossRef]
- Price, N.N.; Hamilton, S.L.; Tootell, J.S.; Smith, J.E. Species-specific consequences of ocean acidification for the calcareous tropical green algae Halimeda. Mar. Ecol. Prog. Ser. 2011, 440, 67–78. [Google Scholar] [CrossRef]
- Rogers, S.I.; Clarke, K.R.; Reynolds, J.D. The taxonomic distinctness of costal bottom-dweling fish communities of the North East Atlantic. J. Anim. Ecol. 1999, 68, 769–788. [Google Scholar] [CrossRef]
- Warwick, R.M.; Light, J. Death assemblages of molluscs on St Martin’s Flats, Isles of Scilly: A surrogate for regional biodiversity? Biodiv. Conserv. 2002, 11, 99–112. [Google Scholar] [CrossRef]
- Warwick, R.M.; Turk, S.M. Predicting climate change effects on marine biodiversity: Comparison of recent and fossil molluscan death assemblages. J. Mar. Biol. Assoc. UK 2002, 82, 847–850. [Google Scholar] [CrossRef]
- Warwick, R.M.; Ashman, C.M.; Brown, A.R.; Clarke, K.R.; Dowell, B.; Hart, B.; Lewis, R.E.; Shillabeer, N.; Somerfield, P.J.; Tapp, J.F. Inter-annual changes in the biodiversity and community structure of the macrobenthos in Tees Bay and the Tees estuary, UK, associated with local and regional environmental events. Mar. Ecol. Progr. Ser. 2002, 234, 1–13. [Google Scholar] [CrossRef]
- Grasshoff, K.; Ehrhardt, M.; Kremling, K. Methods of Seawater Analysis, 2nd ed.; Verlag Chemie: New York, NY, USA, 1983; 419p, ISBN 3527259988. [Google Scholar]
- Strickland, J.D.; Parsons, T.R. A Practical Handbook of Seawater Analysis; Bulletin 167; Fisheries Research Board of Canada: Ottawa, Canada, 1972. [Google Scholar]
- United Nations Educational, Scientific and Cultural Organization (UNESCO). International Oceanographic Table. Wormly 1973, 2, 1–141. [Google Scholar]
- Dickson, A.G.; Sabine, C.L.; Christian, J.R. Guide to Best Practices for Ocean CO2 Measurements; PICES Special Publication; North Pacific Marine Science Organization: Sidney, BC, Canada, 2007; Volume 3, pp. 1–191. ISBN 1897176074. [Google Scholar]
- Sarmento, V.C.; Pinheiro, B.R.; Montes, M.J.F.; Santos, P.J.P. Impact of predicted climate change scenarios on a coral reef meiofauna community. ICES J. Mar. Sci. 2017, 74, 1170–1179. [Google Scholar] [CrossRef]
- Santos, P.J.P.; Castel, J.; Souza-Santos, L.P. Development time of harpacticoid copepods: Some empirical models and implications. J. Mar. Biol. Assoc. UK 1999, 79, 1123–1124. [Google Scholar] [CrossRef]
- Sarmento, V.C.; Santos, P.J.P.; Hale, R.; Ingels, J.; Widdicombe, S. Effects of elevated CO2 and temperature on an intertidal harpacticoid copepod community. ICES J. Mar. Sci. 2017, 74, 1159–1169. [Google Scholar]
- Lang, K. Monographie der Harpacticiden; Nordiska Bökhandeln: Stockholm, Sweden, 1948; pp. 1–1648. ISBN 3874290891, 9783874290890. [Google Scholar]
- Huys, R.; Gee, J.M.; Moore, C.G.; Hamond, R. Marine and brackish water harpacticoid Copepods. Part 1: Keys and notes for identification of the species. In Synopses of the British Fauna (New Series) no. 51; Barnes, R.S.K., Crothers, J.H., Eds.; Field Studies Council: Shrewsbury, UK, 1996; pp. 1–352. ISBN 978-1851532568. [Google Scholar]
- Wells, J.B.J. An annotated checklist and keys to the species of Copepoda Harpacticoida (Crustacea). Zootaxa 2007, 1568, 1–872. [Google Scholar]
- Barreto, A.F. Viability of ASU (Artificial Substrate Unit) in the Evaluation of Anthropic Pressure over the Assemblages of Meiofauna and Copepoda Harpacticoida in Reef Environments. Master’s Thesis, Universidade Federal de Pernambuco, Recife, Brazil, 2014. (In Portuguese). [Google Scholar]
- Nascimento, R.F.M. Temporal Variation of Phytal Meiofauna and Copepoda Harpacticoida Communities in a Climate Change Scenario. Master’s Thesis, Universidade Federal de Pernambuco, Recife, Brazil, 2015. (In Portuguese). [Google Scholar]
- Clarke, K.R.; Warwick, R.M. A further biodiversity index applicable to species lists: Variation in taxonomic distinctness. Mar. Ecol. Prog. Ser. 2001, 216, 265–278. [Google Scholar] [CrossRef]
- Salas, F.; Marcosa, C.; Neto, J.M.; Patrício, J.; Pérez-Ruzafa, A.; Marques, J.C. User friendly guide for using benthic ecological indicators in coastal and marine quality assessment. Ocean Coast. Manag. 2006, 49, 308–331. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; Natural Environment Research Council (NERC): Plymouth, UK, 1994; pp. 1–187. ISBN 9781855311404. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 3rd ed.; Prentice-Hall: Upper Saddle River, NJ, USA, 1996; pp. 1–662. ISBN 9780130845429. [Google Scholar]
- Bastos, R.B. Caracterização de uma zona costeira tropical (Ipojuca-Pernambuco-Brasil): Produtividade fitoplanctônica e outras variáveis ambientais. Braz. J. Aquat. Sci. Technol. 2011, 15, 1–10. [Google Scholar] [CrossRef][Green Version]
- Machado, R.C.A. Phytoplankton productivity and hydrology of Porto de Galinhas reef ecosystem (Pernambuco, Brazil). J. Coast. Res. 2013, 30, 371–378. [Google Scholar] [CrossRef]
- Grego, C.K.S.; Feitosa, F.A.N.; Silva, M.H.; Flores-Montes, M.J. Distribuição espacial e sazonal da clorofila-a fitoplanctônica e hidrologia do estuário do rio Timbó (Paulista, PE). Trop. Oceanogr. 2004, 32, 181–199. [Google Scholar] [CrossRef]
- Jales, M.C.; Feitosa, F.A.N.; Koening, M.L.; Bastos, R.B.; Machado, R.C.A. O Ecossistema Recifal de Serrambi (Nordeste Do Brasil): Biomassa Fitoplanctônica e Parâmetros Hidrológicos. Atlântica 2012, 34, 87–102. [Google Scholar] [CrossRef]
- Maranhão, G.M.B.; Fonseca-Genevois, V.; Passavante, J.Z.O. Meiofauna da área recifal da Baía de Tamandaré (Pernambuco, Brasil). Trop. Oceanog. UFPE 2000, 28, 47–59. [Google Scholar] [CrossRef][Green Version]
- Da Rocha, C.M.C.; Venekey, V.; Bezerra, T.N.C.; Souza, J.R.B. Phytal marine nematode assenblages and their relation with the macrophytes structural complexity in a Brazilian tropical rocky beach. Hydrobiologia 2006, 533, 210–230. [Google Scholar] [CrossRef]
- Cai, W.J.; Hu, X.; Huang, W.J.; Murrell, M.C.; Lehrter, J.C.; Lohrenz, S.E.; Zhao, P. Acidification of subsurface coastal waters enhanced by eutrophication. Nat. Geosci. 2011, 4, 766–770. [Google Scholar] [CrossRef]
- Czerniawska-Kusza, I. Use of artificial substrates for sampling benthic macroinvertebrates in the assessment of water quality of large lowland rivers. Pol. J. Environ. Stud. 2004, 13, 579–584. [Google Scholar]
- Somerfield, P.J.; Olsgard, F.; Carr, M.R. A further examination of two new taxonomic distinctness measures. Mar. Ecol. Progr. Ser. 1997, 154, 303–306. [Google Scholar] [CrossRef]
- Bavestrello, G.; Cerrano, C.; Zanzi, D.; Cattaneo-Vietti, R. Damage by fishing activities to the Gorgonian coral Paramuricea clavata in the Ligurian Sea. Aquat. Conserv. Mar. Freshw. Ecosyst. 2007, 7, 253–262. [Google Scholar] [CrossRef]
- Tsounis, G.; Rossi, S.; Gili, J.M.; Arntz, W. Red coral fishery at the Costa Brava (NW Mediterranean): Case study of an overharvested precious coral. Ecosystems 2007, 10, 975–986. [Google Scholar] [CrossRef]
- Coma, R.; Pola, E.; Ribes, M.; Zabala, M. Long-term assessment of temperate octocoral mortality patterns, protected vs. unprotected areas. Ecol. Appl. 2004, 14, 1466–1478. [Google Scholar] [CrossRef]
- Balata, D.; Piazzi, L.; Cecchi, E.; Cinelli, F. Variability of Mediterranean coralligenous assemblages subject to local variation in sediment deposition. Mar. Environ. Res. 2005, 60, 403–421. [Google Scholar] [CrossRef] [PubMed]
- Baldacconi, R.; Corriero, G. Effects of the spread of the alga Caulerpa racemosa var. Cylindracea on the sponge assemblage from coralligenous concretions of the Apulian coast (Ionian Sea, Italy). Mar. Ecol. 2009, 30, 337–345. [Google Scholar] [CrossRef]
- Garrabou, J.; Perez, T.; Sartoretto, S.; Harmelin, J.G. Mass mortality event in red coral Corallium rubrum populations in the Provence region (France, NW Mediterranean). Mar. Ecol. Progr. Ser. 2001, 217, 263–272. [Google Scholar] [CrossRef]
- Giuliani, S.; Lamberti, C.V.; Sonni, C.; Pellegrini, D. Mucilage impact on gorgonians in the Tyrrhenian Sea. Sci. Total Environ. 2005, 353, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Sousa, W.P. Experimental investigations of disturbance and ecological succession in a rocky intertidal algal community. Ecol. Monogr. 1979, 49, 227–254. [Google Scholar] [CrossRef]
- Whited, D.C.; Lorang, M.S.; Harner, M.J.; Hauer, F.R.; Kimball, J.S.; Stanford, J.A. Climate, hydrologic disturbance, and succession: Drivers of floodplain pattern. Ecology 2007, 88, 940–953. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.B. Ecological extinction and evolution in the brave new ocean. Proc. Nat. Acad. Sci. USA 2008, 105, 11458–11465. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Kroeker, K.J.; Gambi, M.-C.; Micheli, F. Community dynamics and ecosystem simplification in a high-CO2 ocean. Proc. Nat. Acad. Sci. USA 2013, 110, 12721–12726. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.E.; Milazzo, M.; Rastrick, S.P.; Hall-Spencer, J.M.; Therriault, T.W.; Harley, C.G. Natural acidification changes the timing and rate of succession, alters community structure, and increases homogeneity in marine biofouling communities. Glob. Chang. Biol. 2017, 24, e112–e127. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Reich, P.B.; Knops, J.M. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reef Areas | PMS | Salinity | Temperature | DO | pH | TA | SiO2 | DIN | DIP |
|---|---|---|---|---|---|---|---|---|---|
| Tamandaré | 45.1 (±3.9) | 40.5 (±2.1) | 29.5 (±0.7) | 133.4 (±7.8) | 8.20 (±0.07) | 2340.9 (±23.4) | 9.40 (±4.30) | 1.63 (±0.21) | 0.15 (±0.04) |
| Serrambi | 33.1 (±6.2) | 38.5 (±0.7) | 29.95 (±1.3) | 153.8 (±33.5) | 8.31 (±0.11) | 2315.7 (±51.1) | 5.38 (±1.57) | 2.80 (±1.56) | 0.22 (±0.08) |
| P. Galinhas | 38 (±1.4) | 37 (±1.41) | 30.15 (±1.6) | 119.4 (±2.6) | 8.22 (±0.03) | 2352.1 (±0.6) | 5.24 (±1.38) | 2.36 (±1.36) | 0.19 (±0.02) |
| Paiva | 46.6 (±13.3) | 38.5 (±0.7) | 29 (±1.4) | 147.5 (±13.2) | 8.40 (±0.10) | 2313.5 (±29.6) | 16.17 (±11.98) | 8.95 (±11.23) | 0.30 (±0.10) |
| R. Doce | 61.2 (±37.3) | 38 (±1.4) | 28 (±1.4) | 140.6 (±24.1) | 8.22 (±0.05) | 2317.0 (±56.3) | 39.39 (±11.54) | 11.68 (±2.77) | 1.03 (±0.12) |
| Piedade | 40.9 (±4.9) | 36 (±1.4) | 27 (±1.4) | 120.5 (±18.7) | 8.30 (±0.03) | 2331.9 (±7.9) | 19.91 (±19.08) | 4.61 (±3.05) | 0.40 (±0.08) |
| Sub-Order | Family | Genus | Species | Control | Impact |
|---|---|---|---|---|---|
| Polyarthra | Longipediidae | Longipedia | Longipedia sp. | 1.9 | 1.3 |
| Oligoarthra | Ameiridae | Ameira | Ameira parvula | 3.4 | 0.0 |
| Ameira sp1 | 5.5 | 0.0 | |||
| Ameira sp2 | 2.2 | 15.7 | |||
| Ameira sp3 | 0.5 | 2.9 | |||
| Ameira sp4 | 0.4 | 0.0 | |||
| Sarsameira | Sarsameira knorri | 0.9 | 0.0 | ||
| Sarsameira sp1 | 0.0 | 0.4 | |||
| Sarsameira sp2 | 0.7 | 0.0 | |||
| Canthocamptidae | Mesochra | Mesochra sp1 | 0.9 | 0.0 | |
| Mesochra sp2 | 0.2 | 1.5 | |||
| Mesochra sp3 | 0.2 | 0.0 | |||
| Nannomesochra | Nannomesochra sp. | 0.8 | 1.0 | ||
| Dactylopusiidae | Dactylopusia | Dactylopusia sp1 | 0.8 | 0.0 | |
| Dactylopusia sp2 | 1.9 | 1.9 | |||
| Dactylopusia tisboides | 3.2 | 5.8 | |||
| Diarthrodes | Diarthrodes sp1 | 0.4 | 0.0 | ||
| Diarthrodes sp2 | 0.2 | 0.0 | |||
| Paradactylopodia | Paradactylopodia brevicornis | 0.5 | 0.3 | ||
| Ectinosomatidae | Ectinosoma | Ectinosoma sp1 | 3.4 | 33.6 | |
| Ectinosoma sp2 | 1.8 | 1.9 | |||
| Ectinosoma sp3 | 0.4 | 0.0 | |||
| Pseudobradya | Pseudobradya sp. | 1.1 | 0.0 | ||
| Hamondiidae | Ambunguipes | Ambunguipes sp. | 0.5 | 0.0 | |
| Harpacticidae | Harpacticus | Harpacticus obscurus | 5.6 | 0.0 | |
| Harpacticus sp. | 0.0 | 1.4 | |||
| Laophontidae | Applanola | Applanola hirsuta | 0.0 | 0.6 | |
| Echinolaophonte | Echinolaophonte sp. | 0.0 | 0.2 | ||
| Laophonte | Laophonte cornuta | 0.5 | 0.0 | ||
| Laophonte parvula | 3.9 | 0.0 | |||
| Laophonte sp1 | 0.2 | 0.0 | |||
| Laophonte sp2 | 0.0 | 0.2 | |||
| Paralaophonte | Paralaophonte brevirostris | 0.0 | 0.7 | ||
| Paralaophonte congenera | 1.3 | 0.0 | |||
| Paralaophonte sp. | 0.0 | 3.9 | |||
| Miraciidae | Amonardia | Amonardia sp. | 0.4 | 0.0 | |
| Amphiascoides | Amphiascoides sp1 | 5.6 | 0.4 | ||
| Amphiascoides sp2 | 0.7 | 1.5 | |||
| Amphiascoides sp3 | 0.2 | 0.0 | |||
| Amphiascopsis | Amphiascopsis cinctus | 1.8 | 1.5 | ||
| Amphiascus | Amphiascus (Minutus) sp. | 0.6 | 0.3 | ||
| Amphiascus (Varians) sp. | 0.4 | 0.0 | |||
| Amphiascus sp. | 5.3 | 0.0 | |||
| Dactylopodamphiascopsis | Dactylopodamphiascopsis sp. | 0.5 | 0.0 | ||
| Delavalia | Delavalia sp. | 0.5 | 0.0 | ||
| Diosaccus | Diosaccus sp. | 0.5 | 0.0 | ||
| Haloschizopera | Haloschizopera sp. | 0.0 | 0.4 | ||
| Melima | Melima sp1 | 3.2 | 0.8 | ||
| Melima sp2 | 2.9 | 0.0 | |||
| Melima sp3 | 0.2 | 0.0 | |||
| Paramphiascella | Paramphiascella sp. | 4.4 | 0.8 | ||
| Robertgurneya | Robertgurneya sp. | 1.4 | 6.2 | ||
| Robertsonia | Robertsonia knoxi | 3.1 | 0.0 | ||
| Robertsonia mourei | 1.0 | 0.0 | |||
| Robertsonia sp1 | 0.7 | 0.5 | |||
| Robertsonia sp2 | 1.4 | 0.9 | |||
| Stenhelia | Stenhelia sp. | 1.3 | 0.0 | ||
| Parastenheliidae | Parastenhelia | Parastenhelia spinosa | 13.1 | 0.5 | |
| Peltididae | Alteutha | Alteutha sp. | 3.4 | 0.0 | |
| Eupelte | Eupelte sp. | 0.5 | 2.4 | ||
| Pseudotachidiidae | Xouthous | Xouthous sp. | 1.6 | 2.0 | |
| Thalestridae | Parathalestris | Parathalestris sp. | 0.4 | 0.0 | |
| Tisbidae | Scutellidium | Scutellidium sp. | 1.4 | 8.6 |
| Control | Impact & Control | ||
|---|---|---|---|
| Average similarity: 24.44 | Average dissimilarity = 86.52 | ||
| Species | Cum. % | Species | Cum. % |
| Parastenhelia spinosa | 24.38 | Ectinosoma sp1 | 17.47 |
| Laophonte parvula | 31.79 | Ameira sp2 | 26.68 |
| Amphiascoides sp1 | 38.84 | Parastenhelia spinosa | 33.59 |
| Harpacticus obscurus | 45.66 | Scutellidium sp. | 38.16 |
| Paramphiascella sp. | 51.53 | Robertgurneya sp. | 41.66 |
| Robertsonia knoxi | 56.79 | Dactylopusia tisboides | 44.89 |
| Dactylopusia tisboides | 61.13 | Harpacticus obscurus | 47.94 |
| Melima sp1 | 65.33 | Amphiascoides sp1 | 51 |
| Ectinosoma sp1 | 69.4 | Ameira sp1 | 53.99 |
| Amphiascus sp. | 73.37 | Amphiascus sp. | 56.95 |
| Alteutha sp. | 77.09 | Laophonte parvula | 59.55 |
| Ameira sp1 | 79.39 | Alteutha sp. | 61.97 |
| Melima sp2 | 81.61 | Paramphiascella sp. | 64.34 |
| Longipedia sp. | 83.66 | Paralaophonte sp. | 66.36 |
| Ameira parvula | 85.7 | Ameira parvula | 68.34 |
| Amphiascopsis cinctus | 87.49 | Melima sp1 | 70.26 |
| Xouthous sp. | 89.18 | Robertsonia knoxi | 72.08 |
| Ameira sp2 | 90.71 | Ectinosoma sp2 | 73.75 |
| Ameira sp3 | 75.43 | ||
| Dactylopusia sp2 | 77.08 | ||
| Impact | Amphiascopsis cinctus | 78.53 | |
| Average similarity: 49.76 | Melima sp2 | 79.98 | |
| Xouthous sp. | 81.39 | ||
| Species | Cum. % | Eupelte sp. | 82.76 |
| Ectinosoma sp1 | 46.21 | Longipedia sp. | 84.07 |
| Ameira sp2 | 69.18 | Robertsonia sp2 | 85.22 |
| Scutellidium sp. | 77.84 | Amphiascoides sp2 | 86.26 |
| Dactylopusia tisboides | 86.2 | Nannomesochra sp. | 87.25 |
| Robertgurneya sp. | 90.46 | Mesochra sp2 | 88.23 |
| Paralaophonte congenera | 89.11 | ||
| Harpacticus sp. | 89.84 | ||
| Stenhelia sp. | 90.53 |
| Source | Df | SS | MS | Pseudo-F | P (MC) |
|---|---|---|---|---|---|
| Impact | 1 | 21,768 | 21,768 | 3.6876 | 0.0097 |
| Reef (Im) | 4 | 23,612 | 5903 | 4.5396 | 0.0001 |
| Res | 18 | 23,406 | 1300.3 | ||
| Total | 23 | 68,786 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barroso, M.S.; Da Silva, B.J.; Flores Montes, M.J.; Santos, P.J.P. Anthropogenic Impacts on Coral Reef Harpacticoid Copepods. Diversity 2018, 10, 32. https://doi.org/10.3390/d10020032
Barroso MS, Da Silva BJ, Flores Montes MJ, Santos PJP. Anthropogenic Impacts on Coral Reef Harpacticoid Copepods. Diversity. 2018; 10(2):32. https://doi.org/10.3390/d10020032
Chicago/Turabian StyleBarroso, Marina Siqueira, Brenno J. Da Silva, Manuel J. Flores Montes, and Paulo J. P. Santos. 2018. "Anthropogenic Impacts on Coral Reef Harpacticoid Copepods" Diversity 10, no. 2: 32. https://doi.org/10.3390/d10020032
APA StyleBarroso, M. S., Da Silva, B. J., Flores Montes, M. J., & Santos, P. J. P. (2018). Anthropogenic Impacts on Coral Reef Harpacticoid Copepods. Diversity, 10(2), 32. https://doi.org/10.3390/d10020032